Изгнание из Эдема [Стивен Оппенгеймер] (fb2) читать онлайн
- Изгнание из Эдема (пер. Андрей Михайлович Голов, ...) (а.с. Тайны древних цивилизаций) 6.83 Мб, 594с. скачать: (fb2) - (исправленную) читать: (полностью) - (постранично) - Стивен Оппенгеймер
[Настройки текста] [Cбросить фильтры]
[Оглавление]
СТИВЕН ОППЕНГЕЙМЕР ИЗГНАНИЕ ИЗ ЭДЕМА ХРОНИКИ ДЕМОГРАФИЧЕСКОГО ВЗРЫВА
Посвящается моей дочери Мэйлин и сыну Дэвиду, моей жене Фреде и моим отцу и матери.
«Чтобы знать, куда мы идем, необходимо понять, кто мы; чтобы понять, кто мы, необходимо знать, откуда мы родом».Филиппинский вариант пословицы одного из народов Океании
ПРЕДИСЛОВИЕ
Представьте себе, что вы стоите в очереди в пункте таможенного контроля в аэропорту Чикаго или Лондона. Рядом с вами — семеро точно таких же пассажиров. Один из них — афроамериканец с Карибских островов, другой, точнее, другая — светловолосая девушка, уроженка одной из стран Северной Европы. Третий ваш сосед — специалист по компьютерной технике родом из Индии. Четвертый — подросток-китаец, слушающий музыку, надев наушники своего плейера. Пятый, шестой и седьмой ваши соседи — участники научной конференции по наскальным рисункам, прибывшие соответственно из Австралии, Новой Гвинеи и Южной Америки. Все семеро ведут себя тихо и скромно, избегая смотреть друг другу в глаза, поскольку незнакомы и чувствуют себя совершенно чужими друг другу. И тем не менее нам не составит труда доказать, что все они — дальние родственники, ибо у всех них были общие предки, причем как по женской, так и по мужской линии. В каждой клетке нашего тела присутствуют гены. Гены состоят из ДНК (дезоксирибонуклеиновой кислоты), особого, похожего на длинную цепь, «кода» жизни, который заключает в себе информацию о том, кто мы и откуда, и описывает все наши свойства и характерные особенности — от строения ногтей до врожденной одаренности. И если мы проанализируем гены этих семи соседей-пассажиров, мы сможем проследить пути и маршруты их далеких предков в пространстве и времени, вплоть до локуса появления их общего прапредка в Африке на заре существования рода человеческого. Более того, если мы выберем наугад двух пассажиров и сопоставим их гены, мы без труда обнаружим, что у них есть и более поздний общий предок, живший, скорее всего, уже за пределами Африки. Кроме того, мы без проблем сможем определить, где именно жили их предки и когда покинули свою древнейшую прародину. Подобная система доказательств стала возможной лишь в последнее десятилетие благодаря новаторским разработкам целого ряда ученых. Многие из нас были бы немало удивлены, если бы очутились на пресловутой машине времени, которая унесла бы нас в глубины минувшего, через бесчисленные поколения наших далеких и близких предков. Интересно, куда бы она принесла нас? А вдруг мы оказались бы родственниками некой знаменитости или выдающейся личности? Сколько поколений нам пришлось бы отсчитать вспять, чтобы столкнуться лицом к лицу с первыми людьми на нашей планете? А что, если общая линия наследственности, перешагнув порог человечества, пойдет дальше — к обезьянам, червям или простейшим одноклеточным организмам, как полагал Дарвин? Разумеется, мы помним об этом еще со школьной скамьи по урокам биологии, однако, учитывая неопределенность участи, которая ожидает наши души после смерти, в это очень непросто поверить. Мы до такой степени привыкли к постоянно убыстряющемуся темпу технического прогресса, что с каждым новым его достижением чувство удивления и восторга все более и более слабеет. И все же вплоть до самого последнего времени генетики могли лишь мечтать о том, чтобы использовать гены в качестве маркеров, позволяющих во всех деталях проследить историю заселения мира человечеством. Главная причина их пессимизма заключалась в том, что большинство изученных ими генов как бы заново перемешиваются в каждом новом поколении и присутствуют у подавляющего большинства людей. Задача генетиков была невероятно сложной, напоминая попытку восстановить предыдущую сдачу карт по колоде после того, как она была тщательно и многократно перетасована. Таким образом, казалось почти немыслимым выстроить сколько-нибудь точное семейное генетическое древо, уходящее всего на несколько веков назад, не говоря уж о прослеживании такого же древа от самых корней — с момента появления человека современного типа. Изнутри подавляющее большинство современных людей выглядят практически одинаково. Так с чего же, собственно, начать? Однако методика использования тендерной дифференциации генов и их линий, выделения так называемых генов Адама и Евы, возникшая в последнее десятилетие, коренным образом изменила ситуацию в этой области. В отличие от всех прочих генов, митохондриевую ДНК (целый свод генов в пределах одной клетки) мы наследуем только от матери, а Y-хромосому — только от отца. Эти два набора генов, маркированных по признаку пола, передаются без всяких изменений из поколения в поколение, не смешиваясь и не «перетасовываясь», как карты, и благодаря этому их можно проследить вплоть до самых отдаленных наших предков, первых млекопитающих, червей и даже хуже того... Мы можем также реконструировать два семейных генетических древа: одно — наших отцов и другое — наших матерей-прародительниц. В результате в любой группе людей, сколь бы малочисленной она ни была, по одному из этих генетических древ мы можем проследить родословную любой пары вплоть до общего предка, на какой бы ветви он ни находился. Так, общие предки тех или иных людей могли жить 200, 5000 или 150 тысяч лет назад, и тем не менее они тоже занимают свое строго определенное место на реконструированном древе Адама или Евы. Это — вполне реальные генеалогические деревья генов современных людей, имеющие конкретные ветви и отростки. При этом можно определить даже возраст каждой ветви на любом из этих деревьев (хотя надо признать, что точность такой датировки оставляет желать много лучшего). Сегодня уже установлены контуры многих региональных генных деревьев, причем создается впечатление, что они «пропилены» огромной пилой по тем или иным регионам с учетом вполне конкретных ориентиров. Именно таким путем за последнее десятилетие была составлена сводная картина распространения генных линий Адама и Евы, протянувшихся в самые отдаленные уголки света. И вот настал момент, когда, совсем как при выпиливании, вся картина вдруг выходит из плоскости и приобретает узнаваемые очертания. И хотя древо остается пока еще далеко не полным, недостающие фрагменты появляются на карте все чаще и с меньшими усилиями. Темпы прогресса здесь настолько стремительны, что ученые, работающие на переднем крае в одном географическом регионе, могут и не подозревать о прорывах, достигнутых в других областях. Зато мы можем наложить это необъятное генное древо на карту мира и проследить, где, обживая неведомый древний мир, прошли наши далекие предки, оставляя цепочки и линии генов. Эти новые знания позволили решить некоторые бросающиеся в глаза парадоксы, обусловленные вопиющими контрастами между культурной и биологической историей человечества за последние 150 тысяч лет. Сегодня мы можем даже поместить ту или иную находку-окаменелость, относящуюся к определенной эпохе, на соответствующую ей ветвь или отросток на генном древе жизни. Действительно, ученым удалось найти ответ на многие вопросы. Как оказалось, несмотря на то что наш мир представляет собой нечто вроде гигантского плавильного котла, в котором с доисторических времен переплавляются и смешиваются друг с другом всевозможные человеческие потоки, подавляющее большинство ныне живущих представителей человеческой диаспоры с незапамятных времен живут на тех же самых местах, которые избрали их далекие предки. Они живут в этих местах со времен Последнего Ледникового периода. Более того, мы можем даже проследить даты переселения тех или иных народов за последние 80 тысяч лет. Таким образом, от картины, характеризовавшейся многовариантностью и неточностью критериев идентификации, мы неожиданно получили возможность обратиться к точной и предусматривающей региональное деление схеме, отражающей все ветви расселения человека в мире. Приведем несколько примеров того, как благодаря генному древу удалось дать ответ на целый ряд вопросов, издавна не дающих покоя археологам. Один из них — противоречие между «африканским» и «мультирегиональным» характером происхождения человечества. Гипотеза об африканском происхождении утверждает, что все современные люди, живущие ныне на Земле, являются потомками общих предков, совершивших около 100 тысяч лет тому назад массовый исход из Африки. Этот исход привел к исчезновению всех ранее существовавших типов человека во всем мире. Сторонники регионалистской теории, напротив, стремятся доказать, что древнейшие люди на Земле, представители вида Homo neanderthalis (неандертальцы) в Европе и Homo erectus (человек прямоходящий) на Дальнем Востоке в результате длительной эволюции создали те расы, которые существуют сегодня в мире. На сегодняшний день верх в этом диспуте явно одерживают сторонники гипотезы об африканском происхождении, поскольку генное древо человечества, охватывающее последние 100 тысяч лет, прямо указывает на Африку[1] . Никаких следов более древних видов человека на генном древе Адама и Евы не прослеживается, за исключением, естественно, самого его корня, относительно которого мы можем отсчитывать, насколько далеко наши гены отстоят от неандертальцев. Генетический тип неандертальского человека определяет митохондриевая ДНК, и неандертальцы, по всей видимости, были скорее нашими двоюродными братьями, нежели прямыми предками. С неандертальцами нас объединяет общий предок — так называемый Homo helmei. Современные приверженцы теории африканского происхождения обычно подкрепляют свои взгляды утверждениями о том, что австралийцы, жители Азии и европейцы представляют собой потомков различных ветвей Homo sapiens — выходцев из Африки. Однако это не совсем так: лишь одна из линий и на мужском, и на женском генетическом древе имеет бесспорно африканское происхождение. В этом и заключается ключевой аргумент моей книги. Я утверждаю, что в древности имел место всего лишь один великий исход из Африки, и поэтому представители обоих полов имеют только одного общего генетического предка, который и стал отцом или матерью всех прочих народов за пределами Африки, расселившихся по всему свету. Это повлекло за собой крах многих прежних предвзятых мнений. Некоторые европейские археологи и антропологи еще не так давно полагали, что именно европейцы первыми освоили навыки рисования и резьбы, создали первую сложную культуру и даже первыми обрели дар речи — словом, возникало впечатление, будто европейцы обладают неким явным биологическим преимуществом перед другими расами. Между тем структура генетического древа опровергает подобную точку зрения. Аборигены Австралии в генетическом отношении тесно связаны с европейцами и имеют с ними даже общего предка, жившего примерно 70 тысяч лет назад — вскоре после исхода народов из Африки в Йемен. После этого будущие обитатели Австралии отправились вдоль береговой линии побережья Индийского океана, совершив растянувшийся на несколько тысячелетий марш-бросок через острова Индонезии в Австралию, где, оказавшись в полной изоляции от внешнего мира, создали свою собственную, уникальную и сложную художественную культуру. Самые ранние наскальные рисунки, найденные в Австралии, по меньшей мере не уступают в древности творениям первых европейских художников. Это означает, что люди, совершившие исход из Африки, уже владели навыками рисования. Другой загадочный парадокс, давно не дававший покоя археологам, заключался в неожиданном и быстром распространении своеобразной неолитической культуры по всей Европе из Турции. Произошло это около 8000 лет тому назад. Быть может, земледельцы с Ближнего Востока оттеснили и уничтожили древних европейских охотников и собирателей или новые идеи распространялись мирным путем, постепенно получая признание у туземных европейских общин эпохи палеолита? Генетический ответ на этот вопрос совершенно ясен: 80% жителей современной Европы являются потомками представителей генного типа древних охотников и собирателей, и лишь 20% происходят от ближневосточных охотников и собирателей. Таким образом, древние туземцы Европы вовсе не были безвольными слабаками[2]. Наконец, если перенестись на другой конец света, надо признать, что там всегда существовала масса споров об этническом происхождении полинезийцев. Увы, Тур Хейердал оказался здесь далеко не первым (в сущности, капитан Кук оказался куда ближе к истине, полагая, что полинезийцы имеют близкие родственные связи с Малайским архипелагом). Лет пятнадцать тому назад археологи были убеждены, что полинезийцы — это выходцы с острова Тайвань. Однако сегодня генетическое древо опровергает эту версию: предки мореходов на больших каноэ были выходцами из другого региона — Восточной Индонезии[3]. А теперь вернемся в очередь в аэропорту. Давайте вспомним, что мы тоже — участники этой генетической истории, поскольку 99% работ по реконструкции нашего древнего генного древа было выполнено с генетическим материалом и пробами ДНК, которые сдали добровольцы, проживающие в самых разных концах света. И поэтому эта история касается каждого из нас.
ПРОЛОГ
Сегодня многие антропологи утверждают, что весь род человеческий — это потомки выходцев из Африки. Откуда же им это известно? Если у нас действительно был некий общий прапредок, почему же тогда существуют столь разные человеческие расы? Как эти расы соотносятся между собой? Правомерно ли вообще само понятие «расы»? Являемся ли мы, люди, частью общей семьи, или у африканцев, аборигенов Австралии, европейцев и жителей Восточной Азии имеются разные корни и прошли они в своем развитии параллельные этапы эволюции? Но откуда же мы? Какие движущие силы в ходе нашей эволюции побудили потомков приматов, совсем недавно спустившихся с деревьев на землю, отправиться в скитания по необъятным африканским саваннам, а затем, через какую-нибудь пару миллионов лет, позволили им совершить полет на Луну? Нашумевшие книги Джейкоба Брауна «Происхождение человека» и Ричарда Лики «Сотворение рода человеческого» явились этапными вехами на пути привлечения интереса широкой публики к проблемам эволюции человека. И тем не менее они, как и любые книги подобного рода, оставили без ответа множество вопросов. Отсутствие достоверных материальных свидетельств, относящихся к той эпохе, представляет собой как бы зазоры и просветы, неизбежно зияющие между нашими представлениями и реальными знаниями. Недавние археологические находки и открытия в области биологии позволили нам заполнить хотя бы некоторые из этих лакун и в то же время выдвинуть целый ряд новых вопросов. Сегодня мы в состоянии внести коррективы в наши искаженные представления о том, когда и как произошел решающий перелом, отделивший нас от наших предков и двоюродных собратьев, крупных приматов, а также развеять многие мифы, стоящие за привычными представлениями о прогрессе человечества. Анализ ДНК позволил нам достичь невиданного прогресса в осмыслении факторов региональной биологической истории эволюции человека современного типа. Как мы увидим в дальнейшем, так называемые гены Адама и Евы действительно позволяют нам совершить путешествие в пространстве и во времени и перенестись в далекое прошлое, чтобы проследить пути странствий предков человека по земному шару. Однако далеко не все факторы прогресса нашли свое отражение в молекулярной биологии. Так, например, палеоантропология, научная дисциплина, изучающая наших далеких предков, с момента выхода в свет книги Лики, опубликованной двадцать лет тому назад, достигла значительного прогресса сразу в нескольких важнейших областях. Прежде всего целый ряд недавно открытых черепов древнего человека, найденных как в Африке, так и в других частях света, позволил определить временные рамки и географические координаты периодически повторявшихся миграций человека из Африки за последние 2 миллиона лет. Находки других черепов позволили выявить черты родства и преемственности между нашими древнейшими предками и шимпанзе, жившими в еще более ранние эпохи. Во-вторых, после создания компьютеров сравнительный анализ строения и формы черепов обрел более достоверную научную базу, и в результате стало возможным разместить основные типы черепов доисторических людей, найденные по всему миру, на соответствующих ветвях генетического древа, а черепа их современных потомков — на его побегах. Сам принцип формулирования ветвей биологического древа современного человека на основании сравнительного анализа черепа с трудом пробил себе дорогу сквозь тучи псевдонаучных взглядов, связанных с расизмом и деятельностью нацистских антропологов в середине XX в. Начиная с 1960-х гг. возникли новые, более объективные и доказательные математические методы, позволившие палеоантропологам построить более детальную модель генеалогического древа гоминида и даже человека современного типа. Заметно возросший интерес публики к этой теме, а также опасения, что выводы подобных исследований могут быть использованы нацистами и расистами в своих неблаговидных целях, во многом завели эти исследования, образно говоря, на минное поле пресловутых «политически корректных» эвфемизмов, устроенное западными антрополагами под предлогом защиты от предвзятых мнений. Хотя такой язык явно имел своей целью более четко сформулировать различия между человеческими расами, на практике он привел к путанице и неясностям, что дало импульс дискуссиям и спорам. Так, например, понятие «раса» сегодня считается политически некорректным термином, а в некоторых кругах так же воспринимается и термин «этническая принадлежность». Подобную гиперчувствительность следует рассматривать как вызов, а не как реальное препятствие. Обсуждение проблемы различий между расами носит расистский характер, если посредством таких различий пытаются подчеркнуть собственное превосходство и исключительность или принизить другие народы. Однако оно вполне допустимо в контексте позитивной оценки многообразия рода человеческого. В этой книге я хотел бы в первую очередь затронуть вопросы истории эволюции человечества за последние 200 тысяч лет, ответить на которые можно, только проследив вспять, до самых истоков, линии эволюции наших генов. Но чтобы решить эту задачу и осознать, что же побудило наших далеких предков обойти буквально все уголки нашей планеты, мы должны прежде всего задать себе вопрос: «Кто мы?» Силы эволюции, включившие в нас механизмы адаптации к трудностям жизни на просторах африканских саванн — это и есть ключи к нашей природе и невероятно увлекательной истории человечества. Мы не были спущены с небес в этот мир, так сказать, «в готовом виде» — прекрасно сложенными мыслящими существами, наделенными даром речи и выделяющимися среди всех прочих животных. На нас воздействовали силы целенаправленного отбора и слепые, беспощадные и безличные факторы окружающей среды. Как и у всех прочих видов живых существ, у нас были предки и ближайшие родичи, которые были наделены теми или иными из наших врожденных качеств, однако они не сумели воспользоваться ими и вымерли. Наша физическая и поведенческая адаптация была сконцентрирована на выживании в борьбе с нашим заклятым врагом и беспощадным учителем — климатом. Одна из бессмертных повестей Редьярда Киплинга из цикла «Просто сказки» (1902) рассказывает о малыше Слоненке, у которого в те времена еще не было хобота и который донимал своих дядюшек и тетушек назойливыми вопросами. Однажды он совершил ошибку: спросил коварного Крокодила, что тот обычно кушает на обед. Не успел малыш договорить, как Крокодил вцепился ему челюстями в нос и изо всех сил потащил Слоненка в реку. После упорной борьбы малышу удалось вырваться, но его нос при этом сильно вытянулся, превратившись в длинный и очень удобный хобот. Эта притча Киплинга[4], как и другие повести того же сборника, образно говоря, показывает нос эволюционной теории Жана-Батиста Ламарка, объясняющей передачу приобретенных признаков, но в ней содержится и намек на теорию Дарвина. Малыш Слоненок, хотя и весьма жестоким путем, но все же получил новый орган — видоизмененный нос, оказавшийся невероятно удобным, хотя малышу понадобилось немало времени, чтобы осознать это. Точно так же изменения нашего мозга, свободных конечностей (рук, ног), семейной и общественной жизни возникли путем отбора под воздействием жестко неблагоприятных погодных факторов за последние несколько миллионов лет, и мы нашли им совершенно новое применение. Один из наиболее поразительных выводов был сделан на основе детального изучения воздействия периодически повторяющихся циклов оледенения на эволюцию человека и миграционные потоки из Африки за последние 2,5 миллиона лет. Наряду с тем, что суровые климатические условия повлекли за собой массовую гибель и полное вымирание представителей мегафауны (гигантских и особо крупных видов животных), появление новых и лучше приспособленных типов человека, по всей видимости, совпало с суровым оледенением и ухудшением климата в африканских саваннах. Однако изменения климата могли стать движущей силой, стоящей за процессом эволюции человека и определяющей ее в куда более длительной перспективе. Для приматов, как правило, характерны хорошо развитые и приспособленные верхние конечности, сравнительно крупный мозг, разнообразный рацион питания и куда более сложные социальные отношения в группе, чем у большинства современных млекопитающих. 10 миллионов лет назад Африка была настоящим земным раем, где шумели пышные леса — родной дом и колыбель некоторых видов человекообразных приматов. В те времена не только эти человекообразные приматы, но и многие другие виды обезьян активно экспериментировали, пытаясь приспособиться к жизни на земле и поросших высокой травой луговинах у окраин лесов. С тех пор, по мере того как климат в мире становился все более прохладным и засушливым, площадь таких безлесых пространств (саванн) в Африке многократно расширилась, но сами изменения носили циклический характер, повторяясь с нарастающей частотой и интенсивностью.
Приматы, способные ходить на двух ногах
Как мы уже знаем благодаря изучению воздействий Последнего Ледникового периода, наиболее негативная фаза климатического цикла, несмотря на свою непродолжительность, способна привести к вымиранию многих видов животных. Выжившие во время подобных климатических катаклизмов могут иметь уже свой особый генетический тип, сохраняющий определенные специфические черты, которые и помогают им приспособиться к внешним факторам. Примерно 7—8 миллионов лет назад произошло резкое сокращение числа видов человекообразных приматов, совпавшее по времени с расширением площади безлесых травяных степей и глобальным похолоданием, которое продолжалось несколько миллионов лет. Некоторые ученые утверждают, что эта недолгая ледниковая эпоха ознаменовалась тем, что во время нее окончательно разошлись пути эволюции предков современного человека и предков нынешних шимпанзе. Возможно, именно тогда появилось и наиболее важное изменение физического облика наших предков — бипедализм (способность передвигаться на двух ногах). Впрочем, наиболее ранними на сегодняшний день свидетельствами бипедализма являются останки скелетов Australopithecus anamensis — примата, передвигавшегося на двух конечностях. Эти скелеты, датируемые 4 млн. лет назад, были найдены в 1995 г. на берегах озера Туркана в северной Кении[5]. Многие ученые убеждены в том, что существует непосредственная связь между расширением площади безлесых пространств и переходом приматов от квадрипедализма (передвижения на четырех конечностях) обитателей лесов к бипедализму жителей саванн. Возможно, в этом есть доля истины, но, судя по обилию зубастых хищников, с которыми им приходилось делить свое жизненное пространство, первые двуногие приматы, по всей видимости, не рисковали заходить на равнину слишком далеко от спасительных ветвей островков родного леса. Во всяком случае, другие приматы, в частности — предки бабуинов, на удивление хорошо приспособились передвигаться по саваннам на всех четырех конечностях (как поступают и современные бабуины). Другие исследователи выдвигали альтернативные теории, объясняющие, почему передвижение на двух конечностях было более предпочтительным. Это якобы позволяло — в самом буквальном смысле — сохранять голову холодной[6] или, подобно африканским сурикатам (мангустам), наблюдать за окрестностями, следя, не приближается ли хищник. Однако хотя мозг наших предков был значительно крупнее, чем у большинства сухопутных млекопитающих, он не превосходил по величине мозг наших ближайших сородичей — шимпанзе, так что сколько-нибудь реальной опасности перегрева мозга не существовало. Точно так же не давало никаких особых преимуществ и передвижение на двух конечностях. На это способен не только человек, но и многие другие млекопитающие, в том числе обезьяны, шимпанзе, медведи и сурикаты, которые могут длительное время передвигаться на двух конечностях. С точки зрения эволюционного учения более привлекательной выглядит идея о том, что приматы начали освобождать верхние конечности (руки) для того, чтобы совершать ими какие-либо полезные действия, например, держать тяжелые дубины на охоте (или, что более вероятно, орудия защиты от хищников, поскольку наши предки были преимущественно вегетарианцами). К сожалению, у нас нет прямых доказательств этой гипотезы, поскольку дерево — материал недолговечный, а каменных орудий, относящихся к тому времени, пока что не обнаружено. За этими первыми приматами, передвигавшимися на двух конечностях, о которых сохранились весьма скудные сведения, последовали представители известного семейства «Люси» — Australopithecus afarensis. Люси — это древняя самка примата, фрагменты скелета которой были обнаружены в 1974 г. Дональдом Джексоном в Гадаре, Эфиопия. Представители этого вида, жившего примерно 3—4 млн. лет назад, имели рост от 1 до 1,5 м, более прямую осанку и передвигались на двух конечностях, а их таз был более близок к нашему. Однако подобное сходство заканчивалось на уровне шеи, поскольку череп и мозг у представителей этого вида были более близки шимпанзе (так, объем мозга у них составлял 378—500 куб. см), хотя клыки у них были значительно меньше (см. рис. 1). Самки этого вида, как и самки горилл, были значительно меньше, образуя нечто вроде гаремов при сильных самцах. Другой вид прямоходящих приматов (Australopithecus africanus), живший между 2 и 3 млн. лет назад, имел такой же рост, но несколько больший размер мозга, чем шимпанзе, а именно — от 420 до 500 куб. см. Зубы у них также были меньше и более походили на наши. Следует заметить, что эти два вида прямоходящих приматов не обязательно являются этапами эволюционного развития на пути к появлению человека современного типа. Наши предки в те времена вполне могли представлять собой пока что не открытых собратьев этих приматов. Так, например, нашим ближайшим предком может считаться недавно обнаруженный археологами вид, живший около 3,6 млн. лет назад и названный Kenyanthropus platyops, что буквально означает «плосколицый » (рис. 0.1). Плоское лицо, признак, присущий человеку, возможно, представляет собой связующее звено между прямоходящими приматами и нами, людьми современного типа. Можно с уверенностью сказать, что на протяжении нескольких миллионов лет, когда представители австралопитеков («южных приматов») и их непосредственные предки бродили по травянистым саваннам Африки, у них имело место очень небольшое, а отнюдь не резкое увеличение размеров мозга.
 Фото 1. Реконструкция внешнего облика Australopithecus afarensis, из семейства прямоходящих приматов Люси, живших 3—4 млн лет назад. Они одними из первых научились передвигаться прямо на двух ногах (бипедализм), однако объем черепа и мозга у них были такими же, как у шимпанзе.
Фото 1. Реконструкция внешнего облика Australopithecus afarensis, из семейства прямоходящих приматов Люси, живших 3—4 млн лет назад. Они одними из первых научились передвигаться прямо на двух ногах (бипедализм), однако объем черепа и мозга у них были такими же, как у шимпанзе.
 Рис. 0.1
Рис. 0.1
Увеличение размеров мозга в великую засуху
Коренные изменения начались тогда, когда около 2,5 млн. лет назад в мире произошло глобальное похолодание. За несколько миллионов лет влажный и теплый плейстоценовый геологический период уступил место плейстоценовой ледниковой эпохе. Эта эпоха представляла собой своего рода климатическую мельницу, то есть цикл чередующихся сухих ледниковых периодов, во время которого происходили то расширения, то резкие сокращения площадей африканских травяных саванн. Так продолжалось вплоть до Последнего Ледникового периода, кульминация которого имела место около 18 тысяч лет тому назад. Вскоре после начала этого неустойчивого, холодного и засушливого периода на просторах африканских саванн появились первые люди (носители генов Homo) с древнейшими каменными орудиями и значительно более крупным мозгом. Подобно переходу от хождения на четырех конечностях и ходьбе на двух, имевшему место несколько миллионов лет назад, это явилось новым этапом эволюции прямоходящих приматов. У одной из их ветвей, так называемого Paranthropus, развились крупные и сильные челюсти, позволявшие представителям этого вида пережевывать жесткую растительную пищу. Представители другой ветви, Homo, научились делать каменные орудия. У них развился крупный мозг, что позволило им выйти на прямой путь эволюции, на котором они сначала стали охотниками и собирателями, а затем — людьми современного типа. Разумеется, мы склонны думать, что человек — это представитель некоего особого вида, приципиально отличного от прочих приматов. Многие полагают, что тем, кем мы сегодня стали, нас сделал наш крупный мозг. Другие считают даже, что именно заметное увеличение объема мозга побудило древних приступить к созданию примитивных орудий, однако этот аргумент представляется малоубедительным. Да, действительно, тщательно обработанные орудия — это признак, отличающий человека (а возможно — и представителей вида Paranthropus[7]), однако даже шимпанзе способны делать из дерева примитивные, но эффективные орудия. Между тем шимпанзе обладают значительно меньшим по объему мозгом и очень давно отделились от прямоходящих приматов. И хотя до нас, естественно, не дошли остатки их примитивных орудий, созданных около 7 млн. лет назад, объем мозга у шимпанзе до сих пор примерно такой же, как у нашего общего предка, жившего на заре цивилизации. Это говорит о том, что между умением делать простейшие орудия и развитием крупного мозга не существует непосредственной связи. Равно как из сказанного отнюдь не следует, что общие предки человека и шимпанзе в глубокой древности умели делать подобные орудия. У одного из древнейших видов человека, действительно делавшего орудия труда, так называемого Homo habilis[8], средний объем мозга составлял около 650 куб. см. Однако среди известных сегодня черепов Homo habilis один, возраст которого составляет 1,9 млн. лет, имел объем мозга, сравнимый с мозгом шимпанзе: 500 куб. см, что аналогично верхнему пределу мозга австралопитеков[9]. Небольшой рост и объем мозга, а также другие приматоподобные черты внешнего облика Homo habilis позволили некоторым антропологам высказать утверждение, что этот вид был своего рода исключением из генофонда Homo или «клуба предков человека», однако, несмотря на этот широко известный случай, он выглядит скорее предрассудком, нежели научно обоснованным аргументом. Маловероятно, что представители вида Homo habilis были нашими предками (впрочем, то же самое можно сказать и о большинстве гоминидов); тем не менее они умели делать каменные орудия. Сама идея о том, что мы сначала сумели обзавестись крупным мозгом, а затем стали решать, как с ним быть, представляет собой отказ от основополагающих принципов дарвинизма. Появлению любых новых навыков поведения всегда должна предшествовать физическая адаптация, лежащая в основе таких новых навыков. Еще задолго до начала плейстоценовой ледниковой эпохи в нашем поведении должны были существовать некоторые аспекты, в частности — обуславливавшие нашу способность адаптироваться к изменениям климата, которые и способствовали активному развитию и увеличению объема мозга. Проблема поиска пищи в условиях все более засушливой среды обитания могла быть решена благодаря изобретательности и находчивости наших предков. В этом им, несомненно, помог более крупный мозг. Подобное поведение в известной степени сохранилось и у нас, современных людей, поскольку за последние 2,5 млн. лет в результате каждого сколько-нибудь крупного оледенения в Африке появлялись новые виды человека с большим объемом мозга и широким диапазоном навыков. После того как после очередного ледникового максимума наступило короткое потепление климата, Сахара на несколько тысяч лет покрылась обильной растительностью, и люди нового вида отправились с ее просторов в дальний путь, чтобы попытать счастья на землях Евразии. Около 1 млн. лет назад объем мозга у представителей различных видов человека, живших как в Африке, так и за ее пределами, увеличился с 400 до 1000 куб. см и даже практически достиг современных размеров. Другими словами, объем мозга человека увеличился на добрых три четверти еще задолго до того, как на исторической сцене появились люди современного типа[10].
Почему у нас развился крупный мозг?
Ученые и исследователи высказывали различные предположения о том, что же явилось ключевой предпосылкой для развития крупного мозга. Постоянные ледниковые периоды вызвали в Африке крайнюю засуху и наверняка побуждали немногочисленных обитателей саванн проявлять изобретательность и стремление к коллективизму. Нам нетрудно понять, сколь велика была в подобных условиях ценность крупного мозга, и все же остается вопрос: почему столь крупный мозг развился только у нас, людей, а не у других млекопитающих, тоже живших на границе саванн? Один из важных аспектов, которому в наши дни придают большое значение, — это появление в рационе питания древних людей, наряду с растительной пищей, мяса. В частности, видный лондонский антрополог Лесли Эйэлло и его коллега Питер Уилер (основоположник теории охлаждения черепа) утверждают, что для активного роста и развития мозга нашим предкам было просто необходимо есть мясо[11]. Дело в том, что для полноценного питания мозга необходимо множество калорий, а для роста и развития — особые высококалорийные питательные вещества. Тем не менее, как подчеркивают Эйэлло и Уилер, потребление мяса — это скорее средство, чем причина увеличения объема мозга. Эти же исследователи доказывают, что параллельное уменьшение длины кишечника у человека, как о том свидетельствует изменение формы грудной клетки у Homo ergaster, является бесспорным доказательством изменения рациона питания и употребления в пищу большего количества мяса и сокращения доли овощей. Другими словами, древнейшие люди современного типа утратили чисто вегетарианский желудок, столь характерный для австралопитеков. Однако это изменение строения грудной клетки произошло задолго до начала быстрого роста мозга. Представители древнейших видов человека, таких, как Homo habilis, Homo rudolphensis и Homo ergaster, были скорее собирателями, нежели охотниками. Вероятно, со временем они научились использовать в качестве оружия палки и камни, защищаясь от нападения крупных хищников, а также применяли такое же оружие в совместной охоте на травоядных, что сплачивало людей и придавало им больше уверенности в своих силах. Однако без реальных материальных доказательств и артефактов все утверждения подобного рода о влиянии климата и употребления в пищу мяса на активный рост мозга у древнейшего человека остаются не более чем кабинетными домыслами. Так, нам известно, что по крайней мере в Африке Homo erectus пользовался каменными орудиями для разделки мяса[12], но для того, чтобы выявить наличие непосредственной связи между употреблением мяса, ухудшением климата и активным ростом мозга, нам придется сопоставить биологические параметры древнейших людей с чистыми вегетарианцами-приматами, жившими в ту же эпоху и при точно таких же климатических условиях. Не так давно Сара Элтон[13], антрополог, сотрудница Кентского университета в Кентербери, высказала примерно такие же мысли, но результаты ее исследований во многом поколебали нашу веру в уникальность человека как биологического вида. Она измерила объем черепа у ископаемых останков некоторых видов приматов, живших в период от 2,5 млн. (предполагаемое начало глобального похолодания) до 1,5 млн. лет тому назад. Элтон провела исследование двух основных ветвей гоминидов, которые разделились именно в тот период, — представителей Homo и Paranthropus, объединяющих в общей сложности шесть видов. В качестве объекта для сравнения со стороны приматов она выбрала представителей нескольких крупных травоядных обезьян, сородичей бабуинов — Theropithecus, живших в тех же условиях и примерно в то же самое время. Результаты сопоставления оказались просто ошеломляющими. У крупных травоядных обезьян за весь период сравнения не было выявлено никаких признаков увеличения объема мозга, тогда как у гоминидов обоих ветвей Homo (ergaster и habilis) и Paranthropus (boisei) такие изменения имели место. Так, в тот период не только появилось несколько новых видов Homo и Paranthropus со значительно большим объемом мозга, но и, что весьма показательно, объем мозга значительно увеличился у всех гоминидов в пределах каждого вида. Последнее наблюдение находит еще более явное подтверждение в целом ряде навыков поведения, присущих общим предкам ветвей Homo и Paranthropus, но не свойственных их современникам-приматам. Относительное увеличение объема черепа у обоих ветвей гоминидов удивительно еще и в том смысле, что представители ветви Paranthropus с его характерными огромными челюстями считались чистыми вегетарианцами, питавшимися исключительно растительной пищей. За период в миллион лет, рассмотренный Элтон в ее исследовании, средний объем мозга у гоминидов увеличился более чем вдвое — с 400 до 900 куб. см. Если сопоставить эру феноменального роста и увеличения объема мозга с развитием линии человека в более поздний период, бросается в глаза прерывность развития при переходе от древних гоминидов к человеку современного типа. Для сравнения достаточно сказать, что увеличение объема мозга между древнейшим Homo habilis, жившим примерно 2 млн. лет назад, и Homo rhodesiense, жившим 1,07—1,3 млн. лет назад[14], то есть за период около 700 тысяч лет, составило более чем 2,5 раза. В последующие же 1,2 млн. лет, несмотря на тот факт, что большинству типов человека, обитавших за пределами Африки, например, Homo erectus из Азии и неандертальцам, жившим в Европе, было присуще некоторое увеличение объема мозга, от достижения объема мозга современного человека их отделяло всего 6%. (Фактически же за последние 150 тысяч лет у человека современного типа имело место общее снижение объема мозга; см. рис. 0.2.) Таким образом, с физической точки зрения самый ранний период эволюции человеческого древа был наиболее бурным и драматичным. Эти результаты свидетельствуют о том, что наиболее ранний период нарастания неблагоприятных климатических факторов — с конца плиоценового периода и на протяжении климатического рубежа между плиоценовым и плейстоценовым периодами — происходил выборочный рост и увеличение объема мозга у ряда новых видов гоминидов, тогда как у приматов, живших в тех же условиях, подобной эволюции мозга не наблюдалось. Что же это означает? Во-первых, это — аргумент в пользу точки зрения о том, что все эти гоминиды принадлежали к ветвям Homo и Paranthropus и, следовательно, их общий предок обладал рядом новых навыков поведения, способствовавших активному росту мозга, который стал характерным для этих видов к началу периода глобального похолодания. Другими словами, прямоходящие обезьяны, жившие около 2,5 млн. лет назад, уже могли нести в своих генах семена будущего невероятно активного развития мозга. Во-вторых, этот факт ставит под сомнение обоснованность гипотезы о роли мяса, хотя Элтон выдвинула в ее поддержку ряд аргументов, сводящихся к тому, что представители вида Paranthropus вовсе не были ни строгими вегетарианцами, ни примитивными существами, не способными создавать орудия, которые помогли бы им добывать пищу из целого ряда ее новых источников[15]. В-третьих, естественный отбор, направленный на увеличение объема мозга, по всей видимости, наиболее быстро и интенсивно протекал на заре существования рода человеческого — свыше 2 млн. лет тому назад. Что касается теории, будто «мозговитых охотников породило мясо», то с ней связана и другая проблема. Высшие приматы были далеко не единственными охотниками (т.е. хищниками), кормившимися на необозримых просторах африканских саванн. Однако мы почему-то не видим львов, гиен или африканских диких собак, бродящих по вельду[16] с огромными черепами. Справедливости ради надо признать, что эти прирожденные плотоядные имеют относительно крупный мозг и отличаются большей смышленостью и расчетливостью, чем их жертвы, однако они не идут ни в какое сравнение ни с человеком, ни даже с обезьянами. Это сильные, мускулистые существа, настоящие мастера клыка и когтя. В отличие от них, гоминиды всегда отличались интеллектуальной и физической гибкостью в использовании всеновых и новых источников пищи. Мы и сегодня едим массу растительной пищи, в том числе всевозможные фрукты, коренья, листья, семена, орехи и ягоды. Наши руки и зубы развиты куда лучше, чем клыки и лапы животных, специализирующихся на какомлибо одном виде пищи. Единственная физическая особенность, развившаяся у наших рук в процессе эволюции, помимо оппозиции большого пальца, заключается в том, что контроль за их манипуляциями осуществляет определенная область мозга. Значительное число поведенческих различий в рационе питания, резко отличающих нас от плотоядных, в сущности, представляют собой черты, общие для нас и наших ближайших сородичей-приматов — шимпанзе. Мы, как и они, признаем особую социальную значимость совместной охоты. Интереснейший ряд видеофильмов, запечатлевших сцены охоты шимпанзе на обезьян-колобусов в Африке, позволил выявить различия в поведении охотящихся шимпанзе и плотоядных хищников. Мы узнали, что эффективность охоты у этих небольших отважных приматов гораздо выше, чем у львов. Добытое ими мясо, хотя шимпанзе очень высоко ценят его, все же не является главной составляющей их рациона. При этом охотятся далеко не все стаи шимпанзе, и не все участники охотящейся стаи действительно едят мясо. Те из них, которые принимают участие в мясной трапезе, — либо верные исполнители, либо самки, с которыми доминирующие самцы-охотники намерены совокупиться. Таким образом, охота у шимпанзе — это скорее дело престижа, чем общей стратегии выживания, как это имеет место у людей. Сексуальные предпочтения говорят о том, что сильные самцы-охотники выбирают более надежные объекты передачи генов. Нам хорошо известно, что средства привлечения партнеров, обусловленные сексуальным отбором, могут быть весьма разными — от павлиньих хвостов до крупного черепа. Однако оставим вопросы сексуального отбора. Я хотел бы подчеркнуть, что нам следует более пристально понаблюдать за поведением наших ближайших сородичей по биологическому древу. История эволюции приматов на протяжении последних 10 млн. лет — это не история жвачных млекопитающих, которые однажды вздумали отказаться от растительной пищи и начали питаться вегетарианцами, становясь все более и более дерзкими. Это — история эволюции разумных всеядных, наделенных крупным мозгом и обитающих в лесах, где они смогли обрести физическую гибкость, сохранившуюся и после того, как эти существа переселились в другую среду обитания. У них сохранилась удивительная пластичность всех пяти пальцев, а зубы их в большинстве случаев уменьшились, а не увеличились. По какой-то причине численность одной из групп приматоподобных предков обезьян и людей резко увеличилась. Превосходно чувствуя себя в привычной среде обитания, они охотно употребляли большинство видов растительной пищи, которые давали им леса. Проявляя всеядность, они экспериментировали с диетой, пробуя употреблять в пищу более мелких, чем они сами, животных. В процессе борьбы за сексуальных партнеров охота могла стать средством укрепления собственного престижа в рамках сложившейся культурной практики. Будучи существами смелыми и контактными, они легко адаптировались к любым условиям, однако ни шимпанзе, ни наши далекие предки никогда не отказывались ни от гибкости рациона и осознания важности всеядности для выживания, ни от особой роли социального взаимодействия, благодаря которым они так успешно приспосабливались к среде обитания. Единственной и наиболее важной особенностью физического облика, унаследованной нашими предками от австралопитеков, явилась уникальная среди всех прочих млекопитающих способность постоянно передвигаться на двух ногах. Независимо от того, было ли это фактором адаптации к передвижению в саваннах, поросших высокими травами, необходимостью охлаждения массивного черепа и мозга или, что более вероятно, желанием освободить руки, этот переход совершился за миллионы и миллионы лет до того, как началось постепенное развитие и увеличение объема их мозга. Когда, примерно 2,5 млн. лет тому назад, произошло серьезное глобальное ухудшение климата, поведение и физический облик наших предков оказались вполне подготовленными к новому этапу эволюции. Руки у наших предков были свободны, голова стала крепкой и прохладной, а их разум, сообразительность и готовность к сотрудничеству в освоении новых источников пищи, включая мясо, оказались как нельзя более кстати. Засушливый климат всего лишь сместил вектор гастрономических пристрастий приматов, вынудив их уменьшить долю растительной пищи в своем рационе. Вместо того чтобы подражать крупным представителям семейства кошачьих, отращивая, подобно им, огромные клыки и усиливая и без того мощные челюсти, чтобы превратиться в настоящих плотоядных, приматы сделали то, что уже не раз делали в прошлом: прибегли к помощи мозга и рук. Причем происходило это на фоне давних традиций приспособляемости и социального взаимодействия. Новые навыки поведения, связанные с быстрым развитием мозга и появившиеся около 2,5 млн. лет назад, совпали с началом плейстоценовой ледниковой эпохи. Этот новый комплекс поведенческих навыков обладал громадным потенциалом, позволившим справиться с массой неблагоприятных климатических факторов. О том, что все эти факторы были налицо около 2,5 млн. лет тому назад, прежде чем появились первые люди, свидетельствует быстрый рост мозга, имевший место не только у предка человека, но и у его ближайшего сородича из рода Paranthropus.
Все новые и новые модели
Хотя по просторам африканских саванн примерно 2,5 млн. лет тому назад продолжали бродить и другие виды мыслящих приматов, в том числе — несколько видов рода Раranthropus, нас интересуют исключительно представители рода Homo, то есть древнейшие люди. Эти люди были представителями новой концепции эволюции сразу в нескольких отношениях: не только благодаря более крупному объему мозга, смешанной диете и более мелким зубам, но и благодаря способности усваивать адаптационные навыки, включая изготовление первых каменных орудий представителями самых древних видов человека. Если в качестве отправной точки выбрать людей вида Homo habilis, то Homo erectus можно считать видом, определившим направление развития, чем-то вроде знаменитой модели «Форд-Т» — родоначальницы нового класса. Оказавшись типом даже более успешным, чем «Форд-Т», они занимали доминирующее положение на нашей планете более полутора миллионов лет. Люди вида Homo erectus с их суровыми, угрюмыми лицами, плоским носом и быстро увеличивающимся объемом черепа очень напоминали человека современного типа, можно сказать — с ног до головы.
 Фото 2. Реконструкция головы Homo erectus. Эти люди с воинственным лицом и увеличенным объемом мозга покинули Африку, воспользовавшись первой предоставившейся возможностью, с успехом доминируя па планете на протяжении 2 млн лет.
Фото 2. Реконструкция головы Homo erectus. Эти люди с воинственным лицом и увеличенным объемом мозга покинули Африку, воспользовавшись первой предоставившейся возможностью, с успехом доминируя па планете на протяжении 2 млн лет.
Они использовали каменные орудия: поначалу это были слегка подправленные камешки, а затем — гораздо более сложные каменные топоры. Их африканский предок, Homo ergaster, стал первым человеком, покинувшим Африку примерно 1,95 млн. лет назад, чтобы стать уже в Азии представителем вида Homo erectus. Люди этого вида были ростом несколько меньше нас и быстро расселились по необозримым просторам Средней Азии, России, Индии, Дальнего Востока и Юго-Восточной Азии, всюду принося с собой навыки изготовления так называемых орудий из мелкого камня[17]. Существуют противоречивые утверждения относительно того, что в ту же эпоху продолжали существовать и их предшественники на генеалогическом древе человеческой эволюции — люди вида Homo habilis. Однако куда более достоверны свидетельства того, что представители всех будущих видов и ветвей рода человеческого вышли из Африки в первый же промежуточный период потепления между ледниковыми эпохами. Люди вида Homo erectus занимали доминирующее положение на нашей планете на протяжении примерно миллиона лет до тех пор, пока новая ужасающая полоса ледниковых периодов около 1 млн. лет тому назад не привела к иссушению большинства территорий африканского континента. В результате возникло новое, более приспособленное семейство людей. Первым представителем этого семейства в Африке был так называемый Homo rhodesiense. Представители этого вида, не уступавшие нам в росте и имевшие объем мозга около 1250 куб. см, использовали более сложные каменные орудия так называемого ашолийского типа, названного так по названию французской деревни, в окрестностях которой были впервые найдены образцы этого стиля. Орудия ашолийского типа включали в себя крупные плоские камни, обтесанные с обеих сторон, чтобы получился ручной топор с округлой (каплеобразной) рубящей кромкой. Эти выходцы из Африки впервые предприняли переселение в Европу и, возможно, в Китай во время первого глобального потепления, наступившего около полумиллиона лет тому назад, и принесли с собой свою ашолийскую технологию[18]. Затем, примерно 350 тысяч лет назад, наступил очередной суровый ледниковый период, в результате которого около 300 тысяч лет назад на исторической сцене в Африке появился новый вид. Одним он известен под своим архаическим названием — Homo sapiens[19], другим — под названием Homo helmei. Чтобы избежать путаницы, я в дальнейшем буду использовать второе название (Homo helmei). Представители этого вида, имевшие густые брови и такой же рост, как и современные люди, а также несколько больший объем мозга, составлявший у них около 1400 куб. см, представляли собой нулевую, заключительную фазу увеличения объема мозга. Как мы увидим в Главе 2, люди этого типа тесно связаны с началом одной из важнейших революционных эпох в технико-технологической области за всю историю человечества, известной под названием Среднего палеолита. Некоторые ученые в своих работах заходят настолько далеко, что утверждают, что, если бы эти густобровые создания появились на свет в современной семье, они вполне вписались бы в наше общество[20]. Взглянув на долгий и протяженный путь после исхода из Африки во время стабильного глобального потепления, мы увидели бы, как представители Homo helmei примерно 250 тысяч лет назад расселяются по просторам Евразии. Люди Homo helmei, вполне возможно, стали предками вида Homo neanderthalensis[21] — знаменитых неандертальцев, живших в Европе и Азии (см. табл. 3) и, возможно, имевших в ту эпоху ряд близких родственников в Индии и Китае. Между тем семейство видов человека, к которому принадлежали и наши предки, в те времена пока что оставалось в Африке и имело целый ряд физических отличий от своих двоюродных братьев-неандертальцев, обитавших в Европе[22] . Что касается нашего вида, Homo sapiens, то он появился более 170 тысяч лет назад. Его предком был некий вымерший вид, общая численность которого с учетом всех ледниковых периодов не превышала 10 тысяч особей[23]. Хотя Homo sapiens тоже совершил исход из Африки в район Леванта во время следующей паузы между ледниковыми периодами, наступившей около 120 тысяч лет тому назад, данные генетических исследований свидетельствуют о том, что его представители полностью вымерли там во время следующего ледникового периода. (Левант — термин несколько устаревший, но весьма уместный в данном контексте. К Леванту относились современные Сирия, Ливан, Израиль, Палестина и Иордания, то есть практически весь Средиземноморский Ближний Восток, за исключением Египта и Турции.) Когда люди современного типа примерно 70—80 тысяч лет назад начали активно расселяться из Африки по всему остальному свету, в Евразии еще жили представители других ветвей человека. Так, европейский неандерталец и, вполне возможно, азиатский Homo erectus еще жили каких-нибудь 30 тысяч лет назад, однако никаких следов их генофонда в организме современного человека не обнаружено. Весьма показательно, что как неандертальцы, так и люди современного типа, жившие еще до последнего ледникового периода (примерно 20—30 тысяч лет тому назад), имели значительно больший объем мозга, чем мозг современного человека[24]. Порой складывается впечатление, что магический эффект увеличения объема мозга, присущий ледниковым периодам, утратил свою чудодейственную силу задолго до нашего рождения на ветви подвида Homo helmei (рис. 0.2). Быть может, дальнейший рост объема мозга начал представлять серьезную угрозу. Так или иначе, но объем мозга сам по себе перестал играть роль фактора, определяющего успех вида, и на первый план вышло умение пользоваться мозгом. А это повлекло за собой появление новых навыков и культурных инноваций. После того как мы, люди, покинули просторы Африки, несмотря на то что дальнейшее увеличение объема мозга полностью прекратилось, климатические факторы продолжали играть доминирующую роль в процессе расселения человечества и появления все новых и новых изобретений и открытий вплоть до нашего времени. Не будет преувеличением сказать, что волны технических инноваций человечества, распространявшиеся по просторам Евразии начиная примерно с 80 тысяч лет тому назад, были в большей степени результатами стрессов и преодоления их, чем новых биологических факторов, способствовавших развитию человеческого биокомпьютера (мозга).
 Рис 0.2
Рис 0.2
Так, например, появление новых технических достижений и инноваций, прослеживаемое археологами на протяжении Раннего, Среднего и Позднего Верхнего палеолита, а также мезолита и неолита, неизменно совпадало с катастрофическим ростом влажности климата в Европе и миграциями населения на все новые и новые территории. Эти же факторы в Юго-Восточной Азии нашли свое выражение в активном развитии техники строительства лодок и судов и росте мореплавания, ставших ответом человечества на затопление обширных земель континентального шельфа в результате резкого подъема уровня моря. Таким образом, быстрое увеличение объема мозга явилось ключевым фактором, выделившим человека из всех прочих прямоходящих приматов, живших на Земле около 2,5 млн. лет назад. С тех пор объем нашего мозга претерпел ряд изменений и колебаний. Его увеличение не носило стабильного характера, причем большая часть роста объема мозга (в два раза) произошла еще у Homo erectus, жившего около 2 млн. лет назад. Другими словами, самое быстрое и радикальное увеличение и развитие мозга произошло у наших предков примерно 1,5 млн. лет тому назад, и с тех пор его темпы постепенно замедляются. Главный парадокс заключается в том, что сегодня бурными темпами идет накапливание все новых инноваций и поведенческих навыков, приобретшее в последние годы взрывной характер.
Идея Болдуина
Решение парадокса — противопоставления быстрого роста объема мозга в древности и культурно-технологического взрыва, переживаемого человечеством сегодня, — заключается в том, что человеческая культура, образно говоря, питает сама себя, чем и обусловлены все убыстряющиеся темпы ее развития. Как мы вскоре убедимся, история эволюции человеческой культуры — это отнюдь не виртуальная копия биологического древа человечества, на котором каждый последующий вид заметно прибавляет в разуме и сообразительности, что сразу же находит свое выражение в использовании все более сложных орудий. В отличие от биологической эволюции, движущие силы создания все новых и новых культурных достижений всегда развивались совершенно иным путем, и хотя наш мозг давным-давно перестал увеличиваться в объеме, наша культура продолжает и продолжает развиваться. В основе революционных изменений, переживаемых сегодня человечеством, лежит совместная эволюция культуры и генофонда человека. И хотя эта концепция выглядит достаточно простой, она способна преодолеть все наши этнические и расовые предрассудки. Механизмы, посредством которых поведенческие инновации или «новая культура» влияют на ход эволюции, впервые были сформулированы американским физиологом Марком Болдуином около ста лет тому назад[25]. Болдуин предложил особую, поведенчески обусловленную интерпретацию взглядов Дарвина на различные проявления эволюции. Интерпретация эта столь же проста, как утверждение, будто длинная шея у жирафа появилась в результате того, что его предкам приходилось поедать листья, растущие на верхних ветвях деревьев и кустарников. Болдуин утверждал, что гибкость моделей поведения и способность к обучению могли усиливать и изменять само действие механизмов естественного отбора. После того как новые, самостоятельно обретенные или заимствованные поведенческие навыки приводили к изменению среды обитания или условий жизни конкретного сообщества животных, естественный отбор мог отдавать предпочтение генетически обусловленным факторам поведения и физическим особенностям, благоприятствовавшим адаптации к новым условиям. Этот достаточно простой аргумент, известный как «коэволюция» или «генетическая ассимиляция», позволяет избежать сползания к давно развенчанной теории Ламарка о наследовании приобретенных признаков, сохраняя при этом забытое, но куда более здравое и впечатляющее ядро его идей. Коэволюция свойственна отнюдь не только истории развития нашего вида. Если погрузиться в толщу времени и пройтись назад по ветвям древа жизни, нетрудно заметить, что новые, целенаправленно развитые или, возможно, случайно приобретенные навыки поведения часто влекли за собой генетические изменения, которые определили последующий ход развития специфических черт внешнего облика, позволяющих использовать эти новые навыки. Все дарвиновские земляные вьюрки происходят от обыкновенных центрально-американских вьюрков, которые на протяжении своей долгой истории опробовали различные средства, позволяющие выжить и адаптироваться к меняющимся условиям среды обитания на Галапагосских островах. Впоследствии на основе этих изменений появились различные виды вьюрков, лучше приспособленные к выживанию благодаря усвоению этих новых признаков. Любопытно, что, подобно далеко отстоящим от нас представителям генеалогического древа позвоночных, в начале каждого поколения молодняк многих и многих видов заново усваивает и воспринимает так называемые врожденные навыки своих родителей. Так, мы знаем массу ситуаций, свойственных высшим позвоночным, когда сами родители принимают активное участие в обучении молодняка. Таким образом, на самом деле эти новые «развитые» особенности поведения передавались молодняку не через гены, а посредством общения с родителями и другими «учителями», то есть путем обучения. Впоследствии гены, благоприятствующие закреплению новых навыков, начали отбираться посредством биологической эволюции, позволяя представителям формирующегося вида эффективнее использовать эти факторы. Другими словами, гены и культурные аспекты коэволюционировали. Развитие культуры не обязательно тесно связано с генетической наследственностью. На протяжении эволюции большинства видов млекопитающих такое обучение элементам «культуры» было строго ограничено членами данного семейства или группы; в результате поведение представителей данной группы оказывалось тесно связанным с общностью генов. Однако необходимо подчеркнуть, что у многих социальных животных (животных, образующих стаи и имеющих иерархию) навыки выживания передавались и при контактах с представителями других социальных групп (стад, стай), которые далеко не всегда были родственниками. Таким образом, на каком-то этапе за последние несколько миллионов лет биологической эволюции приматов эволюция культуры их общения достигла определенной независимости от генетических кодов животных, являющихся ее носителями. Прибегнув к аналогии, можно сказать, что движущими факторами эволюции скрипки в равной мере могли быть и гильдия мастеров, делавших скрипки, и семейство мастеров, в котором секреты ремесла передавались от отца к сыну. О чем же все это говорит? Некоторые приобретенные, а не врожденные культурные традиции имеют такую географическую локальность, что их можно считать независимыми от генетических связей. Так, например, мы знаем, что японские макаки обычно моют потаты (сладкий картофель) в море. Это — местная культурная традиция, имеющая вполне конкретные исторические и географические корни, традиция, передававшаяся из поколения в поколение. Крайне маловероятно, что подобные особенности поведения могут зависеть от новых генетических факторов; однако, если следовать логике этого тривиального примера, можно сказать, что, если бы практика мытья свежих потатов обеспечивала вполне конкретные преимущества в выживании и именно такие мытые потаты являлись основным источником питания для данной местной популяции макак на протяжении многих поколений, тогда естественный отбор случайных генетических признаков у представителей этих будущих поколений мог бы тем или иным путем закрепить практику мытья свежих потатов. Это и было бы проявлением коэволюции[26]. Особенно выразительно географическая локализация приобретенных культурных навыков у высших приматов прослеживается у шимпанзе. В стаях шимпанзе возникали навыки создания примитивных орудий, носителями которых были как члены некой небольшой группы, так и представители соседних стай, не являющихся ее родственниками. Подобные технические навыки приобретаются культурным путем и не имеют генетической заданности и, следовательно, не обязательно должны быть распространены в других регионах. В определенный момент, возможно, еще до появления гоминидов, культура перешагнула через межвидовой барьер и стала достоянием различных видов приматов. Однако эволюция культуры, образно говоря, вступила в свой подростковый возраст задолго до этого момента и имела к тому времени длительную предысторию, развивавшуюся параллельно с генетической эволюцией. С точки зрения взглядов Болдуина, мы можем сделать любопытное наблюдение и высказать прогноз. Прогноз заключается в том, что, если сложное и многоуровневое общение требует высокоразвитого мозга, это означает, что появлению такого мозга должно было предшествовать длительное и более примитивное общение. Наблюдение же состоит в том, что возникновение письменности имело место около 5000 лет тому назад, а нотная запись была изобретена и вовсе гораздо позже, по историческим меркам — совсем недавно. Эти две кодифицированных системы внеустного общения отражают два высших достижения человеческого разума, однако нам и в голову не придет утверждать, что для того, чтобы ими пользоваться, должен появиться новый вид человека, имеющий особые гены.
Как развивался наш мозг и почему его объем сыграл столь важную роль?
Наиболее существенное различие между людьми современного типа и прочими животными связано именно с большим мозгом, характерным для человека. Здесь необходимо отметить несколько факторов. Действительно, объем мозга очень важен, но это еще далеко не все. Большой не обязательно означает более продуктивный. Так, например, у свиней и кабанов мозг гораздо больше по объему, чем у небольших хищных животных, например, диких котов. Люди, которым в раннем детстве по медицинским показаниям приходится удалять половину мозга, обладают практически нормальным человеческим интеллектом, хотя объем мозга у них не превышает 700 куб. см. Очевидно, значительное место в развитии способностей мозга принадлежит внутренним связям, и мы, люди, по числу внутримозговых связей далеко превосходим любых млекопитающих. Как же они возникли? Как правило, более крупное тело нуждается в большем по объему мозге. Грубо говоря, это обусловлено тем, что для контроля за более крупными органами и мускулами, составляющими крупное туловище, требуется более внушительный объем мозга или как минимум значительно больше внимания, уделяемого управлению всей этой глыбой мускулов. Подобная взаимосвязь между объемом мозга и пропорциями тела, хотя она и является прогнозируемой у большинства млекопитающих, не носит характера прямой зависимости. Если бы подобная зависимость имела место, то мыши, например, обладали бы куда меньшим — в пропорциональном отношении — мозгом, чем тот, который мы видим у них. У высших млекопитающих эта взаимосвязь становится еще менее прямолинейной, так как соотношение размеров тела и мозга у них искажено сразу по многим направлениям. К примеру, приматы во взрослом возрасте обладают пропорционально большим объемом мозга, чем любые другие млекопитающие, поскольку скорость роста их тела, начиная с самого раннего возраста, существенно ниже абсолютной скорости роста мозга. У людей билогические часы взросления мозга идут значительно медленнее, чем у приматов. Как известно, у всех млекопитающих рост мозга прекращается еще до того, как все функциональные факторы тела взрослого животного достигнут полного развития. Человек отличается от всех прочих приматов тем, что его биологические часы предусматривают развитие мозга значительно дольше, чем этого можно было бы ожидать исходя из пропорций тела, как это имеет место у приматов. В результате продолжительной пренатальной и детской стадий развития мозг у человека на этих этапах достигает объема около 1000 куб. см, что примерно соответствует объему мозга вымершего вида Gigantopithecus[27] . Другой несложный пример различий, контролируемых генами и присущих человеку, состоит в том, что отделы мозга, расположенные в задней части черепа человеческого эмбриона на ранних этапах его развития, в итоге вырастают сравнительно более крупными, чем у других приматов[28]. Именно этим объясняется тот факт, что у взрослых людей непропорционально велики мозжечок и кора головного мозга. Между тем известно, что именно эти два отдела мозга отвечают за координацию движений и высшую нервную деятельность. Генетические изменения, лежавшие некогда в основе этих важнейших факторов, по всей вероятности, были достаточно просты и касались нескольких генов, отвечающих за рост и развитие. В результате относительные различия в объеме разных отделов мозга приобрели выраженный характер. Все эти эффекты искажения пропорций генетически запрограммированы и «включаются» в человеческом эмбрионе на ранней стадии его развития, еще до того, как возникают и формируются большинство связей между клетками мозга. Увеличение объема коры головного мозга влечет за собой создание гораздо большей по объему нервной ткани, чем это необходимо для решения повседневных бытовых потребностей и обеспечения нормальной жизнедеятельности организма. Другими словами, у человека (и в значительно меньшей степени — у высших приматов) формируется огромный резерв объема коры головного мозга, не играющий никакой роли для нужд повседневной жизни. Если у эмбриона чрезмерное разрастание коры головного мозга происходит задолго до формирования взаимосвязей между различными отделами мозга, как это может повлиять на качество таких связей у человека в будущем? Ответ будет следующим: когда нервные клетки далеких друг от друга отделов мозга начинают прорастать друг в друга, при дальнейшем развитии эмбриона объем мозга играет важную роль в интенсивности и количестве таких связей, формирующихся как внутри самой коры головного мозга, так и вне ее, с остальными отделами головного и спинного мозга. Возникающее в итоге чрезмерное разрастание связей в коре головного мозга можно охарактеризовать как весьма влиятельное «министерство без портфеля», которое в самом буквальном смысле всюду имеет хорошие связи и агентов влияния во всех ветвях исполнительной власти. Необычно большое число внутрикорковых связей, в частности, предопределяет нетривиальное поведение, творческую одаренность и ассоциативное мышление. Увеличение же числа внешних связей коры головного мозга дает нам возможность непосредственно контролировать движение ядер клеток в стволе мозга, ответственном за речь. Раньше эти ядра контролировались подкоркой, так сказать, на режиме автопилота. И все это — результат «перенастройки» какой-нибудь полудюжины генов-регуляторов[29]. Большая часть подобной «гипертрофии» произошла задолго до того, как мы, люди, появились на исторической сцене. Простое сравнение объема мозга и размеров тела у древнейших людей показывает, что по мере эволюции вида Homo erectus эти изменения привели к невиданному развитию мозга. Итак, зная, что буквально считанные генетические изменения привели к громадному расширению функционального потенциала человеческого мозга, мы вправе вернуться к вопросу о том, какие же невиданные прежде изменения навыков поведения запустили 2,5 млн. лет тому назад процесс быстрого роста мозга?
Пища для разума или разговоры о роли пищи?
Видный физиолог Робин Данбар из Ливерпульского университета, сторонник эволюционной теории развития, высказал утверждение, что животные с относительно крупным мозгом способны помнить большое число особей в рамках социальной структуры своего вида и соответствующим образом общаться с ними. По его мнению, существами, обладающими теоретически наибольшей «социальной емкостью памяти», являются люди. По сравнению с другими видами животных мы, люди, можем выделить и запомнить в общей сложности более 300 человек — объектов для общения, причем эта способность присуща как людям современного типа, так и неандертальцам. На основе моего личного опыта должен признать, что хотя в школьные годы я, вероятно, знал в лицо более тысячи человек, эта цифра не идет ни в какое сравнение с числом людей, с которыми я сегодня поддерживаю регулярные контакты. Если перенести этот пример в более привычный контекст, надо сказать, что существуют вполне конкретные ограничения по уровню плотности населения, которое способна прокормить определенная площадь саванны. Исследования охотников-собирателей племени кунг в Южной Африке показали, что количество членов семейных групп в среднем составляло несколько десятков, а максимальное число жителей поселения в засушливый сезон не превышало сорока человек. Понятно, что социальные отношения в крупных общинах могут иметь более сложный характер, чем в малочисленных. Клайв Гэмбл, видный специалист по истории палеолита из Саутгемптонского университета, утверждает, что наши предки (а впоследствии и наши общественные группы) образовывали группы самой разной численности, выполнявшие различные функции. Так, в составе группы ближайших родственников, по большей части состоявшей из ядра семьи, насчитывалось не более пяти человек; численность более крупной и эффективной в функциональном отношении группы не превышала двадцати членов, и, наконец, наиболее многочисленная семейно-родственная община, члены которой не столь уверенно узнавали друг друга в лицо, могла насчитывать от 100 до 400 человек. Реальные возможности обмена навыками и материальными ценностями могли существовать лишь в двух первых группах, тогда как в третьей подобные действия уже были средством извлечения выгоды. Таким образом, эту третью группу саму по себе нельзя считать примером общительности, способствующей развитию мозга[30]. В то время как способность запоминать и узнавать в лицо большое число сородичей вполне может ассоциироваться с крупным мозгом, трудно допустить, что подобный эффект коллективизма мог послужить источником все новых и новых прорывов в развитии человеческого мозга за последние 2,5 млн. лет, особенно если контакты в рамкам коллектива сводились в основном к ухаживанию посредством вычесывания друг у друга из шерсти блох и вшей да обмена любезными гримасами. Время, которое приходилось тратить на более серьезное дело — поиски пищи, поневоле сводило подобные контакты к минимуму. Робин Данбар и Лесли Эйэлло установили, что в этом контексте наиболее доступным средством ухаживания в социуме первоначально были язык, речь[31], хотя та же речь служила и средством обмена информацией. Действительно, большинство из нас тратят значительную часть своего времени на общение в рамках социума. И все же мне трудно согласиться с утверждением, что дар речи — этот уникальный дар, которым наделены только мы, люди, — использовался в первую очередь как средство ухаживания и установления сексуальных контактов, нежели средство повышения эффективности общения (на охоте, при поисках пищи) и обучения наших отпрысков посредством сообщения им практической информации. Из робких собирателей, бродивших по просторам африканских саванн, род человеческий превратился в одного из наиболее грозных хищников, и произошло это задолго до того, как предки людей современного типа покинули Африку. Я предлагаю взглянуть на этот вопрос с другой точки зрения. Мне кажется, что именно дар речи был тем уникальным навыком, обладателями которого 2,5 млн. лет тому назад стали родственные виды Homo и Paranthropus, что и позволило им в самом тесном сотрудничестве друг с другом выжить во время засушливых циклов плейстоценовой ледниковой эпохи и тем самым обеспечить дальнейший рост и развитие мозга. Согласно сформулированной Болдуином теории коэволюции в результате «появления новых навыков, предшествующих адаптационным физическим изменениям», представители обоих видов должны были обладать некой исходной формой языка. Трудно возразить что-либо конкретное против того, что этот символический кодифицированный язык, а также его сложный синтаксис и те возможности для широкого общения, которые он открывал, способствовали быстрому развитию сообразительности и интеллекта. Проще говоря, вполне вероятно, что еще более 2,5 млн. лет назад мы обладали способностью общаться при помощи неких знаков, что это способствовало увеличению объема нашего мозга и что наш мозг рост и развивался до некоего порогового уровня, пока мы, совсем как малыш Слоненок из сказки Киплинга, у которого нос превратился в хобот, внезапно не обнаружили, что можем разговаривать друг с другом.
Образное мышление и язык: действительно ли они присущи только человеку?
Нет никаких сомнений, что общение между особями в той или иной форме возникло у животных в глубокой древности. Наиболее сложной и развитой формой общения у животных, бесспорно, являются вокативные, или голосовые, сигналы, необходимость издавать которые обусловила ряд физических изменений в организме человека. Вокативная речь имеет массу преимуществ по сравнению с простейшим языком жестов, не говоря уже о способности обмениваться сложными и даже отвлеченными идеями. Так, благодаря речи мы можем общаться в абсолютной темноте, пробираясь сквозь чащу леса и не видя человека, с которым говорим. Переходя на другой язык, нам гораздо проще скрыть свои истинные намерения, солгать и даже обвести вокруг пальца незнакомых людей. Дети начинают говорить заведомую ложь примерно в возрасте четырех лет. Некоторые ученые полагают, что склонность мужчин шутить в обществе женщин, заставляя последних то и дело смеяться, тоже могла быть в древности элементом сексуального отбора. Впрочем, язык, как и все прочие составляющие культуры, как бы заново создается и внутренне возрождается в сознании каждого ребенка, еще только учащегося говорить. Мы, современные люди, по праву отделяем себя от всех прочих видов наших ближайших сородичей в животном мире — приматов, живущих сегодня на Земле. К сожалению, мы никак не можем отказаться от сравнений качественного характера. Подобно тому, как мы склонны подчеркивать религиозные и этнические различия у представителей своего собственного вида, мы пытаемся выделить и сформулировать те факторы, которые отличают нас от приматов, чтобы определить рамки понятий «мы/не мы». Крайним выражением подобных претензий на исключительность и различия между современными расами можно считать термин унтерменш (нем. букв. «недочеловек» или «человек низшей расы»), использовавшийся нацистами применительно к народам, которые они подвергали преследованиям и истреблению. Мы убедили себя в том, что обладаем неисчислимыми интеллектуальными и манипулятивными навыками, которые, словно частокол, отделяют нас от всех прочих представителей животного мира. Всякий, кто пытается заглянуть сквозь этот частокол, неизбежно навлекает на себя упреки в «антропоморфизме». Тем не менее примерно с начала XX в. наш ближайший сородич в животном царстве, шимпанзе, привлекает к себе все более пристальное внимание исследователей, что позволяет извлекать из этого частокола предрассудков колышек за колышком. Однако устаревшие мифы об уникальности навыков человека отмирают с большой неохотой. Прежде бытовало убеждение, будто люди были единственными существами, умевшими пользоваться орудиями. Когда эта идея утратила былой авторитет, предрассудки, лежавшие в ее основе, были перетолкованы в том смысле, что люди — это единственные существа, способные совершенствовать орудия труда и охоты. А когда была отвергнута и эта версия, нам пришлось утешаться тем, что только мы, люди, способны совершать изобретения и открытия и делать орудия. Но шимпанзе вновь опровергли и этот миф. Большинство приводимых здесь фактов заимствованы из исследований Вольфганга Кёлера, проведенных в начале 1920-х гг. в колонии шимпанзе на острове Тенерифе. Кёлер, психолог по образованию, пошел гораздо дальше простой демонстрации того, что шимпанзе способны решить эту проблему. Ему удалось весьма элегантно доказать, что шимпанзе способны мыслить не только рационально, но и абстрактно[32]. К сожалению, в те годы нашлось мало ученых, способных по достоинству оценить результаты этих экспериментов. И потребовалось лишь невероятное терпение Джейн Гудолл и других исследователей, чтобы рассеять старые предрассудки ученых и широкой публики в отношении выводов Кёлера. Это произошло уже в конце XX в. Одним из самых удивительных открытий второй половины XX в. явилось получение убедительных доказательств того, что шимпанзе, представители неговорящих видов приматов, отделившиеся от наших предков на древе эволюции более 5 млн. лет назад, обладают потенциальной способностью говорить, находящейся в стадии формирования. Многих шимпанзе удавалось обучить общаться с человеком. Еще более поразительно, что они способны легко усваивать приобретенные навыки и использовать их в общении друг с другом посредством символического кодифицированного языка. Но самой яркой звездой этой истории по праву считается бонобо по кличке Канзи (бонобо — ближайшие сородичи шимпанзе, а некоторые черты их поведения скорее напоминают людей). Канзи научился общаться с окружающими при помощи сложного языка кодифицированных сигналов. Кроме того, он без труда научился понимать разговорный английский и даже правильно интерпретировать его синтаксис. Уровень его успехов отчасти был обусловлен тем, что он был бонобо (оказывается, бонобо в генетическом отношении ближе к человеку, чем другие виды шимпанзе), однако не исключено, что они были усвоены им в раннем детстве от своей приемной матери-шимпанзе, уроки «обучения» у которой он использовал с максимальной пользой. Эта самка шимпанзе, в свою очередь, была объектом обучения языковому общению и во взрослом возрасте пыталась освоить эти новые символы. Как известно, шимпанзе обладают способностью к абстрактному, символическому и рациональному (предметному) мышлению, а также могут совершать так называемые символические выводы и символические манипуляции, хотя вполне понятно, что в этих действиях они далеко уступают нам, людям. И тем не менее это — вопрос уровня развития. Что же касается языка, речи, то шимпанзе испытывают явные трудности, связанные с отсутствием вокативного (голосового) контроля, и либо недооценивают, либо вообще не понимают важности расширения средств невербального (неречевого) общения[33]. Удивительно, но лингвисты до сих пор упорно игнорируют важность выводов этих экспериментов. Чтобы понять причины этого упорства, мы должны признать дихотомический характер современных теорий происхождения речи и мышления. Начиная с середины XVIII в. в этой области параллельно сосуществуют две линии аргументов. Первая из них, в которой язык рассматривается как некое изобретение, была предложена еще в XVIII в. Этьеном Бонно де Кондильяком, философом эпохи Просвещения. Он утверждал, что разговорная речь развилась на основе языка жестов (langage d'action) и что обе эти системы представляли собой изобретения, возникшие первоначально на основе простейших ассоциаций. Точка зрения Кондильяка поддерживает концепцию культурной эволюции, и основные ее постулаты, естественно, в несколько модифицированном виде прослеживаются и в учении дарвинизма, и в трудах мыслителя середины XX в. Рональда Энглефилда, а в наши дни — в работах новозеландского физиолога Майкла Корбаллиса и ряда других ученых. Эта теория рассматривает язык жестов как систему, первоначально возникшую среди приматов, а затем получившую название конвенциональной или кодифицированной по мере эволюции самих навыков. Впоследствии вербальные сигналы, некоторые из которых присутствуют во «врожденном» арсенале приматов, выделились в отдельную группу, и на их основе была выработана сознательно организованная кодифицированная система общения. Эволюционные факторы способствовали развитию специального голосового аппарата и отдела мозга, расположенного рядом с отделом, отвечающим за язык жестов. Этот речевой центр часто называют подмозолистым полем (обонятельным полем Брока)
Первое прикосновение к дару речи?
Другая точка зрения на корни зарождения языка, преобладающая в наши дни, носит почти креационистский характер в своем отрицании процесса эволюционного развития. Согласно ей, язык или, точнее говоря, произносимое слово рассматривается как некий феномен, внезапно появившийся от 35 до 50 тысяч лет тому назад в качестве своего рода Большого взрыва. По мнению Ноама Хомски, а впоследствии и Стивена Линкера, способность произносить слова и пользоваться синтаксисом была заложена в нашем мозге на генетическом уровне в качестве особого центра или органа речи. Эта точка зрения на язык вполне соотносима с давней идеей о том, что логическое или рациональное мышление неким образом зависит от речи, слов. Эта концепция восходит к учению Платона; она была весьма модной среди интеллектуалов XIX в., таких, как Якоб Гримм («Животные не говорят, потому что они не способны мыслить») и Макс Мюллер («Язык — это наш Рубикон, и никому из дикарей не дано преодолеть его», а также «Без речи не может быть разума, а без разума нет речи»)[35]. Креационистский постулат о резком качественном прорыве в мышлении человека получил дальнейшее развитие в современной интерпретации памятников искусства эпохи Верхнего палеолита в Европе. Резные фигурки и наскальные рисунки на сводах пещер, созданные, согласно принятой датировке, более 30 тысяч лет назад, с этой точки зрения можно рассматривать как первое наглядное сочетание символического и абстрактного мышления, а также языка. Однако зрелая сложность древнейших наскальных рисунков в пещере Шове на юге Франции опровергает эту точку зрения[36]. В любом случае подобная европоцентрическая интерпретация игнорирует бесспорные свидетельства того, что жители другого полушария, австралийцы, создавали свои наскальные рисунки в ту же эпоху, к которой принято относить древнейшие произведения жителей Европы. Поэтому было бы абсурдом утверждать, что и австралийцы, и обитатели других частей света обладали первыми навыками речи и живописи благодаря общению с европейцами. У нас, как сможет убедиться читатель этой книги, есть все основания полагать, что общий африканский прапредок современного человека уже владел навыками речи, изобразительного искусства и символической передачи мыслей задолго до того, как будущие родоначальники человеческих рас около 80 тысяч лет тому назад покинули просторы Африки. Другая проблема, связанная с креационистской теорией, или уподоблением внезапного появления уникальных способностей у человека Большому взрыву, заключается в том, что существуют вполне реальные доказательства того, что неандертальцы имели практически такое же строение голосовых органов, как и мы. Для голосового аппарата неандертальцев были характерны наличие такой же подъязычной кости, увеличенного грудного (торакального) отдела спинного мозга и увеличенная щель для подъязычного нерва. Согласно гипотезе Болдуина, эти атрибуты указывают, что предок неандертальцев (как и наш собственный предок), Homo heidelbergensis[37], еще свыше 500 тысяч лет тому назад обладал даром членораздельной речи. Поскольку Homo heidelbergensis также имел увеличенную щель для подъязычного нерва и некоторые другие важнейшие анатомические элементы голосового аппарата, вполне возможно, что сказанное можно отнести и к более древнему виду — Homo erectus, у которого обнаружены признаки асимметрично развитого мозга. Последний фактор считается исключительно важным для возникновения феномена языка. По мнению некоторых антропологов, в останках черепов вида Homo habilis обнаружена зона, напоминающая подмозолистое поле Брока. Это подтверждает точку зрения о том, что процесс целенаправленного увеличения объема мозга, необходимого для возникновения речи, начался еще более 2 млн. лет тому назад[38]. Эти анатомические факторы уводят нас в тьму времен, к первым людям и поразительному увеличению объема мозга у видов Homo и Paranthropus. И если вообще право мерно говорить о большом взрыве среди примитивных гоминидов, то он мог произойти только тогда. Тим Кроу, профессор психиатрии из Оксфордского университета, утверждает, что можно проследить черты двух тесно связанных мутаций Y-хромосомы, происшедших через некоторое время после разделения ветвей шимпанзе и человека. Есть основания полагать, что одна или даже обе этих мутации могли быть связаны с церебральной асимметрией и, возможно, возникновением языка. А если это так, то мы вправе предположить, что первая из этих мутаций Y-хромосомы произошла у общего предка Homo и Paranthropus, а вторая — у Homo erectus, поскольку именно у представителей этого вида обнаружены первые признаки церебральной асимметрии[39]. Новейшие исследования в области нейрофизиологии с использованием целого ряда современных технологий, включая прямое сканирование, еще более ослабили позиции биологического детерминизма во взгляде на возникновение и эволюцию мышления и языка. Сегодня мы знаем, что синтаксис различных типов языка связан с разными отделами мозга. Дело в том, что синтаксис языка не является чем-то врожденным. Он осознается маленькими детьми, которые, по сравнению со взрослыми, изучающими незнакомый язык, обладают куда большей гибкостью и способностью подсознательно улавливать правильный смысл синтаксических построений. Люди — далеко не единственные существа, пережившие критический период в процессе развития после того, как они усвоили первые навыки языка. Тот же феномен можно наблюдать не только у приматов, но и у разного рода «поющих» животных, будь то птицы или киты. Сложные, часто совершенно уникальные «песни», исполняемые этими животными в поздние годы жизни, воспринимаются ими в процессе обучения в раннем детстве, интерпретируются на свой лад и сохраняются в памяти. Более того, результаты исследований показывают, что речевой центр отнюдь не обязательно ограничивается каким-то одним или несколькими отделами мозга[40]. Эти нейрофизиологические исследования позволили предложить альтернативу теории эволюции языка, выдвинутой Хомски. Такая альтернативная теория включает в себя идеи Кондильяка, Энглефилда и Корбаллиса. Согласно ей, разговорный язык, как и навыки изготовления орудий, был изобретен единственно и только приматами, что способствовало быстрой эволюции их мозга и голосового аппарата. Однако, будучи изобретением культурного плана, язык начал жить собственной жизнью за пределами нашего тела, в рамках обособленных сообществ — носителей речевой культуры. Уникальное сочетание лексических и синтаксических составляющих языка, такого, например, как французский, является культурным достоянием всего франкофонного сообщества, а вовсе не результатом некоего биологического аспекта, связанного с принадлежностью к французской нации. Любой язык и его синтаксис, передаваясь от поколения к поколению, постоянно адаптируется к новым реалиям и еще только формирующемуся сознанию каждого нового поколения молодежи, которая учится говорить на нем. Таким образом, из всего многообразия интеллектуальных и практических навыков, выделенных философами в качестве решающего отличия человека от шимпанзе, единственный, сохранившийся с глубокой древности, — это, бесспорно, человеческая речь. Да, разумеется, между интеллектуальными способностями современных людей и наших далеких предков существует громадная количественная разница, однако интеллект человека достиг того невиданного расцвета, который произошел около 35 тысяч лет тому назад у европейцев, живших в эпоху Верхнего палеолита, далеко не сразу и не на пустом месте. Для этого ему потребовалось пройти длительный — более 4 млн. лет — процесс эволюции. За последние 2 млн. лет предки людей эволюционировали от прямоходящих приматов, научившись активнее пользоваться мозгом, но в этом им помогала коэволюция речи и языка, способствовавшая развитию объема мозга. И, подобно гибкому и удобному хоботу малыша Слоненка из притчи Киплинга, резко расширившийся диапазон возможностей нашего мозга с точки зрения манипулирования символическими представлениями и понятиями позволил нам перейти к решению множества сложных задач помимо речи. Тот факт, что мы способны рассуждать о происхождении и устройстве Вселенной, и даже начинаем изучать ее просторы, свидетельствует о том, что гибкость нашего интеллекта и его открытость к восприятию новых идей практически безграничны. Как мы уже знаем, циклы сильнейших засух, поражавших всю Африку, чередовавшиеся с периодами оледенения, около 2 млн. лет тому назад способствовали увеличению объема мозга человека. То же самое наблюдалось и у нашего ближайшего сородича — вида Paranthropus, но никак не коснулось крупных обезьян, путешествовавших по просторам саванн. Это свидетельствует о том, что уже в ту эпоху существовали некие навыки поведения, присущие только этим видам гоминидов (человекообразных приматов). У целого ряда ученых возникла тенденция рассматривать быстрое увеличение объема мозга как особенность, появившуюся у человека совсем недавно, но если обратиться к реальным фактам, придется признать, что дело обстояло как раз наоборот. Наиболее быстрый пропорциональный рост объема мозга имел место в период от 2 до 1 млн. лет тому назад. К моменту же появления на исторической сцене человека современного типа увеличение объема мозга резко замедлилось. Зато имел место любопытный парадокс: взрывной рост поведенческих навыков, число которых нарастало в геометрической прогрессии. И тогда, вместо того чтобы признать очевидный факт, что человеческая культура питает сама себя, создавая и регулируя темпы собственного развития, многие антропологи, археологи и лингвисты выдвинули предположение, будто в развитии и функционировании нашего мозга за последние 100 тысяч лет произошел некий «генетический» сбой, в результате которого якобы появился новый тип человеческого мозга, обладающего новыми возможностями. Некоторые пошли еще дальше, утверждая, что наиболее вероятным кандидатом на роль этой новой и уникальной способности следует считать именно язык[41]. На мой взгляд, в этих утверждениях сквозит какая-то извращенная, совершенно не дарвинистская логика. Сложная кодифицированная коммуникативная система, или «язык», — это, вне всякого сомнения, важный и во многом уникальный элемент поведения. Насколько же проще и примитивнее был тот язык, благодаря которому наши предки стали выделяться из всех прочих крупных приматов, обитавших в саванне, и который 2,5 млн. лет назад дал первичный импульс эволюции нашего мозга... Тот самый язык, благодаря которому они могли лучше понимать друг друга и более эффективно приспосабливаться к нарастанию неблагоприятных климатических факторов... Я уже касался темы генетики, которая занимает в этой книге достаточно видное место. Основную часть ранней истории человечества за последние 2,5 млн. лет можно реконструировать благодаря объединению выводов исследований ископаемых костных останков и изучения климатических периодов. Все виды человека, кроме одного, вымерли, причем некоторые из них — очень давно, поэтому в данном исследовании мы не учитываем их гены. Это не означает, что генетика не в силах поведать нам ничего существенного о темных эпохах эволюции человечества, предшествовавших выходу на арену истории людей современного типа. В 1970-е годы некоторые генетики начали применять сырые иммунологические средства для замеров аналогий уровня содержания протеина у представителей разных видов и установили, что человек и шимпанзе являются даже более близкими сородичами, чем это считалось прежде. Это заявление в свое время было встречено в штыки, но затем, когда методы сравнительного исследования переключились с иммунологии на демонстрацию базового генетического сходства, а затем и на замеры конкретных генетических различий, их авторы были оправданы и реабилитированы. В ученой среде созрело понимание того, что мы, люди, гораздо ближе к шимпанзе, чем к другим крупным приматам — гориллам и орангутангам, с которыми пути наших предков на генеалогическом древе разошлись немногим более 5 млн. лет назад[42]. Было бы несправедливо утверждать, что мы, люди, вообще не унаследовали никаких генов от исчезнувших видов гоминидов. Наоборот, большинство генов в наших клетках как раз унаследовано нами от вымерших видов гоминидов и приматов. Некоторые гены человека встречаются в останках некоторых видов приматов, вымерших задолго до нашего появления на Земле. Не так давно ученые выделили слабые фрагментарные следы митохондриевой ДНК в целом ряде костей неандертальца, и сегодня они практически в состоянии ответить на главный вопрос: в каком именно родстве мы с ними находимся и присутствуют ли хотя бы некоторые их гены в клетках людей современного типа? Впрочем, в этой книге нас в первую очередь интересует подлинная революция в понимании генетической предыстории человечества за последние 200 тысяч лет. Новые открытия в области генетики вторглись в ту туманную и неизученную сферу, в которой прежде доминирующее положение занимали маловыразительные коллекции каменных орудий из Европы и Африки да немногочисленные фрагменты скелетов, плохо поддающихся датировке. Прежде чем обратиться к детальном рассмотрению следов генов, на мой взгляд, будет небесполезно познакомиться с некоторыми идеями, связанными с генетической наследственностью и ее принципами. Эти концепции по большей части достаточно просты и связаны с нашими расхожими представлениями о наследственности, однако их нередко истолковывают в превратном свете, что объясняется либо сознательным обманом, либо тем, что они изложены на невразумительном «научном» жаргоне.
Секреты горошин
Идея генетической наследственности знакома людям с тех пор, как им удалось одомашнить целый ряд видов диких животных и растений. Скотоводы издревле предпринимали целенаправленные попытки устранять в стаде такие нежелательные признаки, как чрезмерный рост или агрессивность поведения[43]. Селекция зерновых культур осуществлялась путем отбора наиболее крупнозерных и урожайных сортов. Однако большая часть так называемых фермерских навыков была основаны на ошибочных посылках, хотя последние и носили функциональный и практичный характер. Интерес к выяснению точных механизмов и законов, определяющих работу факторов наследственности, резко возрос в середине XIX в. после выхода в свет фундаментального труда Чарльза Дарвина «Происхождение видов»[44], однако по сути понимание принципов наследственности, изложенное Дарвином, оказалось немногим более глубоким, чем у его предшественников. Зато другой его современник, австрийский священник и ботаник Грегор Иоганн Мендель, живший в XIX в., впервые сформулировал логические принципы осмысления того, каким образом осуществляется наследственная передача тех или иных наследственных признаков. Его выводы были основаны на обширных математических расчетах принципов наследования цвета и формы гороха. В принципе Мендель показал, что для передачи любых физических особенностей, или признаков, например, цвета цветков, любое растение гороха использует два гена (правда, он использовал здесь другой термин), которые и определяют выраженность или интенсивность признака. Вариативность преобладания тех или иных генов и обусловливает проявление признаков у растения. В процессе полового размножения каждое из растений-родителей способно передать своему потомству только один из этих генов. Таким образом, растение наследует от каждого из своих родителей сложную смесь признаков, и комбинированный эффект этих двух генов и определяет физические признаки очередного поколения. Поскольку любой из родителей может иметь функциональные гены различных типов, например, наследовать разное число лепестков применительно к каждой паре генов, но реально объектом произвольного выбора может стать только один из них, пропорциональные доли различных типов потомства в своей совокупности составляют картину, которую можно уверенно прогнозировать, зная признаки, которыми обладает каждое из растений-родителей. Таким путем Мендель сумел показать, что наследование признаков осуществляется путем передачи дискретных наборов информации. И вариативность потомства определяется точным, но произвольным смешением этих наборов, или генов, в каждой особи, будь то растение или человек. Мендель приложил немало труда, отбирая наиболее простые, универсальные и легко распознаваемые признаки и изучая их на индивидуальном уровне. На деле же выраженность некоторых физических признаков определяется, мягко говоря, не одним геном, и поскольку мы являемся достаточно сложными организмами, в наших клетках насчитывается около 30 тысяч пар функционирующих генов. Таким образом, заметные невооруженным глазом различия между единокровными отпрысками одной и той же семьи являются не результатом некоего сбоя в процессе наследования, а возникают в связи с тем, что в процессе полового размножения принимает участие большое число генов, образующих произвольные сочетания. А поскольку число генов и тем более их комбинаций крайне велико, это создает невероятно широкий потенциал для вариантов. Наоборот, удивительное сходство близнецов показывает, насколько точной может оказаться трансформация наследственности в физической оболочке. Выдающимся достижением таких ученых, как Уотсон, Крик и Розалинда Франклин, стал осуществленный ими перенос постулатов открытия Менделя в область биохимии, или молекулярной биологии, как сегодня принято называть эту сферу биохимии.
Картонные ключи к тайнам жизни
«Мы все — производные наших генов». Секретные ключи к этому эдуардовскому трюизму сформулировали и перенесли на простые листы картона двое ученых-первооткрывателей — Джим Уотсон и Фрэнсис Крик. Произошло это в 1953 г.[45]. Их «ключи» представляли собой диаграммы четырех химических соединений (базовых нуклеотидов), уникальные взаимосвязи между которыми, зафиксированные на двойной спирали дезоксирибонуклеиновой кислоты (ДНК), заключают в себе кодированный ключ ко всему живому на Земле. Эти картонные ключи позволили раскрыть секрет механизма, соединяющего постулаты Менделя с теорией эволюции путем естественного отбора, сформулированной Дарвином в его знаменитом труде «О происхождении видов». Уотсону и Крику удалось с исчерпывающей точностью объяснить, каким образом тысячи уникальных свойств, варьирующихся у разных особей, передаются из поколения в поколение. Короче говоря, это открытие явилось едва ли не самым выдающимся достижением биологической мысли в XX в. В каждой из клеток нашего тела присутствует невероятно длинная спираль ДНК. Она состоит из генов. Спираль эта хранит, воспроизводит и копирует все индивидуальные свойства нашего организма — нашу генетическую наследственность. Спираль ДНК хранит коды протеинов (белков) — этих строительных кирпичиков, из которых состоит наше тело. Коды каждого из белков «записаны» посредством различных сочетаний четырех химических соединений, так называемых базовых нуклеотидов (обозначаемых буквами A, G, С и Т), которые подают все указания, связанные со строительством нашего тела. Мы наследуем ДНК каждого из наших родителей, и поскольку и от того и от другого мы получаем уникальные сочетания генов, каждый из нас обладает спиралью ДНК слегка видоизмененной, или, лучше сказать, уникальной формы. Наша собственная ДНК — это как бы отпечаток пальца, но только на молекулярном уровне. При половом размножении ДНК родителей копируется и передается эмбриону в равных пропорциях. Важно понимать, что, хотя большая часть ДНК каждого из родителей при воспроизводстве передается потомкам по отдельности, небольшие фрагменты генов каждого из них в каждом поколении перемещаются и перемешиваются друг с другом. Однако подобное смешение — это отнюдь не произвольное массовое смешение генов, постулированное Менделем, а крохотные взаимопереходы, дублирование, выбросы и обмен между материнской и отцовской составляющими ДНК их ребенка. Это явление обозначается специальным техническим термином — рекомбинация[46]. К счастью для исследователей ДНК, существуют два небольших участка нашей ДНК, которые никогда не подвергаются рекомбинации. По фрагментам спирали ДНК, не подвергшимся рекомбинации, гораздо удобнее изучать признаки вида в далеком прошлом, поскольку информация, заключенная в ней, не претерпевает искажений при передаче от поколения к поколению. Эти два фрагмента спирали ДНК получили название митохондриевой ДНК и нерекомбинированной части Y-хромосомы.
Митохондриевая ДНК: ген Евы
Было бы не совсем правильно утверждать, что ровно половину нашей ДНК мы получаем от своего отца, а вторую, точно такую же половину, — от матери. Дело в том, что существует крошечный фрагмент ДНК, который передается только по материнской линии. Это так называемая митохондриевая ДНК, поскольку она содержит уникальный круглый виток в небольших трубкообразных «пакетах», так называемых митохондриях, которые функционируют как своего рода батарейки в цитоплазме клеток. Некоторые специалисты по молекулярной биологии утверждают, что в древнейшую эпоху развития жизни на Земле митохондрия представляла собой отдельную, независимую особь со своей собственной спиралью ДНК и обладала секретом выработки громадных масс энергии. Впоследствии митохондрии проникли в простейшие одноклеточные организмы и навсегда остались в них, размножаясь, как дрожжи, посредством бинарного деления клеток. Особи мужского пола, хотя они получают и используют митохондриевую ДНК своей матери, не способны передавать ее своему потомству. Мужская сперма имеет свои собственные митохондрии, помогающие ей проделать длительный путь от вагины до яйцеклетки, однако в процессе проникновения в яйцеклетку мужские митохондрии разрушаются и гибнут. Образно говоря, мужчина подобен воину, вынужденному оставить оружие у входа в вечность. Таким образом, каждый из нас наследует митохондриевую ДНК своей матери, которая унаследовала свою митохондриевую ДНК от своей матери, и так далее, на протяжении бесчисленного ряда поколений, вплоть до Евы. Отсюда и популярное название митохондриевой ДНК — «ген Евы». Таким образом, каждый человек, живущий сегодня на Земле, унаследовал свою митохондриевую ДНК от одной-единственной прапрапрабабки, жившей на Земле примерно 200 тысяч лет назад. Эта митохондриевая ДНК представляет собой уникальный момент стабильности среди зыбких песков наследственности ДНК. Однако если все хромосомы Евы, существующие сегодня в мире, были бы абсолютно точной копией генов ДНК Евы, то и все их носители были бы совершенно одинаковыми двойниками. Это было бы поистине чудом и в то же время означало бы, что митохондриевая ДНК не способна передать нам информацию об истории нашей собственной эволюции. Хотя осознание того факта, что родословную всех женщин на свете можно проследить в ретроспективе вплоть до их общего предка — праматери Евы, само по себе выглядит впечатляющим, это не позволяет сколько-нибудь подробно проследить генеалогические линии ее дочерей. Для этого нам необходим более широкий спектр вариантов. Итак, самое время поговорить о мутациях ДНК. Когда мы наследуем от своей матери митохондриевую ДНК (мтДНК), иногда в ней бывает запечатлено случайное изменение или мутация в одном или нескольких «знаках» кода мтДНК — примерно одна мутация на каждую тысячу поколений[47]. Этот новый знак, так называемая точечная мутация, будет передаваться через всех последующих дочерей женщины, у которой она возникла. И хотя новая мутация — явление крайне редкое в пределах генеалогической линии одной семьи, общая вероятность мутаций увеличивается прямо пропорционально числу матерей, имеющих дочерей. Таким образом, в следующем поколении у одного миллиона матерей может быть уже более тысячи дочерей с новой генетической мутацией, причем каждая из них будет в чем-то отличаться от остальных. Вот почему, даже если у всех нас 10 тысяч лет тому назад был общий предок по женской линии, все мы обладаем генетическим кодом, содержащим пусть небольшие, но вполне реальные отличия от окружающих.
Использование мутаций для построения генеалогического древа
За период примерно 200 тысяч лет ряд крошечных случайных мутаций, постоянно аккумулировавшихся в различных молекулах мтДНК человека, был рассеян дочерями Евы практически по всему свету. Это означает, что в родословных каждого из нас по женской линии вплоть до Евы можно найти от семи до пятнадцати таких мутаций. Таким образом, мутации представляют собой нечто вроде сводных досье нашей генеалогии по женской линии за всю историю существования рода человеческого. Основная задача ДНК — передать свою точную копию следующим поколениям. Поэтому мы можем использовать такие мутации для реконструкции генеалогического древа мтДНК, поскольку каждая новая мутация мтДНК в яйцеклетке потенциальной матери будет с неуклонной точностью передана всем ее потомкам по женской линии. Таким образом, каждая женская родословная линия определяется как древними, так и сравнительно новыми генными мутациями. В результате этого, зная все возможные комбинации мутаций у женщин во всем мире, мы можем с достаточной уверенностью реконструировать генеалогическое древо по женской линии вплоть до праматери Евы Хотя набросать на обороте конверта генеалогическое древо и изобразить на нем пару-другую последних мутаций — дело, мягко говоря, нетрудное, эта проблема приобретает куда более сложный характер, если речь идет о составлении генеалогического древа всего рода человеческого со многими тысячами всевозможных комбинаций мутаций. Поэтому сегодня для создания такой реконструкции используются мощные компьютеры. Анализируя код ДНК в генетических пробах современных людей и сопоставляя полученные данные с результатами анализа изменений этого кода за многие и многие поколения, биологи могут проследить, как накапливались эти изменения со времен древнейших предков человека. Поскольку мы наследуем мтДНК только по материнской линии, именно эта линия наследственности и представляет собой объективную картину женской половины генеалогического древа рода человеческого. Сегодня мы можем не только воссоздать это древо, но и, учитывая географические координаты мест, где были взяты те или иные пробы, установить, где именно произошли те или иные мутации: в Европе, Азии или, скажем, Африке. Более того, поскольку такие изменения происходят с хотя и произвольной, но все же поддающейся статистическому анализу частотой, мы можем хотя бы приблизительно определить и время, когда произошли подобные мутации. Таким образом, начиная с конца 1990-х годов для нас, исследователей, стали доступны такие вещи, о которых антропологи недавнего прошлого могли лишь мечтать. Например, мы можем проследить направления и маршруты миграций людей современного типа по всей планете. Оказывается, наиболее древнее из известных изменений мтДНК имело место в Африке примерно 150—190 тысяч лет тому назад. Затем, уже в Азии, около 60—80 тысяч лет тому назад стали происходить все новые и новые мутации (рис. 0.3). Это свидетельствует о том, что люди современного типа впервые появились в Африке, а некоторые племена более 80 тысяч лет назад совершили так называемый исход из Африки. Важно понимать, что в связи с более чем случайной природой индивидуальных мутаций такая датировка носит приблизительный характер. В 1990-е годы было опробовано на практике несколько математических моделей датировки миграций древнейших людей, и эти испытания показали весьма различный уровень достоверности таких моделей. Но лишь один из них, появившийся в 1996 г. и позволяющий датировать каждую из ветвей генетического древа по среднему числу мутаций у женщин данной ветви[48], выдержал испытание временем, и поэтому в этой книге я использую в основном его.
 Рис. 0.3
Рис. 0.3
Y-хромосома: ген Адама
Подобно фрагментам мтДНК, находящимся в ядре наших клеток и передаваемых только по материнской линии, существует набор генов, связанных с ядром клеток, который передается только по мужской линии. Это так называемая Y-хромосома, то есть хромосома-индикатор мужского пола. За исключением небольшого сегмента, Y-хромосома не играет никакой роли в произвольном обмене ДНК с другими хромосомами. Это означает, что, подобно мтДНК, нерекомбинированная часть Y-хромосомы (HPчY) остается неизменяемой из поколения в поколение и по ней можно проследить эволюцию человека вплоть до нашего древнейшего предка по мужской линии. Y-хромосома стала использоваться для реконструкции генеалогического древа значительно позже, чем мтДНК, ибо она создает ряд серьезных трудностей при датировке и определении возраста. Однако если эти проблемы будут решены, методика HPчY может оказаться куда более продуктивной с точки зрения определения времени и места мутаций, чем мтДНК, причем это касается как недавнего, так и весьма отдаленного прошлого. Это объясняется тем, что HPчY гораздо крупнее, чем мтДНК, и, следовательно, обладает большим потенциалом для изменений на генетическом уровне. И тем не менее Y-хромосомы уже оказывают существенную помощь в составлении генетической линии, параллельной линии мтДНК. В основных географических пунктах Y-хромосомы служат подтверждением информации мтДНК: они указывают, что общий предок всех людей современного типа жил в Африке, а более поздний предок всех неафриканских народов — в Азии. Кроме того, поскольку поведение мужчин в целом ряде ключевых аспектов отличается от поведения женщин, история, рассказанная генами Адама, вносит в общее повествование немало интересных деталей. Одно из основных различий заключается в том, что число детей у них варьируется в куда более широком диапазоне, чем у женщин. У некоторых мужчин количество детей на порядок больше, чем у большинства остальных. Что касается женщин, то число отпрысков у них значительно стабильнее и имеет меньший разброс. Главным следствием этого является тот факт, что мужские наследственные линии угасают гораздо быстрее, чем родословные женщин, ибо обычно сохраняется лишь генетическая линия мужчины-лидера, мужчины-победителя. Наконец, последний аспект метода генетического отслеживания: очень важно отличать этот новый подход к генетическому отслеживанию истории бытования молекул на древе ДНК, известный под названием филогеографии (буквально — «древесная география») от математического изучения истории всего рода человеческого, которое используется уже на протяжении многих десятилетий и известно под названием классической генетики. Эти две научные дисциплины основаны на одних и тех же принципах менделевского учения о биологии, но имеют совершенно разные цели и используют разные допущения, и различие между ними во многом объясняется ошибочным пониманием или противоречивостью интерпретации. Простейший способ объяснения этих противоречий заключается в том, что филогеография — это дисциплина, изучающая древнейшую историю индивидуальных молекул ДНК, тогда как классическая генетика изучает доисторическую эпоху развития самого человечества. Однако необходимо иметь в виду, что каждая этническая группа является носителем многочисленных и многообразных вариантов конкретной молекулы ДНК, каждый из которых имеет свою собственную историю и происхождение. Хотя два этих подхода к изучению доисторической эпохи эволюции человечества по определению не могут быть идентичными, их объединяет общая цель — стремление проследить пути миграции различных ветвей человечества. И оказывается, что отследить эволюцию отдельных молекул, присутствующих в нашем теле, гораздо легче, чем попытаться проследить направления миграции целых этнических групп.
Названия генетических линий
В этой книге я поочередно рассматриваю материнские и отцовские кланы, наследственные линии генов, родословные, генетические группы и ветви, и даже макрогруппы. Однако в основе всех этих понятий лежат во многом идентичные вещи: члены макрогруппы носителей определенных генных типов часто имеют общего предка (обычно это можно установить путем отслеживания мтДНК или Y-хромосомы). Численность такой группы до некоторой степени носит произвольный характер и зависит от того, где именно расположено основание данной ветви на генетическом древе. Существует фактор, сразу же бросающийся в глаза при изучении генетического древа: это — отсутствие четкой единообразной системы терминологии для описания этих ветвей. В частности, что касается Y-хромосомы, то каждый новый научный труд выдвигает новую систему наименований и обозначений, основанную на всевозможных доморощенных определениях генов-маркеров, сложившихся в различных научно-исследовательских лабораториях. Это весьма затрудняет прямые сопоставления результатов различных исследований и подвергает серьезному испытанию память читателей. Сам же объект изучения, генетическое древо, практически идентичен в разных лабораториях. Не так давно была опубликована сводная система наименований для Y-хромосомы; в этой системе для описания основных ветвей генетического древа используются буквы от А до R[49]. В таблицах и иллюстрациях, представленных в этой книге, я стремился как можно шире пользоваться этими обозначениям. Однако проблема здесь заключается в том, что эти восемнадцать букв и их расположение на генетическом древе довольно сложно удержать в памяти, по крайней мере — для меня. Мне трудно свыкнуться с тем, что это — всего лишь буквы. К счастью, нашей памяти часто помогают такие факторы, как контекст и ассоциации. По этой и только по этой причине я решил обозначить основные ветви этого древа, которые я упоминаю наиболее часто, именами. Некоторые из этих имен носят региональный характер, например, ветвь Китая и Юго-Восточной Азии я обозначил как Хо (по имени знаменитого китайского флотоводца Чжэн Хо и выдающегося деятеля национально-освободительного движения Вьетнама Хо Ши-мина). Названия других ветвей связаны с именами библейских персонажей: например, Адам (родоначальник всех народов, вышедших из Африки) и три ветви его потомков: Каин, Авель и Сиф (см. Приложения 1 и 2). Еще раз подчеркиваю, что в этих именах не следует искать какого-то глубокого смысла: это — всего лишь подсказки. Несколько проще обстоит дело с мтДНК. Многие научные лаборатории еще на достаточно раннем этапе исследований договорились попытаться использовать единую систему обозначений. (Быть может, все дело в том, что в этом процессе участвовало меньше тестостерона![50]) Так, например, существуют две неафриканские генетические линии, происходящие от единой линии выходцев из Африки, — L3. По согласованной системе названия этих неафриканских линий — «N» и «М» . Так вот, одной из них, относящейся к региону Южной Аравии, я дал имя Насрин, а другой, связанной с Индийским субконтинентом, — имя Манью.
ГЛАВА ПЕРВАЯ ПУТЬ ИЗ АФРИКИ
Одним из наиболее эффектных материалов в области популярной генетики явилась публикация в конце 1980-х гг. на обложке журнала «Newsweek» фото с изображением изысканно-соблазнительных чернокожих Адама и Евы, делящих пополам пресловутое яблоко. На прародителей человечества одобрительно поглядывает змей-искуситель.
 Фото 3. Обложка культового номера «Newsweek» от 11 января 1988 г., где помешена статья, излагающая историю «митохондриевой Евы» мира.
Фото 3. Обложка культового номера «Newsweek» от 11 января 1988 г., где помешена статья, излагающая историю «митохондриевой Евы» мира.
Эта обложка обеспечила номеру журнала рекордный тираж. Однако, несмотря на прием из арсенала массмедиа, «Newsweek» действительно опубликовал информацию о важном научном открытии. И знаменитое фото, и текст вступительной статьи сообщали о двух важнейших достижениях научной мысли. Первым из них явилось опубликованное в 1987 г. исследование в области генетики, где говорилось об открытии генов, которые передаются только по материнской линии. Это исследование, авторами которого были Ребекка Канн и ее коллеги, развеяло в пух и прах устаревшие аргументы о месте появления первых людей современного типа. Согласно новейшим свидетельствам, мы, «современная человеческая семья», произошли от одной-единственной генетической линии, сложившейся за последние 200 тысяч лет в Африке, а не в многочисленных изолированных очагах эволюции, разбросанных по всему миру. Эта единственная линия, восходящая к нашему общему предку с неандертальцами, дала начало примерно полудюжине основных материнских кланов (или ветвей), африканские корни которых четко прослеживаются и в наши дни[51]. Второй причиной использования знаменитой библейской аллегории на обложке «Newsweek» явилось то, что данный генетический метод использовал митохондриевую ДНК (мтДНК), передаваемую только по материнской линии. Десять лет спустя небольшая группа генетиков воспользовалась этим вновь открытым методом и установила, что одна-единственная отрасль дюжины с лишним материнских наследственных генетических ветвей древнейших африканских предков послужила основой для возникновения линии, ставшей родоначальницей всего остального населения земного шара[52]. Другими словами, у всех женских генетических ветвей в Африке существовал некий общий предок, или «митохондриевая Ева», а затем, много позже, сложилась «дочерняя» линия «Евы выходцев из Африки», чьи генетические дочери, то есть потомки по женской линии, заселили весь остальной мир. Это открытие стало поистине эпохальным. Титул «Ева», или «Первая леди», который с такой готовностью подхватили массмедиа тех лет, заключал в себе не совсем тот смысл, который вкладывали в него генетики. Благодаря изучению митохондриевой ДНК ученые сумели выявить древнейшую женскую генетическую линию, охватывающую только крохотную частицу нашей генетической наследственности. Митохондриевая Ева отнюдь не была некой конкретной личностью, «Праматерью», как это часто заявляли представители массмедиа. Отслеживание материнского генетического маркера в глубь далекого прошлого, к древнейшему общему прапредку, вовсе не означает, что абсолютно все наши гены в буквальном смысле происходят от одной-единственной женщины. Дело в том, что в ядре каждой клетки нашего тела имеется более 100 тысяч генетических участков (специальный термин для их обозначения — «локусы»), которые необходимо зафиксировать. Любой из этих участков может использоваться в качестве маркерной системы для отслеживания генетической линии вплоть до древнейшего общего предка[53]. Поскольку эти участки перемешиваются в каждом новом поколении, далеко не все ветви их генетического древа ведут к одному и тому же предку. По сути дела, генетическая наследственность современных людей может быть выведена на основе биологического ядра африканцев численностью от 2000 до 10 000 человек[54], живших около 190 тысяч лет тому назад. Хотя может показаться, что сухая реальность параллельного существования «генетического древа» и «генеалогического древа» развеяла романтическое очарование легенды о Еве, она нисколько не уменьшает исключительной роли генетического отслеживания в величественной истории странствий человечества за последние 200 тысяч лет. И возможность поведать практически ту же историю благодаря целому ряду различных генов лишь подтверждает истинность этого повествования и обогащает его. Так, например, мужские генные линии рисуют куда более яркую и увлекательную картину рассеяния по странам и континентам, чем женские генеалогии. К примеру, недавно в Южной Африке было найдено так называемое исчезнувшее колено Израилево[55], идентифицированное генетиками как этническая группа, имевшая еврейских предков, однако — только по материнской линии[56].
Возражения мультирегионалистов и генетиков
Простой и ясный принцип отслеживания маршрутов миграций человеческого по всему земному шару при помощи генетических маркеров не мог не вызвать возражений. Естественно, первыми отвергли его мультирегионалисты[57]. К ним по большей части относились палеоантропологи, которые по-прежнему считали, что каждая из «рас» на нашей планете произошла от своего собственного архаического предка, например, «яванского человека», «пекинского человека», неандертальца и прочих вымерших видов, живших в различных частях земного шара. Одна из ключевых проблем при этом заключается в том, что современные люди, несмотря на все региональные различия между ними, во многих отношениях гораздо ближе друг к другу, чем были их предполагаемые древнейшие предки. Чтобы хоть как-то объяснить эти несоответствия, мультирегионалисты сегодня утверждают, что различные региональные «подвиды человека» в последующие эпохи активно смешивались и обменивались друг с другом генетическим материалом, в результате чего и сформировались сложные современные расы, которые мы видим сегодня (рис. 1.1). Однако генетические данные не позволяют выявить четких свидетельств такого межрегионального смешения. Географическое распределениеветвей и отраслей современных генетических древ митохондриевой ДНК и Y-xpoмосомы групп выходцев из Африки имеют выраженные региональные отличия и особенности. Отстаивая последние бастионы своей гипотезы, мультирегионалисты расширили концепцию «компромисса межрегионального смешения», чтобы включить в нее гипотезу об исходе из Африки, утверждая, что архаические региональные группы населения, такие, как неандертальцы, могли время от времени смешиваться с пришельцами анатомически современного типа, «совершенно такими же, как мы», то есть, другими словами, с кроманьонцами.
 Рис. 1.1
«Анатомически современные люди» — это представители вида Homo sapiens (sensu stricto)[58], обладавшие комплексом особенностей строения скелета, которые отличали и нас, и наших непосредственных предков, живших в Африке около 200 тысяч лет тому назад, от вымершего архаичного вида Homo sapiens (sensu lato)[59], включая и нашего близкого родича — Homo helmei. Главной особенностью «анатомически современного» человека является высокий круглый череп, выступающий подбородок, небольшие лицо и челюсти и слабо выраженные надбровные дуги. Межрегиональное смешение архаических туземцев и пришельцев представляет собой мягкий компромиссный вариант гипотезы исхода из Африки, позволяющий объяснить присутствие некоторых генов архаических видов человека в генетическом «котле» современного человека. В качестве примера подобной «гибридизации» не раз назывался найденный в Португалии детский скелет с короткими неразвитыми конечностями, возраст которого — 24 500 лет, то есть относящийся к эпохе, когда неандертальцы уже давно вымерли. И все же среди генных проб мтДНК и Y-хромосом, сданных десятками тысяч добровольцев, живущих за пределами Африки, не обнаружено даже минимальных проявлений подобного рода смешения[60].
Честно говоря, история на этом не кончается. Хотя свидетельств наличия таких современных архаических «примесей» в генетических линиях «Адама» и «Евы» не существует, эти линии сами по себе составляют крайне незначительную часть обширной «библиотеки» геномов ДНК. Поскольку каждому последующему поколению передается только одна копия мтДНК и Y-хромосомы, линии мтДНК неандертальцев лишь в крайне редких случаях пересекались и смешивались с кроманьонским типом, а затем окончательно вымерли. Однако среди остатков огромного нуклеарного генома до сих пор прослеживаются следы генного смешения. А поскольку такая нуклеарная ДНК смешивается и расщепляется в каждом поколении, крайне сложно построить генетическое древо для каждой из бесчисленных генных линий и с уверенностью сказать, что на нем принадлежит людям современного анатомического типа, а что является наследием регионального архаического населения[61]. Таким образом, как утверждают некоторые ученые, не исключено, что густые кустистые брови коренастых игроков в регби и футбольных хулиганов представляют собой генетическую прапамять о неандертальцах, а не (что, на мой взгляд, более вероятно) нормальные вариации внешности анатомически современного человека.
Совсем другие возражения выдвигают представители традиционной генетики. Они были удивлены и даже, по всей видимости, обескуражены тем, что митохондриевая ДНК дала столь четкую генетическую картину даже при применении их собственных методов с использованием традиционных генных маркеров ядер (передаваемых как мужчинами, так и женщинами), демонстрируя с годами все менее четко выраженную оппозицию между Африкой и остальным миром. Эта нечеткость объясняется целым рядом факторов. Наиболее важным из них можно считать излюбленный традиционалистами классический подход, заключающийся в реконструкции древа массовых миграций, а не в построении древа изменений самих генных линий. Показательно, что подобные идеи сохранились и в более широкой и математически обоснованной концепции «расовых миграций», а не в отслеживании миграций индивидуальных линий самих генов, выявленных у данной группы населения. Другим важным фактором явилось смешение и перенос генных линий в каждом последующем поколении, чего никогда не происходит с мтДНК. Таким образом, большинство генных маркеров, использовавшихся традиционалистами, имели весьма несложные деревья с небольшим число очень древних ветвей, тогда как отростки на них были общими для многих рас населения.
По всем вышеизложенным причинам традиционный способ сравнения наследственных признаков у жителей разных регионов путем замеров частоты присутствия тех или иных генов в ядре имел коренной недостаток применительно к изучению древнейших миграций. Он со всей наглядностью продемонстрирован в исследовании присутствия генов внутри ядра на основе всего одного генного локуса (т.е. речь в данном случае не шла о генах Адама и Евы), осуществленном Джеймсом Уэйнскотом и его коллегами из Оксфордского университета, сторонниками модели африканского происхождения. Это исследование было опубликовано журнале «Nature» в 1986 г. Таким образом, оно вышло в свет всего за год до появления открытий Ребекки Канн, о которых сообщалось в статье в «Newsweek». В той памятной статье были использованы данные генетических проб, взятых у людей со всех концов света, в том числе и проб, взятых мною в 1984 г. в Новой Гвинее. Вместо сравнения относительной частоты встречаемости многих генов Уэйнскот выстроил генеалогическое древо на основе одного-единственного генного локуса. Результат был в принципе тем же самым, что и выводы Канн, свидетельствуя об африканском происхождении. Другие генетики также выдвинули целый ряд технически аргументированных возражений против анализа генов Евы. С тех пор эти возражения технического характера снимаются один за другим[62].
В результате недавнего анализа другой несмешанной системы, Y-хромосомы, и исследований многих других генных маркеров[63] первоначальная версия о происхождении людей современного типа из Африки, подтверждаемая маркерами митохондриевой ДНК, одержала триумфальную победу, и мультирегионалисты превратились в изолированное, но задиристое и горластое меньшинство.
Рис. 1.1
«Анатомически современные люди» — это представители вида Homo sapiens (sensu stricto)[58], обладавшие комплексом особенностей строения скелета, которые отличали и нас, и наших непосредственных предков, живших в Африке около 200 тысяч лет тому назад, от вымершего архаичного вида Homo sapiens (sensu lato)[59], включая и нашего близкого родича — Homo helmei. Главной особенностью «анатомически современного» человека является высокий круглый череп, выступающий подбородок, небольшие лицо и челюсти и слабо выраженные надбровные дуги. Межрегиональное смешение архаических туземцев и пришельцев представляет собой мягкий компромиссный вариант гипотезы исхода из Африки, позволяющий объяснить присутствие некоторых генов архаических видов человека в генетическом «котле» современного человека. В качестве примера подобной «гибридизации» не раз назывался найденный в Португалии детский скелет с короткими неразвитыми конечностями, возраст которого — 24 500 лет, то есть относящийся к эпохе, когда неандертальцы уже давно вымерли. И все же среди генных проб мтДНК и Y-хромосом, сданных десятками тысяч добровольцев, живущих за пределами Африки, не обнаружено даже минимальных проявлений подобного рода смешения[60].
Честно говоря, история на этом не кончается. Хотя свидетельств наличия таких современных архаических «примесей» в генетических линиях «Адама» и «Евы» не существует, эти линии сами по себе составляют крайне незначительную часть обширной «библиотеки» геномов ДНК. Поскольку каждому последующему поколению передается только одна копия мтДНК и Y-хромосомы, линии мтДНК неандертальцев лишь в крайне редких случаях пересекались и смешивались с кроманьонским типом, а затем окончательно вымерли. Однако среди остатков огромного нуклеарного генома до сих пор прослеживаются следы генного смешения. А поскольку такая нуклеарная ДНК смешивается и расщепляется в каждом поколении, крайне сложно построить генетическое древо для каждой из бесчисленных генных линий и с уверенностью сказать, что на нем принадлежит людям современного анатомического типа, а что является наследием регионального архаического населения[61]. Таким образом, как утверждают некоторые ученые, не исключено, что густые кустистые брови коренастых игроков в регби и футбольных хулиганов представляют собой генетическую прапамять о неандертальцах, а не (что, на мой взгляд, более вероятно) нормальные вариации внешности анатомически современного человека.
Совсем другие возражения выдвигают представители традиционной генетики. Они были удивлены и даже, по всей видимости, обескуражены тем, что митохондриевая ДНК дала столь четкую генетическую картину даже при применении их собственных методов с использованием традиционных генных маркеров ядер (передаваемых как мужчинами, так и женщинами), демонстрируя с годами все менее четко выраженную оппозицию между Африкой и остальным миром. Эта нечеткость объясняется целым рядом факторов. Наиболее важным из них можно считать излюбленный традиционалистами классический подход, заключающийся в реконструкции древа массовых миграций, а не в построении древа изменений самих генных линий. Показательно, что подобные идеи сохранились и в более широкой и математически обоснованной концепции «расовых миграций», а не в отслеживании миграций индивидуальных линий самих генов, выявленных у данной группы населения. Другим важным фактором явилось смешение и перенос генных линий в каждом последующем поколении, чего никогда не происходит с мтДНК. Таким образом, большинство генных маркеров, использовавшихся традиционалистами, имели весьма несложные деревья с небольшим число очень древних ветвей, тогда как отростки на них были общими для многих рас населения.
По всем вышеизложенным причинам традиционный способ сравнения наследственных признаков у жителей разных регионов путем замеров частоты присутствия тех или иных генов в ядре имел коренной недостаток применительно к изучению древнейших миграций. Он со всей наглядностью продемонстрирован в исследовании присутствия генов внутри ядра на основе всего одного генного локуса (т.е. речь в данном случае не шла о генах Адама и Евы), осуществленном Джеймсом Уэйнскотом и его коллегами из Оксфордского университета, сторонниками модели африканского происхождения. Это исследование было опубликовано журнале «Nature» в 1986 г. Таким образом, оно вышло в свет всего за год до появления открытий Ребекки Канн, о которых сообщалось в статье в «Newsweek». В той памятной статье были использованы данные генетических проб, взятых у людей со всех концов света, в том числе и проб, взятых мною в 1984 г. в Новой Гвинее. Вместо сравнения относительной частоты встречаемости многих генов Уэйнскот выстроил генеалогическое древо на основе одного-единственного генного локуса. Результат был в принципе тем же самым, что и выводы Канн, свидетельствуя об африканском происхождении. Другие генетики также выдвинули целый ряд технически аргументированных возражений против анализа генов Евы. С тех пор эти возражения технического характера снимаются один за другим[62].
В результате недавнего анализа другой несмешанной системы, Y-хромосомы, и исследований многих других генных маркеров[63] первоначальная версия о происхождении людей современного типа из Африки, подтверждаемая маркерами митохондриевой ДНК, одержала триумфальную победу, и мультирегионалисты превратились в изолированное, но задиристое и горластое меньшинство.
Два маршрута исхода из Африки?
Несомненно, предки современных людей вышли из Африки, как и их ближайшие родичи — приматы, однако конкретное время и маршруты такого исхода, как и всегда, определялись колебаниями климата. В древности существовало два потенциальных маршрута исхода из Африки — северный и южный, и каким именно воспользоваться в ту или иную эпоху, зависело от погодных условий. Когда оказывался открытым какой-то один из них, он указывал мигрантам дальнейший путь — на север или на восток. Люди современного типа впервые совершили исход из Африки около 120 тысяч лет тому назад, пройдя через северные врата континента. Как мы увидим ниже, этот первый исход завершился трагически. Второй же, успешный исход, открыл им путь через Азию, на юг и восток — путь, проторенный их предшественниками. Что касается Европы, то ее они игнорировали примерно до 50 тысяч лет тому назад. Африка стала родиной всего многообразия видов человека, населяющих нашу планету. Необъятная и изолированная природная лаборатория вновь и вновь переплавляла человеческий материал, пропуская его через бесчисленные циклы появления пустынь и появления зелени. Уникальное по масштабам лоскутное одеяло саванн и лесов, которое представляла собой Африка на пороге Сахары, было надежно изолировано от остального мира двумя комплексами естественных ворот и коридоров. На протяжении двух последних миллионов лет эти коридоры действовали как громадные загоны для переселенцев и их скота, загоны, ворота которых попеременно открывались и закрывались. Когда один комплекс ворот открывался, другой обычно закрывался. Одни ворота вели на север, через Сахару и Левант, в Европу, тогда как другие открывали путь на восток, через устье Красного моря, в Йемен, Оман и Индию. Какие именно из ворот оказывались открытыми в данный конкретный период, зависело от очередного цикла оледенения, который, собственно, и определял, какие виды млекопитающих, включая человека, могут мигрировать из Африки, направляясь на север, в Европу, или на восток, в Азию[64]. В наши дни дни Африка соединена с Евразийским макроконтинентом всего лишь одним из прежних коридоров — знаменитым Синайским полуостровом на северовостоке. Эти суровые места, представлявшие собой засушливую пустыню, служили потенциальным маршрутом через Сахару и Синайскую пустыню — дорогой, которая вела во внешний мир и, словно звездные врата из какого-нибудь научно-фантастического фильма, открывались людям лишь тогда, когда очередное изменение угла орбиты Земли или наклона полярной оси вызывало кратковременный период потепления. Именно такой редкий эпизод в геологической истории Земли и имел место около 100 тысяч лет тому назад, когда усиление солнечной активности повлекло за собой активное таяние ледников, и на нашей планете установился теплый и достаточно влажный климат. Пустыни Сахара и Синай, а также пустыни далекой Австралии превратились в огромные озера, окруженные буйной растительностью и достигшие расцвета во время своей короткой геологической весны (рис. 1.2). Но поскольку эта интерлюдия тепла оказалась очень краткой, климатические врата Северной Африки, впустив мигрантов, оказались для них смертельной ловушкой. На протяжении большей части последних 2 млн. лет люди вдоволь натерпелись голода и холода в суровом плену плейстоценовой ледниковой эпохи, и поэтому непродолжительное, но заметное потепление климата на нашей планете, открывшее им врата Эдема, известно у геологов под названием междуледникового оптимума. Эти краткие, но пьяняще прекрасные паузы резко контрастировали с привычным холодным и сухим климатом ледниковых просторов плейстоцена. Нам, современным людям, довелось пережить лишь два таких райских периода за все время нашего существования на Земле. Последний из таких междуледниковых оптимумов имел место около 8000 лет тому назад, и мы до сих пор наслаждаемся последними лучами его осеннего тепла. На протяжении примерно 2000 лет Сахара представляла собой степь, покрытую густыми травами, и бесчисленное множество всевозможных видов животных мигрировало по ее просторам с юга на север, через Северную Африку на Левант.
 Рис. 1.2
Рис. 1.2
По иронии судьбы в наши дни глобальное потепление, вызванное всеобщим загрязнением окружающей среды, помогает затормозить неизбежный возврат к такому же более засушливому и холодному климату, который господствовал на Земле на протяжении большей части нашего существования на ней. Как правило, свидетельства таких древних климатических драм в доисторические эпохи ученые получают в результате раскопок глубоко залегающих слоев или бурения скважин в полярных ледяных шапках и на морском дне. Однако в данном случае нам не требуются обстоятельные научные доказательства из области археологии и климатологии, чтобы доказать истинность нашей теории. Мы можем увидеть все это собственными глазами, побывав в Сахаре и полюбовавшись удивительными наскальными рисунками, уцелевшими в самом ее сердце. Там более 8000 лет назад безвестными древними художниками были созданы многие и многие тысячи натуралистических изображений вымерших видов буйволов, слонов, носорогов, бегемотов, жирафов , страусов и больших антилоп.
 Фото 4. Найденные в Сахаре наскальные рисунки слонов, бегемотов и жирафов, возраст которых — ок. 8 тыс. лет, аналогичные этим из Феззана, Ливия, реальное напоминание о временах, когда в этих местах простирались травянистые саванны.
Фото 4. Найденные в Сахаре наскальные рисунки слонов, бегемотов и жирафов, возраст которых — ок. 8 тыс. лет, аналогичные этим из Феззана, Ливия, реальное напоминание о временах, когда в этих местах простирались травянистые саванны.
Более того, эта удивительная историческая хроника велась непрерывно на протяжении нескольких тысячелетий. Наиболее поздние рисунки со всей очевидностью свидетельствуют, что обилие видов животных в Сахаре практически исчезло около 5000 лет назад, уступив место верблюдам и тарантулам. Это время, когда животные из Центральной Африки потоком устремились на земли нынешнего Марокко, Египта и дальше, на Левант, представляло собой нечто вроде редчайшего дождя в пустыне, под живительными струями которого просыпаются и оживают семена растений, долгие десятилетия дремавшие в земле. Увы, этот земной рай просуществовал очень недолго; животные покинули эти края, и в них вновь воцарилась безжизненная пустыня. Крупные млекопитающие, в отличие от растений, не имели семян, которые можно было бы надолго укрыть в песке до наступления лучших времен. Не только млекопитающие, но и другие животные, лучше адаптирующиеся к неблагоприятному климату, например пустынный узкорот или двоякодышащие рыбы, могут на непродолжительное время погружаться в некий анабиоз в ожидании следующего дождя. Таким образом, во время предыдущей междуледниковой паузы, первой за время существования человека на Земле, отважные первопроходцы, устремившиеся на север, успели покинуть пределы Африки и попасть на Левант до того, как за их спиной вновь затворились врата Сахары. Более ранняя междуледниковая пауза, известная среди ученых как Эмианская, или Ипсвичская, пауза, наступила около 125 тысяч лет тому назад, вскоре после появления на Земле людей современного типа. Благодаря находкам человеческих останков мы знаем, что древнейшие люди современного типа мигрировали из окрестностей Сахары в Северную Африку и на Левант в очень и очень отдаленные времена. Действительно, наиболее древние останки человека современного типа, найденные за пределами Африки, возраст которых составляет от 90 до 120 тысяч лет, были найдены именно в странах Леванта[65]. Возникает принципиальный вопрос: не оставили ли они там сколько-нибудь заметный след? Судя по всему — нет.
Из Африки в Европу: неудавшийся первый исход
Пока первые научные данные не получили убедительного подтверждения, ученые — сторонники гипотезы исхода из Африки — полагали, что древнейший исход людей современного типа на север Африки и далее, на Левант, сформировал своего рода биологическое ядро, из которого впоследствии возникли народы Европы и Азии. Однако подобные аргументы страдали серьезным недостатком. Дело в том, что следы человека современного типа в этих местах около 90 тысяч лет тому назад практически исчезают. Благодаря климатологическим исследованиям мы знаем, что именно около 90 тысяч лет тому назад на Земле наступил краткий, но разрушительный по своим последствиям период резкого глобального похолодания и засухи, в результате чего весь Левант превратился в безжизненную пустыню. После отступления ледников и нового потепления Левант был быстро заселен, но на этот раз — представителями другого вида, нашего ближайшего «кузена» на генеалогическом древе — неандертальцами, которые, по всей вероятности, оказались оттесненными к югу, в регион Средиземноморья, в результате наступления ледников, надвигавшихся с севера. Мы не располагаем материальными свидетельствами пребывания людей современного типа на Леванте или в Европе на протяжении последующих 45 тысяч лет, до тех пор пока примерно 45—50 тысяч лет тому назад на арене истории не появились кроманьонцы (о чем говорит появление авгурисианской техники изготовления орудий), бросившие вызов неандертальцам, оттеснив их на север, на их древнюю прародину[66]. Таким образом, большинство специалистов сегодня полагают, что первые люди современного типа, выходцы из Африки, вымерли на Леванте в результате резкого похолодания и возврата засушливого климата, под влиянием которого Северная Африка и Левант быстро превратились в бесплодные пустыни. Коридор, пролегавший через Сахару, захлопнулся, словно гигантская ловушка, и мигранты, оказавшиеся в ней, не смогли ни возвратиться назад, ни найти пригодных для жизни земель. Зияющая пропасть в 50 тысяч лет между исчезновением следов первых переселенцев на Леванте и последующим вторжением туда новой волны переселенцев из Европы, вне всякого сомнения, ставит под вопрос обоснованность широко распространенной версии о том, что первый исход из Африки на север якобы завершился успешно и создал биологическое ядро будущих европейцев[67]. Давайте же задумаемся — почему. Чтобы понять, почему многие европейские авторитеты в области археологии и антропологии настаивают, будто европейцы возникли самостоятельно и независимо от первого исхода из Северной Африки, необходимо помнить о том, что здесь мы имеем дело с одним из проявлений культурологического европоцентризма, стремящегося объяснить последствия первого исхода. Наиболее важным проявлением такого мышления является несокрушимая убежденность европейских ученых XX в. в том, что именно кроманьонцы, мигрировавшие в Европу не позднее 50 тысяч лет тому назад, и явились основоположниками людей «современного типа» в полном смысле слова. Эта человеческая эпифания[68], принесшая невиданный расцвет всевозможных искусств, ремесел и технических возможностей и культуры в целом, известна среди археологов под сухим названием «европейский Верхний палеолит». По мнению многих ученых, это было нечто вроде творческого взрыва, знаменовавшего начало эпохи мыслящего человека на Земле. Именно к этой культуре восходят впечатляющие наскальные рисунки в пещерах Шове и Ласко, а также изысканные, тонко проработанные резные фигурки «Венеры», которые археологи находят по всей Европе. При этом обычно можно слышать аргументы типа «если мы действительно вышли из Африки и если та древняя культурная революция, столь красноречиво говорящая о даре абстрактного мышления, пришла в Европу с Леванта, она в лучшем случае могла представлять собой краткий привал на пути из Египта». Ergo[69], «мы, люди Запада» (это «мы» объясняется тем, что сторонники этой гипотезы — исключительно европейцы или имеют европейские корни), — всего лишь потомки выходцев из Северной Африки. Таким образом, северный маршрут для многих специалистов является этакой концептуальной отправной точкой миграции или, лучше сказать, исхода из Африки. В следующей главе мы рассмотрим, почему с точки зрения логики невозможно допустить, что первыми «людьми полностью современного типа» были европейцы, и как случилось, что первыми современными людьми, способными говорить, петь, танцевать и рисовать, стали именно африканцы, причем произошло это задолго до исхода некоторых их групп со своего родного континента. Однако попытки дать убедительное объяснение того, как конкретно предки современных европейцев, жившие некогда в окрестностях Сахары, задумали и осуществили исход из Африки, связаны с целым рядом серьезных проблем. Для начала надо заметить, что, поскольку пустыня Сахара на протяжении последних 100 тысяч лет служила непреодолимой преградой для мигрантов, любые позднейшие вторжения выходцев из Северной Африки в Европу могли начинаться с какого-нибудь зеленого прибежища — островка растительности, еще остававшегося в Северной Африке, например из района дельты Нила, после междуледниковой паузы. Предки европейцев не могли 45—50 тысяч лет тому назад совершить исход из региона Сахары напрямую, кроме как на плотах вниз по течению Нила, однако генетическая история решительно отвергает подобную возможность.
Зеленое прибежище в Египте?
Если на всем протяжении длительного засушливого периода после междуледниковой паузы в Северной Африке действительно существовало подобное зеленое прибежище, оно около 45 тысяч лет тому назад вполне могло послужить временным приютом и перевалочным пунктом для предков будущих европейцев. Да, в древности в Северной Африке действительно существовало несколько обширных зеленых оазисов, в частности, дельта Нила в Египте и Средиземноморское побережье нынешнего Марокко. Недавняя находка детского скелета в погребении на холме Тарамса в Египте, датируемая ориентировочно между 50 и 80 тысячами лет тому назад, свидетельствует о том, что там могли сохраниться реликтовые группы населения. Ряд ведущих сторонников гипотезы исхода из Африки сразу же обратили внимание на эту находку, поскольку она предлагает реальное и вполне убедительное объяснение паузы протяженностью в 45—50 тысяч лет. Наибольшей известностью среди них пользуется Крис Стрингер, убежденный приверженец гипотезы о происхождении современных людей из Африки и один из руководителей Лондонского музея естественной истории. Стрингер утверждает, что ребенок-египтянин из Тарамсы принадлежал к колонии обитателей оазисов Северной Африки и что выходцами именно из таких колоний и были мигранты, покинувшие около 50 тысяч лет тому назад Африку и ставшие предками жителей Леванта и Европы[70]. И все же археологические свидетельства присутствия кроманьонцев в Северной Африке крайне скудны и немногочисленны. Даже те каменные орудия эпохи Среднего палеолита, которые были найдены в захоронении ребенка на холме Тарамса, вполне могли быть созданы неандертальцами, и их никак нельзя считать доказательством взрывного роста новых технологий, проникших в ту эпоху в Европу.
Проблема Австралии
Но, пожалуй, наиболее серьезной проблемой для европоцентрической концепции культурного развития, в основе которой лежит гипотеза о северном маршруте исхода из Африки, является сам факт существования австралийских аборигенов, создавших свою собственную культуру пения, танца и живописи задолго до европейцев и, естественно, без какой бы то ни было помощи с их стороны. Но тогда выходцами из какого района Африки они были? Какой маршрут завел их в такую даль, на край света? Можно ли считать их ветвью того же исхода, в котором принимали участие и предки современных европейцев? И, наконец, самое главное: каким образом и почему они попали в Австралию гораздо раньше, чем предки европейцев — в Европу? Эта загадка породила целый ряд попыток объяснения. Понятно, что ответить на все эти вопросы, исходя из гипотезы об одном-единственном северном исходе из Африки в Европу, имевшем место примерно 45 тысяч лет тому назад, за которым последовало расселение человека по всему остальному миру, как то утверждает чикагский антрополог Ричард Клейн в своем классическом труде «Развитие человека», попросту невозможно. Известный зоолог, знаток Африки, художник и писатель Джонатан Кингдон идет еще дальше, доказывая, что первый, «неудачный» северный исход африканцев на Левант, имевший место около 120 тысяч лет назад, привел к расселению уцелевших мигрантов и колонизации Юго-Восточной Азии, а затем и Австралии примерно 90 тысяч лет назад. Эта версия также допускает всего один исход из Африки, и притом по северному маршруту. Крис Стрингер избрал наиболее легкий путь, утверждая, что Австралия была колонизована независимо от этого исхода и задолго до освоения Европы в результате отдельного исхода африканцев в обход Красного моря (см. рис. 1.3)[71]. Во многом соглашаясь с Крисом Стрингером, археолог Роберт Фоули и палеонтолог Марта Лар из Кембриджского университета тоже утверждают, что цепь зеленых оазисов в Северной Африке, простиравшаяся на всем протяжении северного маршрута через Левант, имела жизненно важное значение для предков европейцев и жителей стран Леванта. Эти исследователи не испытывают никаких проблем с количеством исходов из Африки, утверждая, что в глубокой древности имело место множество больших и малых миграций, отправными точками для которых служили оазисы, разбросанные по Эфиопии и всей Северной Африке. Эта точка зрения учитывает значительный рост численности населения в самой Африке во время междуледникой паузы, около 125 тысяч лет тому назад. Лар и Фоули полагают, что возвращение прежнего холодного и засушливого климата привело к тому, что африканский континент как бы разделился на отдельные обитаемые районы-колонии, совпадающие с границами зеленых оазисов (см. рис. 1.6), обитатели которых на протяжении последующих 50 тысяч лет были разделены непреодолимыми пустынями. Согласно схеме Лар-Фоули, предки аборигенов Восточной Азии и Австралии могли быть выходцами из Эфиопии, которые, переправившись через Красное море, отправились в дальние странствия. Они могли выбрать южный маршрут и двинуться по нему в путь совершенно независимо от предков будущих европейцев. Не так давно Фоули и Лар получили «подкрепление»: ряды сторонников северного и южного исходов пополнил американский генетик Питер Андерхилл, специалист в области изучения Y-хромосомы. Он выступил с исследованием, в котором осуществил синтез генетических доисторических факторов. Все трое ученых постулировали древний исход в Австралию по южному маршруту, признавая, что основным маршрутом исхода из Африки был все же северный путь, через Суэц и Левант, в Европу и остальные регионы Азии (рис. 1.3) и что он имел место между 30 и 45 тысячами лет тому назад[72]. Таким образом, обоснованность высказываемого многими специалистами по Евразии мнения о том, что предки европейцев были выходцами из Северной Африки, зависит от целого ряда факторов. К их числу относятся наличие достаточно обширных прибежищ-оазисов в Северной Африке и либо многочисленные разновременные миграции из Африки, либо очень ранняя протомиграция с Леванта в страны Дальнего Востока. Существует и проблема идеологического плана: это — попытка зарезервировать северный маршрут исхода только за предками будущих европейцев.
 Рис. 1.3
Высказываясь поначалу откровенно и прямолинейно, Джонатан Кингдон утверждал, что ранний северный исход из Африки произошел около 120 тысяч лет тому назад, во время так называемой Эмианской междуледниковой паузы[73]. Поскольку многие коридоры в пустынях Африки и Западной Азии в ту эпоху пышно зеленели буйной растительностью, предполагаемые мигранты в Австралию могли без помех продвигаться все дальше на восток с Леванта до Индии. Разумеется, они могли остановиться на длительный привал в зеленых районах Южной Азии, прежде чем двинуться дальше, в Юго-Восточную Азию, куда они прибыли около 90 тысяч лет тому назад. (Под термином «Южная Азия» я имею в виду те страны, расположенные между Аденом (Йеменом) и Бангладеш, которые выходят на береговую линию Индийского океана. К числу этих стран относятся Йемен, Оман, Пакистан, Индия, Шри-Ланка и Бангладеш, а также государства, расположенные на побережье Персидского залива: Саудовская Аравия, Ирак, Бейрут[74], Объединенные Арабские Эмираты и Иран.)
В качестве доказательства присутствия еще в глубокой древности разумного человека на Леванте Джонатан Кингдон ссылается на многочисленные орудия эпохи Среднего палеолита, найденные в Индии. Возраст некоторых из них — 1бЗ тысячи лет. Однако наиболее серьезной проблемой здесь является полное отсутствие скелетных останков человека современного типа такой древности где бы то ни было за пределами Африки. Кингдон утверждает, что эти орудия могли быть изготовлены людьми предсовременного или архаического типа (или мапа, как он их называет), которые как раз в то самое время обитали в Восточной Азии.
Понятно, что для того, чтобы попасть в Австралию, предки австралийцев должны были пересечь с запада на восток всю Азию, однако у нас нет никаких материальных доказательств того, что анатомические современные люди совершали миграции через всю Азию около 90 тысяч лет назад, не говоря уж о более ранней эпохе — 120—163 тысячи лет назад.
Рис. 1.3
Высказываясь поначалу откровенно и прямолинейно, Джонатан Кингдон утверждал, что ранний северный исход из Африки произошел около 120 тысяч лет тому назад, во время так называемой Эмианской междуледниковой паузы[73]. Поскольку многие коридоры в пустынях Африки и Западной Азии в ту эпоху пышно зеленели буйной растительностью, предполагаемые мигранты в Австралию могли без помех продвигаться все дальше на восток с Леванта до Индии. Разумеется, они могли остановиться на длительный привал в зеленых районах Южной Азии, прежде чем двинуться дальше, в Юго-Восточную Азию, куда они прибыли около 90 тысяч лет тому назад. (Под термином «Южная Азия» я имею в виду те страны, расположенные между Аденом (Йеменом) и Бангладеш, которые выходят на береговую линию Индийского океана. К числу этих стран относятся Йемен, Оман, Пакистан, Индия, Шри-Ланка и Бангладеш, а также государства, расположенные на побережье Персидского залива: Саудовская Аравия, Ирак, Бейрут[74], Объединенные Арабские Эмираты и Иран.)
В качестве доказательства присутствия еще в глубокой древности разумного человека на Леванте Джонатан Кингдон ссылается на многочисленные орудия эпохи Среднего палеолита, найденные в Индии. Возраст некоторых из них — 1бЗ тысячи лет. Однако наиболее серьезной проблемой здесь является полное отсутствие скелетных останков человека современного типа такой древности где бы то ни было за пределами Африки. Кингдон утверждает, что эти орудия могли быть изготовлены людьми предсовременного или архаического типа (или мапа, как он их называет), которые как раз в то самое время обитали в Восточной Азии.
Понятно, что для того, чтобы попасть в Австралию, предки австралийцев должны были пересечь с запада на восток всю Азию, однако у нас нет никаких материальных доказательств того, что анатомические современные люди совершали миграции через всю Азию около 90 тысяч лет назад, не говоря уж о более ранней эпохе — 120—163 тысячи лет назад.
Преграды на востоке
С предложенной Кингдоном датировкой временных рамок колонизации Юго-Восточной Азии — 90—120 тысяч лет — связана и другая серьезная проблема. Если, согласно его гипотезе, первая волна миграции в Юго-Восточную Азию покинула земли Леванта несколько позже 115 тысяч лет тому назад, она, по всей вероятности, бесследно сгинула на необозримых просторах Азии. Анализ массовых миграций человека и других видов млекопитающих из Африки в Азию за последние 4 млн. лет свидетельствует о том, что, за исключением первой междуледниковой паузы, мигрантов, продвигавшихся с Леванта во внутренние районы Азии, ожидало множество труднопреодолимых препятствий. В эпохи, когда мир не был согрет благодатным теплом междуледниковой паузы, переселенцам то и дело встречались высокие горы и иссушенные зноем пустыни, служившие непреодолимыми преградами на пути на север, восток и юг от Леванта. На севере и востоке протянулась огромная горная цепь Загрос-Таурус, которая вместе с Сирийской и Аравийской пустынями изолировала Левант от Восточной Европы на севере и индийского субконтинента на юге[75]. При обычных климатических условиях оледенения это были непроходимые гористые пустыни. Не было удобного обходного пути и на севере, где высились хребты Кавказа и шумело Каспийское море. В древности, как и во времена Марко Поло, самым удобным альтернативным путем из Восточного Средиземноморья в Юго-Восточную Азию было как можно скорее добраться до Индийского океана и далее двигаться вдоль его береговой линии. Однако к югу и востоку от Леванта простирались Сирийская и Аравийская пустыни, и единственно возможный маршрут вел из Турции через долину Тигра и далее на юг, вдоль западного склона горной гряды Загрос, до самого побережья Персидского залива (см. рис. 1.6). Однако этот маршрут, пролегавший через так называемый Плодородный Полумесяц, в периоды похолодания и засухи по окончании междуледниковых пауз также лежал через безжизненные пустыни и, естественно, был закрыт для древних мигрантов. Практическая невозможность для людей современного типа попасть из Леванта в Египет или Юго-Восточную Азию в период от 55 до 90 тысяч лет тому назад означает, что северный маршрут исхода из Африки в те времена позволял покинуть Черный континент только предкам будущих европейцев и жителей Леванта, а никак не праотцам обитателей Юго-Восточной Азии или Австралии. Между тем, как это ни странно, Европа и Левант не подвергались сколько-нибудь активной колонизации примерно до рубежа 45—50 тысяч лет тому назад, тогда как Австралия, лежавшая на другом конце света, напротив, интенсивно заселялась задолго до этой рубежной эпохи. А это означает, что для того, чтобы «зарезервировать» северный маршрут исхода только за предками европейцев, Крис Стрингер, Боб Фоули и Марта Лар должны были принять гипотезу о существовании в древности отдельных южных маршрутов, которыми воспользовались предки австралийцев и даже жителей Азии. Решить эту загадку позволяет только изучение генетической истории.
Что же говорят гены о северном маршруте исхода
Все подобные гипотезы и предположения в прошлом базировались на археологических свидетельствах, состоявших из нескольких человеческих костей, датировка которых вызывала серьезные сомнения, и к тому же разделенных огромной паузой во времени. На рубеже тысячелетия вышел в свет обширный труд таких видных ученых, как Мартин Ричардс, Винсент Маколей и Ханс-Юрген Бандельт, посвященный генетическому отслеживанию корней современных европейцев. Этот труд полностью изменил прежние представления и позволил нам проанализировать маршрут и последствия первого исхода из Африки с гораздо более высокой точностью в пространственных и временных координатах[76]. Данное исследование решает одновременно две задачи. Во-первых, оно подтверждает, что наиболее ранняя «экспедиция» на Левант, имевшая место предположительно около 100 тысяч лет тому назад, канула в вечность, не оставив никаких следов. Точно так же все участники и потомки первого крупного исхода людей современного типа вымерли, как и неандертальцы, около 60 тысяч лет назад, не оставив никаких генетических следов на Леванте. Во-вторых, хотя африканцы, жившие в окрестностях Сахары, оставили свои генные маркеры в генах примерно 1/8 части жителей современных общин берберов, пока что не обнаружено никаких генетических свидетельств того, что предки европейцев и жителей Леванта были выходцами из Северной Африки. Откуда мне это известно? Дело в том, что реконструкция достаточно точного генетического древа при помощи митохондриевой ДНК позволяет достичь куда большего, чем просто идентифицировать наших общих древнейших предков. На рис. 1.4 представлено митохондриевое древо, основанием ствола которого послужили многочисленные африканские кланы. Одна из ветвей этого древа (L3) протянулась очень и очень далеко, в Южную Азию (Индию). Именно от ветви L3 произошла наша азиатская Ева выходцев из Африки, многочисленные ветви-потомки которой заселили Аравию и Индию, а затем Европу и Ближний Восток. Мы можем датировать ветви на этом древе, а затем, проанализировав их географическое распределение, показать, где и когда появлялись на свет родоначальники тех или иных доисторических миграций. По большому счету, этот метод можно считать убедительным доказательством правоты гипотезы о исходе из Африки. Благодаря этому методу мы можем проследить, что древнейшая ветвь предков европейцев, обозначенная на схеме U (ее возраст — около 50 тысяч лет; см. рис. 1.4), которой я дал имя Европа — в честь знаменитой возлюбленной Зевса, возникла и сформировалась где-то неподалеку от Индии, отделившись от ветви R, которую я назвал Рохани — по ее локусу в Индии. В свою очередь, R отделилась от ветви N, которой я дал имя Насрин и которая отделилась от ветви L3 — ветви азиатской Евы выходцев из Африки.
 Рис. 1.4
Если предки европейцев действительно были потомками неких аборигенных этнических групп из Северной Африки, таких, как берберы, и отделились от них около 45 тысяч лет назад, мы вправе надеяться отыскать древнейшие североафриканские генетические линии, отходящие прямо от основания ветви L3[77].
Однако, если взглянуть на представителей Северной Африки, в частности — тех же берберов, которые, как считается, являются исконным аборигенным населением этого региона, мы видим прямо противоположное. Оказывается, генетические ветви жителей Северной Африки показывают, что перед нами — либо позднейшие иммигранты из других регионов, либо потомки ветвей, очень далеко отстоящих от L3. Но в Северной Африке не обнаружено никаких следов представителей ветвей, совершивших первый исход из Африки — Насрин и Рохани. Зато представителей этих ветвей можно встретить в Индии (см. рис.). Действительно, мы видим, что Северная Африка активно заселялась этническими группами мигрантов с юга, имеющих генетические линии, типичные для предков европейцев и жителей Леванта. Древнейшая аборигенная генетическая линия мтДНК в Северной Африке, иногда отождествляемая с берберами, датируется периодом их прибытия с Леванта, что произошло около 30 тысяч лет назад. Эта линия образует отдельную подветвь (U6 — см. рис.) клана предков европейцев и жителей Западной Евразии (Западная Евразия — термин, обозначающий Европу и страны Леванта). На ветви U6 четко видны свидетельства исхода из Леванта или Европы, а не какого-либо окружного пути. Примерно 1/8 материнских генетических линий в Северной Африке восходит к потомкам позднейших миграций из глубинных районов Африки в окрестностях Сахары, и более половины из них — следы еще более поздних переселений народов на юг, преимущественно из Европы. Наконец, гены другого дочернего клана Евы выходцев из Африки, азиатской супергруппы М, представители которой сосредоточены в Индии, полностью отсутствуют в Европе, на Леванте и в Северной Африке. Поэтому крайне маловероятно, что Северная Африка могла послужить источником генетического материала для жителей Азии, как то утверждает схема, предложенная Джонатаном Кингдоном[78].
Все сказанное является аргументом в поддержку точки зрения о том, что Европа и Северная Африка являлись реципиентами (восприемниками) миграций с востока. Другими словами, у нас нет доказательств в пользу того, что первый северный исход был направлен из окрестностей Сахары в Северную Африку. Он скорее происходил в обратном направлении. А теперь давайте кратко рассмотрим, как и где именно на Востоке возникли и сформировались ветви под условными названиями «Насрин», «Рохани» и «Европа».
Рис. 1.4
Если предки европейцев действительно были потомками неких аборигенных этнических групп из Северной Африки, таких, как берберы, и отделились от них около 45 тысяч лет назад, мы вправе надеяться отыскать древнейшие североафриканские генетические линии, отходящие прямо от основания ветви L3[77].
Однако, если взглянуть на представителей Северной Африки, в частности — тех же берберов, которые, как считается, являются исконным аборигенным населением этого региона, мы видим прямо противоположное. Оказывается, генетические ветви жителей Северной Африки показывают, что перед нами — либо позднейшие иммигранты из других регионов, либо потомки ветвей, очень далеко отстоящих от L3. Но в Северной Африке не обнаружено никаких следов представителей ветвей, совершивших первый исход из Африки — Насрин и Рохани. Зато представителей этих ветвей можно встретить в Индии (см. рис.). Действительно, мы видим, что Северная Африка активно заселялась этническими группами мигрантов с юга, имеющих генетические линии, типичные для предков европейцев и жителей Леванта. Древнейшая аборигенная генетическая линия мтДНК в Северной Африке, иногда отождествляемая с берберами, датируется периодом их прибытия с Леванта, что произошло около 30 тысяч лет назад. Эта линия образует отдельную подветвь (U6 — см. рис.) клана предков европейцев и жителей Западной Евразии (Западная Евразия — термин, обозначающий Европу и страны Леванта). На ветви U6 четко видны свидетельства исхода из Леванта или Европы, а не какого-либо окружного пути. Примерно 1/8 материнских генетических линий в Северной Африке восходит к потомкам позднейших миграций из глубинных районов Африки в окрестностях Сахары, и более половины из них — следы еще более поздних переселений народов на юг, преимущественно из Европы. Наконец, гены другого дочернего клана Евы выходцев из Африки, азиатской супергруппы М, представители которой сосредоточены в Индии, полностью отсутствуют в Европе, на Леванте и в Северной Африке. Поэтому крайне маловероятно, что Северная Африка могла послужить источником генетического материала для жителей Азии, как то утверждает схема, предложенная Джонатаном Кингдоном[78].
Все сказанное является аргументом в поддержку точки зрения о том, что Европа и Северная Африка являлись реципиентами (восприемниками) миграций с востока. Другими словами, у нас нет доказательств в пользу того, что первый северный исход был направлен из окрестностей Сахары в Северную Африку. Он скорее происходил в обратном направлении. А теперь давайте кратко рассмотрим, как и где именно на Востоке возникли и сформировались ветви под условными названиями «Насрин», «Рохани» и «Европа».
Исход был только один
Наиболее убедительный аргумент против того, что основной маршрут, по которому предки европейцев и прочих рас современных людей совершили исход с Черного континента, пролегал по Северу Африки, — это структура материнского генетического древа всех народов мира. Как показано на рис. 1.4, лишь весьма и весьма небольшой побег одной из ветвей (Ева выходцев из Африки), объединивший около полудюжины основных материнских кланов Африки, сумел выжить после того, как его представители покинули родной континент, чтобы расселиться по всему остальному миру. Именно от этой малочисленной группы и произошли впоследствии все современные расы, обитающие в мире за пределами Африки. Совершенно ясно, что если имел место только один исход, то его участники должны были избрать какой-то один из двух возможных маршрутов, по которым можно было покинуть Африку. Я лично придаю особую важность тому простому и вместе с тем уникальному факту, что в генах всех неафриканских рас и народов на нашей планете прослеживается одна-единственная африканская генетическая линия. В рамках любого исхода из Африки могла присутствовать смесь генетических линий Евы, восходящих к различным потенциальным основоположникам. То же самое можно сказать и о любой произвольно собранной группе людей. И тем не менее выжить смогла только одна из этих генетических линий. Этот факт позволяет допустить, сколько попыток отдельных групп совершить исход из Африки могли оказаться успешными. Допустим, изначально существовало пятнадцать различных в генетическом отношении типов или линий мтДНК, которые покинули Африку в составе группы мигрантов (их могло быть и больше, и меньше; см. рис. 1.5). Это число вполне реалистично; даже в наши дни существует именно пятнадцать материнских генетических линий, возраст которых превышает 80 тысяч лет.
 Рис 1.5
Эти линии можно представить в виде пятнадцати типов мраморных камешков, оказавшихся в одном мешочке. Из этих пятнадцати линий только одна, митохондриевая ветвь, спустя много поколений превратилась в Еву выходцев из Африки, то есть общую «материнскую линию» для всего остального человечества. Другими словами, уцелел всего один камешек, представляющий только один тип мрамора. Этот произвольный процесс отбора и вымирания получил название дрейфа генов, потому что первичное смешение линий как бы совершило «дрейф» в сторону одного-единственного генетического типа.
Механизм, лежащий в основе дрейфа генов, достаточно прост. Время от времени некоторые материнские генетические линии отмирают, поскольку в них не остается дочерей, способных произвести на свет жизнеспособное потомство. Такое угасание и вымирание генетических линий показано на рис. 1.5. В пределах небольших изолированных этнических групп это со временем приводит к преобладанию какой-либо одной наследственной линии. В небольших группах дрейф играет огромную, решающую роль. Любопытный современный пример дрейфа генов, наблюдаемый, правда, по отцовской линии: в небольших, почти изолированных от внешнего мира, альпийских или валлийских горных деревушках через несколько поколений почти все жители оказываются обладателями одной и той же фамилии — скажем, Шмидт или Эванс — независимо от того, чем занимаются их сегодняшние носители[79].
Если мы возьмем две идентичные этнические группы, участвовавшие в исходе из Африки, и изолируем их в двух разных регионах (два мешочка с камешками, если продолжить нашу аналогию), через какое-то время в каждой из этих групп возобладает одна генетическая линия. Поскольку дрейф генов всегда связан с произвольным вымиранием, дрейф в обеих группах, скорее всего, не сведется к доминированию одних и тех же линий (шанс такого развития событий тоже есть, но они невелики — 1 к 15, если, разумеется, исходное число линий в обеих группах будет одинаковым). Если же мы возьмем две разные этнические группы, происходящие из разных регионов Африки (скажем, мигранты по северному и южному маршрутам; см. ниже) и жившие в разные эпохи, разделенные десятками тысяч лет, картина генетических линий в этих группах будет совершенно разной, ибо она будет отражать принципиально разные типы отбора линий африканских предков. И в данном случае вероятность выделения и преобладания одной и той же линии будет крайне мала.
Не надо обладать выдающимися познаниями в области статистического анализа, чтобы понять: вероятность того, что в двух таких независимых и малочисленных группах мигрантов, совершавших свой исход в разные эпохи и двигавшихся из Африки в разных направлениях, в результате произвольного дрейфа генов может возникнуть доминирование одной и той же материнской генетической линии, исчезающе мала. Она сравнима с вероятностью того, что в результате поочередного извлечения камешков из обоих мешочков в них в итоге останутся совершенно идентичные камешки. Но если миграций было не одна, а две, это со всей неизбежностью означает, что у мигрантов из Африки должно было быть как минимум две Евы. Другими словами, гены ясно говорят нам, что если за пределами Африкивыжила и сохранилась только одна митохондриевая генетическая линия, это значит, что существовала всего одна замкнутая этническая группа, которая совершила исход из Африки и стала впоследствии родоначальницей всего населения земного шара. От этой немногочисленной группы спустя множество поколений произошли и аборигены Австралии, и китайцы, и европейцы, и индусы, и полинезийцы. Но их древнейшие гены покинули Африку одновременно в той самой группе исхода.
Разумеется, если принять этот унитаристский аргумент, получается, что, раз северный маршрут не мог вывести из Африки предков будущих аборигенов Австралии, логично предположить, что от этих мигрантов вообще не мог произойти никакой другой народ за пределами Африки. Но прежде чем отвергнуть северный маршрут миграции как таковой и признать реальность исхода по другому, южному маршруту, давайте хорошенько ущипнем себя и спросим: соответствует ли вся совокупность генетических фактов теории одного-единственного исхода? Самым естественным ответом будет ответ «да», особенно если обратиться к отслеживанию других генетических маркеров, помимо митохондриевой ДНК. Если рассмотреть генеалогию Y-xpoмосомы, нетрудно заметить, что все неафриканские народы происходят от одной-единственной, опять-таки африканской отцовской наследственной линии, которую принято обозначать Ml68. В целом ряде исследований, основанных на использовании генетических маркеров, передаваемых обоими родителями, ученые обнаружили свидетельства одного-единственного исхода из Африки[80].
Другой весьма важный вопрос, который хотелось бы отметить здесь, — это точность датировки. Если обратиться к генетической линии африканской родоначальницы Евы выходцев из Африки, от которой произошли все неафриканские народы, нетрудно заметить, что возраст ее формирования — около 80 тысяч лет тому назад[81] (см. рис. 0.3). Это не совпадает с данными датировки по археологическим и генетическим данным, согласно которым колонизация Леванта и Европы происходила гораздо позже — всего лишь около 50 тысяч лет тому назад.
В отличие от весьма разветвленной структуры генетических линий в Северной Африке, в Африке существует еще один, совершенно особый район, характеризующийся невероятным многообразием генетических линий, располагающихся практически параллельно не только друг к другу, но и основанию ствола древа. Этот район — Эфиопия. Ее зеленые холмы и невысокие горы возвышаются над окрестными пустынями и засушливыми саваннами Африканского Рога[82]. Однако Эфиопия расположена к юго-востоку от Сахары, буквально у самых врат южного маршрута исхода. Генетики в наши дни в спешном порядке изучают генофонд жителей Эфиопии, некоторые из которых вполне могут оказаться потомками того самого этнического ядра группы исхода, покинувшего Африку южным путем. Это вынуждает нас обратиться к рассмотрению южного маршрута.
Рис 1.5
Эти линии можно представить в виде пятнадцати типов мраморных камешков, оказавшихся в одном мешочке. Из этих пятнадцати линий только одна, митохондриевая ветвь, спустя много поколений превратилась в Еву выходцев из Африки, то есть общую «материнскую линию» для всего остального человечества. Другими словами, уцелел всего один камешек, представляющий только один тип мрамора. Этот произвольный процесс отбора и вымирания получил название дрейфа генов, потому что первичное смешение линий как бы совершило «дрейф» в сторону одного-единственного генетического типа.
Механизм, лежащий в основе дрейфа генов, достаточно прост. Время от времени некоторые материнские генетические линии отмирают, поскольку в них не остается дочерей, способных произвести на свет жизнеспособное потомство. Такое угасание и вымирание генетических линий показано на рис. 1.5. В пределах небольших изолированных этнических групп это со временем приводит к преобладанию какой-либо одной наследственной линии. В небольших группах дрейф играет огромную, решающую роль. Любопытный современный пример дрейфа генов, наблюдаемый, правда, по отцовской линии: в небольших, почти изолированных от внешнего мира, альпийских или валлийских горных деревушках через несколько поколений почти все жители оказываются обладателями одной и той же фамилии — скажем, Шмидт или Эванс — независимо от того, чем занимаются их сегодняшние носители[79].
Если мы возьмем две идентичные этнические группы, участвовавшие в исходе из Африки, и изолируем их в двух разных регионах (два мешочка с камешками, если продолжить нашу аналогию), через какое-то время в каждой из этих групп возобладает одна генетическая линия. Поскольку дрейф генов всегда связан с произвольным вымиранием, дрейф в обеих группах, скорее всего, не сведется к доминированию одних и тех же линий (шанс такого развития событий тоже есть, но они невелики — 1 к 15, если, разумеется, исходное число линий в обеих группах будет одинаковым). Если же мы возьмем две разные этнические группы, происходящие из разных регионов Африки (скажем, мигранты по северному и южному маршрутам; см. ниже) и жившие в разные эпохи, разделенные десятками тысяч лет, картина генетических линий в этих группах будет совершенно разной, ибо она будет отражать принципиально разные типы отбора линий африканских предков. И в данном случае вероятность выделения и преобладания одной и той же линии будет крайне мала.
Не надо обладать выдающимися познаниями в области статистического анализа, чтобы понять: вероятность того, что в двух таких независимых и малочисленных группах мигрантов, совершавших свой исход в разные эпохи и двигавшихся из Африки в разных направлениях, в результате произвольного дрейфа генов может возникнуть доминирование одной и той же материнской генетической линии, исчезающе мала. Она сравнима с вероятностью того, что в результате поочередного извлечения камешков из обоих мешочков в них в итоге останутся совершенно идентичные камешки. Но если миграций было не одна, а две, это со всей неизбежностью означает, что у мигрантов из Африки должно было быть как минимум две Евы. Другими словами, гены ясно говорят нам, что если за пределами Африкивыжила и сохранилась только одна митохондриевая генетическая линия, это значит, что существовала всего одна замкнутая этническая группа, которая совершила исход из Африки и стала впоследствии родоначальницей всего населения земного шара. От этой немногочисленной группы спустя множество поколений произошли и аборигены Австралии, и китайцы, и европейцы, и индусы, и полинезийцы. Но их древнейшие гены покинули Африку одновременно в той самой группе исхода.
Разумеется, если принять этот унитаристский аргумент, получается, что, раз северный маршрут не мог вывести из Африки предков будущих аборигенов Австралии, логично предположить, что от этих мигрантов вообще не мог произойти никакой другой народ за пределами Африки. Но прежде чем отвергнуть северный маршрут миграции как таковой и признать реальность исхода по другому, южному маршруту, давайте хорошенько ущипнем себя и спросим: соответствует ли вся совокупность генетических фактов теории одного-единственного исхода? Самым естественным ответом будет ответ «да», особенно если обратиться к отслеживанию других генетических маркеров, помимо митохондриевой ДНК. Если рассмотреть генеалогию Y-xpoмосомы, нетрудно заметить, что все неафриканские народы происходят от одной-единственной, опять-таки африканской отцовской наследственной линии, которую принято обозначать Ml68. В целом ряде исследований, основанных на использовании генетических маркеров, передаваемых обоими родителями, ученые обнаружили свидетельства одного-единственного исхода из Африки[80].
Другой весьма важный вопрос, который хотелось бы отметить здесь, — это точность датировки. Если обратиться к генетической линии африканской родоначальницы Евы выходцев из Африки, от которой произошли все неафриканские народы, нетрудно заметить, что возраст ее формирования — около 80 тысяч лет тому назад[81] (см. рис. 0.3). Это не совпадает с данными датировки по археологическим и генетическим данным, согласно которым колонизация Леванта и Европы происходила гораздо позже — всего лишь около 50 тысяч лет тому назад.
В отличие от весьма разветвленной структуры генетических линий в Северной Африке, в Африке существует еще один, совершенно особый район, характеризующийся невероятным многообразием генетических линий, располагающихся практически параллельно не только друг к другу, но и основанию ствола древа. Этот район — Эфиопия. Ее зеленые холмы и невысокие горы возвышаются над окрестными пустынями и засушливыми саваннами Африканского Рога[82]. Однако Эфиопия расположена к юго-востоку от Сахары, буквально у самых врат южного маршрута исхода. Генетики в наши дни в спешном порядке изучают генофонд жителей Эфиопии, некоторые из которых вполне могут оказаться потомками того самого этнического ядра группы исхода, покинувшего Африку южным путем. Это вынуждает нас обратиться к рассмотрению южного маршрута.
Южный маршрут
Чтобы лучше понять перемены «коридоров» (т.е. маршрутов) исхода, по которым прошли потенциальные эмигранты из Африки, нам придется вернуться далеко назад, к первым людям, дерзнувшим покинуть Черный континент. Многие миллионы лет назад, когда древнейший суперконтинент Пангея[83] находился в процессе активного распада, от него откололась Африка, причем на месте разлома образовалась глубокая трещина в земной коре. Эта громадная трещина в земной коре связана с обширной системой бесчисленных ущелий и долин в Восточной Африке. Наиболее заметное следствие такого разлома — глубокая пропасть, образовавшаяся между Африкой и Аравийским полуостровом и более известная как Красное море. К северу от Красного моря находится Суэцкий перешеек, а у южной его оконечности — узкий перешеек шириной 25 км, лежащий на глубине 137 м ниже уровня моря. Он известен под названием Врата Скорби (Баб-аль-Мандаб) и образован многочисленными рифами.
 Фото 5. Красное море. Внизу — Врата Скорби, где ок. 80 тыс. лет тому назад имел место исход из Африки. (Цвета — компьютерная графика.)
Фото 5. Красное море. Внизу — Врата Скорби, где ок. 80 тыс. лет тому назад имел место исход из Африки. (Цвета — компьютерная графика.)
Около 2 млн. лет тому назад Африка еще соединялась с Аравийским полуостровом, и на месте нынешнего пролива лежала суша. Многие виды млекопитающих, обитавших в Африке и Евразии, могли без всякого труда мигрировать между Африкой и Аравией как у северной, так и у южной оконечности будущего Красного моря. В ту эпоху Африка удалялась от Евразии со скоростью примерно 15 мм в год, постепенно обнажая перешеек и столь же неспешно закрывая южные врата Африки. Сравнительно недавние находки свидетельствуют о том, что в числе последних видов млекопитающих, совершивших исход из Африки еще до того, как Врата Скорби были окончательно затоплены и навсегда закрылись, оказались другие наши кузены — представители вида Homo erectus (см. Фото 2), которые унесли с собой свои простейшие каменные орудия. В ту эпоху, пока и северные, и южные врата Африки еще были открыты, наши кузены быстро расселились в восточном направлении, направившись в Юго-Восточную Азию через Индию и на север, через земли Леванта, где около 1,8 млн. лет назад достигли Кавказа[84]. Затопление Врат Скорби водами океана стало далеко не единственной преградой, возникшей на пути миграций млекопитающих и человека в Азию. Здесь уместно вспомнить как минимум еще две таких преграды. Первая из них, уже упоминавшаяся выше, представляла собой длинную горную гряду — так называемую цепь Загрос-Таурус. Эти молодые горы появились на поверхности Земли в предшествующие несколько миллионов лет, практически одновременно с Гималаями. Этот каменный барьер, протянувшийся от современной Турции на западе до Ирана на востоке, служил практически непреодолимым препятствием на пути из стран Леванта в Индию, Пакистан и на Дальний Восток еще с тех пор, как люди современного типа впервые покинули просторы Африки Второй сухопутный барьер отделил северные врата Африки от южных (см. рис. 1.6). В наши дни он носит название Аравийской пустыни, или, выражаясь более мрачным языком, «пустой стороны света». Одно из самых засушливых мест на нашей планете, Аравийская пустыня резко расширила свою площадь, заняв практически весь Аравийский полуостров к тому моменту, когда нынешняя ледниковая эпоха, плейстоцен, резко заявила о себе. Произошло это около 2 млн. лет тому назад. Ледниковая эпоха обычно означает, что климат становится холоднее и с поверхности океана испаряется значительно меньше влаги, а это, в свою очередь, приводит к тому, что в полосе пустынь начинает выпадать гораздо меньше осадков. Во время одного из самых суровых ледниковых периодов, подобных тому, который человечество пережило около 18 тысяч лет назад, громадные массы воды оказываются как бы законсервированными в приполярных ледяных панцирях, толщина которых может составлять более километра. Во время ледникового максимума уровень океана значительно понизился, в результате чего нормальная циркуляция водных масс между Индийским океаном и Красным морем практически прекратилась[85]. За последние 200 тысяч лет, уже во время существования на Земле человека современного типа, были два кратких периода, когда Красное море сильно мелело[86] и превращалось в активно испаряющееся соленое озеро. Подавляющее большинство видов планктона погибло. И хотя Красное море становилось почти безжизненным, в его устье все же оставался очень узкий, шириной в несколько километров, водный канал, разделенный на множество рукавов рифами и островками.

Эти периоды сильного оледенения, словно оазисы в Сахаре, были довольно редкими и непродолжительными И когда они наступали, в среднем через каждые 100 тысяч лет за последние 2 млн лет, жители Африки могли легко покинуть родной континент по южному маршруту, переправившись через узкое устье Красного моря на примитивных плотах, используя островки в качестве перевалочных пунктов (Разумеется, другие крупные млекопитающие не могли использовать плоты) Но для мигрантов, которым удавалось переправиться на азиатский берег, Аравийский полуостров не выглядел особо привлекательным пристанищем, ибо в периоды активного наступления ледника там всегда царила невыносимая засуха Впрочем, и там, особенно на Йеменском нагорье в районе Адена, тоже встречались редкие оазисы и островки зелени К тому же на южном побережье Аравийского полуострова чувствовалось благотворное влияние муссонов[87]. Таким образом, за последние 2 млн лет люди могли как минимум трижды переправиться из Африки в Азию по этому южному маршруту[88]. После того, как люди около 2 млн лет тому назад впервые перебрались на азиатский берег, врата полосы пустынь, отделявшие Европу и Средний Восток от Центральной и Южной Азии, надолго закрылись. Мигранты, совершившие исход из Африки по южному маршруту, могли попасть в Индию только кружным путем, продвигаясь вдоль побережья, а тем, кто предпочел северный маршрут, оставалось направиться сперва на Левант, а затем на Кавказ и далее в Европу Все эти преграды и препятствия на пути миграции древних людей в Азию и обусловили причудливый характер колонизации человеком остальных континентов нашей планеты. Когда в Африке появился новый вариант генов Homo, некоторые туземные племена во время очередной теплой междуледниковой паузы могли предпочесть северный маршрут исхода, тогда как другие уже в эпоху сильного оледенения могли выбрать южный маршрут. Первые люди, избравшие северный маршрут во время междуледниковой паузы, наступившей около 1,8 млн. лет тому назад, достигли урочища Дманиси в Грузии. По внешнему облику они, видимо, были еще весьма близки к виду Homo habilis, древнейшему и более примитивному африканскому виду, чем его восточно-азиатский современник — вид Homo erectus. И где-то на полпути между ранним африканским видом Homo erectus (Homo ergaster) и Homo habilis возник особый вид, так называемый Homo georgicus[89]. Это параллельное освоение новых территорий людьми двух разных древнейших видов, протекавшее по различным маршрутам, является серьезным аргументом в поддержку гипотезы о том, что исходы человека из Африки по южному маршруту, имевшие место еще до того, как он оказался окончательно закрытым, объясняются скорее чисто географическими возможностями, доступными тогда для любых африканских млекопитающих, чем некими гипотетическими особенностями эволюции Homo erectus, как считалось прежде. Вид, останки которого были найдены в Дманиси, давно вымер на Кавказе и Леванте в результате резкого похолодания. Как мы уже знаем, подобная же трагическая участь постигла людей и спустя 2 млн. лет, после так называемой эмианской междуледниковой паузы. Когда на землях Леванта несколько тысячелетий назад началась страшная засуха, людям, жившим в районе Дманиси, не удалось спастисъ ни на севере, ни на юге, ни на востоке, и они вымерли все до единого. На смену им пришли новые мигранты, представители вида Homo heidelbergensis, потомки которых оказались более удачливыми «колонизаторами» Европы, заселение которой, по всей видимости, происходило в три этапа за последние 800 тысяч лет. Последняя из таких миграций с севера, видимо, имела место примерно 250 тысяч лет тому назад. К тому времени у африканских представителей вида Homo, хотя они по-прежнему имели густые брови и надбровные дуги, развился значительно более крупный мозг, и они начали делать более сложные и эффективные орудия эпохи Среднего палеолита. Фоули и Лар называют предка этих последних выходцев из Африки Homo helmei.[90] Эти исследователи утверждают, что Homo helmei, перебравшись в Европу, эволюционировал, став нашим основным и ближайшим сородичем — Homo neanderthalensis. (См. Фото 6 и главу 2.)

Типы черепов (сверху, слева направо): Australopithecus, Homo rudolfensis, Homo erectus. Homo rhodesiensis, неандерталец и человек современного типа.

Фото 6. Реконструкция головы неандертальца. Тяжелые черты лица, надбровные дуги и крупный нос — это элементы адаптации к холодному климату севера.
Примерно за такой же период (а именно 160—800 тысяч лет) произошло приблизительно столько же массовых миграций предков современного человека на Дальний Восток. Эти азиаты, обладатели крупного мозга, были очень близки, хотя и не абсолютно идентичны своим сородичам из Европы. Подобно тому, как благодаря миграции в Европу у неандертальцев развился очень большой мозг, позднейшие миграции из Африки, имевшие место около 200 тысяч лет тому назад, также могли способствовать появлению видов с большим объемом мозга, таких, как черепа маба и дали, найденные в Китае[91], а возможно, и череп «Мадлен», обнаруженный на Яве. (См. рис. 0.2.) Мы можем даже, хотя и весьма приблизительно, датировать последнюю из миграций из Африки по южному маршруту эпохой до появления человека современного вида. Долина Нармада в центральной Индии была родиной архаического вида человека, череп которого с трудом поддается датировке, однако есть все основания считать, что люди этого вида жили более 150 тысяч лет тому назад. И хотя от той эпохи до нас дошло крайне мало ископаемых останков людей предсовременного вида, Индия буквально изобилует находками древнейших каменных артефактов, созданных ими. Наиболее ранние из них, орудия так называемой ашолийской культуры (Нижний палеолит), найденные в Индии, можно приближенно датировать эпохой от 160 до 670 тысяч лет тому назад, тогда как орудия Среднего палеолита были занесены из Африки в Индию в конце этого периода, то есть около 160 тысяч лет тому назад[92]. Это свидетельствует о том, что последние переселения людей предсовременного типа из Африки могли иметь место примерно 160 тысяч лет назад, во время засушливого ледникового периода, на заре человечества современного типа. Следы окончательного перехода от орудий Среднего палеолита к орудиям Верхнего палеолита можно встретить как в Центральной Нармада, так и в бассейне реки Средней Сон в северной Индии. Находки таких орудий связаны со слоями вулканического пепла, образовавшимися в результате грандиозного взрыва вулкана Тоба на острове Суматра, происшедшего около 74 тысяч лет тому назад. Это свидетельствует о радикальных технико-культурных переменах, происшедших примерно в то же время[93]. Что касается орудий Верхнего палеолита, то они явились отличительной особенностью первых людей современного типа, прибывших на Левант и в Европу, что произошло примерно на 25—30 тысяч лет позже.
Ногам холодно?
Южный маршрут, пролегавший через Красное море, становившийся более доступным, когда на Земле наступало очередное оледенение, в известной мере служит аргументом в пользу наших собственных воззрений на исход из Африки. По моему мнению, это произошло не во время особенно теплой эмианской междуледниковой паузы, наступившей около 125 тысяч лет тому назад, а примерно на 45 тысяч лет позже, уже на пороге длительной ледниковой эпохи. Если мы тщательно проанализируем свидетельства этой древней миграции и попытаемся сузить и уточнить ее датировку, перед нами возникают три очевидных, взаимосвязанных между собой вопроса: ради чего мы (то есть наши далекие предки) переправились через Красное море, когда именно это произошло и почему мы не сделали этого прежде. Ответ на последний из этих вопросов (почему же мы так долго выжидали, прежде чем во время очередного ледникового периода совершить совсем небольшой — какихнибудь 11 км — переход через море? Ведь даже устья многих великих рек имели куда большую ширину) будет, пожалуй, наиболее показательным. Мы, люди, существа не только волевые, агрессивные и нетерпеливые, но и изобретательные и отважные. Нам и в голову не пришло бы устраивать массовые экспедиции на другие планеты или хотя бы отправиться за море, на другой континент, если нас не устраивает тамошний климат, о котором мы знаем по донесениям разведчиков. Мы непременно захотели бы убедиться в том, есть ли у нас шансы выжить на обратном пути, если нам придется возвращаться обратно. Реконструкция поэтапного продвижения предков полинезийцев по островам Тихого океана за последние несколько тысяч лет способна поведать нам массу интересного. Судя по тому, как быстро люди современного типа расселились вдоль побережья Индийского океана и достигли Австралии, предки современных азиатов вполне могли воспользоваться примитивными плотами или лодками, чтобы много раз пересечь узкий пролив в устье Красного моря за те 70 тысяч лет, которые их предки провели в Восточной Африке. Что же им мешало? Что их останавливало? Ответ очевиден: по всей вероятности, на противоположном берегу пролива уже жили другие племена, о существовании которых могли узнать разведчики мигрантов. Очевидно, эти существа были не людьми современного типа, а их ближайшими сородичами — представителями азиатской ветви потомков африканского вида Homo helmei. В пользу этой гипотезы говорят найденные в Индии артефакты и останки, о которых мы уже упоминали, а также многочисленные орудия эпохи Среднего палеолита, встречающиеся по другую сторону моря — в Йемене и на юге Аравийского полуострова. В одной из последних книг о происхождении человека[94] использована сцена соседства многих видов, взятая из знаменитого фильма «Звездные войны». Это было сделано ради того, чтобы подчеркнуть, что за несколько десятков тысяч лет до появления на Земле людей современного типа на ней уже жило немало видов человека, мирно сосуществововших на нашей планете на протяжении длительных исторических эпох, не вступая в соперничество и не истребляя друг друга. Однако это скорее объясняется тем фактом, что представители каждого из этих подвидов обитали на отдельных континентах и островках, а не тем, что они по природе были добрыми соседями. Когда представители различных видов человека предпринимали попытки поделить между собой тот или иной континент, результатом этого обычно становилось быстрое сокращение численности какого-либо вида. Находки человеческих окаменелостей в Африке показывают, что появление человека современного типа ознаменовалось быстрым исчезновением наших прародителей — представителей вида Homo helmei — на той же территории. Останки последних из них датируются около 130 тысяч лет тому назад. Нам остается лишь гадать, явилось ли практически полное исчезновение всех африканских гоминидов во время крайне холодного и засушливого периода около 140—190 тысяч лет тому назад простым совпадением с появлением людей современного типа, оставшихся вскоре единственным видом человека на Земле, или же эти новички попросту провели «зачистку» Африки от бедных Homo helmei. Однако наши предположения основаны на печальном опыте совсем недавних попыток и кровавых актов геноцида, имевших место в Тасмании, Германии, Руанде и на Балканах. Уничтожение коренных жителей Тасмании, происходившее в XIX в., явилось единственным примером полного расового уничтожения, предпринятого в рамках кампании по истреблению целого народа, и происходило сравнительно быстро, по всей вероятности — в связи с тем, что жертвы цивилизованных варваров были окружены со всех сторон и просто не могли спастись бегством с острова, ставшего их огромной братской могилой. Таким образом, судя по современным историческим свидетельствам, мы, люди, готовы и способны без колебания уничтожать своих соседей[95] — собратьев по роду человеческому. А раз так, есть все основания полагать, что за несколько тысяч лет наши предки могли преспокойно истребить всех представителей других видов человека на всем континенте[96]. Первые шаги переселенцев на новом континенте обычно оказывались самыми трудными. Необычайно успешная доисторическая экспансия из Юго-Восточной Азии, в процессе которой предки современных полинезийцев расселились по островам Индийско-Тихоокеанского региона, дает нам некоторое представление о том, как протекали вторжения на новые территории. Хотя аборигены-полинезийцы обладали превосходными навыками судоходства и кораблестроения, о чем свидетельствовал еще знаменитый капитан Джеймс Кук, они неизменно избегали высаживаться на островах, где, как им было известно, существовало значительное туземное население. В тех случаях, когда в ходе длительных плаваний им не оставалось ничего другого, кроме как совершить вынужденную высадку на земле «дружественного» острова, например, на побережье большого и густонаселенного острова Новая Гвинея, они много тысячелетий спустя после великих древних миграций по-прежнему устраивали свои колонии в буквальном смысле слова «на прибрежной полосе»[97]. И лишь в последние 500 лет небольшие отряды захватчиков, обладающие более совершенным вооружением и боевой техникой, стали совершать заокеанские плавания и высаживаться на новом, давно и густо населенном континенте[98]. Став единственным выжившим видом, люди современного типа около 130 тысяч лет тому назад сделались безраздельными хозяевами Африканского континента. Если, что вполне вероятно, древнейшие обитатели Восточной Африки современного типа знали, что на противоположном берегу Красного моря, в Йемене, живут их соседи — люди архаического типа, с тяжелыми надбровными дугами, не уступающие им в силе и сноровке, они могли счесть, что с них вполне достаточно и доминирующего положения в родной Африке и что благоразумная осмотрительность — неотъемлемая составляющая доблести. Нам очень хотелось бы узнать, чего, собственно, ждали обитатели африканского берега Красного моря, но это станет возможно лишь тогда, когда будет проведено детальное и всестороннее исследование останков и артефактов представителей палеолитического населения Йемена. Однако результаты одних из первых раскопок, проведенных под руководством кембриджского археолога Гертруды КэйтонТомпсон, со всей очевидностью показали, что люди жили в Йемене уже в эпоху палеолита[99].
Пора в путь
В последние годы было проведено немало исследований, результаты которых позволили лучше понять образ жизни «обитателей прибрежной полосы», который можно считать одним из побудительных мотивов, заставивших африканцев современного типа покинуть родной континент и двинуться в дальний путь вдоль побережья Индийского океана (см. рис. 1.6). На протяжении львиной доли времени из тех 2 млн. лет, которые человек существует на Земле, люди просто бродили по просторам саванн, ведя малопримечательный образ жизни охотников и собирателей. Подобно современным обитателям пустыни Калахари на юге Африки, они быстро поняли выгоды совместной охоты и собирательства, благодаря которому их пищевой рацион пополняли всевозможные съедобные коренья, плоды и листья растений. Когда во время длиннейшей ледниковой эпохи, наступившей примерно 130—190 тысяч лет тому назад, площади саванн начали быстро сокращаться, кому-нибудь из мудрецов древних людей могла прийти в голову мысль попробовать употреблять в пищу моллюсков и всевозможные виды морепродуктов, которые можно подобрать прямо на берегу. Таким образом, вполне возможно, что собирательство на прибрежной полосе начало активно развиваться в еще более ранние эпохи, но, поскольку древние прибрежные пространства давно ушли под воду, мы этого уже никогда не узнаем. Такая диета, богатая белком, весьма благотворна для умственного развития, да и добыть эти продукты куда проще, чем охотясь на крупных млекопитающих. Итак, прибрежные лакомства имели немалое преимущество перед прочими видами пищи, будучи доступными даже тогда, когда климат и почва в саваннах становились совсем засушливыми в результате очередного ледникового периода. И тем не менее получить материальные доказательства собирательства на прибрежной полосе неожиданно легко, поскольку люди обычно оставляли после себя характерные груды пустых (расколотых) раковин от моллюсков. Однако возникает проблема: как установить, как давно образовались эти груды раковин? Дело в том, что груды раковин обычно находят несколько выше уровня кромки моря во время прилива, тогда как на протяжении большей части последних 200 тысяч лет уровень моря был на много метров ниже современного. Это означает, что у нас есть все основания ожидать, что подавляющее большинство древнейших груд раковин моллюсков бесследно исчезли, за исключением тех, что образовались, когда уровень моря был необычайно высоким, например, во время теплых междуледниковых пауз типа той, что имела место около 125 тысяч лет назад. 60 тысяч лет назад по прибрежной полосе Испании и Италии бродили неандертальцы, кормившиеся собирательством, и вполне возможно, что они занесли этот обычай с собой из Африки. Вплоть до недавнего времени наиболее ранние свидетельства практики собирательства на прибрежной полосе были обнаружены в устье реки Класис в Южной Африке; их возраст — от 100 до 115 тысяч лет. Однако в 2000 г. новые доказательства собирательства на прибрежной полосе были обнаружены в Абдуре (см. Фото 7), на западном побережье Эритреи в Красном море, к северу от Врат Скорби. В открытой там стоянке древнего человека, находящейся на самом берегу, возраст которой — около 125 тысяч лет, были обнаружены раздробленные кости крупных млекопитающих, что свидетельствует о смешанном рационе питания древних людей. Орудия, в числе которых оказался и нож из вулканического стекла — обсидиана, практически наверняка были сделаны руками людей современного типа[100].
 Фото 7. Первый в мире «устричный бар» был обнаружен в Абдуре, Эритрея, на рифе, возраст которого — 125 тыс. лет. На фото хорошо видны окаменелые остатки мяса устриц и обсидиановые орудия.
Фото 7. Первый в мире «устричный бар» был обнаружен в Абдуре, Эритрея, на рифе, возраст которого — 125 тыс. лет. На фото хорошо видны окаменелые остатки мяса устриц и обсидиановые орудия.
Огромный интерес, который вызвала эта уникальная находка на берегу Красного моря, объясняется двумя факторами: во-первых, она является наиболее древним во всем мире свидетельством собирательства на прибрежной полосе, а во-вторых, она расположена совсем близко от крайней точки южного маршрута исхода из Африки. Оба эти фактора хорошо вписываются в весьма заманчивую гипотезу, которую можно назвать «поход собирателей в Австралию». В ней мы имеем вполне правдоподобную историю о собирателях на прибрежной полосе, которые задерживались на одном участке до тех пор, пока берег не мог больше их прокормить, а затем перебирались на следующий необжитый участок, потом — на следующий и так далее. Благодаря подобному способу миграции передовые группы мигрантов, переправившиеся через Красное море, могли продвигаться все дальше и дальше по кромке береговой полосы Индийского океана, «проедая» себе путь на восток, и через каких-нибудь 10 тысяч лет добраться до Индонезии. Низкий уровень моря, установившийся в ту эпоху, вполне мог позволить им совершить сухопутную прогулку из Адена до Явы, а оттуда, перебираясь с острова на остров, попасть в Австралию, где на древнейших стоянках аборигенов археологи находят точно такие же груды пустых раковин. Я абсолютно уверен, что эта гипотеза о раннем заселении Австралии в принципе соответствует действительности, но даты нуждаются в уточнении. Это объясняется не только археологическими находками, но и показаниями молекулярных часов на генетическом древе всех народов и рас, заселивших Евразию. Если, следуя этой гипотезе о «походе собирателей в Австралию», предположить, что источником заселения Австралии и всего остального мира были группы мигрантов, покинувшие Африку в результате одного-единственного исхода, мы можем с достаточно высокой надежностью прогнозировать последовательность этапов и даты колонизации en route[101] в Индию и Юго-Восточную Азию, а также на параллельном маршруте в Новую Гвинею. Эти прогнозы и станут критерием оценки достоверности гипотезы.
Когда и ради чего мы покинули Африку?
Одна из принципиальных проблем, связанных с так называемыми генетическими молекулярными часами, заключается в том, что мы вынуждены учитывать довольно широкие рамки погрешностей. Таким образом, хотя мы можем быть вполне уверены, что генетическое древо свидетельствует о том, что успешный исход был всего один, приблизительные данные генетической датировки показывают, что это могло произойти где-то между 70 тысяч и 95 тысячами лет тому назад[102]. Таким образом, получается, что наши восточноафриканские предки собирали груды пустых раковин на западном берегу Красного моря целых 55 тысяч лет (начиная с условной даты 125 тысяч лет тому назад), прежде чем решиться отправиться в дальний путь. Прежде чем приступить к поиску археологических свидетельств такого исхода, весьма полезно попытаться определить, каковы же были климатические условия и влажность в ту доисторическую эпоху развития человечества. Не могло ли продолжительное усугубление ледниковой ситуации вызвать понижение уровня моря у Врат Скорби до такой степени, что наши предки могли преспокойно перейти на другой берег по рифам, поднимающимся со дна? Усиление засушливости климата на побережье Восточной Африки могло повлечь за собой резкое сокращение запасов и источников питьевой воды, а также радикально изменить муссонный характер климата на побережье южного Йемена и сделать эти места более привлекательными для жизни собирателей на прибрежной полосе. Кроме того, переход вброд через Красное море, к берегам Аравии, мог помочь мигрантам избавиться от последних страхов перед незнакомыми соседями. Впрочем, не исключено, что последние, не обладая навыками и пищевым универсализмом собирателей, могли покинуть Аравию из-за засухи и постоянной нехватки воды. Несмотря на соблазнительность библейских аллюзий при подобном сценарии развития событий, основная проблема, связанная с возможным сухопутным исходом через устье Красного моря, предположительно имевшим место около 60—90 тысяч лет тому назад, заключается в том, что данные океанографии решительно отвергают возможность того, что морское дно в районе Врат Скорби могло полностью пересохнуть в недавнем историческом прошлом. Такие климатические аномалии за последние полмиллиона лет имели место по меньшей мере трижды: 440 тысяч лет тому назад, 140 тысяч лет тому назад и, наконец, во время последнего ледникового максимума — около 18 тысяч лет тому назад[103]. Во время ледниковых периодов пролив в устье Красного моря был гораздо уже, что позволяло мигрантам без особого труда перебираться с островка на островок по отмелям и коралловым рифам у островов Ханиш-аль-Кубра у северной оконечности перешейка. Как мы уже знаем, это — тот самый путь, по которому, по всей вероятности, некогда прошел Homo erectus, предок будущих обитателей Азии, и для наших предков-первопроходцев не представляло особых проблем за какие-нибудь несколько тысяч лет достичь берегов Австралии. Однако, хотя это и не коснулось Красного моря, в эпоху первого исхода из Африки действительно имело место сильное похолодание климата (рис. 1.7). Замеры, выполненные на ледяной «шапке» Гренландии, показали, что второе по интенсивности похолодание за последние 100 тысяч лет имело место между 60 и 80 тысячами лет тому назад[104]. В период максимальной интенсивности, около 70 тысяч лет тому назад, это оледенение привело к тому, что уровень Мирового океана находился примерно на 80 м ниже современной отметки. Способ, посредством которого этот минимальный уровень, как его обычно называют, удалось измерить в Красном море, явился неожиданным ключом к тому, что могло послужить для древних людей стимулом, побудившим их отправиться в путь

 Рис. 1.7 Основные этапы заселения мира
Рис. 1.7 Основные этапы заселения мира
Океанографы могут определять максимальные уровни Мирового океана в междуледниковых паузах, изучая рост коралловых рифов. Минимальные же уровни океана, возникающие в периоды ледниковых максимумов, выявить гораздо труднее. Илко Ролинг, специалист-океанограф из Саутгемптонского университета, похоже, нашел способ, позволяющий решить эту проблему применительно к Красному морю. Ролинг замерял уровни планктона в Красном море в доисторические эпохи. Он установил, что в период ледникового максимума Врата Скорби в устье моря, образно говоря, закрывались, и Красное море оказывалось изолированным от Индийского океана. Интенсивное испарение влекло за собой быстрое повышение солености моря, в результате чего основная масса планктона, служившего основанием цепи питания обитателей моря, гибла. В междуледниковых же паузах, типа той, которую мы переживаем теперь, высокий уровень океана позволял Красному морю растворять и вымывать излишек солей, в результате чего жизнь вновь возвращалась в его сонные воды. Но минимальный уровень океана, аналогичный тому, что имел место около 70 тысяч лет тому назад, не мог полностью заблокировать устье Красного моря, и поэтому планктон в нем хотя и сильно пострадал, однако не исчез полностью. Низкий уровень численности планктона в морских отложениях позволил Ролингу подтвердить прежние оценки снижения уровня Мирового океана в ту эпоху. Напомним, такое снижение достигало 80 м по сравнению с современным уровнем[105]. Массовая гибель планктона усугублялась резким снижением содержания кислорода в морской воде и высокими температурами самой воды на отмелях и мелководных участках, таких, как отмели у берегов Абдура в Эритрее, возле устья Красного моря. А низкие уровни планктона в Красном море, в свою очередь, могли повлиять на шансы выжить и вообще на успех миграции собирателей на прибрежной полосе. Что же касается побережья Аденского залива и Йемена на другом берегу моря, то для них, наоборот, характерны насыщенные кислородом и богатые планктоном воды, приносимые течениями Индийского океана. Другими словами, на южном берегу Аравийского полуострова условия жизни для собирателей, бродивших вдоль кромки воды, были, по всей вероятности, поистине превосходными[106]. Итак, острая нехватка источников пищи на западном побережье Красного моря, заманчивые южные берега Аденского залива и прохладный, влажный климат Йеменского нагорья, способного послужить прибежищем для мигрантов, и стали теми факторами, которые побудили наших предков отправиться в путь. Остается ответить на вопрос: когда это произошло? Выжидали ли они момента, когда прокормиться собирательством на прибрежной полосе стало больше невозможно (это могло случиться около 70 тысяч лет тому назад), или же отправились в дальнее странствие гораздо раньше, когда ситуация с питанием еще только начинала ухудшаться? Уровни планктона в Красном море могут ответить и на этот вопрос. Дело в том, что количество планктона зависело от уровня океана не только в районе Врат Скорби. Изменения розы ветров у северо-восточных муссонов ознаменовали собой максимум оледенения и повлияли на содержание кислорода и уровни испарения вод Красного моря. Один из видов планктона, особенно чувствительный к этим изменениям, начал активно вымирать примерно 80 тысяч лет тому назад[107]. Быть может, это — след, оставленный нашими предками-собирателями на их долгом пути?.. Свидетельства другой крупнейшей климатической катастрофы, имевшей место в эпоху, когда уровень океана был очень низким, были получены в пробах грунта из скважин, просверленных в дне Персидского залива[108]. Этой катастрофой было извержение вулкана Тоба на острове Суматра, происшедшее около 74 тысяч лет тому назад. Извержение это, по праву считающееся самым мощным за последние 2 млн. лет, представляло собой колоссальный взрыв, последствием которого стала долгая «ядерная зима». Грандиозный выброс вулканического пепла, направленный в основном на северо-запад, накрыл Индию, Пакистан и весь регион Персидского залива, усеяв громадные территории слоем пепла толщиной от 1 до 3 м. Если наши предки покинули Африку около 80 тысяч лет тому назад, их потомки спустя 6 тысяч лет наверняка испытали на себе последствия извержения вулкана Тоба на Суматре (см. рис. 1.7), ибо собиратели, продвигавшиеся вдоль прибрежной полосы Индийского океана, неизбежно должны были оказаться в зоне самого грандиозного выпадения вулканического пепла за всю историю человечества. Поэтому извержение вулкана Тоба является уникальной временной вехой, поскольку пепел, усеявший громадные территории Южной Азии, поддается достаточно точной датировке, особенно когда удается обнаружить его нетронутый слой. Малазийский археолог Зуранина Маджид провел специальное исследование материальных остатков культуры, созданной людьми современного типа, в лесистой долине в штате Перак, неподалеку от Пенанга. Возраст древнейшей традиции эпохи палеолита, известной под названием культуры Кота-Тампан, насчитывает как минимум несколько десятков тысячелетий. На одной из стоянок найдены орудия, буквально утопающие в слое вулканического пепла из вулкана Тоба[109]. Если причастность человека современного типа к созданию этих орудий подтвердится, это будет означать, что люди современного типа обитали в Юго-Восточной Азии еще до извержения вулкана Тоба, то есть более 74 тысяч лет тому назад. А это, в свою очередь, делает вполне реальной датировку исхода — 80 тысяч лет тому назад. Генетические и прочие свидетельства присутствия человека в Австралии около 70 тысяч лет тому назад также говорят в пользу этой версии. Извержение вулкана Тоба накрыло Индийский субконтинент толстым слоем пепла. Трудно представить, как первые переселенцы в Индию вообще смогли пережить столь страшную катастрофу. Таким образом, мы вправе предположить, что в результате нее все обитатели обширных территорий между Восточной и Западной Азией погибли. И действительно, резкое разграничение, или «борозда», между востоком и западом четко прослеживается на генетическом древе. Но как подобная ранняя датировка исхода соотносится с генетическими данными? Вполне возможно, что это — наиболее противоречивая и загадочная эпоха в истории человечества. Краткий ответ заключается в том, что генетические данные и расположение ветвей древа хорошо согласуются с ранней датировкой исхода (см. главы 3 и 4). Это позволяет также дать ответ на вопрос о происхождении европейцев, затронутый мною раньше: как могло получиться, что Европа была заселена лишь около 50 тысяч лет тому назад, но ее население происходит от того же общего предка, что и жители Австралии и Азии.
Первые азиатские кланы Адама и Евы
Как мы убедились выше, небольшое число кланов-родоначальников Евы, выходцев из Африки, обосновались в Йемене и оказались изолированными от африканского побережья. Через много поколений все генетические линии свелись к одной-единственной митохондриевой линии Евы выходцев из Африки, известной также под сухим техническим индексом L3. От L3 вскоре произошли два дочерних клана по женской линии: Насрин (N) и сестринский ему клан Манью (М) (см. рис. 1.4). Наиболее древнее и разветвленное семейство потомков Манью обнаружено в Индии, тогда как Насрин, в свою очередь, стала единой праматерью всех европейцев. Дело в том, что дочерей (то есть потомков по женской линии) Манью в Западной Евразии практически нет. Любопытно, что в наши дни потомков Насрин и Манью можно встретить на земном шаре практически всюду, за исключением Западной Евразии (см. рис. 0.3). Этот факт сам по себе свидетельствует о том, что наследственные генетические линии практически всех коренных жителей Австралии, Америки, Сибири, Исландии, Европы, Китая и Индии восходят к одной-единственной линии потомков выходцев из Африки. Он свидетельствует, что в древности имел место всего один исход. После исхода уникальные африканские наследственные признаки ветви L3 подверглись дрейфу генов и были утрачены за исключением малочисленных локальных групп. В итоге остались только Насрин и Манью и их потомки по женской линии. Таким образом, мы можем проследить родословные практически любого жителя Земли, за исключением африканцев, стоящего в очереди у кассы магазина где-нибудь во Внешней Монголии[110], Алис-Спрингс или Чикаго вплоть до древнейшей группы, совершившей исход из Африки. Правда, существует несколько исключений, как обычно, подтверждающих правило. Одно из них — ветвь U6 (уже упоминавшаяся выше), представители которой вернулись с Леванта в Северную Африку. Другое — ветвь M1, представители которой переправились через Красное море в Эфиопию примерно во времена последнего ледникового периода. Откуда нам это известно? Не так давно вышел в свет впечатляющий труд эстонских генетиков Томаса Кивисилда и Рихарда Виллемса, убедительно показавший, что представители ветви М1 вовсе не вернулись в Африку, как клан Манью — в Азию[111]. Другими словами, ветвь M1 — это представители позднейшей волны заселения Восточной Африки, пришедшие из Азии. Родословная линия дочерей Евы, Манью, прослеживается в генах только жителей Азии, а не европейцев. Обратившись к рассмотрению древнейших ветвей Манью в Азии, мы получим такие даты: 74 тысячи лет для клана Манью в Центральной Азии, 75 тысяч лет для аборигенов Новой Гвинеи и 68 тысяч лет — для аборигенов Австралии. В Индии, как я уже говорил, присутствует наибольшее богатство и разнообразие подветвейМанью; более того, вполне возможно, что именно Индия — родина Манью на ветви L3. А локальный возраст одного из субкланов Индии (М2) составляет 73 тысячи лет[112]. Если бросить взгляд на генетический след наших отцов, записанный в Y-хромосоме, нетрудно заметить аналогичную картину. Из всех африканских мужских наследственных линий, существовавших еще до исхода, только одна дала начало всем остальным мужским линиям или кланам неафриканского ареала. Линия Адама выходцев из Африки дала начало трем первичным мужским наследственным линиям за пределами Африки, в отличие от двух основных женских линий (Насрин и Манью). Для простоты и удобства идентификации я дал этим кланам, известным генетикам как линии С, D/E (YAP) и F, названия по именам трех сыновей Адама: «Каин», «Авель» и «Сиф» . Как и у линии Евы выходцев из Африки, у линии Адама есть потомки, живущие в наши дни в Африке, в окрестностях Сахары. В случае Адама это линия Авеля, имеющая ветви как в Азии, так и в Африке и Западной Евразии. Последняя ветвь имеет особенно много представителей, особенно среди народности банту, которая не так давно мигрировала с севера на юг африканского континента, причем ее переселение сопровождалось весьма драматическими событиями. Этим я пока и ограничусь. Более подробная дискуссия о линиях трех сыновей Адама представлена в главах 3—7 нашей книги.
Истоки происхождения европейцев
Генетическое древо сообщает нам о предках европейцев и жителей Леванта нечто весьма любопытное и неожиданное: оказывается, они пришли на эти земли не прямо из Африки, а откуда-то с юга, из районов, граничащих с Индией. Их родоначальница-матриарх Насрин была, по всей вероятности, более западной относительно двух азиатских дочерей Евы, появившихся на свет во время долгого пути вдоль прибрежной полосы. Она отличается от Манью главным образом тем, что ее потомков-дочерей можно встретить во всех неафриканских народах по всему земному шару: в Евразии, Австралии и Северной и Южной Америке. Ее отличие от клана Манью, потомки которого не встречаются в Европе и странах Леванта, означает, что разделение ветви Насрин на жителей Востока и Запада могло произойти примерно в том районе, где собиратели и обитатели прибрежной полосы проникли в Индию. Согласно данным генетической датировки, азиатские и австралийские потомки ветви Насрин как минимум имеют столь же древний возраст, как и потомки Манью[113]. Наиболее вероятным пунктом маршрута с точки зрения рождения Насрин можно считать Персидский залив. Правда, в ту засушливую эпоху это был не столько залив, сколько оазис мелких пресноводных озер, питаемых обширными подземными резервуарами, а также двумя великими реками — Тигром и Евфратом. Это живописное пустынное прибежище, по всей вероятности, существовало и в последующие несколько десятков тысяч лет, ибо, хотя эти западные кланы возникли и сформировались на юге, мы не имеем никаких генетических или археологических подтверждений того, что дочери Насрин и Рохани могли появиться в Европе или на землях Леванта ранее чем 45—50 тысяч лет тому назад. Как обычно, у позднейшей колонизации Европы представителями ветви потомков Насрин — выходцев из района Персидского залива — имелись вполне конкретные климатические причины, и мы действительно можем обнаружить их в пробах грунта, взятых из глубинных скважин, которые были пробурены в дне Индийского океана неподалеку от побережья Индии, точнее — в дельте Инда. Как сказано выше, путь из Сирии к побережью Индийского океана издревле был заблокирован полосой непроходимых пустынь, возникших в ледниковую эпоху. Примерно 50 тысяч лет тому назад в Южной Азии наступило непродолжительное, но весьма интенсивное потепление климата и начался бурный рост растительности, а муссонные ритмы были более благоприятными для жизни, чем в наши дни. Это позитивное изменение климата подтверждают богатые каменным углем слои подводных отложений в той же дельте Инда[114]. Но поскольку такое потепление продолжалось очень недолго, возможно — всего несколько тысяч лет, геологи обычно называют его «промежуточным», а не «междуледниковой паузой». Однако влияние этого потепления на Плодородный Полумесяц в Ираке было не менее интенсивным и благотворным. В результате потепления на какое-то время открылся узкий зеленый коридор (см. рис. 1.8), позволивший обитателям побережья Персидского залива мигрировать в Сирию, а прапраправнуки азиатской Евы двинулись дальше, на северо-запад, на территорию Плодородного Полумесяца на Леванте (см. главу 3). Благодаря генетическому отслеживанию удалось выявить первоначальный южноазиатский источник мигрантов в Европу. Как показывает генетическое древо, одна из ветвей Насрин обращена на северо-запад, в Европу, тогда как другая ветвь Насрин направлена на восток, в Индию.
 Рис.1.8
Такая картина генетических корней европейцев — корней, тянущихся из Южной Азии, опровергает основные постулаты гипотезы северного маршрута исхода из Африки Она также в корне меняет европоцентрическую точку зрения, согласно которой европейцы явились создателями первой культуры людей современного типа за пределами Африки. Чтобы быть совсем точными, позволим себе задать один вопрос: существуют ли археологические свидетельства, противоречащие утверждению о том, что первые европейцы были потомками выходцев из Южной Азии. Таких свидетельств попросту нет, зато доказательств обратного более чем достаточно.
Наиболее ранние археологические свидетельства присутствия в Европе технологий, характерных для Верхнего палеолита, датируются временем не ранее 47 тысяч лет тому назад. Бельгийский археолог Марсель Отте бросил смелый вызов общепринятым взглядам, согласно которым предки современных европейцев были выходцами из Северной Африки, создавшими по пути развитую культуру Верхнего палеолита. Он указывает на горную гряду Загрос на этнической территории современного Курдистана, расположенную к северу от побережья Персидского залива, и выделяет этот район в качестве ядра формирования технических достижений Верхнего палеолита[115].
Шри-Ланка, в те времена — полуостров на юге Индии, является местом во многом уникальным в том смысле, что он опровергает расхожие представления о развитии Востока и Запада. Имеются достоверные свидетельства о создании на Шри-Ланке микролитов, уверенно датируемых 28 тысячами лет тому назад. Эти крошечные, тщательно отделанные каменные ножи появились в Европе и на Леванте лишь около 10 тысяч лет тому назад. Но поистине ошеломляющей находкой можно считать другие ножи, обнаруженные в слое, залегающем гораздо ниже первого. Возраст найденных в нем микролитов кажется просто невероятным — от 64 до 74 тысяч лет тому назад! Если достоверность этой датировки подтвердится, ее можно считать бесспорными следами исхода из Африки, имевшего место около 70 тысяч лет назад[116].
Рис.1.8
Такая картина генетических корней европейцев — корней, тянущихся из Южной Азии, опровергает основные постулаты гипотезы северного маршрута исхода из Африки Она также в корне меняет европоцентрическую точку зрения, согласно которой европейцы явились создателями первой культуры людей современного типа за пределами Африки. Чтобы быть совсем точными, позволим себе задать один вопрос: существуют ли археологические свидетельства, противоречащие утверждению о том, что первые европейцы были потомками выходцев из Южной Азии. Таких свидетельств попросту нет, зато доказательств обратного более чем достаточно.
Наиболее ранние археологические свидетельства присутствия в Европе технологий, характерных для Верхнего палеолита, датируются временем не ранее 47 тысяч лет тому назад. Бельгийский археолог Марсель Отте бросил смелый вызов общепринятым взглядам, согласно которым предки современных европейцев были выходцами из Северной Африки, создавшими по пути развитую культуру Верхнего палеолита. Он указывает на горную гряду Загрос на этнической территории современного Курдистана, расположенную к северу от побережья Персидского залива, и выделяет этот район в качестве ядра формирования технических достижений Верхнего палеолита[115].
Шри-Ланка, в те времена — полуостров на юге Индии, является местом во многом уникальным в том смысле, что он опровергает расхожие представления о развитии Востока и Запада. Имеются достоверные свидетельства о создании на Шри-Ланке микролитов, уверенно датируемых 28 тысячами лет тому назад. Эти крошечные, тщательно отделанные каменные ножи появились в Европе и на Леванте лишь около 10 тысяч лет тому назад. Но поистине ошеломляющей находкой можно считать другие ножи, обнаруженные в слое, залегающем гораздо ниже первого. Возраст найденных в нем микролитов кажется просто невероятным — от 64 до 74 тысяч лет тому назад! Если достоверность этой датировки подтвердится, ее можно считать бесспорными следами исхода из Африки, имевшего место около 70 тысяч лет назад[116].
Резюме
Противоречия, отмечаемые в имеющихся на сегодняшний день археологических и антропологических данных, могут быть сняты путем отслеживания неискаженного генетического материала по линии нашей праматери Евы в обратной перспективе, вплоть до самых первых людей современного типа, появившихся в Африке. Генетические свидетельства позволяют нам сфокусировать внимание на древнейших миграциях наших непосредственных предков В результате мы можем выделить одну-единственную группу наших предков, совершивших около 70 тысяч лет тому назад исход из Африки. Принципиальное отличие от общепринятых взглядов, предполагающих множественные исходы из Африки, охватывает все последующие маршруты миграций в истории человечества, начало которым положил один-единственный исход из Африки по южному маршруту. Первопроходцы, покинувшие Африку, продвигались вдоль побережья Аравийского полуострова к Персидскому заливу, на берегу которого они и основали первую колонию людей с запада, потомки которых много тысячелетий спустя заселили Европу Что же касается пути из Индии на Восток, то это совсем другая история
ГЛАВА ВТОРАЯ КОГДА ЖЕ МЫ СТАЛИ ЛЮДЬМИ СОВРЕМЕННОГО ТИПА?
Узколобая «зацикленность» европейцев на собственном прошлом и своем мнимом превосходстве в области материальной культуры начиная с эпохи палеолита на протяжении последнего столетия не раз приводила к целому ряду необоснованных постулатов. Грубо говоря, они сводятся к тому, что мы (ибо я тоже принадлежу к европейцам) будто бы первыми начали мыслить абстрактными категориями, первыми первыми обрели дар речи, освоили искусство живописи и резьбы, научились шить одежду и ткать и, наконец, начали обмениваться товарами. В главе 1 я уже говорил о том, что подобное ложное допущение о том, будто все эти технико-культурные навыки были неведомы и недоступны нашим африканским предкам, подкреплялось утверждением, что европейцы и обитатели Леванта были потомками исхода по северному маршруту и в этническом отношении резко отличались от мигрантов, ставших родоначальниками австралийцев и азиатов. Однако от подобной точки зрения не останется и камня на камне, если со всей серьезностью отнестись к свидетельствам о том, что генетические линии европейцев и других народов Запада представляют собой дальние ветви потомков одной-единственной южноазиатской группы, расселившейся по всему миру за пределами Африки. В этой главе я доказываю, что стремление любой ценой поместить на переднем крае прогресса человечества людей современного типа, живших в Европе и на Леванте, противоречит другим, более соответствующим истине взглядам на древнейшую историю Востока и Запада. Бесспорные генетические свидетельства того, что люди современного типа — это выходцы из Африки, оставившие на своей прародине ближайших сородичей, потомки которых и сегодня живут в Африке и, естественно, являются людьми современного типа в полном смысле этого слова, наносят сокрушительный удар по устоявшимся взглядам людей Запада на современных обитателей Африки. И хотя предвзятость подобных взглядов очевидна, менталитет некоторых европейских археологов остается неизменным.Спор сторонников исхода из Африки и мультирегионалистов
Археологи продолжают упорно доказывать, что существует целый ряд врожденных и основополагающих особенностей поведения человека, резко отличавших в древности первых европейцев современного типа от их ближайших европейских сородичей, неандертальцев, а также — исходя из той же системы аргументации — от их непосредственных предков в Африке. Одной из причин такого противопоставления, возможно, является стремление опровергнуть гипотезу о мультирегионализме и доказать, что мы происходим отнюдь не от неандертальцев. В подобных аргументах как бы подразумевается предположение не только о том, что древнейшие европейцы современного типа (кроманьонцы) были первыми, кто сумел развить некие новые навыки и передать их представителям следующих поколений, но и о том, что неандертальцы и в биологическом отношении были еще не вполне людьми. Если развивать дальнейшую аргументацию подобного рода, то придется признать, что анатомически современные предшественники кроманьонцев, жившие в Африке, не обладали достаточно развитым интеллектом, чтобы создать технические достижения эпохи Верхнего палеолита. Другими словами, европейцы были далеко не первыми людьми на нашей планете, которые обрели дар речи, овладели искусством живописи и резьбы и впоследствии, много тысячелетий спустя — так, по крайней мере, считали многие европейцы, — обучили всем этим культурным навыкам аборигенов Австралии и Африки, не говоря уж о жителях Азии. Возможно, я в чем-то заблуждаюсь и сгущаю краски, но при чтении ряда недавних публикаций у меня сложилось именно такое впечатление. Прежде чем перейти к рассмотрению внешних аргументов против европоцентричной теории, мне хотелось бы сказать несколько слов о том, что же могло привести к возникновению столь искаженной системы воззрений. Проблема возникла еще в 1856 г., сразу же после находки первого черепа неандертальца. С самого начала неандертальцы, что называется, получили негативные отзывы в прессе: их начали высмеивать, называя бровастыми идиотами. Этот образ нашел яркое воплощение в короткой новелле знаменитого писателя-фантаста Герберта Уэллса «Люди-гризли», написанной в 1921 г. Под пером Уэллса неандертальцы, в отличие от современных и вполне разумных кроманьонцев, предстали тупыми и мрачными монстрами. Попытки реабилитации неандертальцев в качестве наших потенциальных сородичей, ни в чем не уступающих нам, до сих пор неизменно заканчивались неудачей, поскольку даже сами их апологеты, возносившие неандертальцам неумеренные похвалы, невольно бросали тень на них, а всевозможные документальные фильмы неизменно сосредотачивали основное внимание на их внешности, обходя молчанием их огромный мозг. Увы, истина заключается в том, что мы по-прежнему склонны считать неандертальцев людьми более низкой стадии развития, а эксперты не устают подчеркивать культурные различия между нами и ними. Если сопоставить достижения материальной культуры неандертальцев и успехи их современников, первых европейцев современного типа, может сложиться впечатление, что наше предвзятое отношение к ним не лишено оснований. Люди современного типа создавали наскальные рисунки, расписывали стены пещер и даже изображали себе подобных, а также оставили множество резных фигурок. Что касается неандертальцев, то до сих пор нет никаких свидетельств, что они создавали нечто подобное. Люди современного типа создавали произведения не только из камня, но из других материалов — кости, раковин, рога, а также всевозможные резные фигурки, словом — артефакты, создателями которых, как считалось до недавнего времени, неандертальцы быть никак не могли. Между тем такие артефакты находят в разных районах Европы, отстоящих друг от друга на многие сотни километров, что можно считать свидетельством торговли или обмена, которых у неандертальцев опять-таки не было и быть не могло. Чем же объяснить столь широкий разброс? Ведь неандертальцы вообще не использовали такие материалы. Подобные свидетельства контактов и сотрудничества между регионами, находящимися достаточно далеко друг от друга, служили дополнительными аргументами в пользу того, что люди современного типа обладали куда более сложной и развитой системой социальных отношений, чем неандертальцы. Последние, согласно расхожим представлениям, жили небольшими изолированными группами, членам которых попросту не хватило интеллектуальных возможностей, чтобы адаптироваться к изменяющимся климатическим условиям, хотя Клайв Гэмбл сообщает, что особые высококачественные камни-заготовки для изготовления изделий транспортировались в пределах Европы на 300 с лишним километров задолго до эпохи Верхнего палеолита[117]. Люди современного типа устраивали каменные очаги и хоронили умерших, что также было нехарактерно для культуры неандертальцев. Наконец, если присмотреться к самым многочисленным и долговечным посланиям из далекого прошлого — каменным орудиям, мы без труда сможем заметить резкое различие между изделиями неандертальцев и людей современного типа. Последние делали ножи — тонкие отщепы от каменной заготовки, длина которых более чем вдвое превышала их ширину. Кроманьонцы, первые европейцы современного типа, по всем привычным критериям были людьми «поющими и танцующими» по сравнению с мрачными, угрюмыми и малоподвижными неандертальцами. Чем же это объяснить? Стандартный ответ на этот вопрос всегда сводился к тому, что мы, видимо, находимся на более высокой ступени биологического развития — по крайней мере, в том, что касается развития умственных способностей. За неандертальцами же всегда признавалось превосходство в физической силе; у них были более крепкие и толстые кости по сравнению с нашим тонким и хрупким скелетом, что довершало картину противопоставления мускулов и мозга. Резкий контраст в физических данных и культурные различия между двумя типами человека широко использовались, чтобы подчеркнуть наступление биологической эры европейцев современного типа — существ разумных и деятельных, но подобные аргументы не отличались особой логичностью и убедительностью. Что еще мы знаем о наших ближайших сородичах? Среди главных свойств, отличающих человека от животных, следует назвать способность к абстрактному, ассоциативному мышлению, и, разумеется, дар речи. Вправе ли мы предположить, что неандертальцы были лишены этого дара, то есть, другими словами, были существами «немыми», не наделенными речью? Конец подобным измышлениям не положил даже тот факт, что в их черепе имелась так называемая подъязычная кость (гиоид), практически такая же, как гиоиды у людей современного типа. Существует широко распространенное мнение, что все новые навыки, которыми обладают современные европейцы, явились результатом активности особого гена или группы генов, появившегося у них около 40—50 тысяч лет тому назад (см. Пролог). Первые и самые впечатляющие открытия, связанные с проявлением творческого потенциала человека (искусство и разного рода технические инновации эпохи Верхнего палеолита, датируемые примерно 18—35 тысячами лет тому назад), действительно были сделаны в Европе. Однако это объясняется тем, что Европа — тот самый регион, где впервые возникла археология как наука и уроженцами которого были практически все наиболее выдающиеся археологи за последние 150 лет. Нам всем хорошо знакомы удивительное изящество, реалистичность и точность передачи натуры, присущая наскальным рисункам, обнаруженным в Ласко и Шове на юге Франции (см. Фото 8).
 Фото 8. Величественные наскальные рисунки из Шове, например этот носорог, ошибочно считаются знаками появления человека или начала эпохи Европы.
Фото 8. Величественные наскальные рисунки из Шове, например этот носорог, ошибочно считаются знаками появления человека или начала эпохи Европы.
Давно стали общим местом восторженные отзывы об этих памятниках древнего искусства и утверждения, что в Европе в конце позднего каменного века (обычно именуемого европейским Верхним палеолитом) в развитии художественной культуры произошел настоящий взрыв, знаменовавший собой наступление эпохи человека современного типа. Некоторые ученые, переводя эти восторги в более конкретную плоскость, заявляют, что до той эпохи «анатомически современные люди», останки которых, находимые в Африке, датируются около 130 тысячами лет тому назад, возможно, выглядели практически как современные люди, но еще «не были таковыми»[118]. Если, следуя той же системе аргументации, современные европейцы, образно говоря, появились на свет из куколки, обладая столь фантастически развитым гением, у них, вне всякого сомнения, должен был появиться некий биологический (т.е. передаваемый на генетическом уровне) элемент, который в прежние времена отсутствовал в нашем организме. Однако подобная аргументация ведет к весьма опасным выводам о том, что предки современных австралийцев и африканцев в биологическом отношении были людьми отсталыми и менее развитыми, чем предки европейцев. Стоп! Что же мы такое говорим?! Не напоминает ли это ситуацию, когда заносчивый горожанин приезжает в какой-нибудь небольшой поселок в глубинке и заявляет тамошним жителям: «Вы — неотесанная деревенщина, отсталые и биологически примитивные недочеловеки»? Не так ли поступает историк, утверждающий, что изобретение письменности и нотной грамоты, осуществление промышленной и аграрной революции всякий раз было результатом появления и действия новых генов? Будущие историки, сравнивая сложный уровень технократической цивилизации и доминирующее положение развитых стран Запада с культурами народов Папуа и Новой Гвинеи, оставшимися на уровне каменного века, поступят неблагоразумно, если вздумают отнести столь резкий контраст на счет неких биологических факторов. Многие из нас, или, по-видимому, даже большинство, бессознательно тешат себя иллюзиями, что другие расы и этнические группы явно уступают нам в развитии. Дело дошло до того, что видный американский биолог Джаред Дайамонд счел себя вправе назвать вещи своими именами. Он написал свою известную и ставшую бестселлером книгу «Пушки, эмбрионы и сталь»[119], чтобы доказать, что неравномерности в развитии и глобальном могуществе разных держав, скорее всего, являются результатом исторических факторов и благоприятного стечения обстоятельств, нежели врожденных интеллектуальных различий между разными этносами и расами. Он попытался объяснить, как и почему совсем небольшие отряды конкистадоров сумели практически уничтожить многолюдные цивилизации доколумбовской Америки.
Вопрос Яли
В самом начале своей книги Дайамонд приводит основной вопрос, заданный ему мудрым и популярным туземным вождем Яли, представителем одной из последних в мире неолитических культур, издревле существующей на северном побережье Новой Гвинеи (см. Фото 9). Яли спросил его: «Почему сложилось так, что вы, белые люди, производите так много грузов и привозите его в Новую Гвинею, а у нас, черных, грузов совсем мало?» (В том контексте, в котором Яли задал свой вопрос, «грузы» означают «импортируемые товары», например, мешки с рисом из Австралии, холодильники и прочие предметы роскоши, по меркам туземцев.) Сам Яли не был рядовым туземцем. Он был незаурядным человеком, посвятившим большую часть своей жизни осмыслению этого вопроса с магическо-религиозной точки зрения, характерной для культуры его племени. Дайамонд испытал на себе обаяние его личности и описывал его как необычайно восприимчивого, терпимого и деликатного человека.
 Фото 10. Новогвинейский лидер Яли во время совершения церемонии в честь культа товаров (карго) на северном побережье (1956).
Фото 10. Новогвинейский лидер Яли во время совершения церемонии в честь культа товаров (карго) на северном побережье (1956).
Яли одно время был лидером наиболее влиятельного культа товаров, возникшего в Новой Гвинее (в основе культа товаров лежало представление о том, что товары якобы можно создавать посредством особых ритуалов), и его особая притягательность повлияла не только на его собратьев, жителей Новой Гвинеи, но и на одного биолога. Уникальный дар харизматика и лидера, присущий Яли, получил свое отражение в другой книге — «Дорога принадлежит товарам»[120]. Эта книга, написанная известным австралийским антропологом Питером Лоуренсом, представляет собой, пожалуй, лучшее описание всевозможных культов товаров. Три главы в ней посвящены Яли и описанию его собственных культов. Мне лично довелось услышать рассказ о Яли из уст человека, лично знакомого с вождем и знавшего его лучше многих. В начале 1980-х гг. я работал в качестве врача в Маданге, родной провинции Яли в Новой Гвинее. Как-то раз, заболев, я сам стал пациентом провинциального госпиталя. Моим соседом по крошечной палате был старый, согбенный и дряхлый австралийский фермер. Мо Джонсон (так звали старика) страдал диабетом и постоянно жил в больничной палате с тех самых пор, как узнал, что его статус ветерана дает ему право на бесплатное лечение и питание. Симпатичный, но капризный старик, он не имел за душой ни гроша, кроме старенького коротковолнового радиоприемника, с помощью которого он нередко приводил в бешенство местную медсестру-самоанку. Както раз он рассказал мне историю своей жизни. Красношеий упрямец и сквернослов, он не мог обходиться без расистских эпитетов, как только речь заходила об аборигенах Новой Гвинеи. Это выглядело весьма странным в компании, с которой он общался. Дело в том, что его, лежавшего на больничной койке, часто навещали приветливые посетители, которые все до единого были коренными жителями Новой Гвинеи. Мо был одним из немногих выживших участников легендарного отряда «береговых наблюдателей» — радистов «кротов», которые в годы Второй мировой войны в прямом смысле слова зарывались в холмы в джунглях на самой кромке берега на островах южной части акватории Тихого океана. Эти храбрецы, по большей части — бывшие фермеры, добровольно вызвались остаться на оккупированной японцами территории, чтобы передавать по радио о перемещениях кораблей, авиации и войск противника Их донесения, как считается, сыграли важную роль в битве в Коралловом море. Большинство «береговых наблюдателей» были убиты, умерли от тропических болезней или были захвачены в плен и погибли в концентрационных лагерях Что касается Мо, героя войны, то он был обязан жизнью Яли — тому самому, о котором рассказывается в нашей истории. Яли был «туземным помощником» Мо и помог ему остаться в живых во время войны. Он вызволил Мо из японского лагеря для военнопленных, а затем сопровождал его на всем протяжении 300-километрового пути в Маданг, лежавшего через непроходимые джунгли и болота. После войны британская колониальная администрация признала Яли, также известного героя, харизматическим лидером и назначила ему продовольственный паек. Однако отношения между властями и Яли были прерваны, как только чиновники поняли, что имеют дело с туземным Мессией, а не марионеткой в своих руках. Как рассказывал мне Мо и как пишет в своей книге Питер Лоуренс, чиновники решили примерно проучить его. Они посадили Яли в тюрьму. Рассказывая об этом эпизоде, Мо произнес весьма знаменательные слова, невольно отражающие взгляды и язык того культурного социума, к которому он принадлежал, и вместе с тем свидетельствующие о его глубокой искренности. Вот что он сказал мне: «Стив, посадив его в тюрьму, они разбили ему сердце. А ведь Яли был лучше их. Он был выше всех. Да что там — он был настоящим белым...» Знакомые австралийцы уверяли меня, что эта фраза не несет в себе ни малейшего расистского подтекста. В действительности Яли не был ни белым, ни даже австралийцем. Зато он был щедро наделен тем, что Дайамонд и Лоуренс в один голос называли интеллигентной сообразительностью. И тем не менее, не обладая достаточными знаниями о чуждой культуре, он не мог ответить на вопрос, который задал Джареду Дайамонду. Впрочем, в те времена в Новой Гвинее не выпускались холодильники; откуда же Яли мог знать, как их производят? Однако по сути вопрос Яли имел в виду те грузы и предметы роскоши для белых колонистов, которые регулярно доставлялись на кораблях, бросавших якорь на верфи Маданга. Последователи культа товаров, в числе которых был и сам Яли, были убеждены, что все эти товары появляются на свет в результате неких тайных магических ритуалов, которыми обладали только белые. И несмотря на всю свою природную сообразительность и несомненно выдающийся интеллект, магическо-религиозные взгляды, свойственные культуре неолита, и крайне ограниченные познания не позволили ему понять, что за океаном может существовать мощный аграрный и промышленный комплекс, производящий все эти товары. Для объяснения причин возникновения веры в товары иногда используется формула «отсутствие знаний о средствах производства», однако это — явное упрощение сути вопроса Яли, ибо, как мы знаем, знание не влечет за собой автоматический отказ от веры в «сверхъестественные» силы. Жители побережья Маданга — неутомимые садоводы и огородники. Каждый садовод непременно выращивает на своем участке, помимо других овощей и фруктов, до тридцати разновидностей таро (особый вид корнеплодов), которые защищают растения от болезней и паразитов. Однако, несмотря на все свое искусство, садоводы Новой Гвинеи убеждены, что их успех зависит не от опыта и агротехники, а от неукоснительного соблюдения правил магии, связанной с возделыванием и выращиванием растений. Как отмечал Питер Лоуренс, они (то бишь садоводы) свято верили, что если их посевы погибли, то виной тому — неумелое совершение магических обрядов, а не неудачные агротехнические меры. В конце концов, всякому известно, как надо выращивать таро! Такой же была точка зрения аборигенов Новой Гвинеи и на все остальные аспекты культуры и ремесел: по их мнению, все эти знания были исключительно прерогативой знатоков, «посвященных», которые знали все необходимые магические ритуалы. Такие представления о главенствующей роли магии по сравнению с любыми «средствами производства» неизбежно распространялись и на экзотические импортные товары, средства производства которых оставались загадочными или не вполне понятными. Такого рода товары просто обязаны были быть созданы с помощью магии. Все попытки Яли выведать магию производства товаров и воспроизвести ее были обречены на провал, но он не мог осознать корень своих ошибок, ибо считал, что колонисты просто-напросто держат в тайне свои секреты. В свою очередь, белые колонисты считали (и говорили об этом вслух), что Яли и последователи его культа — глупцы и плуты. Естественно, они тоже заблуждались.
Не надо судить о людях по их культуре
Какое же отношение имеет сказанное к истокам происхождения человека и первых европейцев современного типа? А вот какое. Дело в том, что даже некоторые современные историки, изучающие доисторическую эпоху, допускают ту же ошибку — судят о потенциальных возможностях людей по их орудиям, изделиям и уровню «культурного развития». Они, эти историки, совершают серьезную ошибку, сравнивая «современных» кроманьонцев с «древними» неандертальцами, поскольку фокусируют внимание на действительно громадном контрасте в области материальной культуры между новоявленными пришельцами и исконными аборигенами. Более того, они бессознательно повторяют ту же ошибку, когда сравнивают кроманьонцев с более ранними анатомически современными людьми, жившими в Африке около 100 тысяч лет тому назад. Как мы уже знаем, люди современного типа активно заселяли Азию и Австралию задолго до того момента, когда одна из их ветвей двинулась в Европу. Их потомки до сих пор живут в тех регионах и имеют все основания, чтобы отвергать европоцентрическую точку зрения, согласно которой европейцы, осуществившие первую в истории индустриально-техническую революцию, и были первыми «настоящими людьми». Слабость подобной гипотезы очевидна: причина, по которой ученые нашли так много свидетельств в пользу торжества орудий эпохи Верхнего палеолита именно в Европе, заключается в том, что Европа попросту оказалась тем континентом, где люди искали также эти свидетельства с особой тщательностью. Громадные трудности с поиском свидетельств существования ранних форм искусства в Африке обусловлены временем, условиями сохранности и специфическими типами древних пещер. Произведения искусства — вещь крайне хрупкая, и наскальные рисунки и росписи, сделанные в древнейшую эпоху на скалах под открытым небом, просто не имели шансов уцелеть и дойти до нас. Действительно ли неандертальцы были существами «тупыми и примитивными» и правомерно ли сравнение технических достижений? Давайте для начала зададим вопрос: а смогли бы они, при прочих равных уровнях развития культуры, достичь тех выдающихся результатов, которых достигли люди современного типа. Обычная точка зрения сводится к тому, что они — не более чем обреченные неудачники. Они расселились по всей Европе, еще около 200 тысяч лет тому назад сумев хорошо адаптироваться к местным негативным факторам и в первую очередь — холоду, однако не отважились обосноваться в наиболее холодных районах. В отличие от них, люди нового типа, бывшие выходцами из тропической Африки, сумели как нельзя лучше приспособиться к жизни в любых, самых холодным местах, причем обжили их за довольно небольшой промежуток времени. Правда, следует отметить, что людям современного типа очень повезло: они могли заранее создать целый ряд технических и культурных новшеств. Но возникает вопрос: обладали ли люди современного типа способностью делать свои орудия иначе и лучше, чем неандертальцы? Другими словами, если мы бросим взгляд на людей современного типа еще до того, как они проникли в Европу, а также рассмотрим характер заселения ими остального мира, сможем ли мы обнаружить явное преимущество людей этого вида перед анатомически современным человеком и прочими их современниками более архаического типа, жившими в тех же регионах? Ответ будет однозначным: нет и еще раз нет.
Анатомически современные африканцы и технология производства каменных орудий
Люди анатомически современного вида появились на нашей планете как минимум 130 тысяч лет назад. На протяжении первых 100 тысяч лет из них подавляющее большинство людей этого вида создавали и использовали каменные орудия того же класса (или «типа»), что и неандертальцы. Технология производства таких орудий к тому времени уже прошла длительный и многоэтапный период развития. Эта технология, обычно называемая технологией Среднего палеолита, по всей вероятности, была изобретена около 200 тысяч лет тому назад африканскими представителями вида Homo helmei, которые, возможно, были общим предком для нас и неандертальцев (см. главу 1). По недоразумению, каменные орудия эпохи Среднего палеолита, обнаруженные в разных районах, нередко известны под разными терминами. Такого рода орудия, найденные в Европе, Северной Африке и странах Леванта, обычно называют артефактами мустерианской культуры — по названию стоянки древнего человека во Франции, где типичные образцы таких орудий были обнаружены рядом с останками Homo heidelbergensis и неандертальцев. Эти орудия, найденные в районах Африки, прилегающих к Сахаре, известны как орудия Среднего Каменного века[121]. Пожалуй, наиболее характерной чертой эпохи Среднего палеолита было заметное уменьшение размеров каменных орудий, появление черенков и рукояток, а также использование заранее обработанных каменных заготовок, из которых делались собственно орудия. На протяжении предшествующего периода, растянувшегося на добрый миллион лет, африканские представители вида Homo erectus, как правило, делали большие топоры, обрабатывая готовый каменный сердечник-заготовку или отщеп с обеих сторон, в результате чего после долгих трудов у них получался готовый топор. Наоборот, мастера эпохи Среднего палеолита первым делом тщательно обтесывали заготовку специальной формы (подобная техника возникла значительно раньше, предположительно около 350 тысяч лет тому назад), а уже потом откалывали от нее несколько отщепов. Затем производилась окончательная отделка готового изделия, которое сохраняло острую кромку, характерную для отщепа заданной формы[122]. Для удобства крепления у отщепа делался короткий черенок. Не считая пару-другую исключений, африканцы современного типа, жившие к югу и северу от Сахары, примерно до рубежной даты — 50 тысяч лет тому назад — продолжали пользоваться орудиями типа Среднего Каменного века (и более ранних типов). Как мы уже говорили в Главе 1, первые африканцы современного типа, покинувшие континент, основали свою неудавшуюся и недолго просуществовавшую колонию к северу от Сахары, на подступах к Леванту. Это произошло между 90 и 125 тысячами лет тому назад. Они, как этого и следовало ожидать от выходцев из Северной Африки, использовали орудия типа Среднего палеолита, весьма напоминавшие орудия, которыми пользовались неандертальцы, раньше их покинувшие Африку, и, естественно, орудия неандертальцев, которые вскоре вновь заселили земли Леванта.
Мигранты современного типа использовали аналогичные технические приемы
Я подчеркиваю: у нас есть все основания ожидать, что первые люди современного типа, покинувшие Африку около 100 тысяч лет тому назад, использовали по большей части технологию, типичную для Среднего палеолита, поскольку не обнаружено никаких свидетельств того, что в те времена в Северной и Восточной Африке могла существовать принципиально новая техника производства орудий. Датируемые около 125 тысяч лет тому назад орудия, найденные на стоянке собирателей на прибрежной полосе неподалеку от устья Красного моря — последнее, на мой взгляд, было наиболее вероятным местом исхода людей современного типа, — относились к эпохе Среднего палеолита, хотя не обнаружено никаких человеческих останков, которые могли бы помочь идентифицировать их создателей. Те же самые аргументы с равным правом можно отнести и к орудиям, которыми пользовались их непосредственные предки, мигрировавшие в Азию. Орудия типа Среднего палеолита впервые появились в Индии около 150 тысяч лет тому назад, но в большей мере они ассоциируются с последней теплой междуледниковой паузой, наступившей около 125 тысяч лет тому назад. Подобные даты означают либо то, что эти орудия были оставлены еще представителями Homo helmei, либо то, что люди современного типа пришли в Индию гораздо раньше, чем это обычно считается. В любом случае единственные останки скелета, найденные в Индии и относящиеся к периоду между 100 и 200 тысячами лет тому назад, так называемый нармадский череп, явно не соответствует современному типу, как утверждалось ранее[123]. Наиболее древнее неопровержимое свидетельство присутствия людей современного типа в Юго-Восточной Азии, обнаруженное в пещере Ниах на острове Борнео, представляет собой череп современного типа, датируемый около 50 тысяч лет назад и найденный в слое рядом с каменными отщепами, которые, по мнению специалистов, типичны для изделий Среднего палеолита в Индии[124]. Поскольку генетические данные говорят о том, что начало расселению людей современного типа за пределами Африки положила одна-единственная миграция («исход» с Черного континента, это хорошо согласуется с общепринятым мнением, что первые мигранты современного типа из Африки, переправившись через Красное море, продолжали делать орудия образца Среднего палеолита (т.е. аналогичные орудиям неандертальцев). И еще один любопытный факт, касающийся Юго-Восточной Азии и предостерегающий от придания чрезмерного значения технике и материальной культуре: за исключением настоящей «фабрики» по производству орудий из каменных отщепов, найденной в пещере Ниах, и нескольких аналогичных примеров, большинство находок орудий палеолитического типа свидетельствуют о заметном регрессе в технологии. После этого жители Юго-Восточной Азии надолго прекратили производство обработанных заготовок. Другими словами, смело можно говорить о том, что они возвратились к устаревшей технологии создания каменных орудий.
Орудия типа Верхнего палеолита в Европе: почему лезвия так сильно отличались друг от друга?
Честь изобретения новой, следующей за Средним палеолитом, технологии производства орудий — типа Верхнего палеолита — ученые обычно приписывают кроманьонцам. Но если говорить о каменных орудиях, в чем конкретно выражался технический прогресс, достигнутый в Европе людьми эпохи Верхнего палеолита? В целом тут сыграли свою роль различные факторы, но важнейшим признаком инноваций явилось производство и использование ножей. Я, как, впрочем, и большинство простых смертных, считал и считаю весьма сухими и невразумительными эзотерические технические термины, используемые археологами для описания орудий из дробленого камня. И вот я решил обратиться к специалисту в области палеолита, профессору Дереку Роу, директору центра исследований четвертичного периода Оксфордского университета, живущему всего за несколько улиц от моего дома. После чашечки превосходного кофе хозяин, человек весьма любезный и эрудированный, показал мне и даже позволил подержать в руках несколько подлинных отщепов и лезвий. Грубо говоря, в процессе производства лезвия откалывались от особой заготовки, словно щепки или сколы. После отслоения очередного длинного отщепа в руках мастера оставалась призмообразная заготовка, от которой можно было отслоить еще много лезвий (см. рис. 2.1). Само лезвие было уже не округлой или треугольной формы, как прежние сколы, а имело вид длинного каменного отщепа с параллельными, слегка изогнутыми кромками, который можно было довести до нужной формы и получить широкий диапазон орудий: ножи, шила, наконечники и скребки. Потенциальные преимущества, обусловленные различиями в обработке заготовки, оказались поистине огромными. Во-первых, из одной заготовки можно было сделать много орудий, а не одно-единственное. Это обеспечивало невероятную экономию затрат труда, и к тому же, если источников подходящего камня поблизости не было, обработка и изготовление заготовки позволяла экономить силы при транспортировке, ибо глыбы дробились на удобные блоки. Во-вторых, из одной и той же заготовки можно было изготовить самые разные орудия. Другими словами, изобретатели плоских лезвий совершили настоящую техническую революцию, развитие которой пошло быстрыми темпами. Полюбовавшись древними орудиями, я спросил профессора Роу, что, по его мнению, явилось более значимым концептуальным прорывом: дальнейшая доработка и использование заготовок, из которых впоследствии делались различные орудия, имевшие место в Среднем палеолите, или же изобретение каменных пластин-лезвий в Верхнем палеолите? Ответ специалиста был однозначным: более значимым изобретением явилось раннее новшество эпохи Среднего палеолита, поскольку использование готовых заготовок представляло собой многоступенчатый процесс, который требовал от его автора мысленно представлять себе все его этапы. Любая ошибка при обработке — и весь труд приходилось начинать сначала. В то же время изготовление пластин-лезвий вместо отщепов, хотя оно и открывало новые, невиданные возможности, представляло, по сути, всего лишь новыйзавершающий этап уже отлаженной технологии. Другими словами, древние люди, начавшие делать заготовки в эпоху Среднего палеолита, совершили куда более значительный технический прорыв, чем люди современного типа, чьи мастера начали производство пластин-лезвий в Верхнем палеолите, то есть много тысячелетий спустя[125].
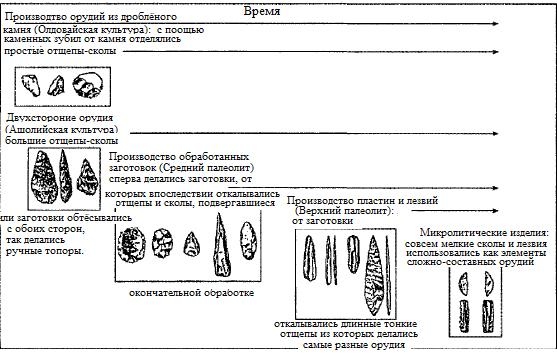 Рис 2.1
Роберт Фоули развивает эту точку зрения, утверждая, что возникновение технологии создания заготовок в Среднем палеолите знаменовало собой появление Homo helmei и эти орудия можно считать даже более надежными маркерами расселения человека современного типа, чем пластины-лезвия эпохи Верхнего палеолита[126].
Чтобы объяснить для себя «концептуальный» парадокс технической революции, связанной с появлением лезвий, я попытался найти в современной жизни пример какого-нибудь совсем простого технического новшества, которое повлекло бы за собой лавинообразное появление всевозможных полезных вещей, и мне сразу же вспомнились застежки-«липучки». При всем уважении к находчивости (и прозорливости) их изобретателя нельзя не признать, что сам принцип изобретения уже давно использовался в семенах репейника, которые, вцепившись в овечью шерсть, отправляются в дальние путешествия. И хотя изобретательская оригинальность застежки-«липучки» вызывает большие сомнения, многообразие возможностей ее применения оказало поистине революционное влияние на многие аспекты нашей повседневной жизни.
Рис 2.1
Роберт Фоули развивает эту точку зрения, утверждая, что возникновение технологии создания заготовок в Среднем палеолите знаменовало собой появление Homo helmei и эти орудия можно считать даже более надежными маркерами расселения человека современного типа, чем пластины-лезвия эпохи Верхнего палеолита[126].
Чтобы объяснить для себя «концептуальный» парадокс технической революции, связанной с появлением лезвий, я попытался найти в современной жизни пример какого-нибудь совсем простого технического новшества, которое повлекло бы за собой лавинообразное появление всевозможных полезных вещей, и мне сразу же вспомнились застежки-«липучки». При всем уважении к находчивости (и прозорливости) их изобретателя нельзя не признать, что сам принцип изобретения уже давно использовался в семенах репейника, которые, вцепившись в овечью шерсть, отправляются в дальние путешествия. И хотя изобретательская оригинальность застежки-«липучки» вызывает большие сомнения, многообразие возможностей ее применения оказало поистине революционное влияние на многие аспекты нашей повседневной жизни.
Неандертальцы отстали от времени. Могли ли они догнать его?
Итак, около 50 тысяч лет тому назад люди современного типа с точки зрения применения каменных орудий находились на том же этапе, что и неандертальцы. Такова одна сторона аргумента. А как насчет эпохи от 28 до 40 тысяч лет назад, когда неандертальцы, как известно, вступали в контакт с людьми современного типа? Мы можем проверить достоверность противоположных гипотез, предполагающих одинаковое и неодинаковое развитие мозга, рассмотрев вопрос о том, что могло бы произойти при общении обеих групп друг с другом. Если неандертальцы, обладавшие более крупным объемом мозга и бывшие ближайшими родичами человека на древе эволюции, не сумели сами выдвинуть продуктивную идею, они вполне могли позаимствовать новые знания у пришельцев-мигрантов. Если же неандертальцы действительно отличались крайней тупостью и не обратили внимания на новые навыки, значит, они так ничему и не научились. В таком случае они сумели усвоить, а в некоторых местах даже развить свои собственные варианты технологии производства орудий Верхнего палеолита. Но такое усвоение, вполне понятно, не могло протекать быстро. Первое, что необходимо отметить, размышляя о возможностях обмена техническими навыками между неандертальцами и людьми современного типа, — это то, что хотя неандертальцы и люди мирно уживались друг с другом в Европе в период между 5 и 12 тысячами лет назад (а в некоторых местах — между 28 и 40 тысячами лет назад), все свидетельства, которыми мы располагаем, говорят о том, что территории их расселения на протяжении этого периода практически не пересекались. Когда люди современного типа, вторгшиеся с востока, быстро заселили Восточную Европу, неандертальцы, компактно обитавшие на западе, постепенно отступали к своим последним оплотам — в Италию, Южную Францию и, наконец, Испанию и Португалию. Недавний тщательный компьютерный анализ стоянок и дат показал, что области совместного обитания неандертальцев и людей современного типа в период после рубежной даты, около 35 тысяч лет тому назад, были крайне ограниченными, и у неандертальцев осталось всего два оплота — в Южной Франции и на юго-западе Испании (см. рис. 2.2). В более поздние времена их осталось уже совсем мало. Нам остается лишь строить догадки о причинах отступления неандертальцев. Было ли оно следствием жестокого конфликта или результатом мирного соперничества? Отсутствие общих территорий расселения на протяжении более 10 тысяч лет свидетельствует о длительном и, скорее всего, не слишком мирном противостоянии.
 Рис. 2.2
Впрочем, возможно, что и более значительные территории совместного проживания не помогли неандертальцам усовершенствовать методы изготовления орудий. В конце концов, на создание и развитие технических новшеств, появившихся в ту эпоху, у людей современного типа ушли многие десятки тысяч лет. Подобно тому, как людям из племени Яли не удалось разгадать секреты появления у европейцев такого невероятного обилия товаров и предметов роскоши, неандертальцы просто не смогли бы в полной мере воспользоваться громадным потенциалом новаторской культуры пришельцев, если они не имели тесных социальных связей с ними. Возможно, неандертальцы вообще редко пользовались возможностью позаимствовать новые технические навыки. Но, несмотря на все эти проблемы, они все же переняли у «современных» кое-какие навыки, главным образом — в районах совместного расселения и в периоды мирного сосуществования (см. рис. 2.2)[127].
Рис. 2.2
Впрочем, возможно, что и более значительные территории совместного проживания не помогли неандертальцам усовершенствовать методы изготовления орудий. В конце концов, на создание и развитие технических новшеств, появившихся в ту эпоху, у людей современного типа ушли многие десятки тысяч лет. Подобно тому, как людям из племени Яли не удалось разгадать секреты появления у европейцев такого невероятного обилия товаров и предметов роскоши, неандертальцы просто не смогли бы в полной мере воспользоваться громадным потенциалом новаторской культуры пришельцев, если они не имели тесных социальных связей с ними. Возможно, неандертальцы вообще редко пользовались возможностью позаимствовать новые технические навыки. Но, несмотря на все эти проблемы, они все же переняли у «современных» кое-какие навыки, главным образом — в районах совместного расселения и в периоды мирного сосуществования (см. рис. 2.2)[127].
Очаги и погребения
Строительство очагов считается одним из признаков, характерных для людей полностью современного типа, однако в России и Португалии были найдены очаги, возраст которых превышает 50 тысяч лет, связанные с применением орудий мустерианской культуры. Это свидетельствует о том, что подобная практика уже существовала в эпоху Среднего палеолита и, следовательно, могла использоваться неандертальцами. Но, пожалуй, одним из самых противоречивых показателей культурного потенциала неандертальцев можно считать погребения. Сложные погребения, в особенности те, в которых находились предметы и орудия, которыми человек пользовался при жизни, — убедительное свидетельство как минимум заботы о посмертии и, не исключено, веры в загробную жизнь. Такую веру можно считать одним из первых проявлений религиозного сознания. Философские аргументы подобного рода делают крайне важным вопрос о том, является ли данная совокупность человеческих останков погребением в подлинном смысле этого слова, а если да, то находились ли в нем традиционные могильные предметы и орудия[128]. Свидетельством настоящих погребений можно считать полные скелеты, да и то не всегда. Полностью сохранившиеся человеческие скелеты, датируемые около 100 тысяч лет тому назад, и, в частности, скелеты неандертальцев, относящиеся к периоду между 40 и 60 тысячами лет назад, вполне могли явиться результатом того, что пещеры, где найдены эти скелеты, были покинуты гиенами и прочими пожирателями падали. Наличие в захоронении остатков цветов, каменных кружков, козлиных рогов и прочих артефактов, присутствие которых объясняется ритуальными или религиозными мотивами, также представляет собой достаточно спорный вопрос[129]. Возможно, наиболее важным свидетельством того, что поначалу это были не более чем культурные инновации локального характера, является тот факт, что наиболее древние погребения встречались только у людей современного типа в Западной Евразии, включая и древнейшие погребения в Кафзехе, Израиль (см. главу 1). Никаких свидетельств существования подобной практики погребений у их современников, обитавших в Африке, не обнаружено. Другими словами, погребения, как и многие другие аспекты технико-культурной революции эпохи Верхнего палеолита, были локальным новшеством, появившимся в Западной Евразии и заимствованным неандертальцами, у которых африканцы современного вида, в свою очередь, переняли обычай предавать погребению мертвецов. Эта последовательность решительно опровергает биологический детерминизм, склонный приписывать те или иные культурные навыки исключительно представителям какого-то одного конкретного вида.
К чему вообще все эти разговоры о неандертальцах?
Мои попытки предстать этаким апологетом неандертальцев, пытающимся сопоставить их культурные навыки с практикой ранних людей современного типа, объясняются отнюдь не желанием доказать, что они, неандертальцы, обладали точно таким же «генетически заданным» интеллектуальным потенциалом. Подобное утверждение невозможно подкрепить имеющимися фактами и свидетельствами. Неандертальцы при всем том, что они были обладателями очень крупного мозга, отличались от людей современного типа в целом ряде других отношений, и поэтому нисколько не удивительно, если их интеллектуальные возможности тоже несколько отличались от наших. Нет, моя цель заключается в том, чтобы доказать, что аргументы о том, будто неандертальцы были существами крайне отсталыми в культурном отношении, поскольку были более медлительными, несообразительными и тупыми, чем пришельцы — люди современного типа, основаны на ложном убеждении, будто пути биологического и культурного развития пролегают совсем близко друг от друга. Во всяком случае, применительно к Европе этот аргумент, что называется, не срабатывает, и его гораздо легче опровергнуть, чем подтвердить материальными свидетельствами. Обитателей Европы эпохи Верхнего палеолита принято превозносить как «революционное человечество», обладавшее такими интеллектуальными преимуществами, как способность к аналитическому мышлению и дар речи. Наиболее частым объяснением этого сценария является концепция биологического прогресса: идея о том, что технико-культурная революция эпохи Верхнего палеолита, имевшая место в Европе, явилась результатом генетически обусловленной мутации, то есть появления гена мышления или речи. Между тем многие из этих радикальных новшеств, которые принесли с собой пришельцы нового вида, были не столь уж новы, и целый ряд инноваций имели вполне конкретную локальную или хронологическую привязку и возникли задолго до появления нашего вида. Эти последние изобретения и обеспечивали пришельцам преимущества в локальном масштабе. Неандертальцы были обезоружены и вытеснены сложной и многообразной культурой, которую принесли с собой пришельцы. В качестве аналогии можно сказать, что никакой антрополог не возьмет на себя смелость утверждать, будто племя неолитической культуры, к которому принадлежал Яли, обладало меньшим биологическим потенциалом, чем мы, сородичи Джареда Дайамонда, живущие в железном веке. Однако совершенно очевидно, что в результате культурной изоляции это племя понятия не имело о множестве технических инноваций, появившихся на Западе за последние 2 тысяч лет, таких, например, как огнестрельное оружие и сталь. Почему мне вздумалось защищать бедных неандертальцев? Мой ответ заключается в том, что сам факт нападок на неумелых и «тупых» неандертальцев, которые хотя и похожи на нас, но все-таки не вполне люди, является весьма симптоматичным для свойственной всем человеческим сообществам потребности изгонять и демонизировать другие группы (см. также главу 5). Я утверждаю, что никем не доказанная «тупость» неандертальцев служит примером точно такого же, бытующего в нашей культуре предубеждения, которое, опираясь на превратно истолкованную географическую логику, обрекает наших анатомически современных африканских предков на роль «недочеловеков». Вполне реальная проблема, обусловленная типичным европоцентрическим мышлением, заключается в том, что современные обитатели Африки являются прямыми потомками тех самых людей, живших в эпоху, предшествующую Верхнему палеолиту, и имеют в своих клетках куда больше общих генов с ними, чем с любой другой расой в мире. Поэтому, унижая их предков, мы унижаем и их самих.
Верхний палеолит в Европе: культурная и биологическая революция?
Клайв Гэмбл — общепризнанный во всем мире специалист по воссозданию основных особенностей поведения людей эпохи палеолита. В своей, пользующейся большим успехом книге «Бредущие сквозь время» он приводит сводный анализ общепринятых взглядов и утверждает, что рубежным, переходным периодом между древними и людьми современного типа следует считать период между 40 и 60 тысячами лет тому назад. Он определяет завершающий этап этого периода, около 40 тысяч лет назад, как время, с которого началось резкое ускорение эволюции. В Европе появились первые произведения искусства, орудия из кости, украшения, погребения, ямы-кладовые для хранения провизии, каменоломни, начал налаживаться обмен товарами, и люди стали создавать постоянные поселения в неблагоприятных климатических зонах. Он развивает свою аргументацию о рубежном периоде, заявляя: «Нет сомнения, что после 35 000 г. [до н.э.] технология производства орудий типа Верхнего палеолита начала быстро распространяться не только в Европе, но и в большей части регионов Старого Света. Исключением из этого правила явилась Австралия...»[130]. Однако подобная «исключительность» Австралии явно не относится к наскальным рисункам, поскольку спустя шесть страниц Гэмбл упоминает о «наскальных рельефах из Каролты в Южной Австралии, которые сегодня можно уверенно датировать 32 тысячами лет тому назад». Такая датировка позволяет считать их почти столь же древними, как и наскальные рисунки в пещере Шове, а это, в свою очередь, дает основание — хотя бы частично — отнести Австралию к регионам, где имело место ускоренное развитие. Другие ученые, и в их числе — чикагский палеонтолог Ричард Клейн, придают этому культурному прорыву еще более важное значение и рассматривают его в качестве свидетельства биологической эпифании человека. В своем известном труде «Развитие человека», написанном в 1989 г., он утверждает:
«Нетрудно доказать, что Верхний палеолит явился своего рода сигналом для наиболее фундаментальных изменений в поведении человека из всех, свидетельства которых удалось обнаружить археологам... Близкая корреляция между артефактами Верхнего палеолита и человеческими останками той эпохи показывает, что именно появление современного в физическом отношении типа человека сделало возможным наступление эпохи Верхнего палеолита (и всего последующего развития культуры). Возникает вопрос: существовала ли сколько-нибудь заметная связь между эволюцией современного человека и развитием новых поведенческих навыков, знаменовавшая наступление Верхнего палеолита?»Затем, привлекая внимание к орудиям людей современного типа той эпохи, характерным для Среднего палеолита, он приходит к выводу:
«Таким образом, анатомические и поведенческие признаки современного типа, по всей видимости, появились в Европе одновременно, тогда как на Ближнем Востоке и в Африке анатомическая «современность» далеко опережала развитие поведенческих особенностей современного типа, по крайней мере — насколько позволяют судить археологические свидетельства. Объяснить подобное наблюдение довольно трудно. Возможно... самые ранние анатомически современные люди в Африке и на Ближнем Востоке не были настолько «современными», как о том свидетельствуют их скелеты. Вполне вероятно, что с точки зрения высшей нервной деятельности они не обладали потенциалом современных людей для создания развитой культуры. Подобные способности могли появиться у них где-то около 40—50 тысяч лет назад, когда, как предполагается, началось быстрое расселение людей современного типа по всему миру»[131].В подобных биологических построениях мы имеем дело с обычной моделью исхода из Африки, согласно которой хронологический и генетический рубеж человека современного типа (в плане поведенческих навыков и высшей нервной деятельности) имел место не ранее 50 тысяч лет тому назад; причем произошел такой перелом в Западной Евразии после исхода из Африки. Эта модель возникла в 1989 г., в то время, когда считалось, что Австралия была заселена всего 40 тысяч лет тому назад. Другими словами, Клейн выступил в поддержку гипотезы о том, что Австралия, а следовательно, и Азия, были заселены людьми анатомически современного типа только после начала эпохи Верхнего палеолита в Европе. Это позволило ему утверждать, что уроженцы Европы, обладатели «новых возможностей высшей нервной деятельности», впоследствии начали активную колонизацию всех континентов за исключением Африки. Второе издание своей книги Клейн опубликовал в 1999 г. К тому времени он уже допускал (о чем говорится на самых последних страницах его книги) возможность более раннего заселения Австралии (и, естественно, Азии), имевшего место около 60 тысяч лет тому назад, и применение гарпунов для промысла рыбы в эпоху между 90 и 155 тысячами лет тому назад. В своих выводах Клейн возвращается к аргументам в пользу эволюционной (т.е. генетически обусловленной) революции в области высшей нервной деятельности, происшедшей в Европе 40—60 тысяч лет тому назад: «На мой взгляд, это показывает, что именно к этому времени [около 50 тысяч лет тому назад] могли сложиться интеллектуальные предпосылки для создания развитой культуры»[132]. Но прежде чем обратиться к рассмотрению доказательств, нетрудно заметить, что подобного рода аргументы предполагают биологически детерминистский подход к культурной эволюции. Они предполагают, что любой фактор прогресса в культуре обусловливается или «включается» в результате генетических изменений. Как я уже говорил в прологе, культура человека (и других приматов) первоначально возникает в зачаточном виде, а затем развивается и приумножается от поколения к поколению. Любой новый прорыв или изобретение никоим образом не связаны с появлением «новых» генов. Наоборот, сперва происходит появление новых поведенческих навыков, а уже затем происходят генетические модификации, как бы «закрепляющие» эти новшества. Другими словами, изменения в культуре предшествуют возникновению физических изменений: именно так, а не наоборот. Более того, в развитии культуры имеют место легко прогнозируемые географические различия. Если появление какого-то изобретения в одном регионе влечет за собой другие локальные инновации, ускоренный темп появления новшеств позволяет этому региону раньше других взять старт в гонке прогресса. Поэтому мы вправе ожидать существенных различий в темпах развития между разными регионами, хотя все они происходят в рамках одного и того же вида человека.
Мог ли европейский ген мудрости распространиться в другие регионы?
В аргументации Клейна существует целый ряд неизбежных логических допущений, сводящихся к тому, что люди полностью «интеллектуально современного» типа появились лишь около 40—50 тысяч лет тому назад. Во-первых, в этой аргументации как бы подразумевается, что ранние африканцы современного типа были гораздо мельче современного человека, то есть, другими словами, они не обладали потенциалом высшей нервной деятельности, необходимым для развития поведенческих навыков, присущих людям современного типа. Этот более чем странный вывод неизменно относился и к людям современного типа, оставшимся в Африке, и к первым мигрантам, переселившимся в Азию и Австралию, поскольку сегодня возобладало мнение, что такая колонизация могла произойти незадолго до рубежной даты — 50 тысяч лет тому назад (наиболее ранний срок установления культуры Верхнего палеолита в Восточном Средиземноморье). О чем же говорят подобные гипотетические выводы? Прежде всего они означают, что прямые предки современных обитателей Африки, жившие в период между 50 и 130 тысячами лет тому назад, с биологической точки зрения были не способны развить и использовать технические и поведенческие навыки эпохи Верхнего палеолита. Они не могли заниматься живописью и резьбой, торговать, организовывать сообщества и т.д. Некоторые утверждают даже, будто они не обладали даром речи, а если и обладали, то их речь была «крайне примитивной». При таких вопиющих недостатках они были бы совершенно не в состоянии, предоставься им такая возможность, управлять автомобилем или пилотировать самолет, сочинять и исполнять духовную и классическую музыку, рок и джаз, и, наконец, не смогли бы стать врачами, финансистами или генетиками. Генетические деревья митохондриевой ДНК и Y-хромосомы показывают, что современные африканцы являются потомками многих генетических линий, возникших гораздо раньше 50 тысяч лет тому назад и к тому же отнюдь не за пределами Африки. Почему же тогда современные африканцы способны с успехом выполнять все эти функции, которые, как полагают некоторые, были генетически недоступны для их предков? Возникает и другая логическая проблема. Если европейцы оказались первыми биологически современными людьми и представляли собой изолированную общность пришельцев, появившихся в сравнительно позднее время, как обстояли дела у жителей остальных регионов земного шара? Каким образом им удалось догнать европейцев? Все живущие на Земле люди являются абсолютно «анатомически современными», и мы можем проследить наши генетические родословные вплоть до совсем небольшой группы предков, которая начала делиться на ветви еще в Африке около 190 тысяч лет тому назад. Ни в какой момент истории после этого общая численность человечества не опускалась ниже 1000 человек[133], и поэтому нетрудно понять, что увеличение численности и образование ветвей этой группы неизбежно должно было привести — и действительно привело — к образованию все новых и новых этнических групп. Итак, древнейшее ядро современного человечества достаточно рано начало дробиться, делиться на ветви и расселяться по земному шару, причем некоторые из этих ветвей никогда более не встречались вплоть до недавнего времени. Этот эффект «необратимого» разрыва никогда не проявлялся с такой очевидностью, как в случае, когда одна-единственная группа мигрантов переправилась через Красное море и двинулась в Индию, а затем дальше — в Австралию. Если же, как полагают многие последователи эволюционизма, в Европе произошло некое позднейшее генетическое изменение, сделавшее нас поведенчески современными людьми, в отличие от поведенчески «архаичных» народов, такая мутация (или мутации) должна была впервые произойти в организме некоторых европейцев во вполне определенное время, значительно позже, чем 45 тысяч лет назад, и, разумеется, это должно было иметь место за пределами Африки. Эта новая мутация должна была передаться всем потомкам, носителям мутированного гена, но ее не должны были унаследовать двоюродные братья и их потомство. Единственное исключение из этого правила могло бы возникнуть, если бы мутированный ген был впоследствии передан в результате близкородственного брака. Но шансы, что такой близкородственный брак мог стать реальностью, были исчезающе малы, ибо группы потомков навсегда расстались и расселились по всему свету. Если бы дело обстояло именно так, то такие навыки, как «дар живописи» или «дар речи», могли бы унаследовать только те, кто были прямыми потомками людей, впервые развивших эти навыки. Таким образом, если набор мутаций, характерных для «поведенчески полностью современных» людей, первоначально сложился в Европе около 40—50 тысяч лет тому назад, получается, что все остальные жители нашей планеты — австралийцы, азиаты и африканцы — просто-напросто не обладали бы способностью заниматься рисованием и резьбой, изготавливать ножи или делать ставки на конных бегах Ясно, что это — полный абсурд, ибо они отлично могут делать подобные вещи. Если следовать логике этого аргумента, единственный способ, посредством которого первоначальные поселения жителей Азии, Африки и Австралии могли догнать европейцев, начиная с эпохи Верхнего палеолита ушедших далеко вперед по пути культурного развития, должен был бы заключаться в том, что эти народы вынуждены были получить своего рода «инъекцию» новых «культуроносных генов». Между тем единственным биологическим путем осуществления генных инъекций или вливаний являются миграции и браки между представителями разных рас. Но для подобной акции недостаточно было бы иметь несколько кузин-иностранок. Чтобы радикальным образом изменить потенциальные способности потомства, гипотетические старые «культуроносные гены» пришлось бы полностью заменить на «новые». Любопытно, что массовый заброс генетического материала — это именно тот аргумент, который обычно используют сторонники гипотезы мультирегионализма, стремясь объяснить, как и почему на основе локальных подвидов единого вида Homo erectus в различных регионах планеты сформировались люди современного типа, похожие друг на друга куда больше, чем представители локальных типов Homo erectus. Основная проблема, связанная с теорией уравнивания генных потоков, заключается в том. что географические характеристики генетических деревьев мтДНК и Y-хромосомы не несут никаких свидетельств подобного крупномасштабного межрегионального смешения. А теперь рассмотрим пример из области культуры. Для того чтобы австралийцы могли создать свои знаменитые наскальные рисунки около 32 тысяч лет тому назад (что они, кстати сказать, и сделали), то есть практически одновременно с появлением аналогичных наскальных рисунков в Европе, потребовался бы мгновенный и массовый «завоз» генетического материала из Европы по всему земному шару, чтобы позволить жителям удаленных регионов достичь столь высокого уровня. Это очень спорная идея, в несостоятельности которой легко убедиться. Взглянув на генетическое древо, нетрудно понять, что, хотя современные австралийцы имеют с европейцами двух общих неафриканских предков по линиям М и N, живших около 70 тысяч лет тому назад, они тем не менее сохранили свои собственные характерные типы М и N. У нас нет никаких свидетельств того, что они, австралийцы, являются потомками европейцев. Нет у нас и никаких сведений о массовом «завозе» генов из Европы в Австралию в эпоху палеолита[134].
Общее правило — отсутствие генетического смешения после исхода из Африки
На самом деле генетические маркеры по мужской и женской линиям, Y-хромосома и мтДНК, демонстрируют нам картину, прямо противоположную массовому «завозу» генов. Одно из наиболее существенных сведений, которые сообщают Y-хромосомы и мтДНК, заключается в том, что после первоначального расселения предков человечества после исхода из Африки были заселены практически все регионы Старого Света и восточного полушария, и вплоть до последнего великого оледенения, наступившего около 20 тысяч лет тому назад, между изолированными регионами не существовало практически никакого обмена генами. Генетические маркеры обеих систем со всей очевидностью показывают четкие разграничения между континентами и регионами. Культурная диффузия (особый тип распространения культуры, не требующий массовых перемещений крупных этнических групп) в эпоху палеолита была скорее способом переноса культуры в отдаленные регионы, не требующим значительного «завоза» генетического материала. Но возможно ли допустить, что австралийские наскальные рисунки, созданные около 32 тысяч лет тому назад, восходят к европейским рисункам, возникшим практически в то же самое время? Наиболее простой ответ на этот вопрос, позволяющий решить этот и аналогичные парадоксы, заключается в том, что африканские предки всех будущих неафриканских народов и рас на момент исхода из Африки уже обладали речью, способностью рисовать, петь и танцевать — и были людьми современного типа в полном смысле этого слова! Таким образом, существует слишком много биологических и просто логических аргументов против некоего генетического эволюционного взрыва, якобы приведшего к появлению около 40 тысяч лет тому назад на Леванте, в Европе и любом другом регионе за пределами Африки человека полностью современного типа. Это требует от нас изучения прямых археологических свидетельств и антропологических аргументов в пользу более простой модели, суть которой — в том, что первые анатомически современные африканцы уже были полностью современными с точки зрения их интеллектуального потенциала.
Свидетельства
Недавно две известные группы антропологов и археологов провели исследование орудий, технических навыков и образа жизни наших древнейших предков, чтобы оценить их потенциальные возможности. Принципы исследований и их хронологические рамки были весьма и весьма различными, но их выводы, на мой взгляд, оказались практически идентичными. Знакомясь с их исследованиями, мы должны постоянно учитывать археологические доказательства изменений в техническом арсенале между различными этнически культурными группами, имея в виду уже знакомый нам вопрос Яли. Существующая система письменности была изобретена около 4000 лет назад в Западной Евразии, но никому и в голову не придет говорить о появлении «гена письменности». То же самое можно сказать и о прочих выдающихся изобретениях нашего времени — радио, телевидении, компьютерах, компьютерных языках, космических летательных аппаратах и т.д., и т.п. Другими словами, мы не можем интерпретировать все более и более высокие уровни сложности новейших технических изобретений человека как вехи его биологического развития. Кроме того, многие свидетельства технократической культуры и культуры, оперирующей символами, например, деревянной резьбы и живописи на дереве, весьма недолговечны, и поэтому до нас дошло несравненно меньше свидетельств творческой активности представителей культуры, использующей дерево. Это означает, что без адекватного контекста мы не можем признать, что технические инновации — это свидетельства нового этапа биологической эволюции. Располагая достоверными знаниями о том, как те или иные культурные новшества распространялись в более близкую к нам историческую эпоху, мы вправе утверждать, что вехами на пути достаточно медленного культурного развития являются локальные «революции», затем происходит их диффузия и взаимопроникновение, и, наконец, наступает общее ускорение развития. При изучении истоков революционного переворота в эпоху Верхнего палеолита, пожалуй, особенно важно определить географический локус, где сложилась культура, непосредственно предшествовавшая ему, — в Северной Африке или Южной Азии?
Каменные следы расселения человека по всему миру
Разумеется, нам лучше всего начать с изучения наиболее многочисленных и долговечных следов далекого прошлого: каменных орудий. Кембриджские ученые — антрополог Роберт Фоули и палеонтолог Марта Лар — провели независимое исследование каменных свидетельств и костных останков практически из всех регионов мира, чтобы установить, с каким именно типом человека, древним или современным, ассоциируются различные технологии производства каменных орудий. Главным выводом этого исследования явилось то, что расселение по всему миру человека современного типа наиболее достоверно устанавливается по возникновению так называемой технологии 3 типа (рис. 2.3) — своего рода технического порога, который преодолели в Африке наши предки, представители вида Homo helmei, около 300 тысяч лет тому назад. Первоначально описанная как техника использования обработанной каменной заготовки, из которой делались отщепы-сколы, технология типа 3 более известна под другим названием — технология Среднего палеолита. Другая характерная черта технологии типа 3 — изменение методов обработки, переход от больших, тяжелых топоров к более мелким каменным орудиям, включая острые отщепы, которые могли иметь черенок[135]. Таким образом, возникновение технологии 3 типа совпадает с появлением вида Homo helmei, имевшего более крупный мозг, — того самого, который, по мнению Фоули и Лар, был общим предком и нас, людей современного типа, и неандертальцев. Тип 3 стал основной технологией, которую использовали все представители этого семейства человека, включая людей современного типа, примерно до рубежной даты — около 50 тысяч лет тому назад. Использование специальных заготовок-сердечников для получения отщепов в концептуальном отношении было наиболее сложной инновацией, появившейся в Нижнем палеолите. Если эти люди были достаточно сообразительными, чтобы обрабатывать заготовки-сердечники, они наверняка догадались делать из них плоские лезвия. Производство пластин-лезвий из обработанных призматических заготовок — это вторая этапная веха на пути технического прогресса.
 Рис.2.3 Географическое распространение типов технологий и соответствующих видов человека
Рис.2.3 Географическое распространение типов технологий и соответствующих видов человека
Однако она куда менее полезна как индикатор расселения людей современного типа, поскольку представляет собой позднейшее и достаточно локальное изобретение, появившееся в Европе и странах Средиземноморья (а впоследствии затронувшее Африку и Азию, но оставшееся неизвестным в Австралии). Несомненно, оно является не слишком надежным маркером расселения людей полностью современного типа за пределами Африки, поскольку имеются свидетельства производства пластин-лезвий другими видами, в том числе людьми не вполне современного типа (например, шательперронийская техника), а также техники ранних африканцев современного типа (такие, как Ховисонс Поорт в Южной Африке, 60—90 тысяч лет тому назад), находимые на стоянках, где встречаются образцы технологий Среднего Каменного века. Надо сказать, что это непродолжительное появление «совершенных» лезвий вскоре вновь уступило место типичным образцам технологии типа 3. Некоторые археологи предполагают, что такие пластины-лезвия могли быть изобретены несколько раз еще до начала Верхнего палеолита, а затем забыты[136]. Пожалуй, двумя наиболее существенными практическими преимуществами этой техники изготовления пластин в Верхнем палеолите были многообразие видов получаемых орудий и экономия сырья. Дуэт американских антропологов и археологов, Салли Мак-Брирти и Алисон Брукс, развили далее аргументацию о технических навыках древних африканцев эпохи Среднего Каменного века. В своей содержательной книге, посвященной возникновению поведенческих навыков человека современного типа и озаглавленной «Революция, которой не было»[137], они бросают вызов общепринятым представлениям о «технической революции», якобы происшедшей примерно 40—50 тысяч лет тому назад. Развивая близкие идеи, но используя подход, резко контрастирующий с методикой Фоули и Лар, которые рассматривали технологию 3 типа (эпохи Среднего палеолита) как отличительный признак поведения человека современного типа, Мак-Брирти и Алисон Брукс рассматривают эволюцию культурных навыков «современных» с точки зрения технологии Среднего Каменного века в Африке. Последняя была почти современницей технологии Среднего палеолита в Европе вплоть до Позднего Каменного века в Африке, причем какой-то период после 70 тысяч лет тому назад между этими регионами существовали крайне скудные контакты и взаимообмен. Так, эти исследователи считают Средний Каменный век в Африке, начавшийся с появления Homo helmei около 250—300 тысяч лет назад, более динамичным и творчески успешным с точки зрения эволюции культуры процессом, чем его современник, Средний палеолит в Европе, подчеркивая, что в нем выделялись черты, более характерные для Верхнего палеолита в Европе. По их мнению, на протяжении нескольких веков в Африке происходил процесс постепенного формирования сообществ и накапливания целого комплекса сложных культурных и материально-технических знаний и навыков. Если обратиться к истории Африки, нетрудно заметить, что лишь немногие из этих технических и социально-общественных новаций могут ассоциироваться с этапами биологической эволюции, когда Homo helmei начал активно вытесняться людьми современного типа. Зато несравненно больше фактов можно отнести к чисто культурной эволюции, которая набирала ускоренный темп развития.
Новаторские африканские орудия
Мак-Брирти и Брукс показали, что, в отличие от Европы, где пластины-лезвия явились отличительной чертой появления человека современного типа около 40—50 тысяч лет тому назад, подобные лезвия использовались в Африке еще в Среднем Каменном веке, около 280 тысяч лет тому назад, опередив таким образом появление людей современного типа на добрую четверть миллиона лет[138]. Другим типом каменных орудий, буквально процветавшим в Среднем и Верхнем палеолите, были каменные наконечники. Эти наконечники представляли собой по большей части длинные отщепы, которые обрабатывались по обеим кромкам, чтобы их можно было насадить на древко копья. История их использования в Африке насчитывает более 250 тысяч лет, что гораздо дольше, чем в Европе, и, как подчеркивают те же Мак-Брирти и Брукс, демонстрирует куда более широкий и разнообразный спектр региональных вариантов (см. рис. 2.4). Один из специализированных типов каменных орудий, первоначально появившийся в Африке, впоследствии был «экспортирован» в Европу в раннюю эпоху ее развития. Тип этот — микролиты. Эти небольшие (25 мм и меньше), но тщательно обработанные орудия, затупленные с одной кромки как перочинный нож, делались из небольших пластин-лезвий или их сегментов. Круг их применения был весьма широк; в частности, они могли использоваться в составных орудиях (ножи, серпы), а также в качестве наконечников для копий и стрел. Отдельный поврежденный микролит было легко извлечь и заменить новым. Микролиты, считающиеся технологией 5 типа — высшим уровнем сложности развития каменных орудий, — знаменовали собой этапную веху: начало Позднего Каменного века в Африке. Первые образцы такого типа были найдены в Мумба Рок Шелтер, Танзания; их возраст — около 70 тысяч лет. В Европе же они появились гораздо позже, в основном — после последнего оледенения, около 8000 лет тому назад. Наиболее ранние находки микролитов, сделанные не в Европе, а, что весьма примечательно, на Шри-Ланке (о. Цейлон), можно датировать временем около 30 тысяч лет назад. Это указывает на непосредственное влияние и «экспорт» идей из Африки по южному маршруту (см. главу 4)[139]. Использование некаменных материалов, таких, как кость и рог, для производства оружия и всевозможных орудий считается еще одним «изобретением» европейцев, сделанным около 30—40 тысяч лет тому назад. Между тем Мак-Брирти и Брукс приводят достоверные свидетельства применения орудий из кости в Африке еще 100 тысяч лет тому назад. Более того, некоторые из наиболее ранних орудий из кости имели острие с зазубринами, напоминающими наконечники гарпунов (см. рис. 2.4).
 Рис. 2.4
Другим сенсационным изобретением, появившимся около 280 тысяч лет тому назад, во времена наших далеких прадедов, Homo helmei, стало использование камней для растирания. Такие камни выполняли две совершенно различные функции: одни из них применялись для растирания пищи, другие — для растирания минеральных пигментов, например, желтой и красной охры, для приготовления красок. Последний вид применения куда более важен для истории нашего интеллектуального развития[140].
Рис. 2.4
Другим сенсационным изобретением, появившимся около 280 тысяч лет тому назад, во времена наших далеких прадедов, Homo helmei, стало использование камней для растирания. Такие камни выполняли две совершенно различные функции: одни из них применялись для растирания пищи, другие — для растирания минеральных пигментов, например, желтой и красной охры, для приготовления красок. Последний вид применения куда более важен для истории нашего интеллектуального развития[140].
Древнеафриканские художники
Использование пигментов считается еще одной этапной вехой истории Верхнего палеолита в Европе[141], так что их появление около 280 тысяч лет назад в Африке со всей очевидностью показывает, что действительно прекрасные наскальные рисунки в пещерах Шове, считавшиеся первым реальным доказательством абстрактно-символического мышления древнего человека, ослепили нас и не позволили заметить такую же перемену в сознании наших куда более отдаленных предков. Пигменты использовались в Африке еще 100 тысяч лет тому назад, причем настолько широко, что добыча минеральных красителей, в частности гематита (красного железняка), можно сказать, велась в промышленных масштабах. Между тем первые подобия карьеров для добычи минералов появились в Европе всего лишь 40 тысяч лет тому назад. Только в одном руднике в Африке с поверхности скальных стенок было добыто не менее 1200 тонн породы, содержащей пигменты-красители[142]. Некоторые антропологи и археологи были поражены, убедившись, что использование пигментов носило систематический характер, поскольку они считали этот факт, наряду с практикой погребений, одним из ранних свидетельств способности к символическому мышлению. Минеральные пигменты применялись в качестве красок для создания рисунков на стенах пещер и других объектах, боевой и ритуальной раскраски тела, при погребении усопших, а также лечении ран. Неандертальцы также использовали пигменты, хотя датировка находок, подтверждающих это, скорее говорит о том, что они заимствовали эту практику посредством культурной диффузии у древнейших людей современного типа. К сожалению, мы уже никогда не узнаем, в какой мере густой волосяной покров на теле неандертальцев служил им защитой от холода, но он, естественно, затруднял нанесение боевой раскраски и снижал ее практическое значение. Люди эпохи Верхнего палеолита использовали пигменты как минерального, так и растительного происхождения, но свидетельства применения растительных пигментов в Среднем Каменном веке в Африке крайне скудны, а со временем исчезнут окончательно. Доказательства существования памятников изобразительной живописи также весьма ограничены из-за их глубокой древности. Это особенно относится к Африке, в которой крайне мало известняковых карстовых пещер типа тех, в которых так хорошо сохранились знаменитые рисунки в Ласко и Шове. Любопытная особенность рисунков пещеры Шове, этогодревнейшего памятника живописи в Европе, заключается в том, что они представляют наиболее совершенные образцы искусства эпохи палеолита. Однако, обладая невероятной экспрессивностью и живописной выразительностью и используя для достижения особо драматического эффекта элементы рельефа самих стенок, наскальные рисунки Шове отнюдь не выглядят первыми робкими проблесками абстрактно-символического мышления. То, что мы видим в них, — это вполне зрелый стиль, вершина целой эпохи в искусстве, по сравнению с которой позднейшие пещерные рисунки кажутся явным шагом назад[143]. Однако самые древние образцы изобразительного искусства были обнаружены все-таки не в Европе, а в одной из пещер в Намибии (Южная Африка), и датируются они, по контексту, Средним Каменным веком, точнее — от 40 до 60 тысяч лет тому назад, то есть они значительно старше европейских рисунков (см. Фото 11). В различных районах Южной Африки были найдены гематитовые «карандаши», датируемые эпохой свыше 100 тысяч лет назад.

Фото 11. Этот кот (или, может быть, бык), найденный на скальной стоянке «Аполло-П» в Намибии, — возможно, самый ранний в мире образец репрезентативного искусства. Его возраст — 40—60 тыс. лет.
Археологические свидетельства такого рода опять-таки противоречат представлениям о Европе как о месте, где произошла «революция символического мышления человечества». Как отмечают Мак-Брирти и Брукс, в Африке вполне могут существовать памятники живописи еще более глубокой древности, но прямые свидетельства их существования либо безвозвратно утрачены, либо их еще предстоит найти, когда поисковые работы в Африке будут проводиться с такой же интенсивностью, как в Европе[144]. Узнаваемые изображения людей, животных и разных предметов в любом случае не являются первыми свидетельствами символического мышления. Правильные линии, поперечные царапины и бороздки на кусках породы или блоках минеральных пигментов, по всей вероятности, имели некое символическое значение. Такие артефакты часто находят на стоянках в Африке; их возраст — более 100 тысяч лет. Но, пожалуй, самые ранние свидетельства таких примитивных «рисунков» на камнях найдены в песчаных пещерах в Индии. Они относятся к рубежу Нижнего и Среднего палеолита, то есть их возраст — от 150 до 300 тысяч лет. Австралийский археолог Роберт Беднарик, напротив, утверждает, что ряды волнистых линий и ложбинок (мелких выемок на поверхности камня) в пещерах Бхимбетка неподалеку от Бхопала являются самыми ранними памятниками символического искусства во всем мире[145].
Древнеафриканские украшения
Индивидуальные украшения, например с использованием бусин и подвесок, являются, по мнению археологов, новым усложнением ритуальных и символических практик в эпоху Верхнего палеолита в Европе. Найдены свидетельства ношения нательных украшений людьми полностью современного типа в Европе примерно 40 тысяч лет назад, но особенно широкое распространение они получили в граветтийских культурах, хотя есть факты, подверждающие их существование около 40 тысяч лет тому назад. Один из самых впечатляющих примеров таких украшений — украшения, найденные в древнем захоронении в 200 км к востоку от Москвы, в Сунгире. Там около 24 тысяч лет тому назад в непосредственной близости от приближающихся ледниковых плит очередного ледникового максимума были сделаны два захоронения, в которых находились останки трех человек: старика и двух детей. Все трое были облачены в одежды, богато украшенные тысячами костяных бусин (см. Фото 12). Пышность наряда покойных довершали браслеты из кости, жезлы, фигурки животных и всевозможные подвески[146].
 Фото 12.
Одно из Сунгирских захоронений: останки пожилого человека, найденные неподалеку от Москвы. (Их возраст — 24 тыс. лет.) Голову украшают бусы из лисьих клыков, на руках — 20 костяных браслетов для запястий и предплечий и многие тысячи костяных бусин. Поразительный пример богатства украшений.
Фото 12.
Одно из Сунгирских захоронений: останки пожилого человека, найденные неподалеку от Москвы. (Их возраст — 24 тыс. лет.) Голову украшают бусы из лисьих клыков, на руках — 20 костяных браслетов для запястий и предплечий и многие тысячи костяных бусин. Поразительный пример богатства украшений.
Мак-Брирти и Брукс установили, что индивидуальные украшения появились в Африке на несколько десятков тысяч лет раньше, чем в Европе. В качестве примера исследователи приводят подвеску из раковины, найденную в Южной Африке в захоронении ребенка и датируемую около 105 тысяч лет тому назад, а также ряд других украшений и подвесок с просверленными отверстиями. Их возраст — более 130 тысяч лет, и они относятся к африканскому Среднему Каменному веку. Бусины, сделанные из скорлупы яиц страусов, — характерная черта орнаментики африканского Позднего Каменного века; их можно отнести к эпохе около 60 тысяч лет тому назад[147]. Как показывает рис. 2.5, где суммированы свидетельства достаточно постепенного накапливания элементов культуры людей современного типа за последние 300 тысяч лет, использование пигментов, нанесение символических меток на скалы и новая техника изготовления лезвий и наконечников вообще не являлись изобретением человека современного типа. Эти навыки и символические действия начали формироваться еще около 250 тысяч лет тому назад — на достаточно раннем этапе развития наших архаических предков. Более хрупкие артефакты — свидетельства сложной и развитой культуры, например бусины и наскальные рисунки, нередко находимые в Африке, датируются эпохой на несколько десятков тысяч лет старше первых аналогичных памятников в Европе. Добыча минералов, костяные орудия и гарпуны из кости появились в Африке около 100 тысяч лет тому назад. Что же касается позднейших навыков, то они в хронологическом отношении ассоциируются с людьми современного типа.
 Рис 2.5
Рис 2.5
Африканские рыбаки
Эти новшества, созданные «современными» людьми, затрагивали все стороны человеческой жизни. Рассмотрев вопрос об их занятиях, мы видим еще более широкий спектр поведенческих навыков, начавший складываться в еще более раннюю эпоху, между 110 и 140 тысячами лет тому назад, — время, к которому относятся наиболее древние ископаемые свидетельства о появлении в Африке людей современного типа. Поскольку мы вовсе не хотели бы попасть в ловушку и автоматически приписывать эти новые навыки неким уникальным генетическим изменениям, есть смысл проанализировать новые черты поведения, которые сразу же ставят нас на целый уровень выше нашего предка, Homo helmei, обладателя крупного мозга. Переход на новый, более разнообразный рацион питания и включение в него рыбы и моллюсков, вероятно, имел место около 150 тысяч лет тому назад. Не будем забывать и том, что неандертальцам, жившим около 60 тысяч лет назад на Средиземноморском побережье, тоже была знакома практика собирательства на прибрежной полосе[148]. Бесспорно, важнейшим поведенческим навыком всякого животного является способность добывать пищу. За последние 2 млн. лет все больше и больше африканских земель превращались в пустыни, так что добывать пищу становилось все труднее. Чтобы выжить, людям приходилось проявлять больше изобретательности, и они превратились в кочевых охотников и собирателей, бродивших по просторам саванн. Именно так кормились наши далекие предки. Люди архаического типа примерно с начала Среднего Каменного века уже были способны вести успешную охоту на крупных травоядных животных[149]. Период сильного оледенения, продолжавшийся между 130 и 170 тысячами лет назад, уничтожил большинство видов дичи, служивших для человека основными источниками калорий и белков. Поэтому нельзя считать простым совпадением, что начиная примерно со 140 тысяч лет тому назад, в эпоху Среднего Каменного века, в Африке возник новый способ добычи пищи — собирательство на прибрежной полосе. Собиратели охотно ели моллюсков, найденных на берегу и служивших обильным источником белка. Следы собирателей обнаружены и в Южной Африке, в устье реки Класис, но наибольший интерес для нас представляет прибрежная стоянка собирателей в Абдуре, Эритрея, на берегу Красного моря. Дело в том, что именно эти места могли стать отправным пунктом на пути собирателей в далекую Австралию (см. главу 1). Здесь, в Абдуре, на самой высокой точке, на которой находился уровень моря в последнюю междуледниковую паузу, в слоях, образовавшихся около 125 тысяч лет тому назад, рядом с костями крупной африканской дичи найдены остатки раковин морских моллюсков и орудия Среднего Каменного века. Мак-Брирти и Брукс утверждают, что переход от спорадического собирательства к настоящему рыболовству произошел в Африке примерно 110 тысяч тому назад[150]. Мне посчастливилось оказаться в числе тех, кому первооткрыватель этих артефактов, эритрейский геолог Сейфе Берхе, фактически сам показал знаменитый риф Абдур. Отправившись в путь вниз по довольно крутому откосу, мы покинули прохладное орошаемое плато, окружающее столицу Эритреи, Асмару, и вскоре достигли жаркого берега Красного моря, оказавшись в пострадавшем от войны[151] порту Массава, процветавшем в эпоху арабского владычества. Сделав краткую остановку, чтобы запастись водой и мороженым, мы направились на юг и около трех часов ехали по не слишком ровной, пыльной и продутой всеми ветрами дороге, тянущейся вдоль побережья. Участки причудливого вулканического рельефа и мощные горные складки чередовались с засушливыми аллювиальными равнинами. Здесь нам пришлось ненадолго задержаться и поменять колесо: мы напоролись на острый вулканический камень. Был конец сухого сезона, и скудные кустарники, по большей части — акации, едва-едва зеленели. Несколько газелей, шакал, мелькнувший вдалеке заяц, дрофа да орел — таковы были скудные остатки того обилия живности, которое мне доводилось видеть в этих местах перед недавней войной. Зато теперь всюду виднелись стада верблюдов и коз, коров и курдючных пустынных овец. Кроме редких пастухов-кочевников, единственными коренными обитателями этой части побережья, простирающейся до Джибути, были афары[152]. Афары, издревле селившиеся в легких хижинах из ветвей кустарников, полностью зависели от своих стад и каждый день добывали воду для своих внушительных стад из колодцев, выкопанных в пересохших речных руслах. Мы остановились на привал на краю одного из таких высохших рукавов в дельте на ровной, как стол, песчаной поверхности пустыни, возвышающейся на добрые 10 м над берегом моря. Вскоре я с удивлением заметил, что мы расположились на том самом знаменитом рифе, где найдены слои, возраст которых — более 125 тысяч лет. Когда, примерно 120 тысяч лет назад, климат в этих местах начал быстро ухудшаться, уровень моря резко понизился, и риф, покрытый всевозможными кораллами и раковинами, живописно поблескивающими на солнце, оказался над водой, прямо у нас под ногами. Устойчивый тектонический подъем продолжался, в результате чего вся обширная полоса рифов поднялась еще на 5 м, образовав утес, протянувшийся на 10 с лишним километров вдоль побережья. Этот дополнительный выступ и защитил риф от волновой эрозии, имевшей место при следующем подъеме уровня океана, который наступил примерно 5500 лет тому назад. На следующий день после приезда мы спустились по склону утеса в сухую долину, образованную высохшим руслом, и встретили нескольких афаров, поивших свои стада. Чтобы защитить колодец от загрязнения, они доставали воду из него кожаными ведрами и выливали в большой, напоминающий мутный прудик резервуар, устроенный прямо в песке. И пока коровы, козы и овцы теснились у края, с нетерпением ожидая своей очереди напиться из этого импровизированного «корыта», пастухи распевали песню, восхваляющую воду. Сразу же после этой идиллической сцены Сейфе Берхе показал мне раковины, возраст которых — более 125 тысяч лет. Остатки устричной трапезы выглядели так, словно они были оставлены не далее как вчера. Нам сразу же бросился в глаза обсидиановый нож, торчащий посреди окаменелых остатков раковин и как бы вмурованный в их массу. Его форма не оставляла никаких сомнений в том, что он был обработан, но в эти места его, видимо, принесли древние мастера, ибо ближайшее месторождение обсидиана находится в 20 км отсюда. Груды раковин, кости животных, нож и прочие артефакты из обсидиана дополняли общую картину трапезы, съеденной древними людьми, которые жили на рифе много тысячелетий назад. Куски коралловых блоков крошились у нас из-под ног, но иначе нам было невозможно вскарабкаться к диковинкам и рассмотреть их вблизи. Среди коралловых обломков на песке вокруг этих огромных блоков тут и там виднелись многочисленные обсидиановые лезвия-сколы длиной 10—40 мм, причем некоторые из них по-прежнему оставались острыми как бритва. При необходимости я вполне мог бы ими побриться. Я вспомнил, что в статье в «Nature», в которой рассказывалось об этой ценнейшей археологической находке, упоминались обсидиановые ножи, обнаруженные на рифе, но это были настоящие микролиты. Главная загадка этой находки заключалась в том, что 125 тысяч лет — это слишком большая древность для микролитов, разумеется, если они не были занесены сюда в гораздо более поздние времена. Тот факт, что они были рассыпаны тут и там, а не торчали из груды окаменевших раковин, делал последнюю гипотезу вполне вероятной. Как показывают данные археологических раскопок, микролиты начали появляться в культурных слоях в Африке лишь около 80 тысяч лет тому назад. Микролиты, поблескивавшие перед нами, были рассеяны в слоях песка, покрывавших утес, и вполне вероятно, что они действительно попали сюда много позже. Интересно, что наиболее ранние микролиты за пределами Африки были найдены как раз к западу от Красного моря, в Шри-Ланке[153]. Наконец, среди особенностей поведения, присущих человеку современного типа, мы видим обмен товарами из весьма удаленных мест, то есть, другими словами, зачаточные формы торговли или доставки важнейших товаров, например обсидиановых блоков и орудий из него, на 300 км и больше. В данном случае мы вновь видим, что и этот вид человеческой деятельности впервые появился в той же Африке по меньшей мере 140 тысяч лет тому назад[154].
Ноу-хау, возраст которого — 2 миллиона лет
Сложная и многогранная картина занятий первых «анатомически современных» жителей Африки, нарисованная Мак-Брирти и Бруксом, показывает, что на момент их появления, около 140 тысяч лет тому назад, практически половина из четырнадцати важнейших критериев — ключей к оценке разумных навыков, которые в ходе дальнейшего развития позволили нам совершить полет на Луну, уже существовала в Африке. Три из них (использование пигментов, камни для растирания и обоюдоострые ножи) были изобретены еще представителями предшествующих видов человека более 140 тысяч лет тому назад. Примерно 100 тысяч лет назад, вскоре после первого исхода на Левант, возникли три четверти этих навыков, а остальные три появились задолго до того, как нога человека современного типа впервые ступила на земли Европы. С точки зрения постепенного накапливания позитивных изменений в культуре на протяжении последних 300 тысяч лет гипотезу о некоей внезапной «культурно-технической революции в Европе», имевшей место всего 40 тысяч лет назад, лопается как мыльный пузырь. И вместо привычных определений статуса человека[155] как «способности к адаптации и изобретательности, сопровождаемых физической эволюцией», мы видим, как эти качества действуют с самого начала истории нашего рода Homo — охотников, изобретателей и создателей всевозможных орудий, — насчитывающей более 2 млн. лет. Несмотря на окончательное ниспровержение его мнимой «особой роли» в эволюции, Верхний палеолит в Европе остается поистине уникальным свидетельством наиболее блистательной эпохи локального самосознания и самовозвеличивания. Но что, собственно, он может поведать нам о нас самих? Да, есть некоторые факты и совпадения, вплоть до распределения дат возникновения наиболее ранних культур эпохи Верхнего палеолита, которые действительно связаны с вторжением в Европу первых людей современного типа (ауриньякская культура и ее преемница, граветтийская культура), и притом таких вторжений было не одно, а два. В следующей главе мы поговорим о том, откуда и почему явились эти пришельцы и как генетический след подтверждает эту точку зрения.
ГЛАВА ТРЕТЬЯ ДВА ТИПА ЕВРОПЕЙЦЕВ
Как мы уже знаем, главный вопрос о происхождении европейцев заключается не в том, мигрировали ли предки будущих европейцев из Африки особым маршрутом, отдельно от предков будущих азиатов и австралийцев, и не в том, чтобы навсегда покончить с мифом о том, будто они были первыми людьми, поведение которых отвечало всем критериям поведения человека современного типа. Вопрос этот куда более серьезен. Чем объяснить столь неожиданный и пышный расцвет культуры? Носила ли их культура аборигенный, местный характер или была откуда-то занесена? Почему некоторые археологи указывают на целый ряд культурных влияний на Европу в период между 20 и 50 тысячами лет назад, причем одно из них исходило с Востока? В этой главе мы поговорим о том, что существуют особые генетические маркеры, прослеживаемые параллельно двум различным волнам культурных влияний в археологически документированной истории Европы, которые имели место около 25 тысяч лет назад, накануне последнего ледникового максимума. Они показывают, что «восточные» корни происхождения европейцев — это отнюдь не нелепый вымысел. В главе 1 мы убедились, что предки европейцев, клан N (или Насрин), принадлежали к одной из первых ветвей, отделившихся от общего ствола мигрантов, совершивших Исход из Африки и появившихся в Аравии, по всей видимости, около 80 тысяч лет тому назад. Несмотря на столь уверенное позиционирование у самого корня генетического древа по материнской линии в Азии, предкам европейцев пришлось провести в Южной Азии несколько десятков тысяч лет. Они пробыли там примерно до рубежной даты — 50 тысяч лет назад, когда влажная и теплая фаза климатического цикла покрыла Аравийскую пустыню ковром зеленой растительности, открыв путь к землям Плодородного Полумесяца — в Турцию и на Левант. Однако эти факторы никак не повлияли на их кузенов — первопроходцев из собирателей на прибрежной полосе, которые продолжали продвижение в Юго-Восточную Азию и Австралию вдоль побережья Индийского океана. Они прибыли в Австралию примерно 60 тысяч лет назад, задолго до того, как началось активное заселение Европы. С точки зрения жителей Азии, Европа была труднодоступным полуостровом, простиравшимся к северо-западу от Старого Света, своего рода географическим тупиком. В генетическом, а также в географическом отношении европейцы были побочной ветвью генеалогического древа мигрантов, совершивших исход из Африки. Поскольку первые неафриканцы современного типа появились в Азии, «полуостровная» Европа была более открытым и благодарным восприемником всевозможных культурных инноваций и изобретений эпохи Верхнего палеолита, чем жители районов, где эти инновации возникли. С этой точки зрения последняя глава была посвящена развенчанию сложившегося археологически-антропологического мифа о великой революции в области биологии человека, якобы совершившейся в Европе и на Леванте, после чего все остальное человечество будто бы последовало за «передовыми» европейцами.
Первые европейцы современного типа
В предыдущей главе мы уже говорили о том, что наши кузены-неандертальцы на пороге своего окончательного исчезновения около 28 тысяч лет назад тоже начали осваивать технические идеи Верхнего палеолита. Это частичная путаница в сфере культурных различий между неандертальцами и людьми современного типа не означает, что археологи не в состоянии обнаружить древнейшие материальные следы и артефакты, свидетельствующие о проникновении в Европу людей современного типа. Напротив, начиная примерно с 30 тысяч лет тому назад по всей территории Европы начали быстро и успешно распространяться сразу несколько таких технико-культурных традиций, связанных с людьми современного типа. Сроки и направления распространения этих традиций были весьма и весьма различными, и поэтому они получили целый ряд названий, связанных со стилевыми особенностями самих орудий и названиями мест, где они были впервые обнаружены. Природа ранних культур человека современного типа в Европе была существенно разной, и некоторые археологи склонны классифицировать их как две основных волны. Концепция переселений может быть развита значительно дальше, чем простое описание диффузии (проникновения) культур, а это — аргументы в пользу двух ветвей миграции, носительниц нескольких различных культур. Первую из этих волн, проникшую в Европу около 46 тысяч лет тому назад, принято называть ауриньякской культурой — по названию деревушки Ауриньяк (Верхняя Гаронна) в южной Франции, где впервые были обнаружены артефакты этой культуры. Более поздняя культура, сложившаяся примерно 21—30 тысяч лет назад, получила название граветтийской — по местности Ля Граветт в районе Перигор (Франция). Для нее характерны односторонние ножи (одна кромка которых притуплена, как у перочинного ножа) и остроконечные лезвия[156]. Весьма соблазнительно приписать распространение этих идей массовым миграциям древних людей. Однако, как показывает история более поздних времен, распространение идей и навыков в широких массах населения может быть более быстрым и всеобъемлющим, чем миграции самих людей. Что же касается первых ветвей заселения Европы людьми современного типа, то можно говорить о существовании генетических свидетельств как минимум двух отдельных миграций, соответствующих появлению новых технико-культурных традиций, которые принесли с собой люди современного типа.
Ауриньякская культура
Ауриньякская культура эпохи Верхнего палеолита впервые появилась в Европе на территории нынешней Болгарии, куда ее носители проникли, по всей видимости, из Турции. Вскоре после этого новый тип каменных орудий начал быстро распространяться вверх по Дунаю. Он появился на стоянках в Исталлоско, Венгрия, а затем к западу от Дуная, в Виллендорфе, Австрия. Неудержимое продвижение ауриньякской культуры вверх по Дунаю и на запад от Черного моря вскоре привело к ее появлению в верховьях Дуная — в Гейссенклёстерле, Германия. Однако задолго до этого ауриньякская культура продвинулась к югу от Австрии и проникла в северную Италию. Оттуда она стала быстро распространяться по всей Ривьере[157], перебралась через Пиренеи, достигла Эль Кастильо в северной Испании и, наконец, около 38 тысяч лет тому назад появилась на Атлантическом побережье Португалии (рис. 3.1)[158]. Ауриньякская культурная традиция в том или ином виде просуществовала до позднейших времен, и ее датировка и атрибуция по останкам человеческих скелетов носит весьма условный характер. Однако это раннее и быстрое проникновение в регионы, в которых прежде отмечались лишь следы культур эпохи Среднего палеолита, свидетельствует о том, что здесь действительно имела место колонизация.
 Рис.3.1
Что касается стилей каменных орудий ауриньякской культуры за пределами Европы, то пока не обнаружено археологически достоверных артефактов, относящихся к периоду ранее 47 тысяч лет тому назад. Наиболее реальный кандидат на роль ее предшественника — Ближний Восток. Так, бельгийский археолог Марсель Отте установил, что горы Загрос (являющиеся частью Плодородного Полумесяца) были прародиной техники создания каменных орудий ауриньякской культуры. Это хорошо согласуется с моей точкой зрения о том, что Плодородный Полумесяц в эпоху палеолита служит своеобразным коридором для проникновения на Левант. Маршрут проникновения первых людей современного типа в Европу, вероятнее всего, пролегал через нынешний Босфорский пролив (который в те отдаленные времена был сухопутным перешейком, а Черное море — пресноводным озером)[159].
Рис.3.1
Что касается стилей каменных орудий ауриньякской культуры за пределами Европы, то пока не обнаружено археологически достоверных артефактов, относящихся к периоду ранее 47 тысяч лет тому назад. Наиболее реальный кандидат на роль ее предшественника — Ближний Восток. Так, бельгийский археолог Марсель Отте установил, что горы Загрос (являющиеся частью Плодородного Полумесяца) были прародиной техники создания каменных орудий ауриньякской культуры. Это хорошо согласуется с моей точкой зрения о том, что Плодородный Полумесяц в эпоху палеолита служит своеобразным коридором для проникновения на Левант. Маршрут проникновения первых людей современного типа в Европу, вероятнее всего, пролегал через нынешний Босфорский пролив (который в те отдаленные времена был сухопутным перешейком, а Черное море — пресноводным озером)[159].
Как согласуется датировка генов и каменных орудий
Самый волнующий вопрос здесь — это вопрос о том, не обнаружены ли генетические следы этого первого проникновения в Европу Как это ни удивительно, некое подобие следов такой древности позволяет обнаружить лишь митохондриевая линия; именно она имела предков на Ближнем Востоке Другими словами, это — ключ, показывающий, что Европа первоначально заселялась потомками одной группы мигрантов. Подобная точка зрения не явилась результатом открытия некоего «справочника по генетической истории», где можно проследить точные даты появления любого гена, ибо такого справочника просто не существует[160]. Туман, которым окутана генетическая история Европы, еще только начинает рассеиваться. Прежде всего надо сказать, что за последние 50 тысяч лет в Европе происходили многочисленные массовые миграции населения, и ее территория подвергалась пагубному воздействию ледниковых периодов. Войны, вторжения и позднейшие миграции в Европу и на Ближний Восток и обратно всякий раз основательно перемешивали варево этносов в генетическом котле европейцев.
 Рис. 3.2
Авторитетная интернациональная группа, состоящая из тридцати шести сотрудников во главе с видным генетиком-эволюционистом Мартином Ричардсом, ныне — сотрудником Хаддерсфилского университета в Англии, собрала все имеющиеся в наличии данные по мтДНК и недавно опубликовала подробное исследование. Ученые изучили практически все имеющиеся данные исследований мтДНК по Европе и, используя эффективную методику выявления и устранения погрешностей, в частности обратные миграции, выявили одиннадцать наследственных линий-родоначальниц для Европы и восемнадцать исходных линий — для Ближнего Востока[161].
С помощью генетических часов Ричардс и его коллеги сумели провести датировку исходных линий на Ближнем Востоке и линий-родоначальниц в Европе (рис. 3.2). Четыре исходных линии (J, T, U5 и I на рис. 3.2) на Леванте могли датироваться периодом от 45 до 55 тысяч лет назад, что свидетельствует о том, что Ближний Восток в те времена сам был объектом колонизации потомками линий, которые были «дочерьми» и «внучками» ветви Насрин.
Рис. 3.2
Авторитетная интернациональная группа, состоящая из тридцати шести сотрудников во главе с видным генетиком-эволюционистом Мартином Ричардсом, ныне — сотрудником Хаддерсфилского университета в Англии, собрала все имеющиеся в наличии данные по мтДНК и недавно опубликовала подробное исследование. Ученые изучили практически все имеющиеся данные исследований мтДНК по Европе и, используя эффективную методику выявления и устранения погрешностей, в частности обратные миграции, выявили одиннадцать наследственных линий-родоначальниц для Европы и восемнадцать исходных линий — для Ближнего Востока[161].
С помощью генетических часов Ричардс и его коллеги сумели провести датировку исходных линий на Ближнем Востоке и линий-родоначальниц в Европе (рис. 3.2). Четыре исходных линии (J, T, U5 и I на рис. 3.2) на Леванте могли датироваться периодом от 45 до 55 тысяч лет назад, что свидетельствует о том, что Ближний Восток в те времена сам был объектом колонизации потомками линий, которые были «дочерьми» и «внучками» ветви Насрин.
 Рис 3.3
Как показано на рис 3.3, это были, разумеется, линии правнучек и праправнучек L3, нашей общей Евы выходцев из Африки. Благодаря достаточно широкому спектру погрешностей для данных мтДНК, период от 45 до 55 тысяч лет назад включает в себя наиболее ранние данные эпохи Верхнего палеолита, что вполне согласуется с временем колонизации Ближнего Востока в Верхнем палеолите[162].
Рис 3.3
Как показано на рис 3.3, это были, разумеется, линии правнучек и праправнучек L3, нашей общей Евы выходцев из Африки. Благодаря достаточно широкому спектру погрешностей для данных мтДНК, период от 45 до 55 тысяч лет назад включает в себя наиболее ранние данные эпохи Верхнего палеолита, что вполне согласуется с временем колонизации Ближнего Востока в Верхнем палеолите[162].
Пятая дочь Европы
Пожалуй, наиболее противоречивым выводом, к которому пришли многие исследователи в рамках этого обширного обзора генетической предыстории материнских линий Европы, является идентификация и датировка первой линии-родоначальницы в Европе, U5. Первоначально в Европу проникла линия U5, генетическая праправнучка одной из четырех исходных линий на Леванте (рис. 3 4). Этот европейский клан вобрал в себя характерные черты Ближнего Востока и Европы. Несмотря на свою глубокую древность, эта линия не встречается в Восточной Азии, будучи ограничена к западу от Центральной Азии Левантом и Персидским заливом, странами Средиземноморья и Европой, а ее древняя дочерняя ветвь, U2i, присутствует в Индии. Общий возраст этого клана на Ближнем Востоке превышает 50 тысяч лет Согласно молекулярным часам, наша искомая пятая дочь Европы, U5, также имеет возраст около 50 тысяч лет и представляет собой наиболее древнюю линию во всей Европе, возникшую за 15 тысяч лет до появления в Европе следующей линии-родоначальницы. Но каким путем генетический сигнал от 54 до 50 тысяч лет назад распространился на Ближний Восток, а оттуда через дочернюю ветвь U5 в Европу, если археологические датировки появления наиболее ранних памятников людей Верхнего палеолита на Леванте и древнейших артефактов ауриньякской культуры в Болгарии составляют соответственно 47 и 46 тысяч лет? Это несоответствие можно объяснить систематическим занижением данных при радиоуглеродном анализе любого времени свыше 40 тысяч лет вследствие так называемого потолочного эффекта[163]. Хотя линия U5, по-видимому, возникла на Ближнем Востоке примерно в то же время, когда она проникла в Европу, ее потомков можно встретить только на весьма ограниченной территории среди представителей национальных меньшинств, проживающих в Турции и Транскавказском регионе Турции и Ирана (см. рис. 3.4).
 К этим меньшинствам относятся турки, армяне, азербайджанцы и курды. Все эти народы и сегодня живут в границах древнего Плодородного Полумесяца, простирающегося от Турции и Транскавказского региона на юго-восток через горы Загрос на территории Ирака и Ирана. Плодородный Полумесяц почти совпадает с землями Курдистана и, наконец, образует своего рода коридор, проходящий параллельно Месопотамии, но к северу от нее, вплоть до побережья Персидского залива — части акватории Индийского океана. Особенно показательно, что линия U5 почти полностью отсутствует в Аравии, что не позволяет считать предков этих народов основным этническим материалом первой волны колонизации Европы носителями ауриньякской культуры.
Есть ли у нас шанс найти генетические следы, согласующиеся с теорией быстрого продвижения создателей орудий ауриньякской культуры на запад, на территорию Центральной Европы, в результате которого около 40 тысяч лет тому назад они достигли Пиренеев и Испании? Хотя линия U5 в наши дни распространена в Европе практически повсеместно, мы знаем, что старейшая правнучка Европы, линия U5а, возраст которой — около 40 тысяч лет, является наиболее распространенной в Стране Басков, находящейся на севере Испании. Будучи одним из немногих прибежищ европейцев во время последнего ледникового периода, Страна Басков[164] сумела сохранить свое исходное генетическое многообразие куда лучше и полнее, чем другие районы Западной Европы.
Таким образом, линия U5 — одна из немногих уцелевших дочерних линий Европы, позволяющая проследить родословную предков первых европейцев вплоть до 50 тысяч лет назад. Эта линия является общей у европейцев с армянами, турками, азербайджанцами и курдами. Что же нам известно о ее семействе, о том, откуда она пришла и кто были ее ближайшие сестры? Изучение древа генетических линий (см. рис. 3.3) дает нам генеалогию, которую мы можем интерпретировать в библейском духе. Европа в генетическом отношении была дочерью Рохани, которая, в свою очередь, была дочерью Насрин, генетической дочери L3 — «Евы выходцев из Африки». Но по какому же маршруту материнский клан Европы прибыл на Левант и где родилась его дочь, линия U5, колонизовавшая и заселившая Европу? Нам неизвестны коренные типы Насрин и Рохани, за исключением Южной Азии, где потомки Насрин встречаются лишь спорадически, а потомки Рохани представлены широким и вариативным генетическим спектром. Большинство типов Рохани, имеющихся в Индии, не встречаются более нигде, но именно это многообразие типов Рохани в Индии и позволяет нам определить, где линия U5 начала ветвиться. Это произошло, видимо, по меньшей мере 55 тысяч лет назад и, таким образом, предшествовало проникновению дочери Рохани, Европы, на Левант. А это дает все основания считать, что прародиной этих линий предков европейцев была Южная Азия. Но даже эта датировка, вполне возможно, недооценивает истинный возраст клана потомков Рохани. Весьма вероятно, что возраст клана Рохани в Азии — значительно старше 55 тысяч лет. Более того, по оценкам китайских исследователей, две азиатские подгруппы потомков Рохани имеют гораздо более почтенный возраст[165].
К этим меньшинствам относятся турки, армяне, азербайджанцы и курды. Все эти народы и сегодня живут в границах древнего Плодородного Полумесяца, простирающегося от Турции и Транскавказского региона на юго-восток через горы Загрос на территории Ирака и Ирана. Плодородный Полумесяц почти совпадает с землями Курдистана и, наконец, образует своего рода коридор, проходящий параллельно Месопотамии, но к северу от нее, вплоть до побережья Персидского залива — части акватории Индийского океана. Особенно показательно, что линия U5 почти полностью отсутствует в Аравии, что не позволяет считать предков этих народов основным этническим материалом первой волны колонизации Европы носителями ауриньякской культуры.
Есть ли у нас шанс найти генетические следы, согласующиеся с теорией быстрого продвижения создателей орудий ауриньякской культуры на запад, на территорию Центральной Европы, в результате которого около 40 тысяч лет тому назад они достигли Пиренеев и Испании? Хотя линия U5 в наши дни распространена в Европе практически повсеместно, мы знаем, что старейшая правнучка Европы, линия U5а, возраст которой — около 40 тысяч лет, является наиболее распространенной в Стране Басков, находящейся на севере Испании. Будучи одним из немногих прибежищ европейцев во время последнего ледникового периода, Страна Басков[164] сумела сохранить свое исходное генетическое многообразие куда лучше и полнее, чем другие районы Западной Европы.
Таким образом, линия U5 — одна из немногих уцелевших дочерних линий Европы, позволяющая проследить родословную предков первых европейцев вплоть до 50 тысяч лет назад. Эта линия является общей у европейцев с армянами, турками, азербайджанцами и курдами. Что же нам известно о ее семействе, о том, откуда она пришла и кто были ее ближайшие сестры? Изучение древа генетических линий (см. рис. 3.3) дает нам генеалогию, которую мы можем интерпретировать в библейском духе. Европа в генетическом отношении была дочерью Рохани, которая, в свою очередь, была дочерью Насрин, генетической дочери L3 — «Евы выходцев из Африки». Но по какому же маршруту материнский клан Европы прибыл на Левант и где родилась его дочь, линия U5, колонизовавшая и заселившая Европу? Нам неизвестны коренные типы Насрин и Рохани, за исключением Южной Азии, где потомки Насрин встречаются лишь спорадически, а потомки Рохани представлены широким и вариативным генетическим спектром. Большинство типов Рохани, имеющихся в Индии, не встречаются более нигде, но именно это многообразие типов Рохани в Индии и позволяет нам определить, где линия U5 начала ветвиться. Это произошло, видимо, по меньшей мере 55 тысяч лет назад и, таким образом, предшествовало проникновению дочери Рохани, Европы, на Левант. А это дает все основания считать, что прародиной этих линий предков европейцев была Южная Азия. Но даже эта датировка, вполне возможно, недооценивает истинный возраст клана потомков Рохани. Весьма вероятно, что возраст клана Рохани в Азии — значительно старше 55 тысяч лет. Более того, по оценкам китайских исследователей, две азиатские подгруппы потомков Рохани имеют гораздо более почтенный возраст[165].
Возраст трех первых из семи «дочерей» Европы — по 50 тысяч лет
У пятой генетической дочери клана Европы были и другие сестры; всего дочерей Европы было семь, но лишь две из них имели такой же возраст, как и U5. Это были ветви U6 и U2i, и весьма характерно, что ни та ни другая ветвь в Европе не представлены. Мы уже проследовали за ветвью U6 в Северную Африку. Уникальный идентификатор берберов, эта ветвь имеет такой же возраст, как и U5, а именно 50 тысяч лет, указывая, что в то же самое время, когда представители ветви U5 отправились из Турции на северозапад, в будущую Европу, ветвь U6 двинулась на северо-запад, вдоль южного побережья Средиземного моря, в Северную Африку. Мы располагаем даже материальными свидетельствами продвижения этих первопоселенцев Северной Африки, прибывших туда с Леванта. Орудия эпохи начала Верхнего палеолита, найденные на побережье Ливии, на стоянке Хауа Фтеах, могут быть датированы примерно 40 тысяч г. до н.э., что свидетельствует о раннем проникновении мигрантов в Африку[166]. Третья из трех генетических дочерей-«сверстниц» клана Европы, возраст которых достигает 50 тысяч лет, по всей видимости, появилась на свет на расстоянии примерно четверти окружности земного шара от берберов Ливии, где-то на побережье Индийского океана. Ветвь U2i, на долю которой приходится около 9,5% всех материнских линий в Индии и свыше 78% всех линий — выходцев из Индии в Европе, несомненно, является коренной уроженкой этого региона. В Индии ее возраст составляет 53 тысячи лет. U2i почти отсутствует на Леванте и в Европе, где мы встречаем небольшое ее ответвление, U2e, возраст которого составляет 2/3 от возраста самой U2i[167]. (См. рис. 3.4.) Другим индикатором, указывающим на южноазиатский регион как на возможный источник генетических предков этого клана Европы, явилось нахождение там древнейших корней европейского типа и зарождение другой ветви, представленной в Европе, — U7, которая впоследствии проникла на Ближний Восток и в Европу[168].
Когда же был открыт коридор Плодородного Полумесяца
Наше материнское генетическое древо свидетельствует о том, что возраст древнейших выходцев из Южной Азии, ставших родоначальниками современных европейцев, составляет не менее 50 тысяч лет. Чтобы проникнуть далее на север, в Анатолию[169], им необходимо было преодолеть Ливийскую и Аравийскую пустыни, воспользовавшись Плодородным Полумесяцем в качестве проходного коридора. Учитывая достаточно широкий диапазон погрешностей, свойственный молекулярным часам, при анализе полосы земель, простирающейся от гор Загрос и болот на побережье Персидского залива на юго-западе Ирана, нам необходимо оценивать их миграции с точки зрения благоприятных климатических циклов, которые, подобно кольцам на срезе дерева, дают наиболее точную и достоверную датировку. Как я уже говорил в главе 1, коридор, пролегавший через Плодородный Полумесяц, был крайне засушливым и оставался закрытым на протяжении последних 100 тысяч лет, открываясь лишь очень ненадолго в результате улучшения климатической обстановки во время так называемых «интерстадиальных» (промежуточных) пауз (см. рис. 1.7). Примерно между 55 и 65 тысячами лет тому назад на нашей планете наступил суровый период невиданных холодов и засухи. В ту эпоху коридор через Плодородный Полумесяц оставался закрытым. Затем, начиная с 56 тысяч лет тому назад, настала полоса из четырех последовательно сменявших друг друга теплых и влажных циклов. Последний из них, начавшийся около 51 тысячи лет назад, оказался самым теплым и длительным; он продолжался примерно 5000 лет. Эта промежуточная пауза оказалась настолько теплой и влажной, что муссоны на побережье Индийского океана в ту эпоху были даже более обильными влагой, чем в наши дни. А это означает, что, помимо открытия коридора через Плодородный Полумесяц, такие засушливые районы Леванта, как пустыня Негев, потенциально могли быть вполне пригодными для обитания для наших создателей каменных орудий эпохи Верхнего палеолита. И если люди и получали в истории благоприятную возможность для умножения своей численности в Южной Азии и миграции на Левант, то это произошло именно в это время. Датировки по климатическим и археологическим свидетельствам также указывают на особо благоприятный период между 45 и 50 тысячами лет тому назад. Таким образом, получается, что датировка по молекулярным часам времени прибытия на Левант наиболее ранних дочерних линий Насрин и их «семейств» не слишком отличается от этих значений[170].
История Адама
Итак, мы познакомились с историей происхождения женского клана предков европейцев, сложившегося в Южной Азии более 50 тысяч лет тому назад. Полное отсутствие в Европе и на Леванте клана Манью, занимающего доминирующее положение в Индии, свидетельствует о том, что Европа могла появиться на свет к западу от самой Индии, у основания Плодородного Полумесяца, где-нибудь в районе древнего города Ур у побережья Персидского залива. Но, естественно, там, где были матери, должны были быть и отцы. В главе 1 я вкратце рассказал линию потомков Адама, совершивших исход из Африки: Каина, Авеля и Сифа, которые покинули Африку по южному маршруту, отправившись заселять весь остальной мир. Все эти три генетических сына являются потомками одного и того же исхода[171]. Не вправе ли мы предположить, что у трех сыновей Адама, выходцев из Африки, существовали свои истории, служащие зеркальным отражением историй двух дочерей внеафриканской Евы на их пути из Южной Азии? Да, так оно и есть, и географическая история потомков как минимум одной линии Y-хромосом, линии Сифа, является еще более захватывающей, чем легенда о Европе, хотя датировать ее время гораздо труднее. (Что касается другой линии, типа Авеля или YAP, то ее аргументация, по-видимому, представляет собой одну из величайших загадок в древнейшей генетической истории мужских кланов; см. главу 4.) Как и в истории дочерей Европы, нам придется проследить два побега вплоть до точки начала их ответвления с ветвей, чтобы установить, в каком направлении развивались генеалогические линии мужчин. Потомки Сифа, сына Адама выходцев из Африки (рис. 3.5), являются самой многочисленной в мире ветвью за пределами Африки. Взяв за основу ветвь Сифа, надо отметить, что один из пяти его сыновей занимает доминирующее положение на Среднем Востоке; на его долю приходится примерно от четверти до половины всех мужских линий в обширном регионе между Ираном и Средиземноморьем. Согласно общепринятой классификации, этого «сына» обычно обозначают буквой J. Я же называю его Джахангир, чтобы подчеркнуть его предполагаемое происхождение из Южной Азии. В Западной и Южной Евразии. Джахангир, как правило, следовал за расселением клана Европы.

 Рис.3.5
Наиболее высокая встречаемость этой генетической линии в Европе отмечена на северо-западном побережье Средиземного моря. Что касается линии Джахангира, то она чаще всего встречается на землях Леванта (30—60%). Наибольшая ее «популяция» в Европе отмечена в Анатолии, где она составляет 40%; затем следуют Балканы и Италия: 20—30%. И хотя потомки линии Джахангира распространены по всей Европе, наиболее высокая их плотность наблюдается в районах, прилегающих к побережью Средиземного моря. Отдельные высокие уровни линии J отмечены и в странах Северной Африки, например, 41% в Алжире[172].
Эта характерная особенность — расселение в прибрежных районах Средиземного моря — отражает расселение линий U5 и U6 в этом регионе около 50 тысяч лет тому назад. Поэтому весьма соблазнительно обратиться вспять и проследить путь линии Джахангира вплоть до самых ее истоков. Последовав за ее ветвью, мы видим, что она, пролегая по Плодородному Полумесяцу через Курдистан, все настойчивее заявляет о себе по мере того, как мы продвигаемся дальше на юго-восток. Имея на всем протяжении Ирана показатель встречаемости не более 35%, мужской клан Джахангир достигает уровня 55% в южном Прикаспии и 59% — еще дальше к югу, в горах Загрос. Кроме того, Джахангир встречается еще далее к югу, в Пакистане и Индии, а также в Центральной Азии и Сибири, но там эти показатели значительно ниже[173].
Понятно, что сам по себе показатель встречаемости не способен поведать нам, где именно была прародина клана, а вот многообразию генетических линий это по силам. Парижский генетик Льюи-Кинтана-Мюрси и его коллеги утверждают, что первоначальной прародиной клана Джахангир были горы Загрос в южном Курдистане — древней стране Элам, расположенной на северо-восточном побережье Персидского залива. И надо признать, их заявления имеют под собой веские основания, ибо именно в горах Загрос отмечено самое высокое в мире многообразие типов линии Джахангир[174]. Как я уже упоминал выше, точность и достоверность методов датировки Y-хромосом, большинство из которых предлагают существенно заниженные данные, являются весьма спорными, но один из этих методов дает такие данные для клана Джахангир: около 55 тысяч лет в Иране и 62 тысячи лет — южнее, в Индии. Ясно, что эти данные не согласуются с генетической историей материнских кланов, поскольку целый ряд методов дает значительно более поздние даты в отношении клана Джахангир, равно как и в отношении других Y-линий.
Итак, мы видели, как согласуются друг с другом археологические, климатические и генетические истории первого появления людей современного типа на Леванте и, следовательно, в Европе и СевернойАфрике, около 50 тысяч лет тому назад. Это происхождение предков европейцев из Южной Азии резко контрастирует с обычными представлениями о маршруте исхода, пролегавшем вокруг восточного Средиземноморья в Европу. Свидетельства подобного маршрута, мягко говоря, весьма скудны.
Вместо этого мы можем проследить даты и локусы стоянок на пути вдоль склонов гор Загрос с юга, от побережья Персидского залива. Климатически благоприятным временем для миграции по Плодородному Полумесяцу через Курдистан была довольно продолжительная теплая интерстадиальная пауза. Имеется целый ряд археологических доказательств того, что прародиной ауриньякской культуры были именно горы Загрос. Наиболее ранняя стоянка представителей ауриньякской культуры в Европе обнаружена на территории Болгарии, неподалеку от Босфора. Ее возраст — около 47 тысяч лет. Если обратиться к генетическим эквивалентам такой миграции, можно найти следы ранней колонизации Ближнего Востока многочисленными материнскими линиями, возникшими некогда, около 50 тысяч лет тому назад, в Южной Азии, а теперь характерными скорее для Западной Евразии. Любопытно, что никаких генетических доказательств присутствия этих линий в Северной Африке не существует, поэтому данный факт вполне логично рассматривать своего рода указанием на маршрут исхода через Южную Азию. Примерно в одно и то же время с колонизацией Леванта три дочерних клана Европы становятся родоначальниками этносов Индии, Северной Африки и Европы. Наконец, дополнительным свидетельством в пользу такого расселения материнских линий можно считать мужской клан Джахангир, который также сформировался в Южной Азии, хотя датировка этого события вызывает разногласия.
Рис.3.5
Наиболее высокая встречаемость этой генетической линии в Европе отмечена на северо-западном побережье Средиземного моря. Что касается линии Джахангира, то она чаще всего встречается на землях Леванта (30—60%). Наибольшая ее «популяция» в Европе отмечена в Анатолии, где она составляет 40%; затем следуют Балканы и Италия: 20—30%. И хотя потомки линии Джахангира распространены по всей Европе, наиболее высокая их плотность наблюдается в районах, прилегающих к побережью Средиземного моря. Отдельные высокие уровни линии J отмечены и в странах Северной Африки, например, 41% в Алжире[172].
Эта характерная особенность — расселение в прибрежных районах Средиземного моря — отражает расселение линий U5 и U6 в этом регионе около 50 тысяч лет тому назад. Поэтому весьма соблазнительно обратиться вспять и проследить путь линии Джахангира вплоть до самых ее истоков. Последовав за ее ветвью, мы видим, что она, пролегая по Плодородному Полумесяцу через Курдистан, все настойчивее заявляет о себе по мере того, как мы продвигаемся дальше на юго-восток. Имея на всем протяжении Ирана показатель встречаемости не более 35%, мужской клан Джахангир достигает уровня 55% в южном Прикаспии и 59% — еще дальше к югу, в горах Загрос. Кроме того, Джахангир встречается еще далее к югу, в Пакистане и Индии, а также в Центральной Азии и Сибири, но там эти показатели значительно ниже[173].
Понятно, что сам по себе показатель встречаемости не способен поведать нам, где именно была прародина клана, а вот многообразию генетических линий это по силам. Парижский генетик Льюи-Кинтана-Мюрси и его коллеги утверждают, что первоначальной прародиной клана Джахангир были горы Загрос в южном Курдистане — древней стране Элам, расположенной на северо-восточном побережье Персидского залива. И надо признать, их заявления имеют под собой веские основания, ибо именно в горах Загрос отмечено самое высокое в мире многообразие типов линии Джахангир[174]. Как я уже упоминал выше, точность и достоверность методов датировки Y-хромосом, большинство из которых предлагают существенно заниженные данные, являются весьма спорными, но один из этих методов дает такие данные для клана Джахангир: около 55 тысяч лет в Иране и 62 тысячи лет — южнее, в Индии. Ясно, что эти данные не согласуются с генетической историей материнских кланов, поскольку целый ряд методов дает значительно более поздние даты в отношении клана Джахангир, равно как и в отношении других Y-линий.
Итак, мы видели, как согласуются друг с другом археологические, климатические и генетические истории первого появления людей современного типа на Леванте и, следовательно, в Европе и СевернойАфрике, около 50 тысяч лет тому назад. Это происхождение предков европейцев из Южной Азии резко контрастирует с обычными представлениями о маршруте исхода, пролегавшем вокруг восточного Средиземноморья в Европу. Свидетельства подобного маршрута, мягко говоря, весьма скудны.
Вместо этого мы можем проследить даты и локусы стоянок на пути вдоль склонов гор Загрос с юга, от побережья Персидского залива. Климатически благоприятным временем для миграции по Плодородному Полумесяцу через Курдистан была довольно продолжительная теплая интерстадиальная пауза. Имеется целый ряд археологических доказательств того, что прародиной ауриньякской культуры были именно горы Загрос. Наиболее ранняя стоянка представителей ауриньякской культуры в Европе обнаружена на территории Болгарии, неподалеку от Босфора. Ее возраст — около 47 тысяч лет. Если обратиться к генетическим эквивалентам такой миграции, можно найти следы ранней колонизации Ближнего Востока многочисленными материнскими линиями, возникшими некогда, около 50 тысяч лет тому назад, в Южной Азии, а теперь характерными скорее для Западной Евразии. Любопытно, что никаких генетических доказательств присутствия этих линий в Северной Африке не существует, поэтому данный факт вполне логично рассматривать своего рода указанием на маршрут исхода через Южную Азию. Примерно в одно и то же время с колонизацией Леванта три дочерних клана Европы становятся родоначальниками этносов Индии, Северной Африки и Европы. Наконец, дополнительным свидетельством в пользу такого расселения материнских линий можно считать мужской клан Джахангир, который также сформировался в Южной Азии, хотя датировка этого события вызывает разногласия.
Вторая волна проникновения в Европу: граветтийская культура
Мартин Ричардс и его коллеги вытянули из запутанного клубка линий Насрин на Среднем Востоке вторую нить миграции будущих европейцев и еще одну, но совершенно отличную от первой историю. Исследователи назвали этот второй клан, проникший путем миграции в Европу, HV, по двум дочерним ветвям, Н и V, которые нередко можно встретить в Европе в наши дни (см. рис. 3.4). Вторая дочерняя линия, V, появилась гораздо позже, где-то на юго-западе Европы. На долю потомков Н приходится практически половина всех материнских линий Западной и Северной Европы, в частности, у славянских, финноугорских и германских народов. Подобный ареал резко отличается от истории развития ветви U5. Активная экспансия HV датируется временем около 335 тысячи лет тому назад, что на 15 тысяч лет позже, чем появление U5, и за 7500 лет до прибытия следующей волны мигрантов. Особенно интересно, что возраст клана HV на Ближнем Востоке (26 500 лет) значительно моложе, чем в Европе, где он составляет свыше 33 тысяч лет. Это позволяет исключить Левант, Анатолию и Болгарию из числа кандидатов на роль прародины клана HV, и поэтому нам следует заняться поисками другого маршрута проникновения HV в Европу, помимо Леванта и Анатолии[175]. Группа эстонских генетиков, возглавляемая Рихардом Виллемсом и Тоомасом Кивисилдом, чей основополагающий труд по изучению мтДНК явился важным шагом вперед в области исследования генетической истории предков индийцев в эпоху палеолита, сумела собрать немало сведений об истоках и расселении HV. Эстонские генетики установили, что наиболее древние корни HV обнаружены в Южной Азии (в северо-западной Индии и Кашмире; их возраст — около 40 тысяч лет), но первым ареалом их распространения в Евразии стал Транскавказский регион (Закавказье)[176]. (См. рис. 3.4.) Этот труд эстонских ученых позволил установить, что горные районы Кавказа, раскинувшиеся между Черным и Каспийским морями, были отправной точкой для целого ряда материнских кланов, проникших в Европу, наиболее крупным и значительным из которых был HV. У многих людей, и в том числе у меня самого, при слове «Кавказ» возникает образ темпераментных и независимых народов, издревле живших на юго-западе бывшего Советского Союза, а также знаменитые горные хребты Кавказа, от которых происходит название всех жителей этого региона — «кавказцы», по непонятной причине использующееся применительно ко всем европейцам. Удивительно колоритная и запутанная картина кавказских языков пленяет своим невероятным многообразием и таинственной древностью. Кавказ, опоясанный с севера и юга ареалами распространения многочисленных индоевропейских и алтайских языков, сохранил две уникальные и очень древние языковые семьи — северо-кавказскую и картвельскую[177]. Этот регион, ограниченный с запада и востока двумя внутренними морями и перерезанный хребтом Кавказа, образует единственный проходимый для человека коридор между нынешними Арменией и Азербайджаном, ведущий в страны Леванта на юге и европейскую Россию на севере. Способны ли мужские хромосомы помочь проследить направление второй миграции в Европу? Несмотря на весьма ограниченную точность датировки при помощи Y-xpoмосом, они благодаря своей межрегиональной специфике являются куда более выразительными, чем мтДНК. Особенный интерес представляет загадочный мужской клан, которому я дал название Инос (по имени сына Сифа, Еноса[178]). Инос — это почти исключительно европейский клан. Согласно данным Зоэ Россер и др., генетиков из Лейчестера, сравнительно равномерный характер распределений линий этого клана свидетельствует о его весьма раннем проникновении в Европу. Итальянский генетик Орнелла Семино и ее американские коллеги более конкретны: они указывают, что преобладание клана Инос на Украине и на Балканах говорит о его связи с материнским кланом HV, а также с граветтийской культурой. Как и HV, клан Инос не имел четко идентифицируемых истоков на Леванте, поэтому вполне вероятно, что он также проник в Европу по маршруту, пролегавшему через Закавказский регион[179].
«Пауза» в обитаемости Ближнего Востока?
Прежде чем вновь погрузиться в генетические изыскания, мы должны попытаться выяснить, какими именно археологическими свидетельствами занятий древнейших людей в Закавказском «коридоре» мы располагаем. В предыдущей главе мы уже видели, что, по крайней мере в Западной Евразии, появление человека современного типа ассоциировалось с переходом от орудий мустерийского типа (Средний палеолит), которыми пользовались еще неандертальцы, к орудиям эпохи Верхнего палеолита, которые использовали европейцы современного типа. На Леванте наиболее ранние свидетельства такого перехода от орудий мустерийского типа к переходным техникам Верхнего палеолита найдены на юге, на землях современных пустыни Негев и Синайской пустыни. Их возраст — от 45 до 47 тысяч лет. Затем в обитаемости Леванта наступила долгая, продолжавшаяся свыше 10 тысяч лет, пауза, после чего на этих землях вновь появились люди, создававшие орудия эпохи Раннего Верхнего палеолита, так называемого ахмарийского типа, относившиеся к культуре, которая просуществовала примерно до 20 000 г. до н.э. Подобная пауза совпадала с резким ухудшением климата, кульминациями которого явились два холодных и засушливых максимума — около 45 и 40 тысяч лет тому назад[180]. Между 34 и 40 тысячами лет тому назад на нашей планете установился холодный, засушливый климат, и сколько-нибудь интенсивных индийских муссонов практически не было. Продолжительное потепление, начавшееся около 34 тысяч лет тому назад, вновь пригласило жителей в прежде засушливые районы Леванта[181]. На этот раз люди обосновались в ином месте, в знаменитой пещере у северной оконечности Плодородного Полумесяца, в северных отрогах гор Загрос — пещере Шанидар. Она расположена у южных врат кавказского «коридора». Пещера Шанидар, расположенная неподалеку от места, где на древних землях Курдистана сходятся сегодняшние границы Турции, Ирана и Ирака, к югу от Армении, получило широкую известность в 1960-е гг. благодаря усилиям Ральфа Солецки, археолога, ставшего ее первооткрывателем. Солецки считал, что ему удалось открыть объекты, которые он сам характеризовал как захоронения неандертальцев с цветами. Неандертальцы действительно жили в этих местах примерно с 50 до 46 тысяч лет тому назад. Затем, после климатической паузы продолжительностью в добрых 10 тысяч лет, обитателями этих мест стали люди другого биологического вида. Переходная техника живописи эпохи Верхнего палеолита, известная под названием барадостийской культуры, появилась в пещере Шанидар примерно около 35 500 лет тому назад и сохранялась там до 28,7 тысяч лет тому назад. Находки в пещере Шанидар — наиболее убедительные на сегодняшний день свидетельства пребывания человека современного вида у южных врат коридора, ведущего в Кавказский регион[182]. Примерно в то же время, 36 400 лет тому назад, возникла первая стоянка людей Верхнего палеолита на европейской территории России. Она обнаружена в верховьях реки Дон, у селения Костенки, в районе к северу от Кавказа[183] . Представители так называемой спицинийской культуры делали свои орудия из кремня, привозившегося из месторождений, которые находились в 150—300 км отсюда. Стоянка у с. Костенки получила широкую известность благодаря находке артефактов исключительно сложной культуры, использовавшей кости мамонтов и возникшей между 33 и 24 тысячами лет назад, но первые люди спицинийской культуры явно не знали этих артефактов. Некоторые археологи, специалисты по изучению Европы до Ледникового периода, считают, что древнейшая история европейцев эпохи палеолита состояла из нескольких четко разграниченных этапов активизации всевозможных занятий и видов деятельности, прямо связанных с улучшением климата. Первый из этих этапов, Ранний Верхний палеолит (45—33 тысячи лет тому назад), характеризовался распространением древнейших образцов ауриньякской технологии, о которых мы уже говорили. Второй этап, охватывающий время от 30 до 24 тысяч лет тому назад, знаменовал собой начало целого ряда культурных взлетов эпохи Верхнего палеолита. Совокупность разнообразных культур, сложившихся на этом этапе около 30 тысяч лет тому назад, известна под общим названием «граветтийского технокомплекса», хотя локальные названия отдельных его составляющих порой вносят путаницу в сложившуюся терминологию. И хотя эти культуры относятся уже скорее к третьему этапу заселения и развития культуры в Европе, видный эксперт по эпохе палеолита Клайв Гэмбл утверждает, что важнейший пороговый рубеж нового этапа в Европе в целом правильнее отождествлять с появлением около 33 тысяч лет тому назад стоянок в Северо-Восточной Европе, например в с. Костенки на Дону, а не с граветтийской культурой, сложившейся далеко на юго-западе около 30 тысяч лет тому назад[184]. Основными технико-культурными новшествами граветтийских культур явились систематическая добыча высококачественных источников сырья (в первую очередь — камня для орудий), наскальные росписи в пещерах, достигшие высокого уровня развития, практика сложных погребений, крупные орудия из кости и широкое использование кости, в частности — мамонтов, для строительства жилищ (см. Фото 13). Таким образом, культуры эпохи Раннего Верхнего палеолита не только демонстрируют резкое ускорение развития репрезентативного (изобразительного) искусства в Европе, использование различных экзотических материалов и практики погребений, но и свидетельствуют о проникновении на новые территории людей, принесших с собой навыки новой культуры из Восточной Европы. Подобное вторжение культуры и новых этнических групп, естественно, не ограничилось Восточной Европой, поскольку некоторые из этих культур получили наиболее совершенное воплощение в Южной Франции и Северной Испании[185].
 Фото 13. Слева: Эта женская фигурка, найденная возле Мальты неподалеку от озера Байкал, вырезана из мамонтовой кости и датируется ок. 23 тыс. лет тому назад. Ее можно считать свидетельством широкого распространения культуры охотников на мамонтов.
Справа: Палеонтологи пытаются создать реконструкцию хижины из ребер мамонта, впервые найденной во время раскопок в Нежине, Украина.
Фото 13. Слева: Эта женская фигурка, найденная возле Мальты неподалеку от озера Байкал, вырезана из мамонтовой кости и датируется ок. 23 тыс. лет тому назад. Ее можно считать свидетельством широкого распространения культуры охотников на мамонтов.
Справа: Палеонтологи пытаются создать реконструкцию хижины из ребер мамонта, впервые найденной во время раскопок в Нежине, Украина.
Вправе ли мы связывать генетический сигнал клана HV, ознаменовавшего проникновение человека современного типа в Северную и Западную Европу, с появлением около 33 тысяч лет тому назад орудий типа Раннего Верхнего палеолита? Самый короткий ответ на это — «да», ибо совершенно ясно, что к этой дате восходит и генетический сигнал женского клана, и время его миграции и миграции соответствующего мужского клана. Таким образом, клан HV может считаться свидетельством наиболее раннего проникновения людей современного типа в Северную Европу через Закавказский регион. Куда же этот клан мог направиться дальше? Да куда угодно. Как мы знаем, HV — это наиболее распространенный клан во всей Европе, но его потомки все же встречаются гораздо чаще в Северной и Юго-Западной Европе, чем на побережье Средиземного моря. У клана была дочь — европейский клан V, — появившаяся на свет гораздо позже, в районе Пиренеев, но данные последних исследований указывают на присутствие более раннего типа — предшественника V в Закавказском регионе, на Северных Балканах, в Южной Испании и Марокко, что свидетельствует о том, что распространение его на запад, в Европу, началось с территории нынешней Украины[186]. (См. рис. 3.4.)
Могли ли предки людей Раннего Верхнего палеолита быть выходцами со Среднего Востока?
Действительно, существует вероятность того, что прародиной клана HV до того, как он мигрировал в Европу, был не Закавказский регион, а районы, лежащие гораздо дальше к востоку: степи Центральной Азии. При подобном сценарии развития событий в Северокавказский регион проникли мигранты из Центральной Азии — территорий к востоку от Каспийского моря, а не из Армении, лежащей на юге. Естественно, это означает, что они двинулись на север, в Россию, по маршруту, обходящему Каспийское море с востока, а не с запада. Очень много информации дала ученым находка превосходно сохранившихся мумий кавказоидов. Эти мумии, возраст которых — около 3000 лет, были обнаружены в Урумчи (Китайский Туркестан), а также в других пунктах Центральной Азии, расположенных вдоль Шелкового пути. Независимо от того, являются ли эти находки свидетельством продвижения на восток племен эпохи неолита, говоривших на индоевропейских языках, или потомков более ранних этносов, мы располагаем реальными археологическими доказательствами присутствия в Центральной Азии людей современного типа, относящимися ко времени около 40 тысяч лет тому назад. Орудия Раннего Верхнего палеолита были найдены на двух стоянках на Алтае, Россия, и Юго-Восточной Сибири, датируемых эпохой около 39 тысяч лет тому назад Это означает, что люди современного типа появились в Центральной Азии достаточно рано, чтобы сформировать крупное миграционное ядро для проникновения в Европу через территорию современной России[187]. Третьей ветвью общего генофонда из Центральной Азии были материнские линии Насрин, распространенные в Западной Евразии. Половина из них приходится на долю клана HV. Обычное объяснение этого присутствия «Европы в Азии» — встречная эмиграция из Европы на восток, вдоль трассы Шелкового пути. Главная проблема, связанная с подобной аргументацией, заключается в том, что V, общая дочь Европы и HV, которая, как можно было бы ожидать, вполне могла присоединиться к этому переселению, полностью отсутствует в Центральной Азии. Более того, большинство других западно-евразийских линий Насрин в Центральной Азии являются, скорее всего, выходцами из Индии, нежели из Европы. Другими словами, клан HV первоначально мог мигрировать из Южной Азии и, продвигаясь на север, обогнуть Каспийское море с востока, а затем двинуться на запад, в Европу (см. рис. 3.4). Данные изучения Y-хромосомы можно считать свидетельством в поддержку точки зрения об альтернативном маршруте генетических кланов Южной Азии, мигрировавших с востока на запад, в Европу, окольным путем — через Центральную Азию[188].
Выходцы из Азии в Европе
Y-хромосома как более крупный носитель информации заключает в себе гораздо больше секретов о нашем прошлом, чем митохондриевая ДНК, этот крошечный след древнейших миграций, сохраненный на уровне микробактерии. Генетики еще только начинают расшифровывать этот секрет, и им предстоит выяснить принцип работы генетических часов потомков Адама. Данные о количестве ветвей у различных исследовательских групп варьируются на несколько порядков и, как правило, оказываются в два-пять раз «моложе», чем данные датировки на древе митохондриевой ДНК. Другими словами, у ученых есть четкий след Y-хромосомы, но они часто чрезмерно занижают его возраст. К счастью, обстоятельный анализ линий-родоначальниц и датировка по мтДНК, выполненный Мартином Ричардсом и его коллегами, а также исследование Тоомаса Кивисилда образуют как бы временные рамки генетически датированных маршрутов отслеживания Y-хромосомы. Для более тщательного изучения географических маршрутов рассеяния мы можем воспользоваться генеалогическим древом Y-хромосомы[189]. За поразительными результатами, полученными в конце 2000 г. Питером Андерхиллом из Стэнфордского университета, в области построения генетического древа человечества на основе использования 150 Y-маркеров очень быстро последовал более детальный анализ генофонда Европы, осуществленный Орнеллой Семино и ее коллегами из той же группы Андерхилла[190]. Это исследование еще более убедительно, чем анализ мтДНК, показало, что колонизация Европы осуществлялась как минимум по нескольким маршрутам. Мы уже знаем, что, параллельно с миграцией кланов дочерей Европы, Джахангир, правнук Сифа, родоначальника линии выходцев из Азии, мог расселиться по всему Плодородному Полумесяцу, отправившись в путь от побережья Персидского залива, и, таким образом, проникнуть через южные врата «коридора» в Турцию, а затем и в Европу. Но есть ли у нас данные о том, что другие мужские линии, помимо Иноса, двигались параллельно клану HV, который возник и сформировался на Кавказе, а может быть — и гораздо дальше на востоке, в азиатской России, и мигрировал на запад, проникнув через Украину в Европу? Ответ будет однозначным: да, есть, ибо здесь мы располагаем вполне конкретными следами. Я уже упоминал об Иносе, называя его типом, который мог стать зеркальным отражением HV на Северном Кавказе. Существует еще целых три генетических линии — кандидата на роль группы, проникшей в Европу. Генетики давно ломают голову над загадкой происхождения, расселения и возраста этих линий. Они, как и Иноc (и Джахангир), тоже происходят от Сифа, третьего сына Адама выходцев из Африки. Две из этих линий, очевидно, происходят с Востока, по всей вероятности — из Центральной Азии, но возникает вопрос: как давно они появились там и где находится их прародина? Надо сказать, что обе они являются потомками самого плодовитого из пяти генетических кланов-«сыновей» Сифа, К, который я для себя называю Кришна — по имени знаменитого индуистского божества.[191] Одна из этих двух генетических линий называется ТАТ, что отражает ее характерную мутацию, а также весьма созвучно имени известного оксфордского генетика Татьяны Зергал, которая активно занималась ее изучением. В Европе ареал ее расселения ограничен восточными окраинами, где эта линия встречается среди финнов и русских. Возникла она, по всей видимости, в Центральной Азии, хотя ее генетические отец и дед, несомненно, происходят из Кашмира и Южной Азии. Изредка представителей клана Кришны можно встретить и на Дальнем Востоке. Что касается датировки, то она достаточно условна, ибо мы имеем дело с Y-хромосомами, однако локальное распространение (по большей части среди населения, говорящего на языках Урала) и минимальное число вариаций в Европе свидетельствуют о ее миграции из Центральной Азии[192]. (См. рис. 3.5.) Другая линия потомков Кришны, также проникшая в Европу, характеризуется мутацией M17. Встречаемость этой линии наиболее высока среди венгров, у которых она достигает 60%. Линия М17 доминирует во всей Восточной Европе, и хотя этот клан распространен практически во всех районах Западной и Восточной Европы, а также на Леванте, его встречаемость там остается стабильно низкой. Его минимальное присутствие на Леванте заставляет отвергнуть версию о том, что этот регион мог служить для М17 воротами на пути в Европу. Льюи КинтанаМюрси, Питер Андерхилл и их коллеги в качестве наиболее вероятного кандидата на роль прародины линии M17, проникшей в Европу, считают Центральную Азию и особенно Алтай. Этот аргумент совершенно необходим для гипотезы Андерхилла и его коллег, которые полагают, что весь клан Сиф совершил отдельный исход из Северной Африки на Левант, а затем направился непосредственно в Центральную Азию[193]. (См. рис. 1.3.) На мой взгляд, а также по мнению Тоомаса Кивисилда, более логично предположить, что древнейшей прародиной линии M17 и ее предков была все же Южная Азия. Ведь именно там, в Пакистане, Индии и восточном Иране и в меньшей степени — на Кавказе, мы находим наибольшее разнообразие вариаций M17. Характерно, что M17 в Южной Азии отличается не просто гораздо более широкой вариативностью, чем в Центральной Азии; это многообразие вариаций характеризует ее присутствие в изолированных племенных группах Южной Азии, опровергая тем самым гипотезу о том, что M17 является маркером «проникновения мужского клана арьев» в Индию. Согласно одной из датировок, эта линия появилась в Индии около 51 тысячи лет тому назад. Все это свидетельствует о том, что М17 первоначально могла совершить исход из Индии и Пакистана через Кашмир, затем проникнуть в Центральную Азию и азиатскую Россию и, наконец, мигрировать на запад, в Европу (см. рис. 3.5). Между прочим, это и подобные наблюдения о характере расселения других линий потомков Сифа составляют ядро моих контраргументов против гипотезы о том, будто клан Сиф проник в Центральную Азию из Северной Африки и Леванта. Я склонен считать, что исход из Африки был только один, и притом — по южному маршруту[194]. Однако вернемся в Европу. Поскольку происхождение европейской линии М17 к востоку от Европы, на Алтае, практически не вызывает сомнений, остается вопрос о датировке подобного исхода. Семино и ее коллеги, отказавшись от генетических данных, обратились к изучению линии М17 и ее «отца», M173, и, рассматривая эти линии как составную часть общей палеолитической миграции с востока на запад, поддающейся археологической датировке, считают, что она имела место 30 тысяч лет тому назад. Итак, мы упомянули M173 — еще одного серьезного кандидата на роль мужской линии, проникшей в Европу. Эта линия относится к очень обширному клану, который я назвал Руслан — по имени героя русского фольклора[195].
Руслан: азиатский прародитель половины мужчин Европы
Клан Руслан, встречающийся в Старом Свете практически всюду — от Англии до Южной и Центральной Азии, в Сибири и даже дальше, вплоть до Австралии и Америки, — Немало постранствовал по свету, и, как говорится в одном исследовании, его коренной тип является самым распространенным типом одинарной Y-хромосомы в мире. Тем не менее ареал расселения генетического «отца» Руслана, клана Р, который я назвал «Поло» — по фамилии семейства купцов-путешественников, странствовавших по Шелковому пути, самым знаменитым из которых был Марко Поло[196], — ограничен Индией, Пакистаном[197], Центральной Азией и Северной Америкой. Исследование географического расселения и многообразия генетических ветвей опять-таки показывает, что Руслан вместе со своим «сыном», кланом M17, сформировался достаточно рано в Южной Азии, где-то в районе Индии, и впоследствии расселился не только на юго-восток, в Австралию, но и на север, непосредственно в Центральную Азию, где и разделился на две ветви, одна из которых устремилась на запад, в Европу, а другая двинулась в Восточную Азию (см. главы 5 и 6)[198]. В отличие от своего сына, клана M17, Руслан не остановился на пороге Восточной Европы. Правильнее будет сказать, что на западе Европы его встречаемость очень высока, достигая 86% в Стране Басков и почти столь же высокого уровня — на Британских островах. Это расселение «отца» и «сына» в разных направлениях свидетельствует о том, что первый мог проникнуть в Европу гораздо раньше последнего. Как мы сможем убедиться в главе 6, это можно объяснить влиянием Ледникового периода. А пока ограничимся тем, что скажем, что, хотя М17 в Европе можно считать сравнительным «новичком», его «отец», Руслан, по-видимому, является наиболее мощной мужской линией-маркером древнейшего проникновения в Европу с востока еще в эпоху Раннего Верхнего палеолита, около 30 тысяч лет тому назад. Итак, на долю этой команды, «отца» и «сына», приходится примерно 50% всех существующих мужских генеалогических линий современной Европы[199].
Азиатские корни европейцев
Наше путешествие во времени в глубины генетической и археологической истории человечества позволяет сделать два важнейших вывода: во-первых, генетической прародиной европейцев первоначально, около 50 тысяч лет тому назад, была Южная Азия, точнее — Пакистан и регион Персидского залива; и во-вторых, предки современных европейцев проникли в Европу, этот прохладный, но богатый сад, по двум совершенно разным маршрутам. Более ранний из этих маршрутов пролегал через Плодородный Полумесяц, который около 51 тысячи лет тому назад был открыт в качестве своего рода коридора, ведущего от побережья Персидского залива. Коридор этот позволил мигрантам проникнуть в Турцию, Болгарию и далее, в Южную Европу. Это вполне совпадает с общим проникновением представителей ауриньякской культуры в Европу. Второй по времени маршрут из Южной Азии в Европу мог пролегать вверх по течению Инда, в Кашмир, и далее, в Центральную Азию, где примерно около 40 тысяч лет тому назад охотники начали охотиться даже на таких огромных животных, как мамонты. Некоторые из этих охотников, обладавшие передовыми по тем временам техническими навыками и орудиями, могли мигрировать на запад, за Урал, в европейскую Россию, а затем — на территорию современных Чехии и Германии. Более консервативная точка зрения на это вторжение с востока сводится к тому, что наиболее ранним маршрутом проникновения человека современного типа на территорию России был путь через Закавказье, а не через Центральную Азию.
ГЛАВА ЧЕТВЕРТАЯ ПЕРВЫЕ ШАГИ В АЗИЮ, ПЕРВЫЙ ПРЫЖОК В АВСТРАЛИЮ
 Фото. Едва различимый, выполненный блекло-пурпурным цветом контур слева от колпачка объектива — древнейший датируемый наскальный рисунок фигуры человека. Его возраст — 17 тыс. лег. Окрестности Кимберли, Австралия.
Фото. Едва различимый, выполненный блекло-пурпурным цветом контур слева от колпачка объектива — древнейший датируемый наскальный рисунок фигуры человека. Его возраст — 17 тыс. лег. Окрестности Кимберли, Австралия.
В главе 1 мы уже знакомились с доказательствами того, что в древности исход из Африки совершила одна генетическая линия, L3, две дочерних линии которой, Манью и Насрин, совместными усилиями заселили все остальные просторы земного шара. Эта единичная генетическая линия имеет ключевое значение для обоснования гипотезы об одном-единственном исходе, совершенном по южному маршруту. Как мы видели в главе 3, тот удивительный факт, что европейцы являются потомками только одного из этих кланов — клана Насрин, позволяет определить локус возникновения этой ветви, появившейся в одной из колоний мигрантов в самом начале пути из Африки, по всей вероятности — где-то в районе Персидского залива. Полное отсутствие каких-либо следов дочерних генетических ветвей Манью и Насрин в Северной Африке и на Леванте и обилие их в Индии исключает возможность северного маршрута миграции в Европу и служит неопровержимым аргументом в пользу версии о южном маршруте исхода из Африки, пролегавшем через устье Красного моря. А теперь давайте обратимся к более детальному рассмотрению генетических и иных свидетельств самой ранней волны колонизации Южной Азии и побережья Индийского океана. С этого момента история Y-хромосомы становится более полной, и в ней появляются некоторые другие генетические маркеры, помимо генов Адама и Евы. К Южной Азии относятся все те страны, которые встретились мигрантам-первопроходцам, кормившимся собирательством на прибрежной полосе, на их пути вдоль северного побережья Индийского океана.
Выжившие в долгом пути: место и время
Если у всех людей, живущих ныне за пределами Африки, был один общий предок, то дата исхода предков аборигенов Австралии будет абсолютно тождественной времени исхода предков европейцев, индусов и китайцев. Более того, все эти многообразные линии должны сходиться в некой общей точке в пространстве и времени, и все поселения, основанные мигрантами на путях скитаний, должны содержать генетические и даже материально-культурные следы, позволяющие определить, кто их оставил. Так, проанализировав пробы мтДНК или Y-хромосомы у любого жителя нашей планеты за пределами Африки, мы можем отыскать их место на соответствующей ветви генетического древа выходцев из Африки. И то, какое место они занимают на ветви древа выходцев из Африки, часто может показать нам, как они попали туда, а порой даже сказать, когда это произошло. Нам не следует поддаваться соблазну делать поспешные выводы, основанные на изолированных генетических данных. Истинная структура ветвей и их географическое распределение часто являются куда более красноречивыми, чем данные по ним, полученные при помощи молекулярных часов[200]. Эти данные носят весьма приближенный характер, и поэтому необходимо брать их средневзвешенные значения с учетом других источников информации, а именно — эволюции климатических факторов, которые показывают, когда конкретно могли сложиться благоприятные для миграции условия. Еще одним источником сведений служат археологические свидетельства, разумеется, если таковые имеются. Археологические свидетельства о миграции по прибрежной полосе неизбежно являются крайне приблизительными. В период между 60 и 80 тысячами лет тому назад колебания уровня Мирового океана составляли от 50 до 80 м ниже современного уровня, так что древние береговые полосы и даже обширные материковые земли, по которым бродили наши далекие предки, сегодня находятся глубоко под водой. Однако существует целый ряд интереснейших исключений из этих эффектов затопления. На некоторых участках побережья Индийского океана нам до сих пор встречаются небольшие поселения так называемых туземных этнических групп, которые вполне могут оказаться прямыми потомками тех самых бродячих собирателей. Еще задолго до появления методов изучения мтДНК главным основанием для выделения этих туземных групп был тот факт, что их культура и внешний облик резко отличались от основной массы населения в прилегающих районах. Некоторые из этих туземных групп имели черты, общие с африканцами, например, курчавые волосы и очень темную кожу. В этой и следующей главах я намерен рассмотреть более объективные генетические и физические свидетельства в пользу этой гипотезы, но для начала приведу названия некоторых из этих групп. Представителей таких туземных групп часто обозначают такими противоречивыми и предвзятыми терминами, как «австралоиды», «негроиды» и «негритосы», чтобы подчеркнуть их отличие от жителей прилегающих районов. К числу таких групп — начиная с побережья Южной Аравии — относится племя хадрамаут, представителей которого часто описывают как австралоидов, но которые, однако, отличаются куда более выраженной примесью африканской крови. На побережье Пакистана, в районе устья Инда, найдены генетические и исторические свидетельства в пользу гипотезы о заметной — и к тому же очень поздней — примеси африканской крови в жилах местных жителей, что можно считать результатом работорговли и завоза в эти места рабов из Африки. Иначе обстоит дело с другими, так называемыми негроидными, туземными группами, сохранившимися в Индии, в частности — народностями кадар и паньян. В той же Южной Азии существует целый ряд других туземных групп, включая так называемые прото-австралоидные этнические группы, такие, как корава, йанади, ирула, гадаба и ченьчу в Индии, а также ведды в Шри-Ланке. Недавнее исследование, посвященное двум из этих групп, ченьчу и койя, позволило выявить не только тот факт, что мтДНК и Y-хромосомные наборы их предков являются уникальными среди других народов Южной и Западной Азии, но и то, что они характерны для древнейшего генетического наследия всего этого региона. По всей видимости, их предки-собиратели, мигрировавшие из Африки, образно говоря, заложили генетические семена для последующей дифференциации генофондов Западной и Восточной Евразии, и с тех пор представители этих народов испытали крайне ограниченный приток генов извне[201]. На Андаманских островах, расположенных в Бенгальском заливе, между Индией и Юго-Восточной Азией, живет целый ряд таких туземных групп, сохранивших архаические дописьменные культуры и обладающих резко выраженными чертами внешности так называемого африканского типа. К числу групп, менее всего затронутых внешним влиянием, относятся народности джарава и онге, представители которых живут на большинстве южных островов Андаманского архипелага. Результаты недавних генетических исследований, находящиеся в процессе анализа и изучения, способны помочь выяснить происхождение жителей этих островов как по материнской, так и по отцовской линиям. С материнской стороны онге и джарава имеют две характерные митохондриевые группы, которые позволяют безоговорочно отнести их к суперклану Манью, что можно считать аргументом в пользу версии об одном-единственном исходе (миграции) из Африки. Эти генетические линии демонстрируют непосредственную взаимосвязь с основными ветвями двух древних и в своем роде уникальных кланов Манью в Индии — М2 и М4. М2 — это наиболее древняя и своеобычная группа клана Манью; кроме того, она имеет наиболее общие компоненты мтДНК среди всех вышеупомянутых туземных групп Индии. С отцовской стороны онге и джарава относятся только к клану Авель (наиболее редко встречающемуся из трех линий Y-хромосом выходцев из Африки и известному под общим названием группы D или азиатского YAP). Это вполне согласуется с точкой зрения о том, что онге и джарава остаются изолированными группами еще с тех самых пор, когда их предки, кормившиеся собирательством на прибрежной полосе, бродили вдоль побережья Индийского океана. Другие туземные группы на этих островах, именуемых Большими Андаманскими, также имеют некоторые отличия и в культуре, и во внешнем облике. С генетической точки зрения, поскольку они принадлежат к ветвям тех же уникальных генетических линий клана Манью, преобладающая часть их Y-хромосом восходит к Сифу, одному из трех сыновей Адама выходцев из Африки [202]. Еще далее на пути древней миграции нас ждет встреча с так называемыми негритосами — коренными жителями Малайского полуострова, которых обычно называют народностью семанг (см. Фото 14). Эти люди — по-видимому, наиболее реальные претенденты на титул потомков древних собирателей на прибрежной полосе. Другая туземная группа, вполне возможно, являющаяся потомками тех же собирателей в Индокитае и на Малайском полуострове — это так называемые туземные малайцы, которые с точки зрения физического облика занимают промежуточное положение между народностью семанг и монголоидами (см. главу 5). Самое поразительное здесь заключается в том, что они являются своего рода хранителями ключей к древнейшим ветвям линии мтДНК, характерной для монголоидного населения этого региона и известной как клан F. Эта материнская линия npe-F, родоначальница многих линий в Восточной Азии, присутствующая у туземных малайцев, является у них общей с другими архаическими группами, обитающими на других островах Больших Андаманских островов, о которых мы уже говорили выше, а также с жителями Никобарских островов, расположенных к югу от Андаманских[203].
 Фото 14. Автор этой книги проводит генетические обследования у жителей Семанга, которые, весьма вероятно, являются реликтовыми потомками народов первого маршрута собирателей на прибрежной полосе, совершивших исход из Африки.
Фото 14. Автор этой книги проводит генетические обследования у жителей Семанга, которые, весьма вероятно, являются реликтовыми потомками народов первого маршрута собирателей на прибрежной полосе, совершивших исход из Африки.
В этой и следующей главах я расскажу более подробно о различных народах и поселениях, оставленных мигрантами — участниками первого великого переселения вдоль береговой линии Индийского океана. Как всегда, главное здесь заключается в том, что генетическое древо говорит нам: это переселение отнюдь не было ранним исходом из Африки, как то полагают некоторые археологи и палеонтологи. Это было продвижение передовых групп на пути заселения всего Старого Света. Самое забавное заключается в том, что некоторые (если не сказать — единственные) наиболее эффектные археологические свидетельства, которые можно использовать для датировки этой миграции собирателей вдоль побережья Индийского океана, обнаружены не в Индии, не в Южной Аравии или в Африке, а почти в самом конце этого долгого пути — на Малайском полуострове, на о. Новая Гвинея и в Австралии. Поэтому мы отправимся сначала в гости к тем, кто проделал наиболее долгий путь вокруг побережья Индийского океана, — к аборигенам Новой Гвинеи и Австралии (см. Фото 15).
 Фото 15. Лица нашего времени (сверху, слева направо): бабушка-китаянка с внуком, мальчик с нагорий Новой Гвинеи, англичанин и австралийский абориген из земли Арнхема.
Фото 15. Лица нашего времени (сверху, слева направо): бабушка-китаянка с внуком, мальчик с нагорий Новой Гвинеи, англичанин и австралийский абориген из земли Арнхема.
Первые австралийцы
Пожалуй, из всех проблем, связанных с археологией этого региона, самые жаркие споры в научных кругах вызывал вопрос о том, какие именно племена первыми проникли в Австралию и когда это произошло. Вопрос «какие», звучащий вызовом мультирегионалистам, которые настойчиво повторяют свои аргументы о региональных различиях в форме черепов древних людей (см. главу 1), выглядит все менее и менее актуальным перед лицом массы генетических свидетельств того, что все современные туземные жители Австралии и Новой Гвинеи принадлежат кланам Насрин или Манью[204]. Другими словами, они вовсе не являются каким-то изолированным исключением и тем более людьми более архаического типа или же потомками от браков людей современного типа с местными представителями вида Homo erectus. Однако эти неоспоримые свидетельства не мешают ретроградам-мультирегионалистам типа Шпильберга предпринимать все новые и новые попытки отстоять свои устаревшие взгляды, используя для этого древнейшие пробы ДНК. Тем не менее сегодня наука располагает свидетельствами, полученными на основе проб генетического материала той же самой ветви L3 потомков выходцев из Африки (более подробно о дискуссии, посвященной новейшим исследованиям в этой проблемной области см. примечание[205]). Зато вопрос «когда» привлекает все большее внимание. Историки просто обожают нашпиливать события на даты вместо того, чтобы довольствоваться простой хронологической последовательностью, так что мне придется сначала поговорить о датах, хотя этот вопрос уместнее было бы затронуть в последнюю очередь. Наиболее ранние общепризнанные археологические свидетельства расселения людей современного типа за пределами Африки вплоть до недавнего времени были найдены только в Австралии. Однако ситуация в этой области меняется на редкость быстро. До начала 1990-х гг. ученые не располагали доказательствами присутствия человека в Австралии и Новой Гвинее в эпоху ранее 40 тысяч лет тому назад. И вот теперь оказалось, что эти датировки, как и датировки артефактов в Европе, были обусловлены ограниченными возможностями радиоуглеродного метода. Однако теперь появились новые методы. Один из них, так называемый метод датировки по люминесценции кремния, позволил исследователям заглянуть гораздо дальше рубежной для радиоуглеродного метода даты 40 тысяч лет назад. В 1990 г. австралийские геологи и археологи Ричард Роберт и Рис Джонс, исследуя возраст первого поселения в пещерах на побережье Земли Арнхема в Северной Австралии, получили цифры от 50 до 60 тысяч лет тому назад . Земля Арнхема расположена прямо напротив острова Тимор — ближайшего к Австралии острова Индонезийского архипелага и, таким образом, наиболее вероятной отправной точки для первых мигрантов, направлявшихся в Австралию[206]. После этого датировки австралийских артефактов приняли весьма драматический характер и, образно говоря, пробили брешь в стене. В 1996 г. археолог Роберт Фаллагар проводил исследования наскальных рисунков на стоянке Джинмиум. Он обнаружил, что стена пещеры была сплошь покрыта плюсками[207], и, как сообщает Фаллгар, возраст обнаруженных в отложениях выкрошившихся фрагментов песчаника, испещренного древними рисунками, согласно оценке по двум независимым методам датировки может составлять 50 и 73 тысячи лет. Более того, ученый обнаружил даже каменные артефакты в культурных слоях, датируемых по методу термолюминесценции эпохой между 116 и 176 тысячами лет, что позволяет считать их наиболее ранними следами деятельности человека на Австралийском континенте, в два или даже три раза более древними, чем уже упоминавшиеся находки в пещерах на Земле Арнхема. Однако эта проблема была практически снята после того, как было доказано, что контаминация (загрязнение)песчинками из камней, обвалившихся со сводов, дала датировки совершенно фантастической древности для слоя земли вокруг этих артефактов. Другими словами, эти датировки были недостоверными, и анализ отдельных песчинок помог выявить и устранить неточности, связанные с контаминацией. Таким образом, возрастнаиболее ранних следов деятельности человека в Австралии остался прежним — около 60 тысяч лет[208]. Впрочем, недавно были опубликованы работы, где приводятся близкие датировки следов деятельности человека: 62 тысячи лет тому назад. Эти датировки основаны на применении различных методов для анализа почвы вокруг скелета стройного анатомически современного человека, найденного неподалеку от озера Мунго в районе Вилландровых озер на юго-востоке Австралии. В одной из этих работ говорится, что для того, чтобы иметь столь почтенный возраст, первые обитатели Австралии должны были попасть на континент в глубокой древности, когда уровень океана был гораздо более низким. Подобная возможность проникнуть в Австралию благодаря низкому уровню океана могла предоставиться около 70 тысяч лет тому назад, что свидетельствует о том, что анатомически современные люди, обитавшие на берегах озера Мунго, были в числе древнейших обитателей континента, поскольку предыдущий столь же низкий уровень океана имел место на 80 тысяч лет раньше, то есть около 150 тысяч лет тому назад, еще до появления человека современного вида[209].
Одна-единственная возможность в истории
Пока ученые выдвигают всевозможные аргументы и контраргументы, мы с полным основанием можем говорить, что датировки находок на многих стоянках близки или совпадают, указывая, что человек современного типа уже присутствовал в Австралии примерно 60 тысяч лет тому назад. Примерно в то же время закончился период очень низкого уровня Мирового океана, когда огромные массы воды были сосредоточены в ледниковых полярных шапках и уровень моря у берегов Австралии был примерно на 80 м ниже современного. И хотя считается, что точная дата наиболее низкого уровня океана — около 70 тысячи лет тому назад, уровень Мирового океана оставался очень низким вплоть до 65 тысячи лет назад, после чего начался его быстрый подъем[210]. Присутствие человека на северо-западном побережье Австралии 62 тысячи лет тому назад не вполне согласуется с реальными условиями для его прибытия морским путем, поскольку период низкого уровня Мирового океана завершился примерно за 3000 лет до этого. Таким образом, если дата прибытия человека в Австралию, равная 62 тысячам лет тому назад, реальна, это означает, что мигрантам пришлось столкнуться с рядом проблем. Крайне низкий уровень океана, имевший место несколько ранее, делал шансы мигрантов перебраться с Тимора в Австралию несравненно более высокими. На контурной карте глубины моря у северного побережья Австралии мы видим, что расстояние, разделявшее Австралию и Тимор, увеличилось со 170 км при уровне океана —80 м и 220 км при уровне —40 м до 470 км при уровне —20 м (см. рис. 4.1). Различие между 170 км и 220 км на первый взгляд представляется не слишком существенным, но дело в том, что за последние 20 тысяч лет расстояние 180 км было абсолютным пределом досягаемости для плаваний между островами во всем мире. Древнейшим свидетельством этой максимальной дальности плавания, которая подтверждается археологическими находками, можно считать заселение острова Манус, расположенного к северу от Новой Гвинеи. Это заселение имело место около 20 тысяч лет тому назад. Таким образом, если говорить о заселении Австралии в результате длительного плавания, то единственная реальная возможность для подобного плавания могла предоставиться людям где-то между 65 и 70 тысячами лет тому назад. Тогда они могли успеть создать те самые памятники древнего искусства, которые обнаружены археологами в Австралии и датируются временем между 55 и 62 тысячами лет тому назад. В любом случае заселение Австралии в результате случайного дрейфа представляется крайне маловероятным сценарием, если принять во внимание генетические свидетельства присутствия в Австралии многочисленных материнских линий-родоначальниц. Подобная дата появления человека на континенте вполне согласуется с генетической датировкой расселения мигрантов в Австралии, составляющей 68 тысяч лет тому назад[211].
 Рис. 4.1
Нам представляется вполне уместным перенести свидетельства деятельности человека, датируемые временем около 60 тысяч лет тому назад, в другой контекст. Дело в том, что на некоторых древнейших стоянках человека в Австралии найдены характерные груды раковин морских моллюсков. Это дает основание предполагать, что первые люди, прибывшие на северо-западное побережье Австралии, по-прежнему кормились собирательством на прибрежной полосе. А если это так, то у нас нет практически никакой возможности найти следы их первоначального присутствия на берегу океана, так как Тиморское море сегодня продвинулось на добрых 500 км в глубь прежнего материка, накрыв своими водами древнейшие поселения человека на берегу. Первые реально датированные свидетельства присутствия человека на северо-западе Австралии после периода минимально низкого уровня океана, естественно, были получены на горизонтах, расположенных выше высокого уровня океана. Подобный высокий уровень отмечался уже между 55 и 60 тысячами лет тому назад, когда уровень океана был ниже современного всего на 20 м, а береговая линия была уже близка к своим современным очертаниям. Береговая линия поблизости от пещер на Земле Арнхема в эту эпоху хорошо согласуется с расчетами предполагаемого уровня океана и датировкой следов деятельности человека в этих местах. Эта датировка, выполненная геологом Ричардом Робертом, дает цифры около 50—60 тысяч лет тому назад. При подобной, обусловленной изменением уровня океана, паузе между заселением и археологическими свидетельствами о деятельности человека следующий минимум уровня океана, отмечавшийся около 55 тысяч лет тому назад, был уже слишком запоздалым для трансокеанских плаваний и археологически документированных данных о присутствии человека в эпоху ранее 50 тысяч лет тому назад[212].
Хотя заселение ближайшего соседа Австралии, огромного острова Новая Гвинея, по всей видимости, представляло такие же проблемы для потенциальных колонистов, как и заселение Австралии, между ними все же были существенные различия. Есть вполне реальная вероятность того, что Новая Гвинея была заселена даже раньше, чем Австралия. Прежде всего возможны два пунктирных маршрута через острова Восточной Индонезии (см. рис. 4.1). Как мы уже знаем, для того чтобы плавание по прямому пути в Австралию с острова Тимор, через Тиморское море, имело шансы на успех, уровень океана в ту эпоху должен был быть предельно низким. Между тем маршрут в Новую Гвинею через о. Сулавеси и Молуккские острова позволял преодолевать гораздо меньшие расстояния между островами и практически не зависел от колебаний уровня океана.
Все острова на пути в Австралию и Новую Гвинею находились в пределах прямой видимости друг от друга. У этих островов издревле существует очень крутая береговая линия, и маршрут путешествия на Новую Гвинею включал в себя очень мало участков, пролегавших по континентальному шельфу. В результате перепады уровня океана почти не сказывались на расстоянии между островами, и этот маршрут можно считать одинаково легким (или трудным) в любую эпоху после того, как наши предки прибыли на о. Борнео сухопутным путем[213].
Можно задать вопрос: имеются ли доказательства того, что предки современных аборигенов Австралии не воспользовались этим альтернативным северным, немногим более протяженным маршрутом, ибо в ту эпоху Австралия могла быть соединена сухопутным перешейком с Новой Гвинеей? Генетические данные со всей определенностью показывают, что между аборигенами Австралии и Новой Гвинеи не существует общих генеалогических клановых линий и что жители Новой Гвинеи в генетическом отношении столь же далеки от аборигенов Австралии, как и от любого другого народа за пределами Африки Более того, оценки времени расселения туземных кланов мтДНК Новой Гвинеи дают цифру 77 тысяч лет тому назад, что значительно старше периода активного расселения австралийцев. Это говорит о том, что Новая Гвинея, по всей вероятности, могла быть заселена людьми гораздо раньше Австралии, а также о том, что австралийские генетические кланы не являются потомками кланов Новой Гвинеи. Другими словами, первые мигранты в Австралию могли отдать предпочтение маршруту от одного необитаемого острова к другому, хотя это и создавало ряд трудностей[214].
Однако при всей своей заманчивости эта генетическая история, хорошо согласующаяся с исключительной древностью и взаимосвязью между колонизацией Австралии и Новой Гвинеи, не согласуется с надежно датируемыми археологическими находками, сделанными на островах Восточной Индонезии или в Новой Гвинее. Вплоть до недавнего времени ученые не располагали свидетельствами о присутствии человека на всем протяжении обоих этих трансостровных маршрутов через Восточную Индонезию около 40 тысяч лет назад, не говоря уж о 70 тысячах лет назад. Однако новые поразительные находки на острове Флорес, одном из самых крупных островов на южном маршруте, способны в корне изменить сложившиеся представления. Глубокая (около 7 м) яма в пещере Лянг Буа, находящейся на острове Флорес, таила в себе удивительные находки, возраст которых — около 840 тысяч лет! Лянг Буа может сыграть значительную роль в качестве источника Ценнейших культурных и палеонтологических свидетельств, относящихся к большей части этого периода, включая и дату прибытия на остров человека современного типа. В ту эпоху в пещере обитали представители двух видов человека. Первые жители явно не были людьми современного типа. В настоящее время даты появления первых людей современного типа уточняются, однако они могут оказаться наиболее ценными для датировки наиболее раннего присутствия человека современного типа в Восточной Азии[215].
Таким образом, если, следуя гипотезе о климатическом «окне», допустить, что фактически колонизация Австралии имела место в период между 65 и 70 тысячами лет тому назад, а острова Флоры и даже Новая Гвинея были заселены около 75 тысяч лет тому назад, мы вправе задать вопрос: а за сколько тысяч лет до этого момента люди современного типа должны были покинуть Африку, чтобы проделать столь дальний путь вдоль побережья южных морей. Понятно, что ответить на такой вопрос крайне сложно. Прежде всего, мы не можем с уверенностью сказать, сколько времени могло потребоваться нашим собирателям, чтобы в буквальном смысле «проесть» себе путь к берегам Тимора или Сулавеси, если древние люди не оставили на нем никаких следов своего продвижения. Как показывают данные оценки уровня океана в ту эпоху, прибрежная полоса, по которой брели наши далекие предки 60—80 тысяч лет тому назад, давно ушла под воду, и нам трудно рассчитывать найти следы древних миграций, не опустившись на дно морское.
Рис. 4.1
Нам представляется вполне уместным перенести свидетельства деятельности человека, датируемые временем около 60 тысяч лет тому назад, в другой контекст. Дело в том, что на некоторых древнейших стоянках человека в Австралии найдены характерные груды раковин морских моллюсков. Это дает основание предполагать, что первые люди, прибывшие на северо-западное побережье Австралии, по-прежнему кормились собирательством на прибрежной полосе. А если это так, то у нас нет практически никакой возможности найти следы их первоначального присутствия на берегу океана, так как Тиморское море сегодня продвинулось на добрых 500 км в глубь прежнего материка, накрыв своими водами древнейшие поселения человека на берегу. Первые реально датированные свидетельства присутствия человека на северо-западе Австралии после периода минимально низкого уровня океана, естественно, были получены на горизонтах, расположенных выше высокого уровня океана. Подобный высокий уровень отмечался уже между 55 и 60 тысячами лет тому назад, когда уровень океана был ниже современного всего на 20 м, а береговая линия была уже близка к своим современным очертаниям. Береговая линия поблизости от пещер на Земле Арнхема в эту эпоху хорошо согласуется с расчетами предполагаемого уровня океана и датировкой следов деятельности человека в этих местах. Эта датировка, выполненная геологом Ричардом Робертом, дает цифры около 50—60 тысяч лет тому назад. При подобной, обусловленной изменением уровня океана, паузе между заселением и археологическими свидетельствами о деятельности человека следующий минимум уровня океана, отмечавшийся около 55 тысяч лет тому назад, был уже слишком запоздалым для трансокеанских плаваний и археологически документированных данных о присутствии человека в эпоху ранее 50 тысяч лет тому назад[212].
Хотя заселение ближайшего соседа Австралии, огромного острова Новая Гвинея, по всей видимости, представляло такие же проблемы для потенциальных колонистов, как и заселение Австралии, между ними все же были существенные различия. Есть вполне реальная вероятность того, что Новая Гвинея была заселена даже раньше, чем Австралия. Прежде всего возможны два пунктирных маршрута через острова Восточной Индонезии (см. рис. 4.1). Как мы уже знаем, для того чтобы плавание по прямому пути в Австралию с острова Тимор, через Тиморское море, имело шансы на успех, уровень океана в ту эпоху должен был быть предельно низким. Между тем маршрут в Новую Гвинею через о. Сулавеси и Молуккские острова позволял преодолевать гораздо меньшие расстояния между островами и практически не зависел от колебаний уровня океана.
Все острова на пути в Австралию и Новую Гвинею находились в пределах прямой видимости друг от друга. У этих островов издревле существует очень крутая береговая линия, и маршрут путешествия на Новую Гвинею включал в себя очень мало участков, пролегавших по континентальному шельфу. В результате перепады уровня океана почти не сказывались на расстоянии между островами, и этот маршрут можно считать одинаково легким (или трудным) в любую эпоху после того, как наши предки прибыли на о. Борнео сухопутным путем[213].
Можно задать вопрос: имеются ли доказательства того, что предки современных аборигенов Австралии не воспользовались этим альтернативным северным, немногим более протяженным маршрутом, ибо в ту эпоху Австралия могла быть соединена сухопутным перешейком с Новой Гвинеей? Генетические данные со всей определенностью показывают, что между аборигенами Австралии и Новой Гвинеи не существует общих генеалогических клановых линий и что жители Новой Гвинеи в генетическом отношении столь же далеки от аборигенов Австралии, как и от любого другого народа за пределами Африки Более того, оценки времени расселения туземных кланов мтДНК Новой Гвинеи дают цифру 77 тысяч лет тому назад, что значительно старше периода активного расселения австралийцев. Это говорит о том, что Новая Гвинея, по всей вероятности, могла быть заселена людьми гораздо раньше Австралии, а также о том, что австралийские генетические кланы не являются потомками кланов Новой Гвинеи. Другими словами, первые мигранты в Австралию могли отдать предпочтение маршруту от одного необитаемого острова к другому, хотя это и создавало ряд трудностей[214].
Однако при всей своей заманчивости эта генетическая история, хорошо согласующаяся с исключительной древностью и взаимосвязью между колонизацией Австралии и Новой Гвинеи, не согласуется с надежно датируемыми археологическими находками, сделанными на островах Восточной Индонезии или в Новой Гвинее. Вплоть до недавнего времени ученые не располагали свидетельствами о присутствии человека на всем протяжении обоих этих трансостровных маршрутов через Восточную Индонезию около 40 тысяч лет назад, не говоря уж о 70 тысячах лет назад. Однако новые поразительные находки на острове Флорес, одном из самых крупных островов на южном маршруте, способны в корне изменить сложившиеся представления. Глубокая (около 7 м) яма в пещере Лянг Буа, находящейся на острове Флорес, таила в себе удивительные находки, возраст которых — около 840 тысяч лет! Лянг Буа может сыграть значительную роль в качестве источника Ценнейших культурных и палеонтологических свидетельств, относящихся к большей части этого периода, включая и дату прибытия на остров человека современного типа. В ту эпоху в пещере обитали представители двух видов человека. Первые жители явно не были людьми современного типа. В настоящее время даты появления первых людей современного типа уточняются, однако они могут оказаться наиболее ценными для датировки наиболее раннего присутствия человека современного типа в Восточной Азии[215].
Таким образом, если, следуя гипотезе о климатическом «окне», допустить, что фактически колонизация Австралии имела место в период между 65 и 70 тысячами лет тому назад, а острова Флоры и даже Новая Гвинея были заселены около 75 тысяч лет тому назад, мы вправе задать вопрос: а за сколько тысяч лет до этого момента люди современного типа должны были покинуть Африку, чтобы проделать столь дальний путь вдоль побережья южных морей. Понятно, что ответить на такой вопрос крайне сложно. Прежде всего, мы не можем с уверенностью сказать, сколько времени могло потребоваться нашим собирателям, чтобы в буквальном смысле «проесть» себе путь к берегам Тимора или Сулавеси, если древние люди не оставили на нем никаких следов своего продвижения. Как показывают данные оценки уровня океана в ту эпоху, прибрежная полоса, по которой брели наши далекие предки 60—80 тысяч лет тому назад, давно ушла под воду, и нам трудно рассчитывать найти следы древних миграций, не опустившись на дно морское.
Древнейшие реликты маршрута собирателей
Существует и другая проблема. Если наши праотцы покинули Африку более 70 тысяч лет тому назад, они вполне могли использовать и создавать каменные орудия старого типа, аналогичные тем, которыми пользовались представители архаического вида Homo sapiens, которые, вполне возможно, покинули Африку по меньшей мере 100 тысяч лет тому назад (см. Пролог). Такие орудия эпохи Среднего палеолита во множестве встречаются в Индии и датируются временем около 160 тысяч лет тому назад, поэтому они не могут считаться надежными маркерами прихода людей анатомически современного типа Орудия эпохи Среднего палеолита в изобилии встречаются в песках береговой полосы южного побережья Аравийского полуострова, однако они не поддаются сколько-нибудь надежной датировке. Йемен, являющийся, по моему убеждению, пунктом прибытия мигрантов после исхода из Африки, пока что не привлек к себе достаточного внимания археологов, однако там тоже были найдены каменные орудия, аналогичные африканским орудиям позднего Среднего Каменного века[216]. Археологическая датировка ранних находок в Австралии и свидетельства еще более ранней колонизации Новой Гвинеи получили серьезную поддержку в результате открытия палеолитической культуры Кота Тампан в долине Ленггонг, что в Пераке на Малайском полуострове, на расстоянии примерно 2/3 пути из Африки в Австралию. Вначале эта культура была идентифицирована по находкам крупных, необычной формы и достаточно грубо сделанных каменных орудий, обработанных только с одной стороны, которые, как считали археологи в 1960-е гг., были делом рук представителей более ранних видов человека. Но когда была проведена переоценка возраста геологических слоев, в которых были найдены эти орудия, стало ясно, что они относятся к гораздо более позднему времени. Более заметный интерес к этим находкам возник в 1975 г., когда Том Харрисон, колоритный куратор Саравакского музея, попытался связать эти находки с грандиозным извержением вулкана Тоба на о.Суматра. Его предположение, основанное на исследовании вулканического пепла вокруг найденных на стоянке орудий, вызвало настоящую сенсацию, лихорадочный интерес к которой пошел на спад лишь совсем недавно[217]. Пожалуй, никто из ученых не отдал изучению культуры Кота Тампан и долины Ленггонг столько сил и времени, как археолог Зурайна Маджид из университета Сайнс Малайзия в Пенанге. Ее обширные исследования целого ряда стоянок этой культуры в долине Ленггонг показали, что местная культура каменных орудий, вполне возможно, просуществовала со дня грандиозного извержения вулкана Тоба до 7 или даже 4 тысяч лет тому назад[218]. А если это так, то находки артефактов этой культуры могут дать ответ на один из самых волнующих вопросов, касающихся этих странных орудий овальной формы: кто были их создатели? Дело в том, что эти орудия не идут ни в какое сравнение с гораздо более сложными артефактами. Орудия, обработанные гораздо тщательнее их, создавались в Африке и Европе людьми архаического типа гораздо раньше, так что с какой стати мы должны считать, что каменные сколы, найденные в вулканическом пепле, были сделаны людьми современного типа, жившими в эпоху извержения Тоба? Двое наиболее авторитетных специалистов по эпохе палеолита в Юго-Восточной Азии, австралийские археологи Питер Веллвуд и Сандра Боудлер, согласны с Зурайной Маджид и Томом Харрисоном в том, что эти орудия скорее всего были созданы людьми анатомически современного типа. Прежде всего надо сказать, что датировки большинства каменных орудий, найденных в долине Ленггонг, относятся к слишком позднему времени, чтобы их могли создать люди более архаического типа. Во-вторых, на Малайском полуострове, не говоря уж о долине Ленггонг, не найдено ни единого образца останков человека типа, предшествовавшего современному. Визитной карточкой Зурайны Маджид стала вызвавшая массу откликов находка так называемого перакского человека, сделанная членами ее группы в 1990 г. в пещере Гунуг Рунту, которая расположена в долине Ленггонг. Этот полный скелет человека современного типа, окруженный каменными орудиями этого класса, был описан специалистами как останки, имеющие характерные австрало-меланезийские признаки. Его возраст — около 10 тысяч лет. Поэтому недвусмысленная связь этих каменных орудий с человеком современного типа решительно опровергает утверждение о том, будто каменные орудия культуры Кота Тампан слишком грубы для того, чтобы считаться делом рук человека современного типа. Таким образом, по крайней мере — на сегодня, перакский человек бесспорно является наиболее убедительным свидетельством того, что орудия архаического типа, найденные в вулканическом пепле, были созданы человеком современного типа[219]. Другой уважаемый специалист в области археологии Юго-Восточной Азии — Ричард Шатлер. Он выразил более обобщенное мнение, что подобные орудия были впервые принесены на острова Юго-Восточной Азии людьми вида Homo sapiens примерно 70 тысяч лет тому назад. (Понятие «островная Юго-Восточная Азия», или ОЮВА, в отличие от континентальной Юго-Восточной Азии, включает в себя все острова Индонезии и Филиппин, за исключением Западной Новой Гвинеи, но включая все острова Борнео.) Шатлер предостерег против поспешного вывода о том, будто такие орудия являются отражением технической отсталости, соглашаясь с другими исследователями в том, что качество наличного сырья для производства орудий во многом определяло их уровень и что для изготовления более сложных орудий, например ножей, вполне мог использоваться бамбук[220]. Так каков же возраст пепла на стоянках культуры Кота Тампан? Когда он оценивался впервые, несколько десятилетий тому назад, тогдашние методы показали такую цифру: около 31 тысячи лет тому назад. Однако подобная датировка извержения вулкана Тоба вызывала возражения со стороны геологов и даже археологов, в частности Питера Беллвуда[221]. Главная проблема здесь заключалась в том, что в те времена у вулкана Тоба не было сколько-нибудь значительных извержений. Последнее крупное извержение вулкана Тоба, крупнейшее в мире за последние 2 млн. лет, произошло гораздо раньше, примерно 71—74 тысячи лет назад. Не так давно ряд видных геологов, включая и автора первоначальной датировки, пришли к согласию во мнении о том, что возраст пепла, в котором были найдены эти орудия, действительно составляет 74 тысячи лет. Эта датировка имеет огромную важность. Если каменные орудия культуры Кота Тампан были созданы людьми современного типа, то в таком случае они являются наиболее древними надежно датируемыми свидетельствами деятельности человека современного типа за пределами Африки. Таким образом, складывается впечатление, что предки аборигенов Австралии покинули Африку и, продвигаясь вдоль береговой полосы, прибыли в Малайзию задолго до катастрофического извержения Тоба. Но, пожалуй, фактором более важным, чем точность датировки, является то, что подобная взаимосвязь между каменными орудиями и вулканическим пеплом на стоянках в Малайзии позволяет утверждать, что древнейшие жители Индии и Пакистана тоже оказались непосредственно в зоне этой величайшей природной катастрофы в истории человечества. Извержение вулкана Тоба явилось самым грозным бедствием за последние 2 млн. лет. Подхваченное ветром громадное облако пепла, взметнувшееся над вулканом, двинулось на северо-запад и накрыло практически весь Индийский субконтинент. Даже в наши дни в этом регионе находят слои пепла толщиной в несколько метров, а в слое пепла на двух древних стоянках в Индии обнаружены орудия, датируемые эпохой Среднего и Верхнего палеолита[222]. Важный вывод, который можно сделать на основе этой взаимосвязи между орудиями и пеплом, заключается в том, что в результате извержения Тоба Восток и Запад разделила глубокая и генетически стерильная борозда. Население Индии было восстановлено путем волн повторной колонизации как с Востока, так и с Запада. И подобная борозда, как мы вскоре увидим, действительно существует на генетической карте Азии. Несмотря на близость вулкана Тоба к Пераку, облако вулканического пепла, взметнувшееся над ним, лишь вскользь задело Малайский полуостров. Люди культуры Кота Тампан оказались в числе несчастливцев, но большая часть полуострова осталась практически незатронутой извержением. Некоторые исследователи на основе сравнительного анализа морфологии черепов высказывают предположение, что туземные «негритосы» племени семанг, издревле кормившиеся охотой и собирательством, которые и сегодня живут в той же самой части густых влажных тропических лесов на севере Малайского полуострова, являются потомками той этнической ветви, к которой принадлежал и перакский человек. Непрерывный характер развития культуры Кота Тампан, версию о котором выдвинула Зурайна Маджид, позволяет провести прямую линию, ведущую к каменным орудиям 74-тысячелетней древности, обнаруженным в вулканическом пепле Тоба. В следующей главе мы рассмотрим материальные свидетельства и артефакты, связывающие нынешних людей племени семанг с древнейшими первопроходцами современного типа, мигрировавшими вдоль побережья Индийского океана[223]. В процессе съемок документального фильма «Истинная Ева», с которым перекликается наша книга, канал «Дискавери» осуществил финансирование исследования генетического наследия туземных групп Малайского полуострова. Это исследование я провел в тесном сотрудничестве с английским генетиком Мартином Ричардсом и рядом ученых из Малайзии. Это исследование явилось составной частью гораздо более обширной программы, осуществляемой генетиками ряда стран Азии. Результаты исследований мтДНК оказались впечатляющими: у 3/4 представителей этнической группы семанг (принадлежащий к различным типам «негритосов») были выявлены уникальные локальные варианты линий Манью и Насрин, почти не имеющие внешних примесей, что хорошо согласуется с мнением о том, что их предки могли прибыть в эти края вместе с первыми мигрантами — собирателями на прибрежной полосе. Найденные у них линии восходят непосредственно к самым корням Манью и Насрин (двух первых дочерей Евы за пределами Африки — см. главу 1). Выявленная у них линия Манью не встречается более ни у какой этнической группы в Юго-Восточной или Восточной Азии (и вообще ни у какого другого этноса) и, хотя она и подверглась активному генетическому дрейфу в результате сокращения численности ее носителей, она тем не менее сохранила достаточно широкое многообразие, что позволяет утверждать, что ее возраст составляет около 60 тысяч лет. Что касается их линии со стороны Насрин, R21, то она происходит непосредственно от Рохани, генетической дочери Насрин. Подобное отсутствие каких бы то ни было связей с другими этническими группами Евразии хорошо согласуется с гипотезой о том, что эти люди, прибывшие в эти края много тысячелетий назад, оставались в полной генетической изоляции в джунглях Малайского полуострова[224]. Есть и другие свидетельства из данного региона, позволяющие отнести появление анатомически современного человека на Дальнем Востоке ко времени около 70 тысяч лет тому назад. Это прежде всего знаменитый люцзяньский скелет, найденный на юге Китая. Люцзяньский скелет (см. Фото 16), от которого уцелели хорошо сохранившиеся череп и несколько костей, был найден в 1958 г. в пещере Тонтяньянь в Гуанси-Чжуаньском автономном районе[225]. Его обнаружили крестьяне, собиравшие гумус и удобрения. Нет никаких сомнений том, что этот скелет принадлежал человеку современного типа, но в вопросе о его возрасте сразу же возникли разногласия во мнениях.
 Фото 16. Люйцзяньский череп Китай) с толстокостными, но современными чертами. По данным уточненной датировки , его возраст составлят по меньшей мере 68 тыс. лет, что свидетельствует о весьма раннем распространении человека анатомически современного типа из Африки, имевшем место еще до извержения вулкана Тоба.
Фото 16. Люйцзяньский череп Китай) с толстокостными, но современными чертами. По данным уточненной датировки , его возраст составлят по меньшей мере 68 тыс. лет, что свидетельствует о весьма раннем распространении человека анатомически современного типа из Африки, имевшем место еще до извержения вулкана Тоба.
Есть данные о датировке по изотопам урана, согласно которой возраст скелета составляет около 67 тысяч лет, однако подобная датировка вызывала сомнения из-за ее несоответствия возрасту того надежно датируемого геологического слоя, в котором был обнаружен скелет. В декабре 2002 г. группа китайских исследователей во главе с геологом Шень Гуаньюнем сообщила о результатах проведенного ею повторного анализа стратиграфических параметров пещеры (а также нескольких соседних пещер) и датировки черепа. Ученые пришли к выводу, что череп следует отнести к эпохе между 70 и 130 тысячами лет тому назад и что он в любом случае не моложе 68 тысяч лет. Череп был найден в так называемой интрузивной брекчии — яме с грудой обломков и отходов вторичного происхождения, содержащей материальные следы различных эпох. Благодаря тому, что их статья об этих исследованиях была опубликована в журнале «Journal of Human Evolution», нижняя временная граница этого периода (68 тысяч лет) представляется вполне солидной датировкой, поскольку она основана на многочисленных датировках возраста каменных капов, закрывающих брекчию. (Каменный кап образуется, когда вода, стекающая со стен, приводит к осаждению кальцита на стене или полу.) Наиболее вероятный возраст этих капов — от 111 до 139 тысяч лет. Эти данные, полученные на основе изучения фрагментов капов и кальцитов в самой брекчии, представляются чрезмерно завышенными. Но главный вывод этих исследований заключается в том, что вполне реальная нижняя граница появления на Дальнем Востоке анатомически современного человека является дополнительным аргументом в пользу гипотезы о раннем исходе человека из Африки через устье Красного моря еще до извержения вулкана Тоба, имевшего место около 71—74 тысяч лет тому назад. Дело в том, что для того, чтобы попасть в Китай около 68 тысяч лет назад, предкам этих людей необходимо было проделать путь длиной в четверть окружности экватора[226]. Некоторые ученые по-прежнему убеждены в том, что аборигены Австралии представляют собой потомков миграции из Африки, более ранней, чем исход родоначальников будущих народов и рас Европы, Азии и Америки. Однако наши генетические следы говорят совсем иное. Ряд исследований присутствия генетических линий материнских кланов в Австралии показал, что все они принадлежат к двум крупнейшим неафриканским суперкланам — Манью и Насрин. А обширные исследования Y-хромосом позволили установить, что все мужские линии аборигенов Австралии относятся к тому же клану Адама выходцев из Африки, что и все прочие обитатели земного шара за пределами Африки, и принадлежат к линиям Сифа и Каина. Та же картина наблюдается и при анализе генетических маркеров, передаваемых через обоих родителей. Другими словами, вся совокупность генетических свидетельств с полной определенностью говорит о том, что австралийцы также являются потомками того самого единственного исхода из Африки. Логика такого подхода в сочетании с рассмотренными выше археологическими свидетельствами позволяет отнести дату прибытия людей современного типа на Малайский полуостров ко времени до 74 тысяч лет тому назад, а дату их появления в Австралии — до 70 тысяч лет тому назад. Это вполне согласуется с датой исхода из Африки, определяемой на основе следов собирательства на прибрежной полосе. Как мы уже говорили в главе 1, одним из мотивов исхода с африканского континента могла стать массовая гибель даров моря на всем побережье Красного моря, вызванная резким повышением солености его вод, начавшимся около 80 тысяч лет тому назад. По моим оценкам затрат времени на миграцию из Африки вдоль побережья Индийского океана, собирателям на прибрежной полосе могло потребоваться около 6000 лет, чтобы «проесть» себе путь вдоль побережья до Перака, и еще около 4000 лет, чтобы достичь берегов Австралии. Такие временные рамки вполне соответствуют паузе между исходом из Африки около 80 тысяч лет тому назад и появлением в Австралии около 70 тысяч лет тому назад. Эта первая дата хорошо согласуется с оценками времени исхода африканского комплекса L3 на основе датировки по молекулярным часам (см. также главу 1)[227].
Взглянуть в лицо матери — родоначальницы миллиарда семейств
Главный вопрос, не дающий покоя журналистам и кинематографистам после выхода в 1988 г. знаменитой статьи в «Newsweek» об «истории» митохондриевой Евы, заключается в том, как могли выглядеть она и ее потомки. Вполне возможно, что все они имели весьма и весьма непохожую внешность и отличались друг от друга даже больше, чем случайно взятые представители разных рас и народов, живущие ныне за пределами Африки. Дело в том, что они, по крайней мере, происходят от одного и того же древнейшего прапредка, и все имеющиеся свидетельства говорят в пользу того, что вариации в пределах индивидуальных групп даже в наши дни уменьшаются по мере удаления из Африки. Однако что касается различий между разными группами, например, китайцами, коренными жителями Америки и аборигенами Полинезии, Меланезии и Австралии, то они, наоборот, нарастают по мере удаления от Африки; таким образом, можно говорить о том, что различия между так называемыми расами по мере удаления от Африки усиливаются. А теперь позвольте мне взглянуть на вопрос с несколько иной стороны. Моя дражайшая теща, которая была китаянкой, однажды сказала мне (причем, что называется, на полном серьезе), что европейцы кажутся ей совершенно одинаковыми, так сказать, все на одно лицо. Но европейцы с полным правом могут сказать то же самое о самих китайцах. Таким образом, налицо явный парадокс, который необходимо решить, прежде чем продолжить составление карты генетической диаспоры за пределами Африки. Суть вопроса заключается в том, каким образом возникло это разделение. То, что на первый взгляд выглядит парадоксом, на самом деле является результатом многократного и неравномерного дробления групп мигрантов, сопровождавшегося активным генетическим дрейфом. Это можно объяснить следующим образом: по мере того как мигранты-переселенцы разделялись на все большее и большее количество ветвей, первоначальные генетические вариации между ними оказывались как бы сосредоточенными в пределах различных групп, предков будущих этносов, которые живут в наши дни на территории Евразии и Америки. За последние 50 тысяч лет члены этих разделившихся и изолированных групп диаспоры выходцев из Африки начали внешне более походить на своих сородичей, чем на представителей других групп. Так возникли различия между группами. И хотя впоследствии я попытаюсь доказать, что многие из подобных различий были не более чем эволюционно обусловленными проявлениями адаптации к новым условиям окружающей среды, некоторые из них выходят далеко за рамки случайного разлома этнического «пирога». Чем же их можно объяснить? Ответ заключается в том же самом процессе, который «сработал» во время исхода из Африки первой небольшой группы мигрантов. Давайте представим — как мы поступили в одной из предыдущих глав, — что многообразие представителей рода человеческого подобно мешочку с камешками разных пород. Предположим, на момент исхода из Африки, имевшего место около 75 тысяч лет тому назад, в мешочке было 10 тысяч камешков. После того как мигранты переправились через Красное море, многообразие резко сократилось, и в колонии-родоначальнице осталось всего 250 камешков. Подобное сокращение известно под названием «эффекта родоначальников». И если члены этой первой колонии жили на юге Аравии изолированной группой на протяжении многих тысяч лет, одни из «камешков» не оставили себе подобных и их линии пресеклись, а другие линии, наоборот, заняли доминирующее положение. В результате число камешков в мешочке уменьшилось еще больше. Как мы уже говорили в главе 1, это и есть генетический дрейф. В дальнейшем население колонии дробилось на неравные части. Когда численность группы стала слишком высокой и данный участок берега более не мог прокормить ее, от нее одна за другой отделялись небольшие группы первопроходцев, которые покидали обжитое побережье и направлялись в глубь материка, продвигаясь в основном вдоль русел рек. Эти небольшие группы несли с собой в мешочке еще меньшее число первоначальных генетических камешков мигрантов-выходцев из Африки. Затем они подвергались воздействию того же «эффекта родоначальников» и мигрировали дальше, продолжая генетический дрейф. А теперь позвольте задать вопрос: что является кроной генеалогического древа и что — его стволом? Были ли собиратели на прибрежной полосе всегда и везде большинством, а первопроходцы, продвигавшиеся вдоль русла рек, — меньшинством, или же основная масса мигрантов в какой-то момент устремлялась в глубь материка? Такой вопрос может показаться уделом академической схоластики, но если наши журналисты действительно хотят заглянуть в лицо людям современного типа, жившим как можно ближе к моменту исхода, вопрос, на который мы должны ответить, будет звучать так: какие именно представители неафриканских народов в наши дни сохранили более всего вариантов тех самых «камешков» из мешочка древнейших обитателей Африки?
Нуклеарные генетические маркеры: чтобы найти больше древних маркеров, нужно иметь больше камешков в мешочке
Чтобы продолжить рассмотрение вопроса о многообразии, мы не можем полагаться на одни только генетические маркеры нашей митохондриевой ДНК или Y-хромосомы. Если уж говорить о них, надо признать, что в отцовских и материнских линиях ныне живущих на земном шаре народов уцелело всего по одному камешку от каждого из их африканских прародителей. Как я уже говорил, именно этот факт является доказательством одного-единственного исхода из Африки (см. главу 1). Понятно, что у одной линии нет и не может быть многообразия. Нет, если мы действительно хотим сравнить генетическое многообразие наших предков-африканцев с наследием остального населения земного шара, нам необходимо найти такие типы генетических маркеров, которые сумели пережить пресловутый эффект родоначальников и генетический дрейф исхода, сохранив достаточно широкое многообразие и количественный состав представителей каждой линии. Поэтому нам придется заняться анализом остальных компонентов нашей генетической структуры, которые присутствуют в ядрах клеток нашего тела и не связаны с тем или иным конкретным полом. Приводимая мной аналогия с камешками, естественно, была нарочито упрощенной. Если обратиться к рассмотрению той на удивление малой части нашей ДНК, которая действительно несет некую полезную нагрузку, нетрудно заметить, что у каждого гена существует от одного до целой дюжины вариантов, несколько отличающихся друг от друга Такие варианты обычно могут быть объединены в составе семейного генного древа. Основание каждого генного древа и его нижние ветви обычно являются по преимуществу африканскими, тогда как верхние ветви и побеги, а иногда и некоторые из нижних ветвей встречаются в основном за пределами Африки. Кроме того, мы можем рассмотреть и нефункционирующие части нашей ДНК, где накапливается громадная масса совершенно бесполезной информации, которая, однако, может использоваться генетиками в качестве своего рода почтового ящика. Эти излишние массивы данных могут накапливаться до тех пор, пока они не начнут составлять около 10% общего объема нашей ДНК. В то же время отсутствие таких специфических массивов у того или иного человека означает, что он сохраняет большинство признаков своего родового или африканского типа[228]. Если мы посмотрим, у представителей каких именно этнических групп более всего сохранилось генетическое наследие наших далеких африканских предков, нам придется вернуться в Австралию и Новую Гвинею. Эти два обширных массива суши образуют остатки грандиозного и сегодня частично затопленного водами океана континентального шельфа материка Сахул, где сохранилось значительно больше первичных генетических типов африканских предков, чем в любом другом регионе мира за пределами Африки[229]. Однако в наши дни аборигены Новой Гвинеи и Австралии мало походят друг на друга, и сегодня единственной крупной этнической группой, по внешнему облику напоминающей современных африканцев, являются аборигены Меланезии, имеющие кожу очень темного цвета и густые вьющиеся волосы. Но и в этом случае аборигены горных районов Новой Гвинеи, которые считаются «наименее изменившимися» представителями меланезийской расы по сравнению с людьми плейстоценового периода, имеют крепкокостное строение (толстые кости и выраженные надбровные дуги — см. главу 5), чем сами современные африканцы. Однако в таком случае возникает вопрос: а как выглядели африканцы в эпоху исхода людей современного типа, около 80 тысяч лет тому назад? Можно практически наверняка утверждать, что их внешний облик был мало похож на современных тонкокостных банту, жителей долины Нила, пигмеев или людей народа кунг. Ископаемые останки черепов кафзехского и скульского человека, датируемые временем между 90 и 120 тысячами лет тому назад, являются куда более крепкокостными, чем черепа современных африканцев. Это свидетельствует о том, что жители Африки могли измениться куда больше, чем горцы Новой Гвинеи. Прожив несколько лет в Новой Гвинее и Австралии, я могу сказать, что человек с внешностью, воссозданной при реконструкции одного из черепов кафзехского человека в ходе съемкок нашего документального фильма, был бы сегодня практически неотличим от аборигенов Новой Гвинеи, но ему вряд ли нашлось бы место среди жителей Африки (см. Фото 14 и 15).
Генетические следы исхода среди реликтовых этнических групп, живущих недалеко от Африки
Какие же еще народы в генетическом отношении являются более близкими к африканцам в результате того, что их исконные земли занимают особое положение на пути собирателей-мигрантов? Что может нам дать сопоставление встречаемости и сохранения архаических наследственных генотипов у африканцев и неафриканских народов? Здесь следует выделить два региона. Ближайшими (помимо аборигенов Австралии и Новой Гвинеи) в генетическом смысле к африканцам народами следует считать архаические туземные группы, живущие в Пакистане и на юге Аравийского полуострова. За ними следуют европейцы и жители Индии. И в самом конце этой «очереди» находятся народы Восточной Азии и Америки, проделавшие долгий путь миграции и сохранившие меньше всего следов генетического многообразия своих африканских предков[230]. Ключевые позиции в сохранении древнего генетического многообразия Африки занимают туземные группы Пакистана и Южной Аравии, то есть народы, издревле живущие на пути предполагаемого южного маршрута исхода из Африки. В пользу этой версии говорит целый ряд свидетельств. На южном побережье Аравийского полуострова живет изолированная народность хадрамаут, которую некоторые исследователи относят к австралоидам. Их материнская генетическая структура включает в себя 40% африканских генных линий, но хотя некоторые из этих маркеров связаны с родоначальницей-Евой выходцев из Африки, большинство этих линий проникли сюда из Африки в более поздние времена. Далее, на всем протяжении океанского побережья Индийского субконтинента встречаются этнические группы, занимающие в генетическом отношении более близкое положение к африканским корням исхода, чем жители районов Азии, лежащих дальше к Востоку. Этнические группы Индии, как кастовые, так и родоплеменные, стали объектом детального исследования наличия у них нуклеарных аутосомных (не связанных с половой сегрегацией) маркеров. Они, как было установлено, сохранили более высокую встречаемость африканских наследственных типов, чем европейцы и любые другие расы и этнические группы Азии[231]. Существуют и другие признаки того, что в Пакистане в значительной мере сохранилось генетическое многообразие Древней Африки. В то время как жители Пакистана в Целом имеют общие древние составляющие ДНК с Индией, Европой и Средним Востоком, они обладают также некоторыми уникальными маркерами, которые не встречаются более нигде за пределами Африки. Существуют этнические группы, имеющие очень древние наследственные корни. Одна из архаических групп так называемых негритосов, макрани, живет в дельте Инда и на океанском побережья Белуджистана в Пакистане. Есть предположение, что предки макрани пришли в эти места после исхода из Африки. У макрани выявлен специфически африканский Y-хромосомный маркер, который ранее встречался только в Африке и считался характерным для региона, прилегающего к Сахаре. Тот же генетический маркер, правда, несколько реже, встречается и у других этнических групп южного Пакистана, Саудовской Аравии и Объединенных Арабских Эмиратов. Кроме того, он имеет весьма высокую встречаемость в Иране. Есть и другой уникальный Y-хромосомный маркер, который встречается за пределами Африки только в этом регионе Пакистана. Наконец, третий древний Y-хромосомный маркер указывает на Пакистан как на очень ранний пункт, где началось разделение ветвей. Именно здесь присутствует ранняя ветвь потомков Адама выходцев из Африки, отличающаяся высокой встречаемостью в Пакистане и несколько меньшей — в Индии (особенно у изолированных племен). Кроме того, она отмечается и далее к северу, на Среднем Востоке, в Кашмире, Центральной Азии и Сибири. Тот факт, что этот маркер не выявлен далее к востоку в Азии, свидетельствует о том, что единственным путем, которым он мог проникнуть в Центральную Азию, была миграция с юга, вверх по течению Инда в Кашмир и далее на север[232].
Истоки происхождения европейцев: где появилась на свет Насрин?
Ключевые позиции Южной Азии в качестве средоточия генетического многообразия африканских линий заставляют предположить, что именно там, в Южной Азии, произошло разделение ветвей мигрантов. Теперь нам следует сосредоточить внимание на том, какой биологической силой обладали генетические древа отцовских и материнских линий, оставившие следы древнейших переселений народов. И хотя находка корней и ранних ветвей линий Манью и Насрин на побережье Южной Аравии и Белуджистана является убедительным аргументом в пользу гипотезы о том, что один-единственный исход из Африки по южному маршруту прежде всего привел мигрантов в Южную Азию, она не обязательно является свидетельством того, что именно там появились на свет сама Насрин и ее шесть западных «дочерей», в том числе Рохани, или западно-евразийские «дочери» самой Рохани (U, HV и JT) (рис. 4.2). Хотя Рохани, вне всякого сомнения, появилась на свет где-то в Южной Азии (см. ниже), место рождения ее западно-евразийских дочерей менее ясно и представляет огромный интерес. Однако, если мы рассмотрим генетическую структуру митохондриевой ДНК во всем Ближневосточном регионе (включая Левант, Анатолию, Армению, Азербайджан, Северный Курдистан, а также более южные районы. Йемен, Саудовскую Аравию, Ирак и Иран), приводимую в анализе линий-родоначальниц современной Европы, мы безтруда заметим огромное генетическое многообразие линий западных дочерей Рохани в Ираке — государстве на побережье Персидского залива (см. главу 3). Для Ирака характерно большое количество неклассифицированных генетических типов и отсутствие других западных дочерних групп Насрин, таких, как W, I или X. Таким образом, вполне возможно, что западные «дочери» Насрин появились на свет далеко на юге, в Месопотамии или возле побережья Персидского залива, и произошло это либо накануне первой, направленной на север, миграции в район Плодородного Полумесяца, либо во время нее[233]. Этот вопрос можно решить только посредством формального анализа линий-родоначальниц, такого, которым воспользовались Мартин Ричардс и его коллеги при отслеживании линий предков европейцев (см. главу 3), но в данном случае необходимо сопоставить Южную Азию с Левантом. На мой взгляд, для того чтобы индийская внучка Рохани, U2i, могла иметь такой же возраст (около 50 тысяч лет), как и ее европейская сестра, линия U5, проще всего предположить, что Рохани и ее «дочери» (Европа, HV и JT) появились на свет южнее, в районе Персидского залива, на полпути между Индией и Европой(см. рис. 4.2). Понятно, что, как мы уже говорили в главе 3, имеется веский аргумент в пользу того, что несколько ранних неафриканских Y-хромосомных групп возникли, сформировались именно в Южной Азии, а не на Леванте[234].
 Рис. 4.2
Рис. 4.2
Южная Азия: исток всех азиатских линий?
Хотя проблеме генетических маркеров на юге Аравийского полуострова посвящено не слишком много публикаций, по этой же тематике применительно к Индийскому субконтиненту существует обширный массив самой разной информации. Большую работу по исследованию митохондриевой ДНК провели эстонские генетики, которым, а также их коллегам из стран Азии мы выражаем глубокую признательность за труд. Ряд групп европейских и американских генетиков ведут активные исследования в области изучения Y-хромосом. И хотя их разработки в известной мере ставят под вопрос наш интерес к Пакистану и Индии, мы все же считаем, что корень проблемы следует искать именно там. Пакистан — это древнейшая прародина индусов, основной путь, ведущий из Южной Азии в Центральную, в регион, лежащий к западу от Гималаев. По территории Индии, Бангладеш и Бирмы также протекают великие реки Азии — Ганг, Брахмапутра и Салуин, поящие влагой обширные районы к югу и востоку от Гималаев. Индийский субконтинент, где отмечается широкое многообразие типов внешнего облика и культур населяющих его народов, представляет собой богатейшую этническую и географическую «транзитную зону» между Западной и Восточной Евразией. К северу и западу от нее, по мере продвижения из Индии в Пакистан, из Пакистана в Афганистан и Иран и далее в Европу, мы наблюдаем плавные и постепенные изменения внешнего облика коренных жителей, включая и пигментацию (цвет) кожи (см. главу 5). Внешнее сходство между европейцами и многими народами Южной Азии просто поразительно.
 Рис. 4.3
Этот процесс постепенного изменения физического облика у жителей Северной Индии и Пакистана являет собой как бы параллель относительной частоте встречаемости двух генетических дочерних суперкланов, Насрин и Манью, на которые делится линия Евы выходцев из Африки (L3). (См. рис. 4.3.) Если обратиться к рассмотрению генофонда народа хадрамаут, живущего в Йемене, мы увидим, что соотношение встречаемости линий Насрин и Манью у них составляет 5 к 1, что хорошо согласуется с точкой зрения о том, что ветвь Насрин возникла далее к востоку, чем Манью, примерно в районе Персидского залива. В районе между Красным морем и устьем Инда в Пакистане западноевразийские генетические потомки Насрин продолжают превосходить в численности представителей второго азиатского суперклана — Манью.
Миновав устье Инда, оказавшись в Белуджистане и приближаясь к самой Индии, мы увидим, что картина начинает быстро меняться. Соотношение линий Насрин и Манью здесь, на западе Индии, в штатах Раджастан и Гуджарат, снижается до 2 к 1. А когда мы достигнем Бенгали и проникнем в Бангладеш, это соотношение изменится на противоположное и клан Манью будет доминировать там в пропорции 5 к 1. Таким образом, мы установили, что доминирующим кланом в Индии является Манью, причем его численность по мере продвижения с запада на восток постоянно возрастала[235].
Что касается районов на севере и востоке Индии, то там такие изменения выражены еще более резко. В Непале, Бирме и восточной Индии уже можно встретить первые лица монголоидного типа. Жители этих мест в основном говорят на языках Восточной Азии, что составляет резкий контраст с их ближайшими соседями, говорящими на индо-арийских или дравидийских[236] языках. К тому времени, как мы достигнем территории восточной Бирмы и окажемся в Тибете, лежащем у северных склонов Гималаев, переход внешнего облика и этнолингвистического типа жителей уже можно считать завершенным, поскольку здесь имеет место быстрое и полное изменение соотношения представителей митохондриевых субкланов Манью и Насрин. В частности, на Тибете соотношение представителей Манью и Насрин составляет 3 к 1, и у нас нет достоверных данных о присутствии этих субкланов в Индии. На Тибете выявлено примерно 70% субкланов Манью и Насрин, типичных для Европы и Юго-Восточной Азии, а остальное приходится на долю неклассифицированных типов Манью местного происхождения. Таким образом северо-восточная часть Индийского субконтинента демонстрирует наиболее четкую и резко выраженную границу между востоком и западом[237]. Эта граница, вполне возможно, отражает глубокую генетическую борозду, проходящую через всю Индию и связанную с облаком вулканического пепла, накрывшим Индийский субконтинент в результате извержения вулкана Тоба около 74 тысяч лет тому назад.
На юге Индийского субконтинента основной тип внешнего облика меняется, и преобладающим становится так называемый дравидский тип, для которого характерны курчавые волосы, очень смуглая кожа и круглые глаза (см главу 5). Сравнение формы черепа указывает на связь между тамилами, крупной этнической группой юга Индии, и представителями народности сеной, туземной группой живущей на Малайском полуострове и занимающей в этническом отношении промежуточное положение между народностью семанг и аборигенами Малаи (см. выше)[238].
Рис. 4.3
Этот процесс постепенного изменения физического облика у жителей Северной Индии и Пакистана являет собой как бы параллель относительной частоте встречаемости двух генетических дочерних суперкланов, Насрин и Манью, на которые делится линия Евы выходцев из Африки (L3). (См. рис. 4.3.) Если обратиться к рассмотрению генофонда народа хадрамаут, живущего в Йемене, мы увидим, что соотношение встречаемости линий Насрин и Манью у них составляет 5 к 1, что хорошо согласуется с точкой зрения о том, что ветвь Насрин возникла далее к востоку, чем Манью, примерно в районе Персидского залива. В районе между Красным морем и устьем Инда в Пакистане западноевразийские генетические потомки Насрин продолжают превосходить в численности представителей второго азиатского суперклана — Манью.
Миновав устье Инда, оказавшись в Белуджистане и приближаясь к самой Индии, мы увидим, что картина начинает быстро меняться. Соотношение линий Насрин и Манью здесь, на западе Индии, в штатах Раджастан и Гуджарат, снижается до 2 к 1. А когда мы достигнем Бенгали и проникнем в Бангладеш, это соотношение изменится на противоположное и клан Манью будет доминировать там в пропорции 5 к 1. Таким образом, мы установили, что доминирующим кланом в Индии является Манью, причем его численность по мере продвижения с запада на восток постоянно возрастала[235].
Что касается районов на севере и востоке Индии, то там такие изменения выражены еще более резко. В Непале, Бирме и восточной Индии уже можно встретить первые лица монголоидного типа. Жители этих мест в основном говорят на языках Восточной Азии, что составляет резкий контраст с их ближайшими соседями, говорящими на индо-арийских или дравидийских[236] языках. К тому времени, как мы достигнем территории восточной Бирмы и окажемся в Тибете, лежащем у северных склонов Гималаев, переход внешнего облика и этнолингвистического типа жителей уже можно считать завершенным, поскольку здесь имеет место быстрое и полное изменение соотношения представителей митохондриевых субкланов Манью и Насрин. В частности, на Тибете соотношение представителей Манью и Насрин составляет 3 к 1, и у нас нет достоверных данных о присутствии этих субкланов в Индии. На Тибете выявлено примерно 70% субкланов Манью и Насрин, типичных для Европы и Юго-Восточной Азии, а остальное приходится на долю неклассифицированных типов Манью местного происхождения. Таким образом северо-восточная часть Индийского субконтинента демонстрирует наиболее четкую и резко выраженную границу между востоком и западом[237]. Эта граница, вполне возможно, отражает глубокую генетическую борозду, проходящую через всю Индию и связанную с облаком вулканического пепла, накрывшим Индийский субконтинент в результате извержения вулкана Тоба около 74 тысяч лет тому назад.
На юге Индийского субконтинента основной тип внешнего облика меняется, и преобладающим становится так называемый дравидский тип, для которого характерны курчавые волосы, очень смуглая кожа и круглые глаза (см главу 5). Сравнение формы черепа указывает на связь между тамилами, крупной этнической группой юга Индии, и представителями народности сеной, туземной группой живущей на Малайском полуострове и занимающей в этническом отношении промежуточное положение между народностью семанг и аборигенами Малаи (см. выше)[238].
Ветвь Манью возникла в Индии, а Насрин — несколько западнее, в районе Персидского залива
Ветвь Манью, которая практически полностью отсутствует в Западной Евразии, дает немало оснований утверждать, что ее древней прародиной является Индия. Дело в том, что именно там, в Индии, Манью имеет наибольшее многообразие и древность. Более нигде в мире она не обладает таким множеством вариаций и обилием основных ветвей. Возраст древнейшей из ее «дочерей» в Индии, ветви М2, составляет около 73 тысяч лет. И хотя эта датировка времени распространения М2 не слишком точна, она вполне может отражать местное восстановление населения после гибели в результате катастрофического извержения вулкана Тоба около 74 тысяч лет тому назад. Ветвь М2 широко представлена у ченчу, народа охотников и собирателей, принадлежащего к австралоидной группе и живущего ныне в штате Андхра-Прадеш. Ченчу обладают целым рядом уникальных местных вариантов ветви М2, а также имеют общего предка с типами М2, выявленными в других районах Индии. В целом есть все основания, чтобы считать местом возникновения Манью именно Индию, а не какой-либо другой район, находящийся к западу от нее или в Африке[239]. Спектр типов Насрин, выявленных в Индии, также резко отличается от типов, встречающихся к западу от нее. Хотя типы, характерные для ее «дочери» Европы, выявлены у 13% индийцев, у которых были взяты пробы ДНК, практически все они приходятся на долю двух подветвей Европы, U7 и U2i (локальных индийских версий клана U2). Наряду с рассеянием других кланов Европы, выявленным к западу от Индии, там почти нет коренных типов Насрин. Это свидетельствует о том, что хотя U7 и U2i являются очень древними кланами в Южной Азии, они проникли туда с запада, во времена первой волны заселения Европы из Южной Азии, около 50 тысяч лет тому назад, то есть явно после первоначального исхода из Африки. У нас также нет данных, которые позволили бы поместить локус возникновения Насрин в Индии, хотя там и наблюдается рассеяние первого поколения ее «дочерей», которые имеют двойников в Западной Евразии[240]. Самой удивительной и любопытной среди всех родичей клана Насрин в Индии является ее «дочь», Рохани. Если читатель помнит, Рохани — самая плодовитая «дочь» Насрин, ставшая праматерью большинства людей Запада, включая Европу, не говоря уже о двух ее «дочерях» на Дальнем Востоке, каждая из которых тоже обладает обширным и равзветвленным потомством. При анализе присутствия Рохани в Индии создается впечатление, что ее плодовитость напоминает планирование семьи. Эстонские генетики сумели идентифицировать многочисленные ветви потомков Рохани, берущие начало в Индии, причем ни одна из них не встречается ни в каком другом регионе. Эти новые ветви настолько богаты, что удалось даже датировать время их распространения — около 73 тысяч лет тому назад[241]. Нигде более, ни на западе, ни на востоке, мы не находим столь широко разветвленного многообразия генетической линии Рохани. Одного этого вполне достаточно, чтобы считать Южную Азию исходной прародиной Рохани. Казалось бы, более логично предположить, что Рохани и ее европейские дочери появились на свет именно в районе Персидского залива, а не в Индии, но у нас нет никаких генетических оснований для подобного взгляда. Принимая во внимание сравнительно небольшую удаленность и огромную дистанцию во времени, можно признать, что возможен любой сценарий развития событий. И все же представляется крайне маловероятным, что Рохани могла появиться на свет на Леванте или в Европе. Дело в том, что в этих регионах не обнаружено никаких следов Рохани, а это аргумент в пользу версии о том, что врата для любой миграции на север из районов, прилегающих к Сахаре, в ту эпоху были уже закрыты. Но, пожалуй, самое интересное здесь заключается в том, что невиданно пышный расцвет линий кланов Манью и Насрин в Индии говорит о том, что они увековечили генетическую память о людях, которым посчастливилось выжить во время катастрофического извержения вулкана Тоба около 74 тысяч лет тому назад. Обезлюдевшая Индия, вполне возможно, начала заселяться с запада представителями разных типов клана Рохани, а с востока — представителями ветвей Манью. Дополнительными аргументами в поддержку этой гипотезы являются результаты недавнего исследования Т. Кивисилда и его коллег, изучавших две племенные группы на юго-востоке штата Андхра-Прадеш[242]. Одна из этих групп, охотники-собиратели племени ченчу, относящиеся к австралоидному типу, почти целиком принадлежит к клану Манью и сохраняет некоторые ветви Манью, как характерные для Индии, так и и уникальные для нее. Другая этническая группа, народность койа, не принадлежащая к австралоидному типу, сохранила столь же богатый спектр индийских ветвей Манью (60% от общего числа линий) и, кроме того, имеет 31% типов Рохани. Племенные группы ченчу и койа представляют собой как бы древнюю библиотеку генетических линий Рохани и Манью в Индии, которые являются предками большинства материнских генетических линий из всего их многообразия, отмечаемого на остальной территории Индийского субконтинента. Ни у одного из этих типов не выявлено параллельное присутствие западно-евразийских типов Насрин. Наличие типов Рохани у народности койа, а не у австралоидов ченчу, возможно, объясняется некоторыми аспектами повторного заселения Индийского субконтинента с запада. Любопытно, что у этих двух племенных групп, несмотря на всю их древность и независимую друг от друга эволюцию, а также несомненно индийские генетические корни и локальный характер, нет общих материнских генетических типов (т. е. нет общих точек соприкосновения).
Первые люди Азии: три источника
Можем ли мы проследить маршрут и восстановить историю миграции линий-родоначальников Адама выходцев из Африки вдоль побережья Аравийского полуострова и далее через Пакистан в Индию? Задача была бы куда легче, если бы на пути миграции остались представители Адама выходцев из Африки, но корневая линия Адама, как и Евы выходцев из Африки, полностью вымерла, и у него остались лишь три непосредственных «сына» — Каин, Авель и Сиф, три мужские линии, которые охватывают все остальное человечество за пределами Африки[243]. Одна из этих трех корневых ветвей, С (или RPS4Y), которой я дал название Каин — по имени первородного сына Адама, представляет ветвь, встречающуюся только у неафриканского населения и в небольшом количестве присутствующую у коренных жителей Южной и Восточной Азии, Океании, а также Северной и Южной Америки. С еще более низкой (около 5%) встречаемостью линия Каин присутствует в разных районах Индии, включая и наиболее австралоидное из ее этнических меньшинств — племя ченчу. Линия Сифа, F (или М89), также является практически полностью неафриканской, за исключение разве что нескольких корневых типов, которые, как и ветви материнской линии Манью (M1), простираются вплоть до Северной Африки и, в частности, присутствуют в Марокко. Сиф — бесспорно, самая распространенная из этих ветвей; она встречается во всем мире за пределами Африки, причем ее встречаемость особенно высока в Австралии и Новой Гвинее, что со всей определенностью демонстрирует, что она представляет собой соединение двух генетических сыновей Адама на пути их эпических странствий по прибрежной полосе. На долю Сифа приходится около 95—97% всех мужских линий Индии, и он образует там множество ветвей[244]. Три наиболее распространенные в Индии ветви не встречаются практически более нигде в мире (единственное исключение — некоторые районы Центральной Азии к северу от Индии) и, таким образом, словно эхо отражают это пышное цветение материнских линий в Индии. Две из них представляют собой линии потомков группы К, Кришны, наиболее плодовитого сына Сифа, тогда как третья — это ветвь первого поколения, идущая от Сифа и встречающаяся не только в упомянутых районах Центральной Азии, но и, что особенно важно, далеко на юговостоке, в Камбодже. Все эти типы активно представлены у племен и этнических групп Южной Индии, особенно — у австралоидных ченчу. Люди племени койа, кроме того, сохранили и редкий корневой тип Сифа, встречающийся у четверти койа. Этот тип ограничен Индийским субконтинентом и соседними районами Центральной Азии, что является аргументом в поддержку версии о том, что именно Индия явилась для Сифа важнейшей «опорной базой» на его южном пути из Африки[245]. Согласно другой концепции исхода из Африки, полученной на основе данных по Y-хромосоме, в первоначальной группе мигрантов-собирателей на прибрежной полосе продвижения присутствовали только линии Каин и Авель, Сиф же появился много позднее, пройдя через Северную Африку и Левант и направившись в Центральную Азию, а уже оттуда — в Индию и Юго-Восточную Азию. Как сказано в этой и следующей главах нашей книги, я не разделяю подобную точку зрения, доказывая посредством целого ряду аргументов (включая близкое географическое и генетическое совпадение линий-родоначальниц), что объяснить все имеющиеся находки лучше всего позволяет версию об одном-единственном, южном, маршруте исхода. Что же касается присутствия линии Сифа в Центральной Азии, то наиболее распространенный индийский тип Y-хромосом является ключом к решению вопроса о том, берут ли многочисленные Y-хромосомные линии, присутствующие в Индии и Центральной Азии, свое начало в Индии или наоборот. Например, тип M17, который я уже упоминал в главе 3 как одного из главных фигурантов в Европе, пришел именно из Центральной Азии. Кроме того, M17 имеет в Индии среднюю встречаемость порядка 27%, но достигает в одном из ее штатов, штате Пенджаб, уровня 47%. Наибольшего многообразия — куда более широкого, чем в Центральной Азии или Европе, — М17 достигает в Иране, Индии и Пакистане. Это относится и к Южной Азии, которая является его древней прародиной. Ключевой вопрос здесь — поиск М17 при высокой встречаемости и широком многообразии у представителей племенных групп Южной Индии, в частности у австралоидных племен ченчу и ядхава. Если бы M17 у представителей этих племенных групп была привнесенной, а не коренной линией, ее многообразие было бы значительно более низким[246].
Вопрос о противоречивости линии YAP: африканская она или азиатская?
Третьей основной линии, часто называемой YAP, я дал имя Авель — по имени второго сына Адама. Одна из ветвей этой линии — небольшая, глубокая и очень древняя азиатская ветвь, распространение которой носит очаговый, дискретный характер, в основном — на Дальнем Востоке (более подробно о ней см. ниже). Другая, западная, ветвь YAP образует несколько ответвлений, представители которых заселили многие районы Северной Африки и Среднего Востока, хотя в небольшом количестве она присутствует у жителей Пакистана, Индии и Центральной Азии. Некоторые подветви YAP относятся к числу наиболее распространенных в Африке, тогда как другие характерны только для Среднего Востока[247]. Здесь можно вспомнить аналогию с Библией: в первой ее книге, книге Бытия, сказано, что Сим и Хам были двумя братьями, потомки которых заселили соответственно Средний Восток и Северную Африку. К большому сожалению для наших изысканий, первоначальная мужская линия Авеля, возникшая еще до разделения YAP+ на западную и восточную ветви, по всей вероятности, не сохранилась и полностью вымерла, так что от нее остались лишь два «сына». Все, что оставила нам эта ветвь, — это тайну своего происхождения. Где же она возникла: в Эфиопии или Аравии? Вопрос, вынесенный в заглавие параграфа «африканская или азиатская?» присутствует и в названии «хамито-семитская», используемом для характеристики языковой семьи, на языках которой говорит подавляющее большинство потомков YAP. Но, независимо от того, какое конкретно название мы используем применительно к ней — старое, восходящее к Библии, или же более новое, «афроазиатская», — главный вопрос сводится к тому, являются ли корни происхождения этой языковой семьи африканскими или азиатскими. Учитывая сравнительно молодой возраст большинства крупных языковых семей, можно предположить, что любая миграция через Красное море, будь то с востока на запад или с запада на восток, по-видимому, имела место около 10 тысяч лет тому назад. Известный американский генетик Майк Хаммер настаивал, что линия YAP возникла и сложилась за пределами Африки и что один из ее «сыновей» впоследствии возвратился на древнюю прародину. Другая группа генетиков во главе с Питером Андерхиллом отстаивает версию об африканском происхождении YAP[248]. В этом вопросе я склонен согласиться с Майком Хаммером и утверждать, что линия YAP присутствовала в первой колонии выходцев из Африки, однако неясность происхождения афроазиатской языковой семьи заключает в себе нечто большее, чем простую аналогию (см. рис. 4.4). Неафриканские генетические линии, которые восходят к Эфиопии и Северной Африке[249], вполне могли сложиться и после последнего ледникового максимума, когда люди начали активно возвращаться на зеленые пространства, еще недавно бывшие пустынями. Большой интерес представляет вопрос о том, когда именно после последнего оледенения имело место это расселение. Некоторые археологи и лингвисты считают, что заметное увеличение крупнейших языковых семей было связано с сельскохозяйственной революцией эпохи неолита, имевшей место в последние 5000—7000 лет[250]. Однако генетические данные показывают, что повторное расселение имело место в эпоху мезолита (т. е. примерно 12 500 лет тому назад), то есть, другими словами, гораздо раньше эпохи неолита[251].
 Рис. 4.4
Загадочным аспектом миграции линии Авеля является зияющая пауза в ее расселении между Западной Евразией и Дальним Востоком и, что особенно важно, ее практически полное отсутствие в Индии. Тот факт, что она тоже присутствовала в группах мигрантов-собирателей, продвигавшихся по прибрежной полосе, заметен благодаря присутствию азиатской YAP у аборигенов Андаманских островов, Камбоджи и Японии (см. главу 5). Данные недавних исследований свидетельствуют о присутствии азиатской ветви YAP на уровне встречаемости 3—6% и такой же частоты на западе, далеко к северу от прибрежной полосы, в частности в Центральной Азии (Узбекистан). Это отражает картину расселения линии Каин, которая хотя и не полностью отсутствует в Индии, тем не менее имеет там гораздо более низкую встречаемость, чем на севере, в Центральной Азии. Учитывая, что азиатская ветвь YAP хотя и редко, но все же встречается по древнему пути миграции собирателей на прибрежной полосе и рядом с ним, вполне возможно, что ее отсутствие в Индии — а также низкую встречаемость линии Каин в Южной Азии — можно объяснить опустошительным влиянием катастрофического извержения вулкана Тоба на весь Индийский субконтинент, — извержения, о котором свидетельствуют данные геологии и генетической истории материнских линий (см. выше). Извержение Тоба могло создать генетическую перемычку, за которой последовало активное местное восстановление линии Сифа за счет двух братьев-собирателей на прибрежной полосе — Каина и Авеля[252].
Рис. 4.4
Загадочным аспектом миграции линии Авеля является зияющая пауза в ее расселении между Западной Евразией и Дальним Востоком и, что особенно важно, ее практически полное отсутствие в Индии. Тот факт, что она тоже присутствовала в группах мигрантов-собирателей, продвигавшихся по прибрежной полосе, заметен благодаря присутствию азиатской YAP у аборигенов Андаманских островов, Камбоджи и Японии (см. главу 5). Данные недавних исследований свидетельствуют о присутствии азиатской ветви YAP на уровне встречаемости 3—6% и такой же частоты на западе, далеко к северу от прибрежной полосы, в частности в Центральной Азии (Узбекистан). Это отражает картину расселения линии Каин, которая хотя и не полностью отсутствует в Индии, тем не менее имеет там гораздо более низкую встречаемость, чем на севере, в Центральной Азии. Учитывая, что азиатская ветвь YAP хотя и редко, но все же встречается по древнему пути миграции собирателей на прибрежной полосе и рядом с ним, вполне возможно, что ее отсутствие в Индии — а также низкую встречаемость линии Каин в Южной Азии — можно объяснить опустошительным влиянием катастрофического извержения вулкана Тоба на весь Индийский субконтинент, — извержения, о котором свидетельствуют данные геологии и генетической истории материнских линий (см. выше). Извержение Тоба могло создать генетическую перемычку, за которой последовало активное местное восстановление линии Сифа за счет двух братьев-собирателей на прибрежной полосе — Каина и Авеля[252].
Три азиатские мужские линии-родоначальницы: все они были собирателями на прибрежной полосе
Давайте возвратимся к трем основным мужским линиям выходцев из Африки. Можем ли мы задать такой же вопрос, который обращали к нуклеарным и митохондриевым маркерам относительно следов древнейшего исхода из Африки? Я думаю — да. Датировка Y-ветвей пока что находится в зачаточном состоянии, однако, если мы обратимся к географическим локусам стоянок предков и потомков, мы сможем оценить географическую последовательность распределения ветвей. Это — так называемый филологогеографический метод. Для начала возьмем линию Каин. Поскольку на ее долю приходится 60% всех типов Y-хромосом в Австралии, она является там доминирующей линией. На таких островах Восточной Индонезии, как Молуккские острова и о. Нуса-Тенггара, которые первыми встретились мигрантам на их пути в Австралию, мы видим наименее модифицированные или даже корневые типы линии Каин, от которой происходит уникальный аборигенный австралийский тип. Единственным местом в мире, где также выявлен этот корневой тип, является Индия, где он имеет довольно низкую встречаемость, в том числе и у представителей туземных племен австралоидного типа. На соседнем острове Новая Гвинея вы встречаем линию С2, близко родственную («братскую») австралийской линии Каин. Взглянув на локусы других древнейших колоний, мы без труда заметим, что Каин практически отсутствует в Западной Евразии, но зато широко распространен к востоку и северу от Индии. Резкое падение встречаемости линии Каин показывает, что она по большей части присутствует на восточно-азиатском побережье Тихого океана и гораздо реже — в глубинных континентальных районах Центральной, Северной и Южной Азии. Эти азиатские типы по большей части принадлежат к одному клану, который впоследствии мигрировал в Америку, явившись родоначальником тамошних аборигенов. Широкое многообразие линий этого клана отмечено в Японии[253]. Распространение линии Каина в прибрежных районах Восточной Азии и выраженные региональные различия у ее потомков, наблюдаемые в Юго-Восточной Азии и Океании (рис. 4.5), свидетельствуют о том, что эта линия была в числе родоначальниц мужских линий на пути первоначальной миграции собирателей по всему южному побережью Азии, хотя ее и нельзя считать единственной, поскольку обе линии-«брата» также оставили следы своего продвижения на побережье Индийского океана в ЮгоВосточную Азию и Японию. Однако длинный путь в Австралию, а также к побережью Восточной Азии преодолела только одна из двух оставшихся ветвей — линия Сиф. Ее следы выявлены благодаря присутствию (правда, достаточно редкого) линии Сиф в ее первоначальном или корневом виде во всех указанных регионах. В Австралии выявлены два основных локальных типа Y-хромосом: один из них происходит от линии Каин (см. выше) и является наиболее распространенным, поскольку его численность составляет 60% от общего числа линий, а другой — от азиатского генетического «сына» Сифа, Кришны, на долю которого приходится до 30% оставшихся линий. Такой уровень встречаемости Кришны (около 30%) сохраняется более или менее стабильно на всем побережье Юго-Восточной Азии и западного побережья Тихого океана вплоть до Кореи. В то же время линия потомков Сифа присутствует на уровне нескольких процентов в Юго-Восточной Азии и Австралии, а также на тихоокеанском побережье, в Корее. Эти данные свидетельствуют о том, что первый маршрут собирателей на прибрежной полосе пролегал не только в Новую Гвинею и Австралию, но и продолжался далее, по всему тихоокеанскому побережью Китая, Японии и Кореи. Третья корневая линия Y-хромосомы, азиатская YAP, доходит до Камбоджи, а затем круто сворачивает в сторону Австралии, вместо того чтобы продолжать путь на север, вокруг побережья Тихого и Индийского океанов. Она имеет эпизодическую встречаемость в Юго-Восточной Азии и очень высокую — на Тибете, в южном Китае и Японии[254].
 Puc. 4.5
Подобная картина распространения мужских генетических линий служит зеркальным отражением миграций материнской мтДНК и очень много, хотя и в косвенной форме, говорит нам об идентификации и возрасте первых мужских линий-родоначальниц. Согласно прогнозам, южный маршрут имел место где-то в самом начале пути, и прежде чем приступать к его отслеживанию, мы должны попытаться найти точку, в которой сходятся все три древнейшие неафриканские линии. В настоящее время ведутся исследования на юге Аравийского полуострова и в районе Персидского залива, но если мы обратимся к рассмотрению этих трех мужских линий-родоначальниц в Пакистане и Индии, мы без труда установим, что там представлены все три эти линии. Мы можем найти и корень, и ветви линий Каин, Сиф и генетических «сыновей» и «внуков» Сифа. Более того, мы найдем и ряд уникальных типов YAP от ветви YAP, которая обычно встречается только в Африке и Западной Евразии[255].
Нигде более за пределами Африки мы не встречаем такого широкого многообразия корней и ветвей Y, если не считать регионов Центральной и Северной Азии, да и там они, надо признать, представлены далеко не столь пышно и обильно. Эта картина, характерная для Центральной Азии как промежуточной зоны между Востоком и Западом, обусловлена активным смешением европейских и азиатских материнских линий мтДНК, представленных в этом регионе[256], что свидетельствует о том, что одним из самых первых этапов разделения после прибытия в Индию был маршрут на север, вверх по течению Инда, ведущий в Центральную Азию. Этот ранний маршрут, устремленный в глубь континента, более подробно рассмотрен в следующей главе.
Таким образом, весь регион Южной Азии, древнейшей прародины участников одного-единственного и успешного исхода по южному маршруту, свидетельствует о присутствии в нем генетических корней, связанных с миграциями не только так называемых туземных этнических групп вдоль побережья Индийского океана, но и многих предков современных народов. Среди этих корней мы можем выявить базовые генетические «лагеря» и опорные пункты большинства последующих миграций групп и племен в глубь континента, по необъятным просторам Евразии. Эти маршруты миграций после небольшой паузы были направлены в Европу, Кавказский регион и Центральную Азию. Создается впечатление, что передовые отряды этих мигрантов по прибрежной полосе сохранили удивительное первозданное генетическое многообразие, характерное группе выходцев из Африки, и двинулись в путь по побережью Индийского океана. Таким путем они довольно скоро достигли Индонезии и островов западной Океании, проникнув в Австралию задолго до того, как их кузины и кузены, отправившиеся на запад, оказались в Европе.
Точная хронологическая взаимосвязь между исходом из Африки и проникновением в Юго-Восточную Азию, с одной стороны, и катастрофическим извержением вулкана Тоба около 74 тысяч лет тому назад, с другой, имеет критически важное значение. Прежде всего, извержение Тоба — это одно из наиболее точно и достоверно датируемых событий эпохи палеолита, и выпадение его вулканического пепла можно считать точным критерием датировки для всей Южной Азии. Во-вторых, пагубный эффект вулканического пепла и последовавшая за ним долгая «ядерная зима» неизбежно должны были оказаться губительными для всего живого на пути выпадения пепла, а фактически захватили куда более обширный регион. Наличие в вулканическом пепле Тоба орудий, созданных, как считается, людьми современного типа на стоянках на Малайском полуострове, свидетельствует, что передовые отряды мигрантов-собирателей на прибрежной полосе проникли на Дальний Восток задолго до извержения Тоба. Триангуляция этой рубежной даты и других дошедших до нас фактов говорит в пользу такого сценария. В числе других «ключей» — новые данные по изучению люйцзяньского черепа, данные датировки по люминесцентному методу объектов в Австралии и оценки минимального уровня Мирового океана, позволявшего совершить миграцию в Австралию около 70 тысяч лет тому назад практически сухопутным путем, генетические данные о расселении группы L3 около 83 тысяч лет тому назад и, наконец, период резкого повышения солености Красного моря, начавшийся около 80 тысяч лет тому назад. Наиболее убедительными свидетельствами очень раннего присутствия человека современного типа в Азии могут стать находки ископаемых останков и исследование всего контекста их культурного слоя. Такие работы активно ведутся на стоянке Лианг Буа на о. Флорес.
Таким образом, если взрыв вулкана Тоба действительно произошел уже после того, как Индия была заселена людьми, мы вправе ожидать массовой гибели населения на всем Индийском полуострове в результате извержения, затронувшего восточную часть субконтинента значительно сильнее, чем западную. Это всего лишь одно из возможных объяснений парадокса генетической картины Индии, где можно выявить генетический след древнейших мигрантов-собирателей на прибрежной полосе. В то же время множество индийских подгрупп линий Манью и Рохани являются уникальными и ограничиваются только самим субконтинентом; в особенности это относится к племенам на юго-востоке полуострова. А это — именно та картина, которую мы вправе ожидать после грандиозной катастрофы. Возраст наиболее древних из этих локальных линий достигает 73 тысяч лет.
В следующей главе мы рассмотрим вопрос о том, как повели себя эти первопроходцы на севере и востоке азиатского континента и каким образом они проникли в эти места.
Puc. 4.5
Подобная картина распространения мужских генетических линий служит зеркальным отражением миграций материнской мтДНК и очень много, хотя и в косвенной форме, говорит нам об идентификации и возрасте первых мужских линий-родоначальниц. Согласно прогнозам, южный маршрут имел место где-то в самом начале пути, и прежде чем приступать к его отслеживанию, мы должны попытаться найти точку, в которой сходятся все три древнейшие неафриканские линии. В настоящее время ведутся исследования на юге Аравийского полуострова и в районе Персидского залива, но если мы обратимся к рассмотрению этих трех мужских линий-родоначальниц в Пакистане и Индии, мы без труда установим, что там представлены все три эти линии. Мы можем найти и корень, и ветви линий Каин, Сиф и генетических «сыновей» и «внуков» Сифа. Более того, мы найдем и ряд уникальных типов YAP от ветви YAP, которая обычно встречается только в Африке и Западной Евразии[255].
Нигде более за пределами Африки мы не встречаем такого широкого многообразия корней и ветвей Y, если не считать регионов Центральной и Северной Азии, да и там они, надо признать, представлены далеко не столь пышно и обильно. Эта картина, характерная для Центральной Азии как промежуточной зоны между Востоком и Западом, обусловлена активным смешением европейских и азиатских материнских линий мтДНК, представленных в этом регионе[256], что свидетельствует о том, что одним из самых первых этапов разделения после прибытия в Индию был маршрут на север, вверх по течению Инда, ведущий в Центральную Азию. Этот ранний маршрут, устремленный в глубь континента, более подробно рассмотрен в следующей главе.
Таким образом, весь регион Южной Азии, древнейшей прародины участников одного-единственного и успешного исхода по южному маршруту, свидетельствует о присутствии в нем генетических корней, связанных с миграциями не только так называемых туземных этнических групп вдоль побережья Индийского океана, но и многих предков современных народов. Среди этих корней мы можем выявить базовые генетические «лагеря» и опорные пункты большинства последующих миграций групп и племен в глубь континента, по необъятным просторам Евразии. Эти маршруты миграций после небольшой паузы были направлены в Европу, Кавказский регион и Центральную Азию. Создается впечатление, что передовые отряды этих мигрантов по прибрежной полосе сохранили удивительное первозданное генетическое многообразие, характерное группе выходцев из Африки, и двинулись в путь по побережью Индийского океана. Таким путем они довольно скоро достигли Индонезии и островов западной Океании, проникнув в Австралию задолго до того, как их кузины и кузены, отправившиеся на запад, оказались в Европе.
Точная хронологическая взаимосвязь между исходом из Африки и проникновением в Юго-Восточную Азию, с одной стороны, и катастрофическим извержением вулкана Тоба около 74 тысяч лет тому назад, с другой, имеет критически важное значение. Прежде всего, извержение Тоба — это одно из наиболее точно и достоверно датируемых событий эпохи палеолита, и выпадение его вулканического пепла можно считать точным критерием датировки для всей Южной Азии. Во-вторых, пагубный эффект вулканического пепла и последовавшая за ним долгая «ядерная зима» неизбежно должны были оказаться губительными для всего живого на пути выпадения пепла, а фактически захватили куда более обширный регион. Наличие в вулканическом пепле Тоба орудий, созданных, как считается, людьми современного типа на стоянках на Малайском полуострове, свидетельствует, что передовые отряды мигрантов-собирателей на прибрежной полосе проникли на Дальний Восток задолго до извержения Тоба. Триангуляция этой рубежной даты и других дошедших до нас фактов говорит в пользу такого сценария. В числе других «ключей» — новые данные по изучению люйцзяньского черепа, данные датировки по люминесцентному методу объектов в Австралии и оценки минимального уровня Мирового океана, позволявшего совершить миграцию в Австралию около 70 тысяч лет тому назад практически сухопутным путем, генетические данные о расселении группы L3 около 83 тысяч лет тому назад и, наконец, период резкого повышения солености Красного моря, начавшийся около 80 тысяч лет тому назад. Наиболее убедительными свидетельствами очень раннего присутствия человека современного типа в Азии могут стать находки ископаемых останков и исследование всего контекста их культурного слоя. Такие работы активно ведутся на стоянке Лианг Буа на о. Флорес.
Таким образом, если взрыв вулкана Тоба действительно произошел уже после того, как Индия была заселена людьми, мы вправе ожидать массовой гибели населения на всем Индийском полуострове в результате извержения, затронувшего восточную часть субконтинента значительно сильнее, чем западную. Это всего лишь одно из возможных объяснений парадокса генетической картины Индии, где можно выявить генетический след древнейших мигрантов-собирателей на прибрежной полосе. В то же время множество индийских подгрупп линий Манью и Рохани являются уникальными и ограничиваются только самим субконтинентом; в особенности это относится к племенам на юго-востоке полуострова. А это — именно та картина, которую мы вправе ожидать после грандиозной катастрофы. Возраст наиболее древних из этих локальных линий достигает 73 тысяч лет.
В следующей главе мы рассмотрим вопрос о том, как повели себя эти первопроходцы на севере и востоке азиатского континента и каким образом они проникли в эти места.
ГЛАВА ПЯТАЯ РАННИЕ АЗИАТСКИЕ ВЕТВИ
Все мы склонны подмечать различия. «Мамочка, почему этот дядя так не похож на того?» — подобные вопросы часто можно услышать из уст малышей. Итак, почему же люди, живущие на разных концах света, столь непохожи друг на друга? И почему взрослых и детей так занимает этот вопрос? Для многих и очень многих проблема этнических различий нашла свое выражение в идеях расизма, который является самым чудовищным и безобразным проявлением массового сознания. Но если попросту закрыть глаза на проблемы, окружающие нас со всех сторон, придется вечно топтаться на одном и том же месте. Этнические различия от этого вовсе не исчезнут. И лишь непредвзятые научные исследования, быть может, позволят нам выяснить, «почему этот дядя так похож на того» и почему нас тревожит вопрос о тех внешних различиях, которые характеризуют различные этнические группы. В предыдущей главе уже была в общих чертах изложена теория формирования этносов огромного азиатского континента. Она состоит в том, что многочисленные этнические группы, которые ныне заселяют просторы Азии, некогда распались на более мелкие, которые в свою очередь разошлись в разные концы обширного континента, что позволило им развиваться независимо друг от друга. Хотя распад крупного этноса на более мелкие кажется совершенно очевидным, а теория изолированного развития этнических групп получила научное подтверждение на основе данных генетики, однако нет пока достаточной ясности в том, каким был конкретно исторический процесс на азиатском континенте: ни окаменелости, ни археологические находки не раскрывают тайн прошлого. Дело в том, что археологические находки весьма немногочисленны, а окаменелостей сохранилось еще меньше, поэтому кажется преждевременным и спекулятивным любой ответ на вопрос, где предки различных современных этнических групп Азии обитали более чем за 70 тысяч лет до нас и когда они расселились на необозримых просторах этого континента. Окаменелости не позволяют получить точные ответы на поставленные вопросы, так обратимся же вновь к археологии, поскольку мы в состоянии датировать первобытные стоянки, опираясь на данные климатологии и генетики, а это в свою очередь поможет нам реконструировать маршруты передвижения первых жителей Азии. Пионеры проникали внутрь гигантского континента с трех сторон азиатского побережья Индийского и Тихого океанов. Выбор одного из трех направлений, изоляция и адаптация к новым условиям — все это сказалось на внешнем облике первопроходцев, глубоко врезалось в их черты и сделало их столь разными. Почему вопрос о внешних различиях волнует нас, как ни один другой? Что мы имеем в виду, когда говорим «разные»? Ответ на первый вопрос, возможно, коренится в самой природе человека, в том, что время отшлифовало в нас способность узнавать и помнить невероятное количество разных лиц. Нам не обойтись без этой способности отчасти уже потому, что круг нашего общения весьма широк[257]. Он значительно шире круга общения наших ближайших родственников из числа животных — шимпанзе, и взаимодействия между членами социальных групп у людей оказываются значительно сложнее. Мы должны уметь узнавать в лицо многих и многих людей. Оказавшись не в состоянии вспомнить имя того или иного человека, которое нам следовало бы помнить, мы смущаемся и краснеем, понимая, что допустили непростительную оплошность. Свою хорошую память на лица можно не только выгодно использовать, вращаясь в определенном обществе, но, обобщая и классифицируя свои наблюдения, можно выделить некие черты и внешние особенности, характерные для определенной группы людей, а затем найти различия между этой группой и какой-нибудь другой. Совершенно очевидно, что, разделив на основании внешних признаков людей на «своих» и «чужих», то есть тех, кто выглядит «по-другому», мы будем вести себя среди «своих» совершенно иначе, чем среди «чужих», относясь к последним с явным пренебрежением. К счастью, мы можем проникнуть внутренним взором в глубины своей совести и увидеть, что душа наша бессознательно стремится делить людей на «своих» и «чужих», что может привести к чудовищным преступлениям против человечности — к организованному расизму, и это понуждает пас следовать закону и усилием воли подавлять зародыши расизма в собственной душе и объявлять их вне закона. К сожалению, подобный самоконтроль не всегда увенчивается успехом. И мы оказываемся не в состоянии учиться на собственных ошибках, забывая злодейские погромы и травлю национальных меньшинств, забывая тех людей, что в прошлом стали жертвами расизма.
Ограниченные возможности слова
Побочным результатом борьбы с расизмом стала дискуссия о целесообразности признать табуированным само слово «раса», репутация которого непоправимо пострадала во времена нацизма. Поскольку в чистом виде рас не существует — они представлены многочисленными местными вариантами, переходными формами и смешанными разновидностями, многие антропологи считают понятие «раса» ненаучным, нарушающим права человека, бессмысленным и производящим впечатление некой абстракции, а следовательно, достойным признания вне закона. Все приведенные выше наблюдения весьма ценны, но факт остается фактом — и дети всегда его замечают: люди из разных регионов одного и того же континента могут иметь выраженные различия во внешнем облике. Короче говоря, ни запрет термина «раса», ни регулярная замена его различными эвфемизмами в такой ситуации не помогут. Наиболее часто в качестве альтернативы этому термину используются слова «население» или «этническая группа», но эти понятия столь расплывчаты, что могут лишь окончательно завести нас в тупик. Чтобы обозначить внешне отличающиеся типы людей, я использую в своей книге следующие термины — я вовсе не настаиваю на том, что они абсолютно точны, но зато они общеупотребительны и общепонятны, — итак, вот эти термины: «кавказцы» (или «кавказоиды»), «монголоиды» и «негритосы». Многие антропологи сочтут их неточными и отвергнут как явление, оскорбляющее личное достоинство человека.
 Рис. 5.0
Читатели, которые разделяют подобные взгляды, могут обратиться к таблице 5.0 — она позволяет перевести привычную для меня терминологию в систему терминов, основанную на географическом принципе и позволяющую обозначить туземное население, для которого характерен широкий спектр специфических особенностей строения организма через посредство зон его проживания.
Хотя я и оставляю за собой право пользоваться привычной для меня терминологией, я вовсе не испытываю презрения к людям, чья внешность не похожа на мою собственную, — напротив, я прекрасно понимаю, что у всех прочих людей земного шара есть одно явное передо мной преимущество — ибо все они так не похожи на меня. Но вместе с тем я уверен, что политически корректные эвфемизмы и более современные термины могут скорее завести в тупик, нежели прояснить суть дела. Всем, конечно, известно слово «монголоиды» — оно часто используется для краткого обозначения целой категории людей, наделенных определенными чертами внешности и телосложения. Однако необходимо всегда помнить, что «традиционные» термины — это лишь неточные обозначения сложных жизненных явлений, которые не могут существовать вне бесчисленных конкретных вариаций в пределах определенной группы и вне системы взаимовлияний между различными народами.
Самым очевидным дифференцирующим признаком, позволяющим отличать одни народы Евразии по внешнему виду от других, является цвет кожи, которая в более солнечных тропических регионах приобретает темный шоколадный оттенок. И это не случайное совпадение обстоятельств. Хотя гены еще таят в себе немало загадок, мы можем утверждать, что темный цвет кожи, обязанный своим появлением пигменту меланину, определяется генетической памятью человека, которая формировалась в процессе длительной эволюции. Кожа людей, которые живут в тропиках и субтропиках, подвергается мощной атаке ультрафиолетовых лучей. В результате этого человек рискует пострадать от чрезмерного загара и покрыться волдырями, а также умереть от рака кожи. Лишь темный цвет кожи способен уменьшить риск и угрозу для здоровья человека. Среди других, хотя и не столь очевидных преимуществ, можно назвать то, например, что пигментация, в которой участвует меланин, позволяет коже более эффективно противостоять перегреву и излучать излишки тепла, а также препятствует разрушению в организме такого важного витамина, как фолиевая кислота.
Итак, в результате длившейся многие и многие поколения эволюции кожа людей в жарких климатических поясах стала темной. Именно темнокожие среди множества своих чад и домочадцев в областях повышенной солнечной активности живут дольше и чувствуют себя как нельзя более счастливыми[258]. В Северной Азии (т.е. в той части Азии, которая охватывает территории от Цзиньхай-Тибетского плато на севере и до Урала на востоке) и в Европе солнечный свет не столь интенсивен, и риск развития рака кожи менее выражен, зато население этих регионов повсеместно страдает от рахита и различных заболеваний костей, вызванных недостатком солнечного света. В самом Лондоне, причем уже в начале двадцатого столетия, от этих заболеваний умерло множество детей. Я сам был очевидцем подобных трагедий. Дело в том, что, попав как-то раз в больницу, а именно в отделение патологии Лондонского королевского госпиталя, я специально узнавал, насколько распространен рахит в наши дни. Во второй половине двадцатого столетия в Великобритании вновь получили широкое распространение такие заболевания, как рахит и остеомаляция[259], вторгшиеся в наши широты вместе с переселенцами с Индийского субконтинента. Именно дети потомственных индийцев чаще всего страдают рахитом. Это отчасти объясняется тем, что их темная кожа продолжает фильтровать солнечный свет, как это было необходимо на родине их предков, а не здесь, где солнечного света и без того недостаточно.
Итак, направление эволюции определяется двумя взаимосвязанными факторами естественного отбора, под влиянием которых сформировалось повсеместно наблюдаемое явление: по мере приближения к нулевым широтам цвет кожи местных жителей становится все более темным. Эти обусловленные солнцем изменения цвета кожи и волос вырабатывались из поколения в поколение в процессе длительной эволюции. Те факторы естественного отбора, которые определяют темный цвет кожи, очевидно, повлияли на генетический код жителей Африки. Если же покинуть Африку и отправиться путешествовать по обширным территориям Евразии, спускаясь изСкандинавии, на севере Европы, и Сибири, на севере Азии, к землям Италии и Юго-Восточной Азии, на которых расположены южные оконечности обоих континентов, можно наблюдать, как постепенно меняется цвет кожи и волос местных жителей.
Подобно африканцам, другие темнокожие народы живут преимущественно в солнечных тропических странах. Вполне очевидно, что мы столкнемся и с исключительными случаями, но они будут представлены потомками тех людей, которые в недавнем прошлом покинули родину и поселились в южных странах, их кожа будет еще слишком светла для этих широт (или наоборот, слишком темна), что и естественно, ведь процесс эволюции длится многие поколения. Таким примером-исключением может служить Австралия, которая заселена потомками недавних эмигрантов, поэтому жители этой страны по большей части до сих пор остаются «бледнолицыми». В Австралии, как нигде на земном шаре, высок процент людей, страдающих раком кожи, но и здесь уже начался постепенный процесс эволюции, который, по всей вероятности, ведет к тому, что потомки выходцев из Европы приобретут естественный для этих широт темный цвет кожи. И наоборот, первые переселенцы, отправившиеся с юга на север в страны Северной Европы и Азии, выглядели неестественно темными среди местных жителей, но со временем кожа их потомков стала более бледной. Однако оставим на время Австралию. Дело в том, что в какой бы точке Земли мы ни оказались, цвет кожи местных жителей будет связан с местным уровнем интенсивности ультрафиолетового излучения (рис. 5.1)[260].
Рис. 5.0
Читатели, которые разделяют подобные взгляды, могут обратиться к таблице 5.0 — она позволяет перевести привычную для меня терминологию в систему терминов, основанную на географическом принципе и позволяющую обозначить туземное население, для которого характерен широкий спектр специфических особенностей строения организма через посредство зон его проживания.
Хотя я и оставляю за собой право пользоваться привычной для меня терминологией, я вовсе не испытываю презрения к людям, чья внешность не похожа на мою собственную, — напротив, я прекрасно понимаю, что у всех прочих людей земного шара есть одно явное передо мной преимущество — ибо все они так не похожи на меня. Но вместе с тем я уверен, что политически корректные эвфемизмы и более современные термины могут скорее завести в тупик, нежели прояснить суть дела. Всем, конечно, известно слово «монголоиды» — оно часто используется для краткого обозначения целой категории людей, наделенных определенными чертами внешности и телосложения. Однако необходимо всегда помнить, что «традиционные» термины — это лишь неточные обозначения сложных жизненных явлений, которые не могут существовать вне бесчисленных конкретных вариаций в пределах определенной группы и вне системы взаимовлияний между различными народами.
Самым очевидным дифференцирующим признаком, позволяющим отличать одни народы Евразии по внешнему виду от других, является цвет кожи, которая в более солнечных тропических регионах приобретает темный шоколадный оттенок. И это не случайное совпадение обстоятельств. Хотя гены еще таят в себе немало загадок, мы можем утверждать, что темный цвет кожи, обязанный своим появлением пигменту меланину, определяется генетической памятью человека, которая формировалась в процессе длительной эволюции. Кожа людей, которые живут в тропиках и субтропиках, подвергается мощной атаке ультрафиолетовых лучей. В результате этого человек рискует пострадать от чрезмерного загара и покрыться волдырями, а также умереть от рака кожи. Лишь темный цвет кожи способен уменьшить риск и угрозу для здоровья человека. Среди других, хотя и не столь очевидных преимуществ, можно назвать то, например, что пигментация, в которой участвует меланин, позволяет коже более эффективно противостоять перегреву и излучать излишки тепла, а также препятствует разрушению в организме такого важного витамина, как фолиевая кислота.
Итак, в результате длившейся многие и многие поколения эволюции кожа людей в жарких климатических поясах стала темной. Именно темнокожие среди множества своих чад и домочадцев в областях повышенной солнечной активности живут дольше и чувствуют себя как нельзя более счастливыми[258]. В Северной Азии (т.е. в той части Азии, которая охватывает территории от Цзиньхай-Тибетского плато на севере и до Урала на востоке) и в Европе солнечный свет не столь интенсивен, и риск развития рака кожи менее выражен, зато население этих регионов повсеместно страдает от рахита и различных заболеваний костей, вызванных недостатком солнечного света. В самом Лондоне, причем уже в начале двадцатого столетия, от этих заболеваний умерло множество детей. Я сам был очевидцем подобных трагедий. Дело в том, что, попав как-то раз в больницу, а именно в отделение патологии Лондонского королевского госпиталя, я специально узнавал, насколько распространен рахит в наши дни. Во второй половине двадцатого столетия в Великобритании вновь получили широкое распространение такие заболевания, как рахит и остеомаляция[259], вторгшиеся в наши широты вместе с переселенцами с Индийского субконтинента. Именно дети потомственных индийцев чаще всего страдают рахитом. Это отчасти объясняется тем, что их темная кожа продолжает фильтровать солнечный свет, как это было необходимо на родине их предков, а не здесь, где солнечного света и без того недостаточно.
Итак, направление эволюции определяется двумя взаимосвязанными факторами естественного отбора, под влиянием которых сформировалось повсеместно наблюдаемое явление: по мере приближения к нулевым широтам цвет кожи местных жителей становится все более темным. Эти обусловленные солнцем изменения цвета кожи и волос вырабатывались из поколения в поколение в процессе длительной эволюции. Те факторы естественного отбора, которые определяют темный цвет кожи, очевидно, повлияли на генетический код жителей Африки. Если же покинуть Африку и отправиться путешествовать по обширным территориям Евразии, спускаясь изСкандинавии, на севере Европы, и Сибири, на севере Азии, к землям Италии и Юго-Восточной Азии, на которых расположены южные оконечности обоих континентов, можно наблюдать, как постепенно меняется цвет кожи и волос местных жителей.
Подобно африканцам, другие темнокожие народы живут преимущественно в солнечных тропических странах. Вполне очевидно, что мы столкнемся и с исключительными случаями, но они будут представлены потомками тех людей, которые в недавнем прошлом покинули родину и поселились в южных странах, их кожа будет еще слишком светла для этих широт (или наоборот, слишком темна), что и естественно, ведь процесс эволюции длится многие поколения. Таким примером-исключением может служить Австралия, которая заселена потомками недавних эмигрантов, поэтому жители этой страны по большей части до сих пор остаются «бледнолицыми». В Австралии, как нигде на земном шаре, высок процент людей, страдающих раком кожи, но и здесь уже начался постепенный процесс эволюции, который, по всей вероятности, ведет к тому, что потомки выходцев из Европы приобретут естественный для этих широт темный цвет кожи. И наоборот, первые переселенцы, отправившиеся с юга на север в страны Северной Европы и Азии, выглядели неестественно темными среди местных жителей, но со временем кожа их потомков стала более бледной. Однако оставим на время Австралию. Дело в том, что в какой бы точке Земли мы ни оказались, цвет кожи местных жителей будет связан с местным уровнем интенсивности ультрафиолетового излучения (рис. 5.1)[260].
 Рис. 5. 1
Поскольку эта особенность эволюционного развития постоянно оказывает влияние на человека во все эпохи и на всех широтах, наш цвет кожи способен поведать скорее о том, где жили наши предки в последние 10—20 тысяч лет, нежели об их генетической дивергенции за предыдущие 60 тысяч лет. Поэтому цвет кожи у современного человека имеет весьма ограниченную ценность как источник, позволяющий проследить древние маршруты миграции разных ветвей человечества после его исхода из Африки. Другими словами, цвет кожи — это далеко не самый надежный индикатор для изучения доисторической эры развития человечества, скажем, между 70 000 и 10 000 гг. до н.э. Может показаться странным, что цвет кожи столь изменчив, поскольку именно он является самым заметным и решающим среди всех «расовых» признаков. Ключом к дискриминационным установкам в поведении человека являются не логические доводы, а стремление отыскать какую-нибудь легко узнаваемую деталь или предлог, позволяющие объединить представителей своей группы и «отсечь» членов соперничающих групп. Различие между ними может носить самый тривиальный характер. Мы регулярно выдумываем все более абсурдные различия, например, различия между всевозможными религиозными сектами, чтобы оправдать преследования или истребление наших соседей.
Рис. 5. 1
Поскольку эта особенность эволюционного развития постоянно оказывает влияние на человека во все эпохи и на всех широтах, наш цвет кожи способен поведать скорее о том, где жили наши предки в последние 10—20 тысяч лет, нежели об их генетической дивергенции за предыдущие 60 тысяч лет. Поэтому цвет кожи у современного человека имеет весьма ограниченную ценность как источник, позволяющий проследить древние маршруты миграции разных ветвей человечества после его исхода из Африки. Другими словами, цвет кожи — это далеко не самый надежный индикатор для изучения доисторической эры развития человечества, скажем, между 70 000 и 10 000 гг. до н.э. Может показаться странным, что цвет кожи столь изменчив, поскольку именно он является самым заметным и решающим среди всех «расовых» признаков. Ключом к дискриминационным установкам в поведении человека являются не логические доводы, а стремление отыскать какую-нибудь легко узнаваемую деталь или предлог, позволяющие объединить представителей своей группы и «отсечь» членов соперничающих групп. Различие между ними может носить самый тривиальный характер. Мы регулярно выдумываем все более абсурдные различия, например, различия между всевозможными религиозными сектами, чтобы оправдать преследования или истребление наших соседей.
Изменение формы и черт лица
Другие, более существенные изменения, такие, например, как форма нашего лица, обусловлены различиями лицевых костей черепа. Эти лицевые кости претерпели существенные изменения у жителей Восточной, Юго-Восточной и Южной Азии, что является отражением достаточно длительного периода, прошедшего после разделения населения этих регионов на разные ветви. На всей территории Восточной Азии мы встречаем монголоидный тип (см. табл. 16) с характерной для него дополнительной складкой, так называемым эпикантусом[261], защищающим верхнее веко, а также широкими скулами и черепом. Этот тип часто делится на два подтипа: северных монголоидов и южных монголоидов (см. рис 5.2), причем для последних характерен менее выраженный эпикантус; в том числе южно-китайский и более смуглокожий монголоидный типы, распространенные в Юго-Восточной Азии[262]. В Южной Азии, в частности — в Индии, можно встретить множество самых разнообразных этнических групп. Большинство индусов, несмотря на темно-смуглый цвет кожи, по телосложению и прочим антропологическим параметрам гораздо ближе к европейцам и жителям Ближнего Востока, чем к обитателям Южной Азии. Европейцев, имеющих длинный узкий череп, круглые глаза и бледную кожу, иногда называют представителями кавказского типа. Чем дальше мы продвигаемся на север Индии и Пакистана, тем ближе жители этих районов к «кавказоидам» — обитателям Леванта и ближневосточного региона. Напротив, в южной Индии преобладают представители смуглокожего, курчавого и круглоглазого этнического типа. В восточной Индии, Ассаме и Непале чаще встречаются люди с более выраженными монголоидными чертами внешности. В главе 4 мы рассматривали другой тип обитателей Азии, которые, возможно, являются потомками племен, селившихся вдоль древнего прибрежного пути в Австралию. По всему побережью Индийского океана, в Пакистане и Индии, на Андаманских и Филиппинских островах, а также в Малайзии сохранились отдельные, так называемые аборигенные, этнические группы, которые по внешнему облику более напоминают африканцев и жителей Новой. Гвинеи. У них очень темный цвет кожи и характерные курчавые волосы. Эти этнические меньшинства принято называть соответственно негроидами, негритосами и меланезийцами. Хотя эти аборигенные группы и составляют ничтожно малое меньшинство среди прочих великих и малых народов азиатского Старого Света, о которых я уже упоминал, они, по мнению многих ученых, в физическом и антропологическом отношении гораздо ближе к первым племенам-выходцам из Африки, чем к другим народам Азии. Это сходство убедительно свидетельствует о тех огромных изменениях, которые претерпело подавляющее большинство этносов Азии. Изучение строения зубов у тех же аборигенных групп населения, которое я использовал в качестве генетических индикаторов (глава 4), позволило получить целый ряд антропологических аргументов в пользу этой точки зрения. Другим подтверждением того, что эти аборигенные группы представляют собой реликты древних обитателей побережья и островов, является тот факт, что ископаемые останки черепов, найденные в Европе, Восточной и Юго-Восточной Азии и восходящие к эпохе до последнего Ледникового периода, по форме нельзя отнести ни к кавказоидному, ни к монголоидному типу. Вместо этого мы видим формы, близкие к древнейшим черепам африканского и ближневосточного типа, возраст которых составляет 100 тысяч лет[263].
Древние люди были более рослыми и крепкими
Известно ли нам, как могла выглядеть первая группа эмигрантов-переселенцев из Африки? Проще всего ответить, что нет. Наиболее естественное и правдоподобное предположение о том, что они были темнокожими и имели курчавые волосы, основано на внешности современных африканцев[264], хотя весьма маловероятно, что африканцы, жившие 80—100 тысяч лет тому назад, выглядели, как их современные потомки. И тем не менее ряд ключей к этой проблеме все же существует. До нас дошло несколько ископаемых черепов людей современного (в анатомическом смысле) типа, возраст которых — около 100 тысяч лет. Один из них, который часто называют типичным примером черепов того периода, найден в районе первого неудавшегося исхода из Африки на Левант (Ближневосточный регион). Он был назван Скхул 5 — по названию пещеры, где он был обнаружен. В тех же местах был найден еще ряд аналогичных черепов, так что Скхул 5 — всего лишь наиболее хорошо сохранившийся из них. Череп типа Скхул использовался для создания реконструкции «лица Евы» (см. Фото 17) в нашем документальном фильме «Истинная Ева». Черепам людей анатомически современного типа того периода присуща широкая вариативность, но они обладают несколькими общими особенностями, которые претерпели изменения в последующих поколениях, в том числе и у современных африканцев. Наиболее характерными из них являются крупный рост и «крепость» черепа. В числе факторов, обусловивших крепость, — крупные, покатые черепа, толстые кости и тяжелые надбровные дуги, резко отличающие древних людей от современного человека. В отличие от «крепкого» типа, так называемый «стройный» вариант используется для описания людей с плоским, высоким черепом, вертикальным лбом, более плавными надбровными дугами и сравнительно более тонкими костями конечностей и черепа. В числе других наследственных признаков черепов эпохи, предшествовавшей южному исходу, можно назвать относительную узость черепа (меньшее расстояние от виска до виска по сравнению с расстоянием между лбом и затылком) и более широкую верхнюю часть лица[265].
 Фото 17. Так выглядела Ева выходцев из Африки. Реконструкция, использованная в документальном фильме «Путь из Эдема»/«Истинная Ева». Реконструкция воссоздана по одному из наиболее хорошо сохранившихся черепов эпохи исхода на Левант. Ее черты отражают толстокостный тип, типичный для той эпохи, относительно узкий череп и широкие скулы и лоб.
Фото 17. Так выглядела Ева выходцев из Африки. Реконструкция, использованная в документальном фильме «Путь из Эдема»/«Истинная Ева». Реконструкция воссоздана по одному из наиболее хорошо сохранившихся черепов эпохи исхода на Левант. Ее черты отражают толстокостный тип, типичный для той эпохи, относительно узкий череп и широкие скулы и лоб.
Крупный рост и крепость скелета тесно связаны друг с другом (это в особенности относится к черепу). Это ставит перед нами серьезную проблему, когда мы пытаемся воспользоваться костями для реконструкции человека доисторического периода, поскольку наиболее заметным изменением со времен исхода народов из Африки явилось все более нарастающее снижение роста и крепости телосложения у всех народов мира, в том числе — африканцев. Хотя некоторые из устойчивых изменений носят, по-видимому, генетический характер, наиболее резко выраженное снижение роста и крепости телосложения у человека за последние 10 тысяч лет обусловлены, как ни парадоксально это звучит, особенностями питания, а не генетическими факторами.
Почему мы стали более мелкими?
Древние охотники и собиратели вели суровую, исполненную опасностей жизнь, и поэтому плотность населения была крайне низкой, но зато их рацион был очень богат белками и минеральными веществами, а также всевозможными овощами и фруктами. Наоборот, хотя земледельцы за последние 8000 лет сумели добиться более высокой урожайности зерновых с гектара и имели большие семьи в результате раннего отнятия детей от груди, их рацион отличался однообразием. Их пища была богата углеводами, но скудна белками, витаминами и кальцием. Раннее отнятие от груди и переход на пищу, богатую углеводами и бедную белками, приводил к задержкам в росте, которые сохранялись и во взрослом возрасте. Следовательно, хотя с развитием земледелия численность населения значительно возросла, рост и физическая выносливость детей и взрослых резко уменьшились. Этот эффект особенно заметен у народов, чей рацион изобилует рисом, но крайне скуден животными белками, примером чему могут служить народы Дальнего Востока. Сегодня мы имеем возможность наблюдать, как сказывается обратное изменение этой культурно-обусловленной формы недоедания. Когда уроженцы стран Восточной Азии, основой питания которых на родине был рис, эмигрировали в США и переходили на совершенно иной — высококалорийный, богатый белками — рацион питания, дети бывших азиатов уже через два поколения почти догоняли в росте американских детей. Известно, что после Второй мировой войны дети жителей Европы и Америки благодаря высококалорийному питанию превосходят своих родителей в росте и физических данных[266]. Таким образом, если крепость телосложения до некоторой степени зависит от пропорций тела, а средние пропорции жителей различных регионов, в свою очередь, зависят от их сегодняшнего рациона питания, получается, что роль этих двух важнейших составляющих нашей внешности в формировании различий между неафриканскими народами за последние 70—80 тысяч лет заметно снизилась. Кроме того, многие негативные изменения крепости, пропорций тела, а также внешних данных современных людей со времени исхода из Африки вполне могут быть выправлены с помощью полноценного рациона и здорового образа жизни. Именно этим объясняется типично «неандертальский» облик некоторых современных «звезд» жестких контактных видов спорта. Со временем формируются устойчивые изменения, среди которых особенно выделяются два фактора: изменения черепа и зубов. Прежде всего надо отметить, что уменьшение размеров черепа в разных регионах было обусловлено различными факторами.
Изменения черепа
Хотя аборигены Австралии и в меньшей степени некоторые племена Новой Гвинеи тоже претерпели негативные изменения — уменьшение роста и пропорций тела, — они, оказывается, сохранили значительно большую крепость скелета, чем практически все остальные народы мира. Уменьшение величины черепа у австралоидов затронуло как его высоту, так и ширину. Аборигенов Австралии иногда называют лучшим примером сохранения архаических наследственных форм, однако еще более полноценными «консерваторами» являются жители Новой Гвинеи, что объясняется целым рядом причин. Что касается аборигенов Австралии, как в прошлом, так и в настоящем, то некоторые из присущих им черт остаются необъяснимыми. Одна из них заключается в том, что древнейшие ископаемые останки черепов, найденные в Австралии, принадлежали к «стройному», а не к «крепкому» типу. Другая причина, позволяющая считать черепа австралоидов несколько «изменившимися» по сравнению с архаическим типом, заключается в том, что у большинства аборигенов Австралии в наши дни волосы не вьющиеся, а курчавые[267]. В наши дни существует всего лишь одна этническая группа, сохранившая такой же уровень крепости черепа, как у австралийцев и жителей Новой Гвинеи. Это — племя тьерра дель фуэган в Южной Америке, сегодня уже практически вымершее. В Японии найден другой череп «крепкого» типа: это знаменитые ископаемые черепа народности минатогава с острова Окинава, субтропического японского острова, датируемые — судя по высоте — последним Ледниковым периодом (см. Фото 18). Эти черепа весьма близки по форме к черепам пренеолитической этнической группы йомонов, также живших на Японских островах, — тех самых йомонов, которые считаются предками айнов, современного коренного населения Северной Японии[268]. У предков европейцев и жителей Среднего Востока (Средней Азии) наблюдались ограниченные и характеризующиеся широкой вариативностью изменения длины и крепости черепа. К ним относятся длинноголовость (дилихоцефалия), а также значительное снижение крепости черепа, что помогает объяснить такую деталь, как мои собственные насупленные брови[269].
 Фото 18. Череп Минатогава I с о.Окинава, датируемый последним ледниковым максимумом, вполне может считаться останками древнейших предков аборигенов Японии — айнов.
Фото 18. Череп Минатогава I с о.Окинава, датируемый последним ледниковым максимумом, вполне может считаться останками древнейших предков аборигенов Японии — айнов.
Наиболее же значительные изменения отмечены у народов, которых в наши дни принято называть монголоидами. У них уменьшение величины черепа приняло характер заметного сокращения расстояния между лбом и затылком при сохранении ширины и высоты черепа. Другое важное изменение, характерное для монголоидной расы, — подчеркнуто плоский тип лица. Эта особенность наиболее четко прослеживается на черепах людей эпохи неолита, живших на восточном берегу озера Байкал в Сибири, а также по всей Монголии (рис. 5.2). Как было установлено, выраженно плоский характер лица не является принципиально новой чертой в общем диапазоне вариативности форм человеческого черепа. Плоское лицо — правда, в менее выраженном виде — характерно для некоторых ранних черепов древнейших жителей Африки, для современных охотников и собирателей народности хойсан, а также для достопочтенного отца-основателя[270] Южно-Африканской Республики после падения режима апартеида. Подобные изменения привели к появлению и распространению высокого черепа с тонкими чертами (брацицефалического типа), преобладающего в наши дни в Восточной Азии, что имеет под собой уже четко сложившуюся генетическую основу. Жители Восточной Азии относятся к числу обладателей самых «стройных» черепов среди современного человечества[271]. Когда же и где возникли и получили распространение физические изменения, характерные для современного облика представителей монголоидной расы? Конкретная дата появления в Китае первых людей современного типа вызывает споры, однако это, скорее всего, произошло около 70 тысяч лет назад. При этом важно помнить, что первые обитатели Китая современного типа не имели ничего общего с внешностью монголоидного типа и что немногочисленные ископаемые останки черепов, относящиеся к эпохе до последнего Ледникового периода и найденные в равнинном Китае, возраст которых составляет около 20 тысяч лет, относятся к архическому «крепкому» типу. Фактически же возраст самых древних в мире останков бесспорно монголоидного типа составляет всего 10 тысяч лет. Хотя эти останки были найдены на Дальнем Востоке, самые древние останки человека современного типа с монголоидными чертами были найдены далеко на западе, в урочище Афонтова Гора II на берегах реки Енисей на юге Сибири, что к западу от озера Байкал. Поскольку эта изолированная находка вызывает целый ряд вопросов и других черепов, относящихся к доледниковому периоду, в Южной Сибири не найдено, все, что можно сказать об этом единичном ископаемом черепе без ссылки на другие источники, — это то, что локусом изменений в направлении монголоидного типа не обязательно был Китай[272].
 Рис. 5.2
Рис. 5.2
Эволюция зубов
Другими, не столь заметными, но не менее важными изменениями явились изменения в форме зубов. Сохранение некоторых архаических особенностей скелета у современных европеоидов стало причиной проведения целого ряда детальных дентологических исследований (рис. 5.3), которые показали, что европейцев тоже можно поставить в один ряд с обитателями прибрежной полосы Индийского океана (включая жителей Южной Индии, негритосов племени семанг (Малайя) и коренное население Новой Гвинеи), сохранившими куда больше архаических африканских особенностей строения зубов и черепа, чем представители любых других народов за пределами Африки. Эта заметная близость в строении зубов у европейцев и других неафриканских народов региона Индийского океана, которым присущи «наименее выраженные изменения», видимо, является следствием одного южного исхода. На мой взгляд, это свидетельство — еще один гвоздь в гроб (если таковой еще нужен) расистских стереотипов, объявляющих европейцев высшей расой, а аборигенов — примитивными низшими существами[273]. К этим «недифференцированным по дентальным признакам обитателям прибрежной полосы» с точки зрения дентального анализа весьма близки представители обширной группы народов. В ее состав входят аборигены Юго-Восточной Азии, принадлежащие к несемангскому (т.е. не связанному с негритосами) типу, в частности — малайцы, представители коренного населения Малайского полуострова, различные архаические этнические группы Тихоокеанского побережья, такие, как аборигены-айны в Японии, а также многочисленные полинезийские племена. Все эти группы сохранили те или иные признаки сходства с представителями ранней формы строения зубов — сундадонтами, название которых происходит от названия геологического района в Юго-Восточной Азии — шельфа Сунды (Сундаленд), где, как предполагается, возник и сформировался этот тип. Сундадонты не встречаются далее к западу вдоль прибрежной полосы, например, в Индии. Монголоидные черты выделяются здесь в особый тип, который можно классифицировать как южных монголоидов. Народности минотагава (палеолит) и йомоны в Японии, которые могут считаться предками современных айну (айнов), также имеют в строении зубов черты, сближающие их с сундадонтами Юго-Восточной Азии и племенами островов Тихого океана[274].
 Рис. 5.3
Особое место в этом ряду (см. рис. 5.3) принадлежит аборигенам Австралии, которые, в отличие от своих соседей из Новой Гвинеи, сохраняют некоторые черты, свойственные сундадонту. Эта тенденция свидетельствует о возможности промежуточного смешения[275].
Вторая из двух крупнейших дентологических групп Евразии, помимо сундадонтов, — это синодонты (букв. «китаезубые»). Для этнических групп синодонтов характерно сохранение некоторых особенностей строения зубов, присутствовавших у сундадонтов, а также общее снижение величины зубов. Синодонтность присуща всем северным монголоидам, обитающим в Центральной Азии, причем ее характерные свойства становятся более выраженными при продвижении на север, что относится и к жителям Америки, переселившимся туда из Азии через Берингов пролив (см рис 5.2). Для Юго-Восточной Азии синодонтность даже в наше время является нехарактерной.
Прибегнув к сильному упрощению, можно сказать, что сундадонтность отражает древнейшие изменения в строении зубов по сравнению с собирателями на прибрежной полосе — выходцами из Африки, и разнообразные комбинации этих черт можно встретить у представителей южной ветви монголоидов, обитающих в Юго-Восточной Азии, Полинезии и Микронезии, и в меньшей мере у некоторых немонголоидных народностей, таких, как йомоны в Японии и изолированные этнические группы жителей Андаманских островов. Синодонтность представляет собой следующий этап дивергенции в том же морфологическом направлении, что и сундадонтность. Она встречается у северной ветви монголоидов, включая и большинство китайцев, и в более выраженном и гипертрофированном варианте — у жителей Северо-Восточной Сибири и коренных жителей Америки[276].
Рис. 5.3
Особое место в этом ряду (см. рис. 5.3) принадлежит аборигенам Австралии, которые, в отличие от своих соседей из Новой Гвинеи, сохраняют некоторые черты, свойственные сундадонту. Эта тенденция свидетельствует о возможности промежуточного смешения[275].
Вторая из двух крупнейших дентологических групп Евразии, помимо сундадонтов, — это синодонты (букв. «китаезубые»). Для этнических групп синодонтов характерно сохранение некоторых особенностей строения зубов, присутствовавших у сундадонтов, а также общее снижение величины зубов. Синодонтность присуща всем северным монголоидам, обитающим в Центральной Азии, причем ее характерные свойства становятся более выраженными при продвижении на север, что относится и к жителям Америки, переселившимся туда из Азии через Берингов пролив (см рис 5.2). Для Юго-Восточной Азии синодонтность даже в наше время является нехарактерной.
Прибегнув к сильному упрощению, можно сказать, что сундадонтность отражает древнейшие изменения в строении зубов по сравнению с собирателями на прибрежной полосе — выходцами из Африки, и разнообразные комбинации этих черт можно встретить у представителей южной ветви монголоидов, обитающих в Юго-Восточной Азии, Полинезии и Микронезии, и в меньшей мере у некоторых немонголоидных народностей, таких, как йомоны в Японии и изолированные этнические группы жителей Андаманских островов. Синодонтность представляет собой следующий этап дивергенции в том же морфологическом направлении, что и сундадонтность. Она встречается у северной ветви монголоидов, включая и большинство китайцев, и в более выраженном и гипертрофированном варианте — у жителей Северо-Восточной Сибири и коренных жителей Америки[276].
Загадка происхождения монголоидов
При изучении формы черепа, строения зубов или других генетических индикаторов нельзя не заметить, что среди всех этнических групп за пределами Африки наиболее сильные изменения со времени исхода затронули именно монголоидную расу. Эта дифференциация по сравнению с древнейшим архаическим прототипом и утрата внутригруппового разнообразия заметно усиливается по мере продвижения на крайний Северо-Восток Азии и далее — в американскую Арктику. Главный вопрос, остающийся неясным в наши дни, — как, когда и где началось активное формирование черт, выделяющих представителей монголоидной расы по сравнению с другими народами Евразии. И это — не праздный схоластический вопрос, занимающий лишь академические круги. В конце концов, именно монголоиды — наиболее многочисленная и распространенная раса на земном шаре. Ее представители появились в Америке еще до последнего Ледникового периода, примерно 20 тысяч лет тому назад (см главу 7), однако что касается равнинных районов Китая, то там этот тип появился лишь после того, как была перевернута последняя страница в истории последнего оледенения. Краткое резюме этой сложной и запутанной истории в нашем изложении выглядит так: 1. Древнейшие обитатели Азии (которые были также и предками европейцев) внешне выглядели практически так же, как и африканцы той эпохи, и имели сходное строение зубов. 2. Эти древнейшие жители Южной Азии, обладавшие «крепким» черепом, впервые появились около 80 тысяч лет тому назад и расселились по всей прибрежной полосе Южной Азии, от побережья Аравийского полуострова через Индию в Юго-Восточную Азию, выйдя к берегам Новой Гвинеи в конце дуги островов на юго-западе Тихого океана. Современные жители районов вдоль этого маршрута расселения сохранили некоторые архаические черты строения зубов, характерные для их далеких предков. 3. Из Индокитая представители этих недифференцированных типов расселились на север, в Китай, в период между 35 и 70 тысячами лет тому назад обогнув западное побережье Тихого океана. 4. В некий исторический момент еще до последнего Ледникового периода среди представителей «премонголоидной расы», живших в Юго-Восточной Азии, сформировался новый комплекс особенностей строения зубов. Эти новые типы строения зубов впоследствии в несколько видоизмененной форме распространились на северо-восток, вдоль побережья Тихого океана, и дальше — в Японию (их носители стали предками современных йомонов и айнов), а также на юго-восток — в Полинезию. (См. рис 5.2 ) 5. Второй основной комплекс признаков строения зубов, синодонтность, по всей видимости, является более поздним и происходит от сундадонтности. Эволюция от сундадонтности к синодонтности и связанные с ней промежуточные формы не выявлены ни в Юго-Восточной Азии, ни на западной окраине Тихого океана, ни среди йомонов или айнов в Японии. Однако степень синодонтности постепенно нарастает по мере продвижения на север у современного монголоидного населения континентальных районов Восточной Азии, достигая своего крайнего выражения у коренных жителей Америки. Таким образом, получается, что источником и предшественником синодонтности была сундадонтность, возникшая в южных районах. В связи с отсутствием ископаемых окаменелостей признаки синодонтности на черепах старше 10 тысяч лет, найденных в Китае и на Японских островах, обнаружить не удалось. 6. Монголоидной форме черепа присущи тонкие «удлиненные» кости, характерное плоское лицо и уменьшение длины. Подчеркнуто плоский нос обнаружен у одного из черепов, относящихся к эпохе до последнего Ледникового периода, найденном в районе возле озера Байкал. Плоский нос наиболее выражен у черепов в этом районе. 7. Как и синодонтность, признаки монголоидной расы на китайских черепах, относящихся к эпохе до последнего Ледникового периода, не выявлены. В более поздние времена они получили широкое распространение в Северной и Восточной Азии и считаются характерной особенностью синодонтов, то есть, другими словами, северной ветви монголоидов, а также большинства коренных народностей Америки. Таким образом, нарастающие изменения формы черепа протекали параллельно с переходом от сундадонтности к синодонтности в строении зубов. Если предками синодонтов действительно были выходцы с юга — сундадонты, то по крайней мере некоторые черты физического облика, характерные для северной ветви монголоидов, также восходят к южным предкам. Если же предки первых типичных монголоидов северной ветви были выходцами с юга, это означает, что они имели общего предка с такими народами, как малайцы — коренные обитатели Малайского полуострова, которые до сих пор живут там. Как мы увидим далее, существуют вполне убедительные свидетельства того, что по крайней мере некоторые основные генетические цепочки Восточной и Северо-Восточной Азии берут свое начало из Индокитая, сохраняя расовые черты и признаки, которые и в наши дни можно встретить у коренных представителей южной ветви монголоидов в этом регионе. В этой связи сделать паузу и рассмотреть различные частные случаи этой модели миграции «юг — север», представляющей собой противоположность модели «север — юг», что прослеживается по изменениям строения зубов и костей. Из двух основных азиатских типов строения зубов, ассоциируемых с представителями монголоидной расы, сундадонтность, по всей видимости, возникла первоначально в Юго-Восточной Азии, представляя собой древнейший тип изменения строения зубов, который и в наши дни встречается у некоторых реликтовых архаических этнических групп немонголоидного происхождения. Синодонтность же, напротив, является самым распространенным типом строения зубов во всей Восточной Азии и Америке, однако она не встречается у представителей коренного населения Юго-Восточной Азии. Последний аргумент говорит в пользу версии направления миграции «юг—север». А как же быть с гипотезой о миграции с севера на юг? Если она справедлива и корни происхождения южной ветви монголоидов именно таковы, мы вправе были бы ожидать массового вторжения носителей синодонтных признаков с севера на юг, в Юго-Восточную Азию. Однако на самом деле ничего подобного не обнаружено. Простой анализ уровня изменений также опровергает теорию о том, что специфические типы строения зубов привели сначала к возникновению сундадонтности на юге, а затем трансформировались в синодонтность, когда носители монголоидных признаков направились в Центральную Азию. Что касается эволюции и усиления признаков сундадонтности, что повлекло за собой формирование синодонтного типа, некоторые признаки монголоидной расы, например характерная плоскостность лица, более выражены у северной ветви монголоидов, чем у южных монголоидов. Таким образом, какими бы ни были движущие силы эволюции, эти специфические изменения должны были накапливаться где-то еще дальше к северу, возможно, в глубине континентальных районов Азии. Ископаемые останки (или, точнее сказать, их отсутствие) свидетельствуют о том, что район «дальнейшего развития» монголоидных признаков находился отнюдь не в Китае, Японии, Юго-Восточной Азии или Индии, а где-то в Центральной Азии к северу от Гималаев, например, в Монголии или в районе озера Байкал в Южной Сибири. Более того, на юге Сибири существует особая этническая группа, среди представителей которой, кстати сказать, резко отличающихся от соседних народностей, встречается немало носителей сундадонтных признаков, что свидетельствует о сохранении в этих местах архаического расового типа (см. рис. 5.3).
Почему представители монголоидной расы стали выглядеть иначе?
Существует целый ряд теорий, объясняющих, как и где начали формироваться особенности скелета монголоидов, обусловившие их внешние отличия. Первое, что необходимо сказать об этих теориях, — это то, что все они носят откровенно спекулятивный и даже спорный характер. Теории, рассматривающие вопрос «где», имеют, как я уже отмечал, чисто гипотетический характер, что объясняется практически полным отсутствием ископаемых останков представителей монголоидной расы. Недостоверность же теорий, отвечающих на вопрос «как», обусловлена тем, что сегодня не существует реальных средств, позволяющих связывать те или иные изменения скелета с конкретными генами-«контролерами». Впрочем, это не мешает нам хотя бы вкратце познакомиться с такими теориями. К сожалению, как это часто случается в научных дебатах, противоречия между сторонами приводят к возникновению настоящей пропасти между ними, тогда как истина обычно находится где-то посередине. Что касается теорий о происхождении, то нам предлагается выбор между южным и северным локусами прародины предков монголоидов, а если говорить о теориях, объясняющих, как могли произойти подобные изменения, нам приходится обращаться к адаптирующей физической эволюции, при которой возможны либо изолированное развитие, либо дрейф (миграция) генов. (Впрочем, между двумя предполагаемыми механизмами изменений существует определенная связь, поскольку адаптационные[277] теории рассматривают именно холодную северную зону в качестве среды, где имела место адаптация.) Географическое деление монголоидов на северян и южан становится еще более заметным при рассмотрении физических (и генетических) факторов, как старых, так и выявленных совсем недавно, свидетельствующих о том, что единая некогда раса действительно разделилась на две ветви — северных и южных монголоидов. Изучение динамики роста у детей в Китае показало, например, что дети жителей Гонконга (Южный Китай), несмотря на то что питались они гораздо лучше детей из континентального Китая, заметно уступали в росте детям из Пекина (Северный Китай). Тот факт, что дети из Гонконга, питающиеся лучше своих сверстников из Пекина, значительно уступают им в росте даже спустя несколько поколений после экономического бума в Гонконге, свидетельствует о том, что между севером и югом сложились устойчивые генетические различия[278].
Являются ли расовые отличия монголоидов результатом дрейфа генов или целенаправленного отбора?
Основную альтернативу традиционным гипотезам о происхождении монголоидов, рассматривающим проблему, «как» они возникли, сформулировать достаточно несложно. Наиболее простой ответ на этот вопрос (рассматриваемый в последней главе) сводится к тому, что немногочисленная группа-основатель, изолированная от прочих этнических групп, самостоятельно эволюционирует, приближаясь к определенному генетическому или физическому типу. Комбинированное воздействие эффектов группы-основателя и накапливания изменений еще более усиливается в результате сексуального отбора. Чтобы упростить этот пример еще более, буквально до абсурда, допустим, что в группе-основателе монголоидной расы, пришедшей с юга, у большинства ее представителей по чистой случайности оказались маленькие носы. В результате подобной групповой самоидентификации и предпочтения, отдаваемому при выборе партнеров лицам с маленькими носами, курносые носы через какое-то время стали основной отличительной чертой внешности членов группы. Это еще более усилило действие эволюционных сил, способствовавших формированию маленьких носов, и у дальних потомков этой группы-основателя носы стали совсем крошечными. В дрейф генов оказались вовлечены и другие факторы, ассоциируемые в генетическом отношении с маленькими носами, например плоские лица. Другая теория, более старая и, возможно, более спорная, заключается в том, что основателям монголоидной расы пришлось адаптироваться к куда более неблагоприятным и сложным условиям новой среды обитания, в данном случае — к холодному и ветреному климату азиатских степей к северу от Гималаев. Эта идея имеет давнюю родословную, но удачнее всех, пожалуй, ее сформулировал американский биолог Дэйл Гатри. Гатри увлеченно описывал среду обитания, которую он называл Мамонтовыми степями Азии эпохи палеолита. Мамонтовая степь представляла собой необъятные пространства, поросшие высокими травами; сердцем этой степи был обширный район, расположенный к северу от 35-й параллели, Гималаев и Тибетского нагорья. Этот комплекс факторов низкотемпературной среды обитания, известный биологам под названием «палеолитический биом», прекратил существование около 10 тысяч лет тому назад, являя собой парадоксальный результат глобального потепления, а вместе с ним навсегда исчезли и множество крупных и мелких видов флоры и фауны[279]. По мнению Гатри, в южной части «сердца» Мамонтовой степи очень рано появились люди, пришедшие сюда из Синьцзяна, Монголии и окрестностей озера Байкал в Сибири. Сегодня в этих местах раскинулись обширные площади пустынь. В эпоху расцвета великая Мамонтовая степь простиралась от побережья Атлантики на западе до острова Хоккайдо на востоке. Это была самая крупная травянистая саванна, когда-либо существовавшая на нашей планете, и в ней спокойно паслись неисчислимые стада крупных травоядных. Небольшие стада некоторых из этих травоядных дожили до наших дней в труднодоступных районах. К числу этих животных принадлежат дикая лошадь, северный олень, мускусный бык и антилопа сайгак, Что же касается более крупных видов, таких, как волосатый мамонт, степной бизон и волосатый носорог, то они давно вымерли. Понятно, что Мамонтовая степь была поистине раем для охотников, которые научились преодолевать холода и сильные ветры. Гатри пишет, что люди эпохи палеолита, охотившиеся на травоядных животных в этой степи, предпочитали селиться вдоль южной ее границы, проходившей примерно по 40-й параллели, где было больше островков леса, дававших людям приют и укрытие (см. рис. 5.2). Нет никаких сомнений, что люди, жившие в столь холодных климатических условиях, нуждались в теплой одежде. Гатри и другие исследователи высказывают предположение, что выжить в суровых условиях будущим монголоидам помог целый ряд факторов физической адаптации. В их числе — двойное верхнее веко, или уже упоминавшаяся складка эпикантус, позволявшая уменьшить потери тепла через глаза, более толстый слой подкожного жира вокруг глаз и на щеках, скулах и подбородке; маленький нос, снижающий риск обморожения; более округлая форма головы и особое строение черепа, позволяющее организму терять меньше тепла; более плотное и приземистое туловище с относительно короткими конечностями; более равномерное распределение жира, что, вместе с меньшей длиной конечностей, обеспечивало снижение на 10% отношение поверхности тела к его объему; бледная, близкая по тону к европейцам кожа, обеспечивающая поглощение солнечных лучей и выработку достаточного количества витамина D для защиты от рахита; и, наконец, повышенная чувствительность кровеносных сосудов в конечностях, позволяющая избежать обморожения. Лично я, несмотря на то что целый ряд аргументов Гатри представляется мне весьма вероятными, нахожу их перечень чрезмерно обширным, да и аргумент о величине носа не кажется мне убедительным. Ведь в Европе, где климатические условия практически такие же, как современные ее жители, так и неандертальцы использовали и используют большие носы как своего рода кондиционеры для согревания и увлажнения воздуха перед тем, как он попадет в весьма чувствительные бронхи и легкие. Это представляется куда более убедительным аргументом в пользу появления крупных носов, и подобные гипотезы высказывались как минимум дважды независимо друг от друга. В самом деле, разве может маленький нос быть столь же приспособлен к жизни в холодном климате? Между тем именно у эскимосов и в меньшей степени — чукчей, жителей крайнего северо-востока Сибири, развился нос, который принято называть «приплюснутым»: крошечный нос, буквально вдавленный в плоское лицо[280]. Дальнейший процесс адаптации монголоидов к условиям жизни на Крайнем Севере свидетельствует, что крошечный нос оказался не слишком эффективным «кондиционером», и климат Арктики потребовал его дальнейшей эволюции. Наиболее достоверным (под словом «достоверность» я имею в виду обоснованность теоретических построений, опирающихся на реальные факты) в рассуждениях Гатри мне представляется место, где он утверждает, что наиболее активно адаптация протекала на севере, где у северной ветви монголоидов наблюдается сравнительно бледная кожа, а самые выраженные изменения внешнего облика, в частности появление особенно плоского лица, наблюдаются в окрестностях озера Байкал как в глубокой древности, так и у современных коренных жителей этого региона[281]. Гатри высказал ряд предположений относительно того, сколь долгим мог быть процесс развития адаптационных изменений и когда он начался. Он вполне обоснованно полагает, что наибольшие изменения подобного рода вызвал последний Ледниковый период, достигший своей кульминации около 20 тысяч лет тому назад, создав так называемый стресс холода, но говорит, что сам процесс мог начаться как минимум 40 тысяч лет тому назад. Аргументы в поддержку столь раннего начала сводятся к тому, что, как мы уже говорили, накапливание изменений и признаков плоского лица началось у будущих монголоидов, живших к западу от озера Байкал, задолго до начала Ледникового периода (см. рис. 5.2). Хотя я должен признаться, что теоретические построения Гатри относительно эволюционного развития представляются мне весьма соблазнительными, меня тревожит проблема отсутствия реальных, материальных доказательств подобного рода — ископаемых останков человека. Кроме того, нерешенным остается и вопрос о том, каким образом у представителей южной ветви монголоидов, живущих в тропиках Юго-Восточной Азии, могли появиться эти черты внешности, если эти люди, как принято считать, являются потомками оседлого коренного населения. Если весь комплекс признаков, общих для северной и южной ветвей монголоидов, в первую очередь обусловлен адаптацией к холодным климатическим условиям, в таком случае, согласно теории миграции с юга на север, он должен был бы отсутствовать у представителей южной ветви монголоидов. Позволю себе привести почти анекдотический пример. Моя дражайшая жена, предки которой были уроженцами южного побережья Китая и которая по классификации, используемой мною в этой книге, относится к южным монголоидам, переносит холод куда хуже, чем я, человек, предки которого были южанами! Если же теории о миграции с юга на север и адаптации к холодному климату соответствуют действительности, нам пришлось бы признать, что первоначальные физические изменения могли явиться результатом действия некоего иного эволюционного механизма, чем простая адаптация к холоду, и что некоторые из этих изменений впоследствии были заметно усилены под воздействием холода.
Неотения у человека
Мне представляется весьма убедительным предложенное Гатри объяснение воздействия эволюционных факторов на севере, но не менее правдоподобным выглядит и продвижение с юга на север, подтверждаемое генетическими данными и особенностями строения зубов. Между тем существует и другой эволюционный феномен, способный связать воедино два этих фактора. Много лет назад видный палеонтолог Стивен Джэй Гоулд выдвинул интересную гипотезу, заключающуюся в том, что комплекс анатомических изменений, характерных для монголоидов, можно объяснить феноменом неотении — явления, при котором инфантильная или детская форма тела сохраняется во взрослом возрасте. Неотения у человека является одним из наиболее простых объяснений того, как и почему у людей за последние несколько миллионов лет так быстро развился непропорционально большой мозг. Относительно большой мозг и обращенность черепа вперед на позвоночнике, исчезновение волосяного покрова на всем теле, характерные для человека, наблюдаются у внутриутробных, неродившихся детенышей шимпанзе. Гоулд обратил внимание на постепенное усиление признаков неотении у монголоидов и даже предложил для этого явления особый термин — «педоморфия»[282]. Такой механизм, по всей видимости, включает в себя всего один ген-«контролер», и поэтому подобный эффект может быть достигнут за относительно короткий промежуток эволюции. Он же вполне может послужить объяснением появления совершенно нелогичного курносого носа и исчезновения основной части волосяного покрова на лице. Существуетнесколько эволюционных механизмов, в результате которых особи с выраженным педоморфизмом могут со временем произвести на свет сравнительно больше потомства, что объясняется дрейфом генов или направленным генетическим отбором, причем оба этих фактора не являются взаимоисключающими. Один из этих механизмов может быть обусловлен генами экономичности, использующимися в холодной степи (сокращение излишней мышечной массы, уменьшение объема зубов, более тонкие кости, меньший рос и физические данные). Это соответствует избирательно-адаптационной модели эволюции монголоидов, которая могла включать в себя уже существующие признаки, возникшие и оформившиеся у сундадонтов, которые мигрировали с юга на север вдоль побережья Тихого океана. Точно так же женщины с педоморфными признаками могли считаться более привлекательными, а это уже аргументы в пользу гипотезы полового подбора и дрейфа генов. И хотя дальнейшие дрейф или подбор могли протекать на севере, сам комплекс признаков вполне мог быть занесен туда с юга. Я склонен полагать, что в эволюции монголоидного типа могли сыграть свою роль оба этих признака. Признаки южной ветви монголоидов первоначально могли сформироваться в результате полового отбора и/или дрейфа генов. Затем, когда некоторые из носителей этих признаков переселились в Центральную Азию, у них началось «дальнейшее развитие «темы», обусловленное теми самыми эволюционными факторами, о которых так увлеченно писал Дэйл Гатри. В этом смысле могло существовать две основных «домашних» платформы для развития признаков монголоидной расы. Эта кабинетная теория не нашла широкой научной поддержки, поскольку генетика пока что находится на пороге определения того, какие именно гены контролируют рост и развитие телосложения, не касаясь вопроса о выявлении основных различий внутри нашего собственного вида. С другой стороны, генетический анализ, вероятно, смог бы оказать немалую помощь в определении направления миграции.
Четыре пути в Центральную Азию
Как и всегда случается в подобного рода походах и экспедициях, осуществляемых как современными людьми, так и их далекими предками, на решение о выборе дальнейшего пути вновь прибывшими переселенцами в Азии повлияли два фактора — географические особенности местоположения и климат. Правила выбора места всегда были достаточно просты: старайся селиться там, где есть вода и выпадает достаточно дождей, а во время миграции избегай пустынь и высоких горных хребтов и следуй вдоль берегов рек. Мы уже располагаем рядом свидетельств того, что маршрут собирателей на прибрежной полосе вдоль побережья Индийского океана на пути в Австралию действительно был наиболее легким и удобным. Чем же это объяснить? Ведь на самом деле маршрут был далеко не из простых; дело в том, что поначалу через каждые несколько сотен километров нашим первопроходцам-мигрантам приходилось переправляться вброд через устья больших рек. Но раз они совершали подобные переправы на пути в Австралию, им, естественно, приходилось проделывать то же самое и вдоль всего побережья Восточной Азии. И на берегу очередной великой реки у переселенцев была возможность повернуть влево и направиться в глубь материка, питаясь дарами речных вод и следуя вверх по ее течению. Как заметил Марко Поло, один из первых европейцев-путешественников, побывавший в Восточной Азии, пустыни и горы представляют собой труднопреодолимые преграды для всякого, кто стремится проникнуть в Центральную Азию. За исключением немногочисленных перевалов, путь туда возможен только вдоль долин, образуемых руслами больших рек. Как мы знаем, первый успешный исход из Африки, увлекший далеких предков всех неафриканских народов, имел место примерно 75 тысяч лет тому назад. Продвигаясь вдоль прибрежной полосы, они могли достаточно рано проникнуть в Китай и Японию. На этом пути они, по всей видимости, обошли стороной весь Центральноазиатский регион. Однако они в любой момент своего дальнего странствия могли свернуть влево и двинуться в глубь материка вверх по течению одной из великих рек. На севере Индии, где в небо вздымаются заснеженные пики Гималаев, дальнейший путь был сопряжен с серьезными трудностями. Гигантские складки горных хребтов, возникшие в результате грандиозной тектонической подвижки, которая произошла в Азии в глубокой древности, высятся по обе стороны высочайших вершин Гималаев, охватывая Непал и Тибет. Высокие горные хребты, практически всюду достигающие высоты 3000 м, наглухо закрывают доступ в Центральную Азию со стороны побережья Индийского океана на протяжении более 6500 км от Афганистана на западе до Ченьду, Китай, на востоке. Там, на востоке, эта гряда расстилается как скомканный ковер, образуя целый ряд хребтов, протянувшихся с севера на юг еще на 2500 км — от начала Шелкового пути в северном Китае до Таиланда на юге. Шелковый путь, получивший широкую известность на Западе благодаря тому же Марко Поло, представлял собой длинный торговый маршрут[283], протянувшийся к северу от Гималаев и проходящий параллельно им. Этот маршрут связывал Восток с Западом. Он проходил через всю Центральную Азию, практически совпадая с южной границей сердца Мамонтовой степи, о которой писал Гатри. Шелковый путь и в наши дни, как и в древности, остается одной из немногих артерий, связывающих Китай с Западом, если не считать протяженный путь, ведущий на юг, к Сингапуру. На восток — по Шелковому пути от западной оконечности Гималаев. В наши дни Шелковый путь проходит по южной и северной границам пустыни Такламакан в Синьцзяне. В эпоху палеолита на месте нынешней пустыни лежали травянистые степи, а еще дальше к северу находился целый ряд водных путей, в том числе — реки Тарим и Джунгария, служившие легким и удобным путем для охотников и собирателей из западных районов Центральной Азии, таких, как Таджикистан, Узбекистан, Киргизстан и Казахстан, в Синьцзян[284] и Монголию. Этими же водными артериями могли воспользоваться и древнейшие люди, стремившиеся проникнуть в Центральную Азию[285]. А теперь обратимся к реалиям Каменного века. Первые люди могли проникнуть в Центральную Азию с юга вверх по течению Инда, протяженность которого достигает 8000 км, и достичь западных районов Китая у западной оконечности Шелкового пути. Предположим, что речь идет о первых мигрантах, оказавшихся в Индии. Важнейшей задачей, стоявшей перед ними после подъема вверх по течению Инда, было преодоление горных хребтов на севере Индии и Пакистана. Хребты эти простирались далеко на запад, до Афганистана. Обойти эти горы с запада и двигаться дальше на север, через территорию современного Афганистана, было делом крайне сложным, если не сказать — невозможным, ибо, не говоря уже о невероятно длинном обходном пути на запад, мигрантам пришлось бы идти практически через пустыни. Марко Поло пересек эти пустыни. Он отправился в путь из Ормуза в устье Персидского залива и, пройдя через Афганистан, оказался в Кашмире, после чего, миновав горный перевал, попал в Китай и направился в Кашгар. Оттуда он, уже по Шелковому пути, двинулся в самое сердце бывшей Мамонтовой степи. Впрочем, Марко Поло мог попасть в Кашмир и гораздо более легким путем. Немного восточнее, неподалеку от побережья современного Пакистана, великий Инд описывает петлю и поворачивает на север, до того места, где начинается водный путь, по которому можно попасть в Кашмир. Другой, тоже относительно равнинный, путь в Центральную Азию через верховья Инда ведет через Хайберский перевал в Кабул, а оттуда — в Узбекистан и Казахстан и далее на восток, в Синьцзян. На запад — по Шелковому пути из Китая. На всем протяжении истории Шелковый путь служил единственным маршрутом, ведущим из Китая на запад — в Центральную Азию. Таким образом, альтернативный путь, ведущий в Мамонтовую степь, мог пролегать от тихоокеанского побережья Восточной Азии. Передовые отряды древнейших мигрантов, двигавшихся вдоль береговой линии, могли пройти по всему побережью Китая и, оказавшись в северном Китае, повернуть на запад и по Шелковому пути проникнуть в Монголию, Синьцзян и Южную Сибирь. На север — из Бирмы в Тибет. Этот, третий путь в Мамонтовую степь, не слишком часто используемый в наши дни в качестве торговой артерии, ведет к восточной оконечности Гималаев. Восточная окраина Гималаев состоит из множества отрогов и складок, образовавшихся в том месте, где Индийская материковая плита уперлась в Азиатскую, что вызвало тектонические подвижки. Как видно на карте (рис. 5.4), эти складки представляют собой как бы естественные каналы для большинства великих рек Южной и Юго-Восточной Азии. Реками этими, если двигаться с запада на восток, являются Брахмапутра, протекающая в Бангладеш, Сальвин, несущая свои воды по территории Бирмы, Меконг, текущий по землям Вьетнама, и Янцзы, которая течет в южном Китае. Эти четыре великие реки, вытекающие из северо-восточных отрогов Тибета, несут свои воды практически параллельно друг другу на протяжении свыше 150 км, а расстояние, разделяющее их на этом участке, составляет буквально считаные километры. Последняя из четырех великих рек, Янцзы, берет свое начало в горах северо-восточного Тибета у северной окраины плато, возле самой границы Мамонтовой степи. Я упомянул об этих руслах великих рек не потому, что многие торговцы и в наши дни активно пользуются этими водными артериями, а потому, что они позволяют попасть в Тибет[286], Монголию и Центральную Азию через одно из четырех устий рек, которые находятся на большом расстоянии друг от друга на побережье Юго-Восточной и Восточной Азии и впадают в Индийский и Тихий океаны. На восток — из России. Наконец, следует назвать еще один, северный маршрут миграции с запада на восток, в Восточную Азию: через земли азиатских районов России, так называемый русский Алтай. Наиболее удобный сухопутный путь из района Алтая в Центральную Азию во время самого мягкого в климатическом отношении периода Позднего Каменного века (30—50 тысяч лет назад) пролегал непосредственно через степи. Продвигаясь на восток через территорию Южной Сибири по многочисленным озерам и рекам, существовавшим в те времена, наши древние мигранты-первопроходцы могли достичь района озера Байкал по маршруту, пролегающему через Синьцзян и Монголию. В те времена степи на этих землях были покрыты густыми высокими травами. Тем не менее для того, чтобы проделать этот путь, мигрантам первым делом надо было попасть на Алтай. Как мы увидим, примерно 40 тысяч лет назад они действительно достигли Алтая и окрестностей озера Байкал в Южной Сибири. Итак, теперь мы знаем, что в древности существовало четыре потенциальных маршрута, ведущих в Центральную Азию: три из них вели от побережья Индийского и Тихого океанов (с запада, юга и востока), а один брал свое начало на территории современной России (с северо-запада).
 Рис. 5.4
В самой же Центральной Азии существовало три параллельных маршрута вдоль водных артерий, соединявших восток и запад, — маршрута, по которым могли пройти первопроходцы древности. Два из них — южные: через Синьцзян и Монголию, и один — северный через Южную Сибирь. Северный маршрут был доступен только в периоды потепления в эпоху палеолита, то есть от 30 до 50 тысяч лет назад, во время так называемых климатических пауз (см главу 3).
Рис. 5.4
В самой же Центральной Азии существовало три параллельных маршрута вдоль водных артерий, соединявших восток и запад, — маршрута, по которым могли пройти первопроходцы древности. Два из них — южные: через Синьцзян и Монголию, и один — северный через Южную Сибирь. Северный маршрут был доступен только в периоды потепления в эпоху палеолита, то есть от 30 до 50 тысяч лет назад, во время так называемых климатических пауз (см главу 3).
Когда люди современного типа впервые отправились в странствия по степям?
На сегодняшний день наиболее ясные и достоверные археологические свидетельства присутствия людей современного типа к северу от Гималаев более 40 тысяч лет тому назад нам удалось обнаружить именно вдоль четвертого и самого северного из этих коридоров, ведущих в степи. Южная Сибирь находится буквально у самого сердца Мамонтовой степи, которое, согласно трудам Гатри, находилось в Синьнцзяне и Монголии. Впечатляющие результаты последних археологических раскопок в пещерах Алтая — Усть-Каракол и Кара-Бом (см .рис 5.4) — со всей ясностью показывают, что новая технология обработки камня эпохи Верхнего палеолита появилась в этих местах около 43 тысяч лет назад. Эти датировки находок на Алтае весьма близки по возрасту к другим находкам, сделанным далее к востоку отсюда, в районе села Макарово на западном берегу озера Байкал, возраст которых составляет около 39 тысяч лет. В период между 30 и 35 тысячами лет тому назад существовал целый ряд подобных стоянок. Варварина Гора и Толбага на восточном берегу озера Байкал и Малая Сия, далее к северу, у берегов Енисея. Это означает, что переход к Верхнему палеолиту на всей территории Южной Сибири произошел как минимум не позже аналогичного перехода в Европе[287]. Не так давно бельгийский археолог Марсель Отте заявил, что центр зарождения новой технологии обработки камня эпохи Верхнего палеолита находился на севере нагорья Загрос (на территории нынешних Ирана и Ирака), откуда эта технология впоследствии распространилась как на северо-запад — в Левант и Европу, так и на северо-восток — в Центральную Азию и на Алтай (Россия), предположительно — по тому пути, который описывал еще Марко Поло, или через Кавказ[288].
Способны ли каменные орудия поведать о том, какие племена первыми появились в Центральной Азии?
Появление новой технологии обработки камня эпохи Верхнего палеолита принято считать решающим свидетельством проникновения человека современного типа в Европу (на Балканы и далее на запад). Она же считается своего рода индикатором перехода от неандертальца, для которого характерны орудия эпохи Среднего палеолита. Как известно, этот переход произошел несколько сотен тысяч лет назад (см. главу 3). Это можно считать свидетельством параллельного прибытия людей современного типа, обладателей новой технологии обработки камня эпохи Верхнего палеолита, с юга, с нагорья Загрос, на Алтай, что произошло около 43 тысяч лет назад[289]. Впрочем, если обладание этой новой технологией не являлось этаким sine qua nоn[290] для появления людей современного типа в Азии, то вполне вероятно, что первые люди могли проникнуть на Алтай и в более ранний период. Правда, надо признать, что к датировке времени прибытия первых людей в любые районы Азии необходимо подходить с особой осторожностью. Во-первых, хотя технология обработки камня, возникшая в эпоху Верхнего палеолита, действительно свидетельствует о присутствий человека современного типа в Европе, а также на территории Леванта и Юго-Восточной Азии, она была отнюдь не единственным техническим знанием, которым пользовались люди современного типа, так что свидетельства о ее применении в тех или иных районах нельзя использовать в качестве опоры для датировки появления первых людей. Дело в том, что эпоха Верхнего палеолита явилась, образно говоря, собранием целого ряда новшеств, первое из которых, согласно новейшим археологическим данным, появилось около 45—50 тысяч лет тому назад, и его принято ассоциировать с Северной и Западной Евразией. Первые люди современного типа использовали целый ряд разнообразных «старых» технологий и навыков, включая традиционные способы изготовления орудий, унаследованные от Среднего палеолита и характерные для неандертальцев. Находки в пещерах Алтая (Россия), в частности — в пещере Кара-Бом, свидетельствуют о наличии традиционных техник Среднего палеолита в слоях, залегающих ниже типичных ауриньякских слоев Верхнего палеолита и датируемых 62—72 тысячами лет назад. Эти нижние слои трудно отнести к деятельности людей современного или архаического типа (в частности, неандертальцев), поскольку они с равным основанием могли быть оставлены и теми и другими, и к тому же в них не сохранилось ископаемых останков и артефактов, позволяющих уточнить датировку. Ученые никогда не находили останки неандертальцев так далеко на севере — на Алтае, хотя на юго-западе, например в Тешикташе (Узбекистан, Центральная Азия), подобные останки — не редкость. Ключевой вопрос здесь состоит в том, отражают ли более ранние слои Среднего палеолита постоянное накапливание технических навыков, позволившее совершить прорыв в технологии изготовления орудий, или культура Верхнего палеолита была занесена откуда-то извне. Последнее утверждение представляется более реальным, по крайней мере, в том, что касается ауриньякской технологии. Таким образом, сохраняется реальная вероятность того, что слои эпохи Среднего палеолита, возраст которых составляет 60—70 тысяч лет, найденные в пещере Кара-Бом, представляют собой свидетельства присутствия человека современного типа на Алтае в еще более раннюю эпоху[291]. Вторая причина, побуждающая проявлять осторожность в датировке, заключается в том, что вплоть до недавнего времени хронология палеолита в Синьцзяне, Тибете и южной Монголии была практически не изучена. Дело здесь не в отсутствии останков и артефактов эпохи палеолита, а в том, что многие из них не поддаются датировке. Один из недавних отчетов о раскопках в Монголии утверждает, что возраст традиционных технических навыков Среднего палеолита, по-видимому, составляет 60 тысяч лет. В другом отчете говорится, что на Тибете существовали две раздельные микролитические традиции эпохи Верхнего палеолита: северная и южная, связанные с жителями и навыками обработки каменных орудий соответственно Северного и Южного Китая. Датировка была не слишком достоверной, поскольку многие находки были обнаружены на поверхности (находки с поверхности, естественно, вырваны из исторического контекста и реального соположения слоев, необходимых для археологической датировки)[292]. Таким образом, мы располагаем весьма убедительными археологическим свидетельствами того, что технология эпохи Позднего палеолита распространялась и северным путем, через всю Южную Сибирь, и начало этому процессу было положено на Алтае (Россия) примерно 43 тысячи лет тому назад. С другой стороны, более чем вероятно, что в Центральной Азии и на юге, в сердце Мамонтовой степи, уже в древности активно селились люди современного типа, однако реальных ископаемых останков, подтверждающих это, пока не обнаружено. Как мы вскоре увидим, данные генетических исследований, полученные в Северной Азии, служат весомым аргументом в пользу того, что люди современного типа появились в Северо-Западной Азии около 40 тысяч лет назад. Люди эпохи Верхнего палеолита, жившие в Северной Азии, во многом близки к представителям богатой граветтийской культуры, возникшей в Мамонтовой степи далеко на западе — в Центральной и Восточной Европе, о чем свидетельствуют находки в урочище Мальта неподалеку от озера Байкал, возраст которых — 23 тысячи лет (см. табл. 19)[293].
Генетические данные
Я проанализировал многие археологические материалы, свидетельствующие о присутствии первых людей современного типа в Центральной Азии, а также на побережье двух океанов — у «входов» в единственные коридоры, ведущие в Центральную Азию, к Шелковому пути и в самое сердце Мамонтовой степи. Эти трое «ворот» находились на северо-западе Азии, в Индокитае и в Северо-Восточном Китае. Что же касается генетических данных, то их я упоминал очень скупо, и на то были свои причины. Структура ветвей митохондриевой ДНК и Y-маркеров является куда лучшими индикаторами древнейших миграций молекул, чем средством датировки или идентификации так называемых рас или этнических групп. Таким образом, куда надежнее — разумеется, там, где это возможно, — прослеживать маршруты предполагаемых миграций с учетом географических, археологических и климатических данных, прежде чем выяснять, в какой мере эти результаты подтверждаются генетическими данными.
Деление на север и юг в Восточной Азии
Я уже упоминал в главе 2 о том, что после первичного рассеяния народов — выходцев из Африки были постепенно заселены все регионы и земли Старого Света и юго-западная часть Тихого океана и что вплоть до наступления последнего великого Ледникового периода (около 20 тысяч лет тому назад) межрегиональная миграция генов была крайне незначительной или отсутствовала совсем. И Азия, несмотря на всю свою богатую историю переселения народов, отнюдь не явилась исключением из этого правила. Некоторые крупные генетические ветви на азиатском континенте стали известны генетикам задолго до появления понятия о митохондриевой ДНК и Y-маркерах. Как и следовало ожидать, регионы распространения кавказоидного типа в Западной Евразии располагались поблизости друг от друга, а некоторые из них «накладывались» на Индийский субконтинент. При этом имело место выраженное отличие от Восточной Евразии, население которой, в свою очередь, подразделялось на северян и южан, что соответствовало северной и южной ветвям монголоидов. Если говорить о происхождении будущих народов, мы должны провести четкую границу между Юго-Восточной Азией (включая южное побережье Китая: провинции Гуандун, Гуаньси и Фудзян) и Северо-Восточной Азией (включая Северный Китай, восточное побережье Китая, Японию, Корею, Монголию, Тибет и некоторые районы Сибири), причем одна из групп населения, живущая на крайнем северовостоке Азии, в этническом отношении занимает промежуточное положение между кавказоидами и северными монголоидами[294]. Подобные исследования мало что дают для выявления локусов формирования народов, но, несмотря на это, показывают, что даже в крупных этносах, таких, как так называемый этнос хань в Китае, прослеживаются выраженные региональные варианты, тяготеющие к дифференциации по принципу север — юг. Эти исследования помогли также развеять целый ряд мифов о генетических истоках происхождения китайцев. Так, например, широко бытовало мнение, что за последние несколько тысяч лет существования императорской власти в Китае[295] имела место массовая экспансия ханьцев с севера на юг, следствием которой якобы были вытеснение, замещение и ассимиляция коренного населения этих регионов. Однако современные исторические исследования показывают, что народности Южного Китая, исходя в первую очередь из прагматических политических соображений, стали называть себя ханьцами совсем недавно — всего каких-нибудь несколько веков назад[296]. Это свидетельствует о том, что большинство миграций и переселений народных масс в Китае носило скорее легендарный, нежели реальный характер, и поддерживает точку зрения о том, что состав региональных этнических групп населения в Китае был куда более устойчивым, чем это считалось прежде. Другой подход к решению вопроса о том, сформировались ли монголоиды как раса на севере или на юге, — это анализ вариативности. Исходя из основных принципов учения Дарвина, несколько видов (или в данном случае новая этническая группа, развившаяся на основе небольшой группы-основателя) должны были иметь общий центр происхождения, сохраняя при этом широкий спектр вариантов (в том числе и закрепленных генетическими изменениями) в пределах такой группы. Сегодня, когда доступен генетический анализ, у южной ветви монголоидов неизменно наблюдается гораздо более широкий спектр вариантов, чем у северной. Изучение изменений Y-хромосомы также позволяет считать родиной монголоидов именно южные районы[297]. Несмотря на всю свою пользу, применение подобного упрощенного подхода к вариативности имеет два потенциальных недостатка. Во-первых, ни один из генных путей или маркеров, использованных в исследованиях, нельзя идентифицировать с признаками «монголоидности»: останки людей отождествляют с монголоидами просто по месту находки, и поэтому подобный аргумент, если применять его с достаточной последовательностью, рискует оказаться в замкнутом круге одних и тех же посылок и выводов. Во-вторых, Северная Азия, в отличие от тропической Юго-Восточной Азии, во время последнего Ледникового периода сильно пострадала от депопуляции (сокращения численности населения), массового вымирания людей и генного дрейфа, так что низкая вариативность, наблюдаемая сегодня на севере, отнюдь не обязательно отражает первоначальную этническую картину. Но, пожалуй, наиболее радикальная гипотеза, описывающая влияние последнего Ледникового периода на население Центральной и Северной Азии, сводится к тому, что в древности весь обширный азиатский регион, простирающийся к северу от Гималаев, был совершенно безлюдным, так что нынешние жители этих мест являются потомками выходцев из Южного Китая и Юго-Восточной Азии, а также Европы. Это, разумеется, означает, что мы так никогда и не узнаем, каким был первоначальный генетический комплекс древнейших жителей Северной Азии. Эта точка зрения представляется не слишком убедительной. Во-первых, детальные палеоклиматологические[298] исследования показывают, что во время Ледникового периода в Центральной Азии существовали обширные пространства степной тундры, способные обеспечить всем необходимым охотников. Это подтверждается археологическими находками, отражающими основные виды занятий человека во время максимальной фазы последнего Ледникового периода (см. главу 6). Дэйл Гатри решительно выступает против гипотезы о полном отсутствии населения на юге, в сердце Мамонтовой степи. Во-вторых, как мы увидим ниже, подобная картина не позволяет объяснить наличие генетического многообразия и поливариантности на юго-западе Центральной Азии (в Узбекистане и Киргизстане) или резко выраженных географических различий между представителями разных ветвей генетического древа в Северной и Южной Азии. В частности, существует целая группа генетических линий, единых для Северной Европы, Северной Азии и Северной Америки, однако не имеющих ничего общего с Южным Китаем или Юго-Восточной Азией[299].
Схема развития генетического древа
Как и в случае с историей исхода из Африки, ответ на интересующий нас вопрос станет более ясным, если мы посмотрим на маркеры Адама и Евы. Большим достоинством построения древа митохондриевой ДНК и Y-хромосомы является то, что они позволяют проследить ветви и даже побеги, соответствующие отдельным молекулам, применительно к каждому конкретному району. Эту увлекательную научную «игру» в гены и районы их распространения можно назвать филогеографией. В принципе она заключается в соединении разрозненных региональных побегов в крупные ветви и размещении последних на генетическом древе. Хотя правила предельно просты, их применение может оказаться достаточно сложным. Чтобы проследить векторы миграции из одного региона в другой, нам необходимо найти ветви-источники в обоих регионах и новый «побег» в искомом районе, который отсутствует на ветви прародины. Для прародины с многообразными векторами миграции в самых разных направлениях нам необходимо выявить общие ветви, имеющие разные «побеги» в других районах. Пожалуй, один из наиболее удачных примеров подобной методики был описан в работе видного итальянского генетика Антонио Торрони и его коллег, опубликованной в 1993 г.
 Правая часть
Правая часть
 Левая часть
Рис 5.5
Эти исследователи выявили четыре основные линии митохондриевой ДНК этнической группы-родоначальницы коренного населения Америки, пометив их буквами А, В, С и D. Истоки каждой из этих буйно разросшихся ветвей, давших обильные «побеги» в Америке, можно возвести к аналогичным ветвям-прародителям А, В, С и D в Азии (см. главы 6 и 7).
Мы можем применить этот же подход и к гораздо более сложной проблеме — идентификации прародины монголоидов в Азии. Взяв за начало митохондриевую ДНК Восточной Азии, мы можем заметить, что на сегодня там присутствуют девять четко сформировавшихся ветвей (гаплогрупп; см. рис. 5.5), которые можно обозначить буквами A, D, С, D, E, F, G, X и Z. За исключением сестринских групп С и Z, ни одна из этих групп не может быть выведена на основе другой. Каждая из них более или менее уверенно выстраивается на основе двух евразийских линий-родоначальниц — Манью или Насрин. Недавно были описаны еще несколько второстепенных групп (М7/М10 и N9), однако их существование никак не сказывается на значимости аргументации. (Более подробно о филогенезе и источниках по распределению гаплогрупп митохондриевой ДНК в Восточной Азии см. примечание[300].)
Из девяти наиболее хорошо изученных восточно-азиатских ветвей митохондриевой ДНК три (С, X и Z, см. рис. 5.9) прослеживаются только к северу от условной линии «запад — восток», проведенной ниже Гималаев, тогда как остальные шесть (А, В, D, E, F и G) были обнаружены у обеих — как северной, так и южной — ветвей монголоидов. На первый взгляд может показаться, что эти уникальные северные ветви — весомые аргументы в пользу гипотезы о том, что прародина монголоидов находилась к северу от Гималаев, но на самом деле у этого феномена есть более простое объяснение. Дело в том, что линия С и близкородственная ей линия Z, а также загадочная группа X не являются коренными фигурантами древнейшей истории Монголии. Они появились среди северной ветви монголоидов откуда-то извне, вероятнее всего — из Сибири или Центральной Азии. Как мы скоро увидим, в поддержку этой версии существует целый ряд аргументов. Распределение генов и структура шести остальных ветвей генетического древа свидетельствуют о том, что генетическая прародина монголоидов находилась к югу от Гималаев. С особенной ясностью на это указывают древнейшие генетические ветви В и F. Таким образом, мы, по всей видимости, имеем дело сразу с несколькими источниками генов у жителей Юго-Восточной Азии, которые впоследствии распространились на север. Наоборот, среди обитателей Северной Азии мы имеем несколько ветвей, никак не связанных с коренным населением Юго-Восточной Азии. Возвращаясь к той же версии о развитии ветвей, можно отметить, что идея о южной прародине и последующей миграции монголоидов на север подтверждается данными исследования материнских генов и вполне соответствует гипотезе об эволюции сундадонтов в синодонтов (см. главу 5).
Чтобы сосредоточить внимание на поисках возможного вторичного северного региона «дальнейшего развития той же темы», нам придется обратиться к обзору географических зон, где присутствуют все шесть ветвей митохондриевой ДНК — А, В, D, E, F и G, — являющихся общими для северной и южной ветвей монголоидов. Таким образом, продвигаясь с запада на восток, мы обнаружим, что в Центральной Азии присутствуют пять основных линий (за исключением G), в Тибете — все шесть, в Монголии — Пять (за исключением Е), в Китае — четыре (за исключением Е и G) и, наконец, в Корее — пять (за исключением[301]). Пожалуй, вряд ли можно считать простым совпадением, что страны с большим числом генетических линий, обычно ассоциируемых с монголоидной расой — Центральная Азия, Тибет, Монголия и Корея, — расположены вдоль 40-й параллели, которая, согласно Гатри, служила своего рода осью Мамонтовой степи (см. рис. 5.6).
Основываясь на этих факторах, мы можем задать вопрос о том, каков примерный возраст этих ветвей, расположенных вдоль 40-й параллели. Тоомас Кивисилд установил, что местный возраст четырех из этих ветвей в Монголии весьма близок: ветвь А насчитывает 35 500 лет, В — 40 500 лет или 33 500 лет (см. ниже), D — 44 500 лет и, наконец, F — 42 500 лет. Эти даты значительно «моложе», чем средний возраст тех же линий на юге, в Южном Китае. Но самое важное заключается в том, что они вполне сопоставимы с датами начала активной колонизации Центральной Азии и относятся к эпохе после улучшения климата в Азии, которое произошло примерно 52 тысячи лет назад (см. главу 3) и благодаря которому первые обитатели Азии смогли проникнуть на просторы Мамонтовой степи[302].
И все же: по каким путям и маршрутам люди современного типа проникли в этот обширный регион? Обратившись к анализу двух древнейших линий Юго-Восточной Азии — В и F (идущих от потомков Насрин), которые тянутся из Юго-Восточной Азии в Тибет, Монголию и далее на северо-восток, вдоль побережья Тихого океана, мы видим, что они проникают в глубь материка в Сибири (Северная Азия), а затем останавливаются[303]. В этом отношении распределение этнических групп В и F отражает древнейшее распространение признаков сундадонтности. Как В, так и F достигают наиболее выраженной поливариантности среди туземных этнических групп Таиланда, Вьетнама и Камбоджи, что свидетельствует о том, что их прародиной является Индокитай.
Весомым аргументом в поддержку версии о том, что группа F возникла и сформировалась на юге, является вновь идентифицированная гаплогруппа «npe-F», которая распространена среди малайцев — коренных жителей Малайского полуострова (выводы сделаны на основе распространения признаков сундадонтности; см. рис. 5.6).
Левая часть
Рис 5.5
Эти исследователи выявили четыре основные линии митохондриевой ДНК этнической группы-родоначальницы коренного населения Америки, пометив их буквами А, В, С и D. Истоки каждой из этих буйно разросшихся ветвей, давших обильные «побеги» в Америке, можно возвести к аналогичным ветвям-прародителям А, В, С и D в Азии (см. главы 6 и 7).
Мы можем применить этот же подход и к гораздо более сложной проблеме — идентификации прародины монголоидов в Азии. Взяв за начало митохондриевую ДНК Восточной Азии, мы можем заметить, что на сегодня там присутствуют девять четко сформировавшихся ветвей (гаплогрупп; см. рис. 5.5), которые можно обозначить буквами A, D, С, D, E, F, G, X и Z. За исключением сестринских групп С и Z, ни одна из этих групп не может быть выведена на основе другой. Каждая из них более или менее уверенно выстраивается на основе двух евразийских линий-родоначальниц — Манью или Насрин. Недавно были описаны еще несколько второстепенных групп (М7/М10 и N9), однако их существование никак не сказывается на значимости аргументации. (Более подробно о филогенезе и источниках по распределению гаплогрупп митохондриевой ДНК в Восточной Азии см. примечание[300].)
Из девяти наиболее хорошо изученных восточно-азиатских ветвей митохондриевой ДНК три (С, X и Z, см. рис. 5.9) прослеживаются только к северу от условной линии «запад — восток», проведенной ниже Гималаев, тогда как остальные шесть (А, В, D, E, F и G) были обнаружены у обеих — как северной, так и южной — ветвей монголоидов. На первый взгляд может показаться, что эти уникальные северные ветви — весомые аргументы в пользу гипотезы о том, что прародина монголоидов находилась к северу от Гималаев, но на самом деле у этого феномена есть более простое объяснение. Дело в том, что линия С и близкородственная ей линия Z, а также загадочная группа X не являются коренными фигурантами древнейшей истории Монголии. Они появились среди северной ветви монголоидов откуда-то извне, вероятнее всего — из Сибири или Центральной Азии. Как мы скоро увидим, в поддержку этой версии существует целый ряд аргументов. Распределение генов и структура шести остальных ветвей генетического древа свидетельствуют о том, что генетическая прародина монголоидов находилась к югу от Гималаев. С особенной ясностью на это указывают древнейшие генетические ветви В и F. Таким образом, мы, по всей видимости, имеем дело сразу с несколькими источниками генов у жителей Юго-Восточной Азии, которые впоследствии распространились на север. Наоборот, среди обитателей Северной Азии мы имеем несколько ветвей, никак не связанных с коренным населением Юго-Восточной Азии. Возвращаясь к той же версии о развитии ветвей, можно отметить, что идея о южной прародине и последующей миграции монголоидов на север подтверждается данными исследования материнских генов и вполне соответствует гипотезе об эволюции сундадонтов в синодонтов (см. главу 5).
Чтобы сосредоточить внимание на поисках возможного вторичного северного региона «дальнейшего развития той же темы», нам придется обратиться к обзору географических зон, где присутствуют все шесть ветвей митохондриевой ДНК — А, В, D, E, F и G, — являющихся общими для северной и южной ветвей монголоидов. Таким образом, продвигаясь с запада на восток, мы обнаружим, что в Центральной Азии присутствуют пять основных линий (за исключением G), в Тибете — все шесть, в Монголии — Пять (за исключением Е), в Китае — четыре (за исключением Е и G) и, наконец, в Корее — пять (за исключением[301]). Пожалуй, вряд ли можно считать простым совпадением, что страны с большим числом генетических линий, обычно ассоциируемых с монголоидной расой — Центральная Азия, Тибет, Монголия и Корея, — расположены вдоль 40-й параллели, которая, согласно Гатри, служила своего рода осью Мамонтовой степи (см. рис. 5.6).
Основываясь на этих факторах, мы можем задать вопрос о том, каков примерный возраст этих ветвей, расположенных вдоль 40-й параллели. Тоомас Кивисилд установил, что местный возраст четырех из этих ветвей в Монголии весьма близок: ветвь А насчитывает 35 500 лет, В — 40 500 лет или 33 500 лет (см. ниже), D — 44 500 лет и, наконец, F — 42 500 лет. Эти даты значительно «моложе», чем средний возраст тех же линий на юге, в Южном Китае. Но самое важное заключается в том, что они вполне сопоставимы с датами начала активной колонизации Центральной Азии и относятся к эпохе после улучшения климата в Азии, которое произошло примерно 52 тысячи лет назад (см. главу 3) и благодаря которому первые обитатели Азии смогли проникнуть на просторы Мамонтовой степи[302].
И все же: по каким путям и маршрутам люди современного типа проникли в этот обширный регион? Обратившись к анализу двух древнейших линий Юго-Восточной Азии — В и F (идущих от потомков Насрин), которые тянутся из Юго-Восточной Азии в Тибет, Монголию и далее на северо-восток, вдоль побережья Тихого океана, мы видим, что они проникают в глубь материка в Сибири (Северная Азия), а затем останавливаются[303]. В этом отношении распределение этнических групп В и F отражает древнейшее распространение признаков сундадонтности. Как В, так и F достигают наиболее выраженной поливариантности среди туземных этнических групп Таиланда, Вьетнама и Камбоджи, что свидетельствует о том, что их прародиной является Индокитай.
Весомым аргументом в поддержку версии о том, что группа F возникла и сформировалась на юге, является вновь идентифицированная гаплогруппа «npe-F», которая распространена среди малайцев — коренных жителей Малайского полуострова (выводы сделаны на основе распространения признаков сундадонтности; см. рис. 5.6).
 Рис. 5.6
Представители этой группы «пре-F» встречаются в Южном Китае (провинции Юннань и Гуаньси), Индокитае (Таиланд и Вьетнам), на Суматре, а также Андаманских и Никобарских островах, что, по всей вероятности, свидетельствует о древнейших этнических связях с обитателями прибрежной полосы в Юго-Восточной Азии[304].
Располагая сведениями о том, распространились ли представители группы В на запад, проникнув из Китая в Монголию вдоль Шелкового пути, или же двинулись на север из Бирмы через Тибет, мы сможем узнать направление маршрута первой волны колонизации, которая, по всей видимости, имела место около 50 тысяч лет тому назад. Это — немаловажные ключи. Тот же Тоомас Кивисилд установил, что возраст двух основных подгрупп В, проникших в Монголию, составляет 40 500 лет (В1) и 33 500 лет (В2) Это говорит о том, что, хотя в Монголии и части западных районов Центральной Азии оба подтипа В могли появиться еще в эпоху палеолита, более древняя из этих ветвей могла пройти через Тибет, тогда как более поздняя, не исключено, проникла на запад, в Центральную Азию, с побережья Тихого океана, где ее представители сегодня занимают доминирующее положение[305].
Надо упомянуть и о трех линиях — D, Е и G, — частично связанных с Манью, линиях, распространение представителей которых в Восточной Азии увеличивается при продвижении с севера на юг, отражая аналогичное распространение признаков синодонтности. Однако остается нерешенным вопрос о том, выходцами откуда могли быть другие основатели «этнической колонии монголоидов» в Мамонтовой степи (С, X и Z) и каким именно из трех возможных коридоров, ведущих в Мамонтовую степь, они воспользовались. К югу от Гималаев представители этих трех линий Северной Евразии встречаются крайне редко. Ветви-сестры С и Z наиболее распространены в Сибири и Северо-Восточной Азии, тогда как в более южных районах (за исключением Монголии, Тибета и Центральной Азии) их представители почти не встречаются. Линии эти тянутся вдоль всего континуума «восток — запад» через азиатские великие степи, и численность их представителей в Центральной Азии вплоть до Турции весьма значительна. Что касается ветви С, то она простирается даже в Америку через Аляску, однако преимущественное ее распространение на севере Восточной Азии свидетельствует о том, что эти люди не могли прийти в Центральную Азию через Китай. Наоборот, куда более вероятно, что они проникли в Азию откуда-то с запада, причем представители восточной ее ветви были обладателями новой технологии создания орудий, характерной для Позднего палеолита, артефакты которой были обнаружены в пещере Кара-Бом на Алтае (Россия). Возраст их — около 43 тысяч лет. Люди, создавшие их, принадлежали к группе Манью, и их древнейшие предки пришли сюда из Индии и Пакистана, проникнув в долину Инда через Кашмир или Афганистан и обогнув по пути западную оконечность Гималаев (см. рис 5.4) Все эти факты свидетельствуют о том, что мигранты проникли в азиатскую степь где-то 40—50 тысяч лет тому назад из западных районов Центральной Азии (см рис 5.6) Подсчеты Тоомаса Кивисилда, находящиеся в полном соответствии с версией о том, что линии С и 2 распространялись по степи и их носителями были обладатели новой технологии эпохи Верхнего палеолита, показывают, что возраст группы С в Монголии составляет 42 тысячи лет[306].
Последняя из северных линий, линия X, обнаружена только у европейцев и туземных жителей Америки (правда, есть сообщение о находке следов линии X в Южной Сибири), однако связь между Старым и Новым Светом существовала еще как минимум 30 тысяч лет назад. Это свидетельствует о том, что группа X распространялась по необъятным пространствам Центральной и Северо-Восточной Азии вместе с представителями культуры охотников на мамонтов, однако полностью исчезла в Азии во время Последнего Ледникового периода, сохранившись лишь на отдаленных окраинах своего ареала в Америке и Европе (см. главу 7)[307].
Столь сложная картина линий распространения митохондриевой ДНК свидетельствует о том, что монголоиды были выходцами с юга, тогда как жители Центральной Азии происходят от некоей западно-азиатской этнической группы, однако в формирование их родовых признаков внесли свой вклад уроженцы Восточной и Юго-Восточной Азии, мигрировавшие в Центральную и Северо-Восточную Азию. Эти генетические данные являются аргументами в поддержку теории о трех этапах колонизации Центральной Азии с «баз» на побережье Индийского и Тихого океанов 40—50 тысяч лет тому назад. Разграничение и выделение генетических составляющих в этносах и этнических группах Северо-Восточной Азии проясняет вопрос о происхождении ее населения и вполне отвечает концепции о том, что центром происхождения монголоидной расы была Юго-Восточная Азия.
Рис. 5.6
Представители этой группы «пре-F» встречаются в Южном Китае (провинции Юннань и Гуаньси), Индокитае (Таиланд и Вьетнам), на Суматре, а также Андаманских и Никобарских островах, что, по всей вероятности, свидетельствует о древнейших этнических связях с обитателями прибрежной полосы в Юго-Восточной Азии[304].
Располагая сведениями о том, распространились ли представители группы В на запад, проникнув из Китая в Монголию вдоль Шелкового пути, или же двинулись на север из Бирмы через Тибет, мы сможем узнать направление маршрута первой волны колонизации, которая, по всей видимости, имела место около 50 тысяч лет тому назад. Это — немаловажные ключи. Тот же Тоомас Кивисилд установил, что возраст двух основных подгрупп В, проникших в Монголию, составляет 40 500 лет (В1) и 33 500 лет (В2) Это говорит о том, что, хотя в Монголии и части западных районов Центральной Азии оба подтипа В могли появиться еще в эпоху палеолита, более древняя из этих ветвей могла пройти через Тибет, тогда как более поздняя, не исключено, проникла на запад, в Центральную Азию, с побережья Тихого океана, где ее представители сегодня занимают доминирующее положение[305].
Надо упомянуть и о трех линиях — D, Е и G, — частично связанных с Манью, линиях, распространение представителей которых в Восточной Азии увеличивается при продвижении с севера на юг, отражая аналогичное распространение признаков синодонтности. Однако остается нерешенным вопрос о том, выходцами откуда могли быть другие основатели «этнической колонии монголоидов» в Мамонтовой степи (С, X и Z) и каким именно из трех возможных коридоров, ведущих в Мамонтовую степь, они воспользовались. К югу от Гималаев представители этих трех линий Северной Евразии встречаются крайне редко. Ветви-сестры С и Z наиболее распространены в Сибири и Северо-Восточной Азии, тогда как в более южных районах (за исключением Монголии, Тибета и Центральной Азии) их представители почти не встречаются. Линии эти тянутся вдоль всего континуума «восток — запад» через азиатские великие степи, и численность их представителей в Центральной Азии вплоть до Турции весьма значительна. Что касается ветви С, то она простирается даже в Америку через Аляску, однако преимущественное ее распространение на севере Восточной Азии свидетельствует о том, что эти люди не могли прийти в Центральную Азию через Китай. Наоборот, куда более вероятно, что они проникли в Азию откуда-то с запада, причем представители восточной ее ветви были обладателями новой технологии создания орудий, характерной для Позднего палеолита, артефакты которой были обнаружены в пещере Кара-Бом на Алтае (Россия). Возраст их — около 43 тысяч лет. Люди, создавшие их, принадлежали к группе Манью, и их древнейшие предки пришли сюда из Индии и Пакистана, проникнув в долину Инда через Кашмир или Афганистан и обогнув по пути западную оконечность Гималаев (см. рис 5.4) Все эти факты свидетельствуют о том, что мигранты проникли в азиатскую степь где-то 40—50 тысяч лет тому назад из западных районов Центральной Азии (см рис 5.6) Подсчеты Тоомаса Кивисилда, находящиеся в полном соответствии с версией о том, что линии С и 2 распространялись по степи и их носителями были обладатели новой технологии эпохи Верхнего палеолита, показывают, что возраст группы С в Монголии составляет 42 тысячи лет[306].
Последняя из северных линий, линия X, обнаружена только у европейцев и туземных жителей Америки (правда, есть сообщение о находке следов линии X в Южной Сибири), однако связь между Старым и Новым Светом существовала еще как минимум 30 тысяч лет назад. Это свидетельствует о том, что группа X распространялась по необъятным пространствам Центральной и Северо-Восточной Азии вместе с представителями культуры охотников на мамонтов, однако полностью исчезла в Азии во время Последнего Ледникового периода, сохранившись лишь на отдаленных окраинах своего ареала в Америке и Европе (см. главу 7)[307].
Столь сложная картина линий распространения митохондриевой ДНК свидетельствует о том, что монголоиды были выходцами с юга, тогда как жители Центральной Азии происходят от некоей западно-азиатской этнической группы, однако в формирование их родовых признаков внесли свой вклад уроженцы Восточной и Юго-Восточной Азии, мигрировавшие в Центральную и Северо-Восточную Азию. Эти генетические данные являются аргументами в поддержку теории о трех этапах колонизации Центральной Азии с «баз» на побережье Индийского и Тихого океанов 40—50 тысяч лет тому назад. Разграничение и выделение генетических составляющих в этносах и этнических группах Северо-Восточной Азии проясняет вопрос о происхождении ее населения и вполне отвечает концепции о том, что центром происхождения монголоидной расы была Юго-Восточная Азия.
История Адама
Итак, после того как мы ознакомились с данными генетической структуры ветвей митохондриевой ДНК, изучение истории заселения Азии на общем древе Y-хромосомы и присутствия ряда миграций в Центральную Азию приобретает более достоверный характер. Древо Адама — это во многих отношениях новое понятие в науке. В этом качестве на нем недостает многих и многих «побегов», пышно зеленеющих на Евином древе жизни, а данные для ветвей развития Y-хромосомы весьма и весьма ненадежны. Так ради чего же мне возвращаться к Адаму и лишний раз выслушивать ту же историю, которую куда лучше и подробнее поведала нам Ева? Все дело в том, что Y-хромосома сама по себе является очень четким и достоверным региональным маркером. Выразительный пример этого — история заселения Европы (см. главу 3), где различные патернальные (отцовские) кланы отличались куда более высокой степенью этнической чистоты в рамках Европы, чем материнские кланы Евы. Распространение последних, несмотря на всю их региональную структурированность, остается куда более туманным. То же самое можно сказать и о заселении Восточной Азии. Генеалогическое древо Адама и Евы свидетельствует о том, что и жители Центральной Азии, и аборигены американского континента имеют давние корни, ведущие в Северо-Восточную, Восточную и Юго-Восточную Азию, однако разграничение этнических групп-родоначальниц обитателей Мамонтовой степи гораздо нагляднее прослеживается на Адамовом древе. Чтобы лучше понять этот феномен, давайте вспомним три основные внеафриканские ветви потомков Адама: Авель (YAP или М145), Каин (или RSP4Y) и Сиф (или М89)[308]. (См. рис 5.5.) Линия Каина, или каинитов, — это линия, быстро распространявшаяся на юго-восток вдоль побережья Индийского океана. Каиниты стали мужчинами-родоначальниками этнических групп в Восточной Индонезии, Австралии и Новой Гвинее. Собиратели на прибрежной полосе не ограничивались контактами с населением стран противоположного полушария и продвигались вдоль побережья Индийского и Тихого океанов вплоть до берегов Японии и Кореи, где они создали наиболее ранние свои поселения. Где-то на пути вдоль береговой линии примерно в эпоху Верхнего палеолита, по всей вероятности — ближе к концу своего долгого пути в Японию, у Каина родился сын-азиат, ставший родоначальником многих народов, и его потомки рассеялись по всей Северо-Восточной Азии, а затем по Шелковому пути проникли в Монголию и Центральную Азию. Часть каинитов двинулась дальше на северо-восток и, переправившись через будущий Берингов пролив, оказалась в Америке[309]. В главе 4 мы уже говорили о том, что западные сыновья Авеля приняли активное участие в заселении Европы, Среднего Востока и Африки. Однако существовала и другая, восточно-азиатская ветвь потомков Авеля. Как и каиниты, азиатские потомки Авеля быстро продвигались далеко на восток вдоль побережья двух океанов и достигли берегов Индонезии, а затем двинулись на северо-запад — к острову Тайвань и Японии. Крайней границей их продвижения на север стала Корея. Между тем сегодня в Индии не встретишь потомков азиатской ветви сынов Авеля. Это можно объяснить тем, что они продвигались очень быстро, а их единичные представители в Индии были стерты с лица Земли облаком вулканического пепла, возникшим в результате катастрофического извержения вулкана Тоба, которое произошло примерно 74 тысячи лет тому назад (см. главу 4). Зато в Тибете число азиатской ветви потомков Авеля достаточно велико, что свидетельствует о том, что некоторые из сынов Авеля сумели пережить извержение вулкана Тоба и направились из Восточной Индии вверх по течению великих рек и попали в Юго-Восточной Азии на обширные горные плато, где и обосновались в степях Монголии и на Алтае (Россия)[310]. Таким образом, получается, что оба клана мужчин-мигрантов прибрежной полосы воспользовались двумя восточными коридорами, чтобы проникнуть в степные просторы Центральной Азии (рис. 5.7). Их вклад в колонизацию Старого Света в количественном отношении был ничтожно мал по сравнению с вкладом ветви третьего потомка Адама — Сифа, семя которого было разнесено буквально по всем уголкам неафриканского мира. Его потомки воспользовались всеми тремя путями, чтобы проникнуть в глубинные районы Центральной Азии. По даннымпоследнего исследования, на долю линии потомков Сифа приходится 93% из общего числа 12 127 обследованных жителей островов Азии и Океании, а остальное составляют потомки линий Каина и Авеля. В этой связи важно подчеркнуть, что, подобно дочерям неафриканской Евы Насрин и Манью, сифиты продвигались вдоль береговой линии двух океанов в компании своих собратьев, сынов Каина и Авеля, и, что самое главное, — одновременно с ними. Представители линии Сифа прошли здесь не в более поздние времена и не другим путем, как утверждают некоторые генетики. Доказательством этого служат следы одновременного присутствия потомков всех трех братьев, в том числе — характерно сифские черты внешнего облика у представителей древнейших этнических групп собирателей на прибрежной полосе Индийского океана, в том числе и у жителей Юго-Восточной Азии[311].
 Рис. 5.7
Именно маршрут потомков Сифа подтверждает использование другого, альтернативного пути: того самого северного маршрута, ведущего в Центральную Азию в обход западной оконечности Гималаев (наряду с южной ветвью пути, которая вела в Индокитай и огибала отроги Гималаев с востока). Мы можем даже вычислить, откуда сифиты начали свой поход. Две трети поливариантности типов внешности, присущей этой крупнейшей ветви-родоначальнице евразийских народов, встречается в Индии. На долю самого Сифа пришлась примерно четверть вариантов Y-хромосомы, а на долю его сынов и потомков — большая часть остальных вариантов[312]. Еще примерно 10% потомков Сифа обитают в Центральной Азии, и, следовательно, все четверо его сынов-потомков в первом поколении имели своих представителей в Центральной Азии, что опять-таки говорит о потенциальной возможности прямого расселения их на севере как раз в то самое время, когда Евразию активно заселяли представители других групп с митохондриевой ДНК: С, X и Z.
Мы уже знаем, что три сына Сифа внесли основной вклад в колонизацию и заселение большей части Индии, Европы, Среднего Востока и стран Средиземноморья[313]. Однако в этой части нашего повествования нас интересует не столько сам Сиф и три его сына — родоначальника народов Западной Евразии, сколько его четвертый (в генетическом отношении) сын, Кришна, и особенно та роль, которую последний сыграл в формировании отцовских генов многих народов Евразии, не говоря уж о том, что его можно считать отцом практически всех коренных народов Америки.
На долю Кришны приходится около 40% типов Y-xpoмосом, обнаруженных на сегодня. Его достаточно широкое распространение в Европе, Азии, Океании и Северной и Южной Америке свидетельствует о том, что он появился на свет в Индии вскоре после первоначального исхода из Африки. Несмотря на достаточно раннее рождение и широкое распространение своей линии, расселение первых «сынов» Кришны носило весьма дискретный характер. Некоторые из них поселились в Пакистане и Индии, другие направились на север, обосновавшись на Леванте и в Центральной Азии, третьих можно встретить только в Меланезии (Новая Гвинея и окружающие острова). Наконец, представители еще одного типа (ТАТ) встречаются только в Центральной Азии и Северо-Восточной Европе[314].
Однако наиболее продуктивными в генетическом отношении сыновьями Кришны — с точки зрения заселения территории — стали два сына Кришны. Каждый из них как бы указывает векторы расселения будущих жителей Евразии и Америки в эпоху палеолита. Одна из этих ветвей, M175, или согласованный тип О, встречается почти исключительно в Восточной и Юго-Восточной Азии. Я назвал бы эту группу Хо, по имени двух выдающихся личностей этого региона: китайского мореплавателя адмирала Чжен Хо (Хэ)[315] и видного деятеля освободительного движения Вьетнама Хо Ши Мина[316]. Если представить, что эта ветвь, как и материнские линии В и F, появилась на свет, когда Кришна, продвигавшийся по прибрежной полосе океанов, прибыл из Индии в Бирму, нетрудно заметить, что Хо, в свою очередь, делится на три ветви. Все три эти ветви сегодня представлены в Китае, Индокитае и Юго-Восточной Азии и отличаются друг от друга лишь степенью расселения на севере. Одна из них осталась в Южном Китае, Индокитае и Юго-Восточной Азии, и на ее долю приходится до 65% аборигенных типов Малайзии. Вторая продолжила вектор расселения до побережья Тихого океана и проникла в Южный Китай, причем наиболее высокая концентрация представителей этой ветви отмечена на о. Тайвань. Третья проникла в глубь континентального Китая и, продвигаясь на север, вдоль побережья, достигла Японии, Кореи и Северо-Восточной Азии, а небольшое число ее представителей проникло по Шелковому пути в Центральную Азию[317]. Характерно, что ни одна из трех этих мужских ветвей, связанных с Восточной и Юго-Восточной Азией, не проникла в Америку (см рис 5.7).
В отличие от Восточной и Юго-Восточной Азии, где доминирующая роль принадлежала Хо, для жителей Северной и Центральной Азии основным источником «поставки» Y-хромосом почти исключительно служили потомки другого азиатского «сына» Кришны — Поло (М45). Характер происхождения Поло от Кришны в Индии (рис 5.8) свидетельствует о том, что ранняя ветвь послужила источником Y-маркеров и потомства, представителей которого и сегодня можно встретить среди коренных обитателей региона озера Байкал — кетов и селькупов. Наиболее любопытная в историческом отношении деталь их происхождения заключается в том, что именно Поло — тот самый «сын» Кришны, который дал импульс проникновению представителей этой ветви в Северо-Восточную Европу через Россию, тесно связан с так называемой граветтийской культурой (см главу 3). Мы вправе сопоставить эти материалы с данными распространения этой ветви на Алтае в эпоху Верхнего палеолита — от 23 до 43 тысяч лет тому назад, а также с историей распространения митохондриевых линий в Северной и Центральной Азии. По всей видимости, представители ветви Поло проникли в Центральную Азию из Пакистана примерно 40 тысяч лет назад и расселились на территориях, образующих гигантскую букву «Т». Они продвигались по Мамонтовой степи как на запад, так и на восток. На востоке они достигли берегов озера Байкал и двинулись дальше, в Америку, а на западе проникли сперва в Россию, а затем — в Восточную Европу[318].
Рис. 5.7
Именно маршрут потомков Сифа подтверждает использование другого, альтернативного пути: того самого северного маршрута, ведущего в Центральную Азию в обход западной оконечности Гималаев (наряду с южной ветвью пути, которая вела в Индокитай и огибала отроги Гималаев с востока). Мы можем даже вычислить, откуда сифиты начали свой поход. Две трети поливариантности типов внешности, присущей этой крупнейшей ветви-родоначальнице евразийских народов, встречается в Индии. На долю самого Сифа пришлась примерно четверть вариантов Y-хромосомы, а на долю его сынов и потомков — большая часть остальных вариантов[312]. Еще примерно 10% потомков Сифа обитают в Центральной Азии, и, следовательно, все четверо его сынов-потомков в первом поколении имели своих представителей в Центральной Азии, что опять-таки говорит о потенциальной возможности прямого расселения их на севере как раз в то самое время, когда Евразию активно заселяли представители других групп с митохондриевой ДНК: С, X и Z.
Мы уже знаем, что три сына Сифа внесли основной вклад в колонизацию и заселение большей части Индии, Европы, Среднего Востока и стран Средиземноморья[313]. Однако в этой части нашего повествования нас интересует не столько сам Сиф и три его сына — родоначальника народов Западной Евразии, сколько его четвертый (в генетическом отношении) сын, Кришна, и особенно та роль, которую последний сыграл в формировании отцовских генов многих народов Евразии, не говоря уж о том, что его можно считать отцом практически всех коренных народов Америки.
На долю Кришны приходится около 40% типов Y-xpoмосом, обнаруженных на сегодня. Его достаточно широкое распространение в Европе, Азии, Океании и Северной и Южной Америке свидетельствует о том, что он появился на свет в Индии вскоре после первоначального исхода из Африки. Несмотря на достаточно раннее рождение и широкое распространение своей линии, расселение первых «сынов» Кришны носило весьма дискретный характер. Некоторые из них поселились в Пакистане и Индии, другие направились на север, обосновавшись на Леванте и в Центральной Азии, третьих можно встретить только в Меланезии (Новая Гвинея и окружающие острова). Наконец, представители еще одного типа (ТАТ) встречаются только в Центральной Азии и Северо-Восточной Европе[314].
Однако наиболее продуктивными в генетическом отношении сыновьями Кришны — с точки зрения заселения территории — стали два сына Кришны. Каждый из них как бы указывает векторы расселения будущих жителей Евразии и Америки в эпоху палеолита. Одна из этих ветвей, M175, или согласованный тип О, встречается почти исключительно в Восточной и Юго-Восточной Азии. Я назвал бы эту группу Хо, по имени двух выдающихся личностей этого региона: китайского мореплавателя адмирала Чжен Хо (Хэ)[315] и видного деятеля освободительного движения Вьетнама Хо Ши Мина[316]. Если представить, что эта ветвь, как и материнские линии В и F, появилась на свет, когда Кришна, продвигавшийся по прибрежной полосе океанов, прибыл из Индии в Бирму, нетрудно заметить, что Хо, в свою очередь, делится на три ветви. Все три эти ветви сегодня представлены в Китае, Индокитае и Юго-Восточной Азии и отличаются друг от друга лишь степенью расселения на севере. Одна из них осталась в Южном Китае, Индокитае и Юго-Восточной Азии, и на ее долю приходится до 65% аборигенных типов Малайзии. Вторая продолжила вектор расселения до побережья Тихого океана и проникла в Южный Китай, причем наиболее высокая концентрация представителей этой ветви отмечена на о. Тайвань. Третья проникла в глубь континентального Китая и, продвигаясь на север, вдоль побережья, достигла Японии, Кореи и Северо-Восточной Азии, а небольшое число ее представителей проникло по Шелковому пути в Центральную Азию[317]. Характерно, что ни одна из трех этих мужских ветвей, связанных с Восточной и Юго-Восточной Азией, не проникла в Америку (см рис 5.7).
В отличие от Восточной и Юго-Восточной Азии, где доминирующая роль принадлежала Хо, для жителей Северной и Центральной Азии основным источником «поставки» Y-хромосом почти исключительно служили потомки другого азиатского «сына» Кришны — Поло (М45). Характер происхождения Поло от Кришны в Индии (рис 5.8) свидетельствует о том, что ранняя ветвь послужила источником Y-маркеров и потомства, представителей которого и сегодня можно встретить среди коренных обитателей региона озера Байкал — кетов и селькупов. Наиболее любопытная в историческом отношении деталь их происхождения заключается в том, что именно Поло — тот самый «сын» Кришны, который дал импульс проникновению представителей этой ветви в Северо-Восточную Европу через Россию, тесно связан с так называемой граветтийской культурой (см главу 3). Мы вправе сопоставить эти материалы с данными распространения этой ветви на Алтае в эпоху Верхнего палеолита — от 23 до 43 тысяч лет тому назад, а также с историей распространения митохондриевых линий в Северной и Центральной Азии. По всей видимости, представители ветви Поло проникли в Центральную Азию из Пакистана примерно 40 тысяч лет назад и расселились на территориях, образующих гигантскую букву «Т». Они продвигались по Мамонтовой степи как на запад, так и на восток. На востоке они достигли берегов озера Байкал и двинулись дальше, в Америку, а на западе проникли сперва в Россию, а затем — в Восточную Европу[318].
 Рис 5.8
Мы располагаем доказательствами столь неординарного «зеркального» распределения митохондриевых Х-маркеров в Северной и Южной Америке, а также в Европе и имеем широкую генетическую базу, свидетельствующую о том, что корни создателей мамонтовой культуры туземцев Америки восходят к жителям Северной Евразии, о чем красноречиво говорит название одного из трудов, опубликованных в 1990-е гг. — «От Костенки до Кловиса: Верхний палеолит и адаптация палео-индийцев»[319].
Представим себе переселенцев, которые двинулись из Индии и Пакистана на север, чтобы основаться на нагорьях Алтая в России. Им пришлось столкнуться с серьезными и негативными климатическими факторами, но усилия их не пропали даром, ибо игра, что называется, стоила свеч. Эта группа-родоначальница населения Северной Азии разделилась на западную и восточную ветви, заняв обширные степи. Те из них, кто направились на восток, впоследствии достигли берегов Америки, а отдавшие предпочтение западу стали прародителями этносов Северной и Западной Европы. Те же, кто решил остаться, во время последнего Ледникового периода оказались у самой его кромки и были вынуждены уйти на юг, в Центральную Азию, а впоследствии совершили повторную экспансию, став теми самыми коренными жителями Сибири и Урала, которых мы видим сегодня на этих территориях.
Итак, теперь мы знаем, что древнейшие обитатели прибрежной полосы часто делились на все новые и новые ветви в различных пунктах своего долгого пути вдоль побережья Индийского и Тихого океанов и что примерно 40 тысяч лет тому назад они заселили большую часть Азии и островов Восточного полушария. Первая из ветвей, двинувшаяся из Индии на север, стала родоначальницей охотников эпохи Верхнего палеолита, обитавших в степях Центральной Азии. Последующие же ветви, расселявшиеся вдоль русел великих рек Центральной Азии, дали начало той самой расе, представителей которой мы сегодня называем монголоидами. Свидетельствами о существовании в Азии так называемого монголоидного типа вплоть до максимума Ледникового периода, имевшего место 20 тысяч лет назад, мы не располагаем. Таким образом, эволюционные факторы, обусловившие возникновение этой физической дивергенции, могли действовать и долгое время спустя после прибытия первых выходцев из Центральной Азии в Северную Азию. А это подводит нас к следующему этапу климатической истории нашей планеты — Великому Оледенению.
Рис 5.8
Мы располагаем доказательствами столь неординарного «зеркального» распределения митохондриевых Х-маркеров в Северной и Южной Америке, а также в Европе и имеем широкую генетическую базу, свидетельствующую о том, что корни создателей мамонтовой культуры туземцев Америки восходят к жителям Северной Евразии, о чем красноречиво говорит название одного из трудов, опубликованных в 1990-е гг. — «От Костенки до Кловиса: Верхний палеолит и адаптация палео-индийцев»[319].
Представим себе переселенцев, которые двинулись из Индии и Пакистана на север, чтобы основаться на нагорьях Алтая в России. Им пришлось столкнуться с серьезными и негативными климатическими факторами, но усилия их не пропали даром, ибо игра, что называется, стоила свеч. Эта группа-родоначальница населения Северной Азии разделилась на западную и восточную ветви, заняв обширные степи. Те из них, кто направились на восток, впоследствии достигли берегов Америки, а отдавшие предпочтение западу стали прародителями этносов Северной и Западной Европы. Те же, кто решил остаться, во время последнего Ледникового периода оказались у самой его кромки и были вынуждены уйти на юг, в Центральную Азию, а впоследствии совершили повторную экспансию, став теми самыми коренными жителями Сибири и Урала, которых мы видим сегодня на этих территориях.
Итак, теперь мы знаем, что древнейшие обитатели прибрежной полосы часто делились на все новые и новые ветви в различных пунктах своего долгого пути вдоль побережья Индийского и Тихого океанов и что примерно 40 тысяч лет тому назад они заселили большую часть Азии и островов Восточного полушария. Первая из ветвей, двинувшаяся из Индии на север, стала родоначальницей охотников эпохи Верхнего палеолита, обитавших в степях Центральной Азии. Последующие же ветви, расселявшиеся вдоль русел великих рек Центральной Азии, дали начало той самой расе, представителей которой мы сегодня называем монголоидами. Свидетельствами о существовании в Азии так называемого монголоидного типа вплоть до максимума Ледникового периода, имевшего место 20 тысяч лет назад, мы не располагаем. Таким образом, эволюционные факторы, обусловившие возникновение этой физической дивергенции, могли действовать и долгое время спустя после прибытия первых выходцев из Центральной Азии в Северную Азию. А это подводит нас к следующему этапу климатической истории нашей планеты — Великому Оледенению.
ГЛАВА ШЕСТАЯ ВЕЛИКОЕ ОЛЕДЕНЕНИЕ
В последней главе мы видели, что Восточная и Центральная Азия были заселены как бы тремя волнами генетической колонизации, берущими свое начало из Индии. Древнейшие поселенцы следовали по пути собирателей на прибрежной полосе, продвигавшимися вдоль побережья из Индии через Индокитай к берегам Японии и Кореи, основывая на пути своей миграции колонии и поселения. Выходцы из этих поселений на побережье впоследствии проникли в глубинные районы Центральной Азии и вышли к берегам великих рек Азии, миновав проходы и перевалы в сплошной стене огромной горной гряды, протянувшейся с востока на запад и как бы обрамляющей Гималаи. Юго-Восточный Тибет и Цзиньхайское плато вполне могли стать первым районом Центральной Азии, куда около 60 тысяч лет тому назад проникли мигранты из Бирмы и Индокитая. В то же время мигранты, продвигавшиеся вверх по течению Инда из Пакистана, достигли предгорий Алтая (Россия), расположенного далеко на западе. Их миграция могла состояться во время мягкой климатической паузы, имевшей место около 43 тысяч лет тому назад. Те же из переселенцев, которые, продвигаясь вдоль побережья, проникли в Северный Китай, могли направиться на запад вслед за течением Хуанхэ (Желтой реки) и попасть в Центральную Азию примерно в то же время или чуть позже. Карта заселения Северной Азии понемногу начала заполняться. Около 30 тысяч лет назад обширная полоса земель на территории бывшего Советского Союза, протянувшаяся от Алтая через озеро Байкал в Южной Сибири до реки Алдан на востоке, подверглась активной колонизации первопроходцами-людьми современного типа, владевшими техническим навыками и орудиями, современными памятникам эпохи палеолита в Европе (рис. 6.1). Мигранты проникли даже в районы, лежащие за Полярным кругом, к северу от Уральского хребта; они поселились там примерно 40 тысяч лет тому назад. Есть данные, свидетельствующие о том, что технические достижения и навыки эпохи Верхнего палеолита, возникшие в Северной Евразии, могли быть перенесены на территорию нынешней Внутренней Монголии вплоть до северной излучины Желтой реки, но, судя по имеющимся находкам, подобное технико-культурное влияние не распространилось далее на юг и не проникло в Китай[320]. Предложенная Дэйлом Гатри концепция, согласно которой в ту эпоху охотники и собиратели бродили по необъятным просторам Мамонтовой степи, простиравшейся от Восточной Сибири до Центральной Европы (см. главу 5), получает реальные подтверждения благодаря археологическим находкам, относящимся к эпохе около 30 тысяч лет тому назад. Однако его предположения о том, что прародиной «монголоидной» расы были южные окраины Великой степи, носит слишком гипотетический характер, учитывая конкретику того времени и реалии Сибири. Для начала напомним, что, как уже было сказано выше, пока не найдено человеческих останков бесспорно монголоидного типа, относящихся к столь отдаленной эпохе. Насколько нам известно, древние обитатели Южной Сибири выглядели практически так же, как кроманьонцы, населявшие Европу. У них сложилась такая же культура; по крайней мере, согласно генетической истории, которую донесли до нас линии Y-хромосом, у них было немало общих генов с обитателями Европы (см. главу 5).

 Рис. 6.1
В качестве аргумента в пользу того, что один из центров культуры той эпохи действительно мог существовать на Западе, можно отметить, что расцвет культуры Мамонтовой степи в те времена имел место далеко на западе, в Центральной и Восточной Европе (см. Фото 19). Следы материальной культуры Мамонтовой степи обнаружены в Мальте, неподалеку от озера Байкал. Они датируются временем около 2000 лет тому назад (см. Фото 13,глава 3). Эта дата всего на 2000 лет старше, чем древнейшие человеческие останки монголоидного типа, найденные чуть западнее, в том же районе Южной Сибири, на стоянке Афонтова Гора[321].
Рис. 6.1
В качестве аргумента в пользу того, что один из центров культуры той эпохи действительно мог существовать на Западе, можно отметить, что расцвет культуры Мамонтовой степи в те времена имел место далеко на западе, в Центральной и Восточной Европе (см. Фото 19). Следы материальной культуры Мамонтовой степи обнаружены в Мальте, неподалеку от озера Байкал. Они датируются временем около 2000 лет тому назад (см. Фото 13,глава 3). Эта дата всего на 2000 лет старше, чем древнейшие человеческие останки монголоидного типа, найденные чуть западнее, в том же районе Южной Сибири, на стоянке Афонтова Гора[321].
 Ранние фигурки граветтийской культуры, выполненные (слева направо) из обожженной глины (Моравия), мамонтовой кости (Франция), известняка (Виллендорфская Венера, Австрия) и кости (Украина).
Ранние фигурки граветтийской культуры, выполненные (слева направо) из обожженной глины (Моравия), мамонтовой кости (Франция), известняка (Виллендорфская Венера, Австрия) и кости (Украина).
 Фото 19. Резные изображения северного оленя Позднего Верхнего палеолита: на конце бивня мамонта — Монтастрюк, Франция (вверху), и на костяном метателе для дротиков — Ложери Бассе, Франция (внизу).
Фото 19. Резные изображения северного оленя Позднего Верхнего палеолита: на конце бивня мамонта — Монтастрюк, Франция (вверху), и на костяном метателе для дротиков — Ложери Бассе, Франция (внизу).
Великое оледенение: льды, озера, пустыни
Когда на часах эпохи палеолита появилась дата 20 тысяч лет тому назад, изменения наклона земной оси и влияния на ее орбиту космических факторов, возникших за многие сотни миллионов километров от нашей планеты, приняли весьма драматичный характер. Три крупнейших центра гравитации нашей Солнечной системы сблизились и совместились, в результате чего даже летом поверхности Земли в Северном полушарии стал достигать абсолютный минимум солнечного тепла[322]. Климат стал гораздо холоднее, а непродолжительные периоды потепления, так называемые междустадиальные паузы, характерные для эпохи между 30 и 50 тысячами лет тому назад, и вовсе отошли в прошлое. Между тем именно эти теплые промежуточные периоды и солнечное тепло способствовали активному таянию льдов в Северном полушарии и препятствовали сползанию ледниковых плит полярной «шапки» планеты по всему Скандинавскому полуострову в Северную Европу. И вот теперь эти ледники получили возможность распространяться по всему северу. Уровень океана начал быстро понижаться, достигнув отметки 130 м ниже современного. Короче говоря, на Земле наступило последнее из пережитых ею оледенений — Ледниковый период. (За последние 100 тысяч лет Земле не раз доводилось испытать похолодания, и поэтому археологи склонны называть это великое оледенение Последним ледниковым максимумом (ПЛМ), а не просто очередным ледниковым периодом.) Последний ледниковый максимум повлек за собой куда более драматические миграции и переселения обитателей северных регионов, чем в любую другую эпоху истории человечества. Анализ климатической карты нашей планеты в эпоху около 18 тысяч лет назад позволяет нам понять, почему это произошло. Громадные пространства на нашей планете вскоре стали полностью необитаемыми. Для начала надо сказать, что громадные ледяные плиты, толщина которых в отдельных районах достигала 5 км, делали территории, на которых распространялся ледник, совершенно непригодными для обитания. Однако белые плиты ледников распределялись по поверхности Северного полушария далеко неравномерно. В Европе, например, они захватили главным образом центральные и северовосточные регионы (см. рис. 6.2). Так, на Британских островах, составлявших в те времена единое целое с континентальной Европой, земли были покрыты сплошным ледниковым панцирем вплоть до нынешнего Оксфорда. А в Скандинавии до сих пор сохранились неизгладимые следы продвижения ледника — озера и фьорды, а также возникшее в ту эпоху резкое проседание земной коры, известное в наши дни под названием Балтийского моря. Северная Германия, Польша и государства Балтии находились у самой кромки мощного ледникового панциря, который простирался на северо-восток через весь Полярный круг, Финляндию и Карелию вплоть до Архангельска и далее, до Северного Урала. Южнее, в Центральной Европе, границами продвижения языков ледника служили горные районы: Пиренеи, Центрально-Европейский горный массив, Альпы и Карпаты. Что касается Восточной Европы, то она, как мы увидим ниже, была затронута ледником значительно меньше, чем Западная. Азии в эпоху оледенения повезло гораздо больше, чем Европе. Большая часть Северной и Центральной Азии оказалась незатронутой ледником (см. рис. 6.3). Лишь у восточной оконечности Североуральского хребта огромная ледниковая шапка полностью накрывала современный полуостров Таймыр и продвинулась к югу от него. Другой частью азиатского континента, где могла возникнуть ледниковая шапка, было обширное Тибетское плато, расположенное далеко на юге. В результате того, что плато было сильно приподнято над уровнем моря, на нем свирепствовали сильные холода. Однако присутствие на нем ледниковых плит вызывает серьезные сомнения, поскольку существуют материальные свидетельства обитания человека на Тибетском плато, относящиеся к эпохе задолго до Последнего ледникового максимума[323]. Ледник сильно затронул Северную Америку, в частности территорию Канады и район Великих озер, и штаты крайнего северо-востока США, то есть, другими словами, его владения простирались на добрых две трети континента. Ледниковые плиты севера как бы уравновешивались двумя мощными ледяными панцирями на востоке, соединенными с ледяной шапкой Гренландии. Что касается Аляски, то она в те времена была связана с Сибирью посредством громадного и свободного от льда моста, роль которого выполнял давно ушедший под воду континент Берингия. На нем, как и в Азии, ледниковых плит практически не было. Большая из двух ледниковых плит американского континента, так называемая Лаврентидская плита, на востоке оставила свой отпечаток в виде глубокой впадины в земной коре, имеющей форму обширного внутреннего моря, более известного как Гудзонов залив. В некоторых регионах Евразии и Америки вокруг прежних ледниковых шапок и плит возникли огромные водоемы, известные под названием ледниковых озер. Наибольшей известностью среди таких озер пользуются знаменитые Великие озера[324] в Северной Америке. Кстати сказать, ледяные плиты тоже не были статичными, а постоянно перемещались и двигались, как глетчеры. Эти застывшие реки не только прорезали в поверхности огромные русла будущих рек и фьорды, но и уничтожили подавляющее число свидетельств того, что люди жили на севере в доледниковый период. Лед в эпоху Великого Оледенения служил не единственной преградой на пути расселения людей. В те времена невиданного прежде размаха достигли и площади пустынь. Громадные пространства вокруг ледяных шапок превратились в ледяную пустыню, в которой могли выжить лишь наиболее жизнестойкие виды растений и животных. На территории Европы полярная пустыня простиралась на восток от южных окраин Англии через всю северную Германию до южной оконечности финно-скандинавской ледниковой плиты. Весь обширный регион от Леванта и Красного моря до Пакистана, и без того достаточно засушливый, превратился в крайне сухую пустыню. Южные районы Центральной Азии, от Туркменистана, Каспийского моря и Узбекистана на западе до Синьцзянь-Уйгурского района (лежащего к северу от Тибета) и Внутренней Монголии на востоке, превратились в полосу засушливых пустынь, простиравшуюся по обе стороны 40-й параллели. Эта пустыня, быстро сменившая легендарную Мамонтовую степь Гатри, навсегда отделила Северную Азию и северные районы Центральной Азии от Восточной и Юго-Восточной Азии.
Беженцы Ледникового периода
Население Африки, как и его далекие предки за последние 2 млн. лет, жестоко страдало во время каждого очередного Ледникового периода. Площадь Сахары в такие периоды резко увеличивалась, накрывая практически всю Северную Африку; пустыня Калахари охватывала почти всю Юго-Западную Африку, а большую часть территории африканского континента к югу от Сахары занимали маловодные травянистые саванны, на которых практически не было лесов и кустарников. Великие тропические леса Центральной и Западной Африки превращались в скудные островки, ограниченные Центральной Экваториальной Африкой и южными районами побережья Гвинеи на западе Африки. В Восточной Африке экспансия засушливых саванн точно так же отрезала жителей востока от охотников и собирателей, обитавших на юге континента. Нам, жителям Северной и Центральной Европы, вместе с американцами обожающим строить ледяные замки, остается только недоумевать и гадать, какая участь постигла древних европейцев анатомически современного типа. Сумели ли они спастись, покинув родные места, или же полностью вымерли, уступив на арене истории место переселенцам со Среднего Востока? Наши двоюродные братья, неандертальцы, вымерли примерно за 10 тысяч лет до последнего ледникового максимума (см. главу 2). Однако археологические находки со всей определенностью говорят о том, что люди действительно жили в Европе до Последнего ледникового максимума, но затем они оказались оттесненными на юг, в три или, может быть, четыре зоны умеренно (по меркам Ледникового периода) холодного климата. Генетические следы позволяют нам получить массу интереснейших сведений о происхождении и этническом составе этих беженцев Ледникового периода, но для начала давайте обратимся к археологическим находкам, чтобы реконструировать материальный фон той эпохи.
 Рис. 6.2
Большая часть территории Северной Европы в эпоху Последнего ледникового максимума (ПЛМ) была необитаемой. В то же время в Южной Европе существовали три основные региона, куда устремлялись беженцы эпохи палеолита (см. рис. 6.2). Если двигаться с запада на восток, то первое из таких прибежищ включало в себя земли современных Испании и Франции по обе стороны Пиренеев, в частности — Страну Басков (Басконию). Для него были характерны тонко проработанные «листообразные» каменные орудия солютрийской культуры (название которой происходит от небольшой французской деревушки Солютр). Обосновавшиеся здесь беженцы, принесшие с собой свои технические знания с северо-западных окраин континента, в культурном отношении резко отличались от других беженцев, обживавших южные районы и бывших носителями традиции, техника обработки каменных изделий у которой получила название эпи-граветтийской культуры.
Вторая область притока беженцев находилась в Италии, и занятия населения в тех местах носили более или менее локальный характер. Наконец, третья зона притока беженцев находилась в Украине, точнее — в обширном регионе к северу от Черного моря, ограниченном двумя великими реками — Днепром и Доном и отделенном от остальной части Южной Европы Карпатскими горами, которые в эпоху ПЛМ были частично накрыты ледниковым панцирем. В Центральной Европе существовали еще как минимум два района, служившие своего рода прибежищами для беженцев. Один из них — западная Словакия, чуть к югу от отрогов Карпат, другой — бассейн реки Днестр в Молдавии, лежащий чуть к востоку от Карпат, на северозападном побережье Черного моря[325].
Именно эти стоянки на территории Восточной Европы стали центрами позднего цветения культур Мамонтовой степи эпохи Верхнего палеолита. Когда же Последний ледниковый максимум достиг своего апогея, центр активности представителей этой культуры переместился из западной Словакии на восток, на территорию нынешних Молдавии и Украины, а также на юг, в Венгрию. Именно в Украине и далее к северу, на берегах Днепра и Дона, на великой Русской равнине мы находим наиболее хорошо сохранившиеся следы не только постоянного присутствия, но и расселения человека в Восточной Европе в эпоху Великого Оледенения[326].
Рис. 6.2
Большая часть территории Северной Европы в эпоху Последнего ледникового максимума (ПЛМ) была необитаемой. В то же время в Южной Европе существовали три основные региона, куда устремлялись беженцы эпохи палеолита (см. рис. 6.2). Если двигаться с запада на восток, то первое из таких прибежищ включало в себя земли современных Испании и Франции по обе стороны Пиренеев, в частности — Страну Басков (Басконию). Для него были характерны тонко проработанные «листообразные» каменные орудия солютрийской культуры (название которой происходит от небольшой французской деревушки Солютр). Обосновавшиеся здесь беженцы, принесшие с собой свои технические знания с северо-западных окраин континента, в культурном отношении резко отличались от других беженцев, обживавших южные районы и бывших носителями традиции, техника обработки каменных изделий у которой получила название эпи-граветтийской культуры.
Вторая область притока беженцев находилась в Италии, и занятия населения в тех местах носили более или менее локальный характер. Наконец, третья зона притока беженцев находилась в Украине, точнее — в обширном регионе к северу от Черного моря, ограниченном двумя великими реками — Днепром и Доном и отделенном от остальной части Южной Европы Карпатскими горами, которые в эпоху ПЛМ были частично накрыты ледниковым панцирем. В Центральной Европе существовали еще как минимум два района, служившие своего рода прибежищами для беженцев. Один из них — западная Словакия, чуть к югу от отрогов Карпат, другой — бассейн реки Днестр в Молдавии, лежащий чуть к востоку от Карпат, на северозападном побережье Черного моря[325].
Именно эти стоянки на территории Восточной Европы стали центрами позднего цветения культур Мамонтовой степи эпохи Верхнего палеолита. Когда же Последний ледниковый максимум достиг своего апогея, центр активности представителей этой культуры переместился из западной Словакии на восток, на территорию нынешних Молдавии и Украины, а также на юг, в Венгрию. Именно в Украине и далее к северу, на берегах Днепра и Дона, на великой Русской равнине мы находим наиболее хорошо сохранившиеся следы не только постоянного присутствия, но и расселения человека в Восточной Европе в эпоху Великого Оледенения[326].
Генетическое постоянство и Последний ледниковый максимум
Способна ли генетическая реконструкция поведать нам нечто большее о жителях того или иного района, то есть рассказать, откуда они пришли и куда направились дальше, а не просто создать обычную культурно-историческую панораму, показывающую, чем они занимались в ту эпоху, когда на них обрушились холодные объятия ледника? Генетическое отслеживание может согласовываться и действительно близко согласуется с археологическими данными по той эпохе, но оно сообщает нам и нечто большее о генетических корнях происхождения европейцев, в частности о том, что 80% генетических линий современных жителей Европы происходят от предков, которые уже жили в Европе задолго до Великого Оледенения. В главе 3 мы уже говорили о том, как материнский клан HV расселился из Восточной Европы, проникнув в Северную и Западную Европу, что, возможно, ознаменовалось появлением граветтийских культур примерно 33 тысячи лет тому назад. Линии клана HV сегодня распространены достаточно широко по всей Европе. Что касается линии Н, то она является наиболее распространенной и общей практически у всех европейцев. Однако так было далеко не всегда, и другая ветвь клана Н, V, которая, по всей вероятности, появилась на свет в Стране Басков, показывает, почему это произошло[327]. Археологические данные указывают, что прибежище на юго-западе Европы, в Стране Басков с юго-запада, стало новой родиной для мигрантов и культур, пришедших с северо-запада Европы на начальном этапе Последнего ледникового максимума. А поскольку Западная Европа отделена от Италии высокими горами, у нас есть все основания ожидать, что по окончании Ледникового периода процесс миграции мог пойти в обратном направлении, когда начался массовый исход уцелевших жителей из Страны Басков на восток и на север, вдоль побережья Атлантического океана. Это именно тот маршрут, на который указывает расселение материнской подгруппы V в послеледниковый период, когда V, отличавшаяся наиболее высоким многообразием и встречаемостью, а также древностью именно в Стране Басков, направилась на север и сегодня в небольших количествах встречается в Италии. V появилась в Стране Басков вскоре после Последнего ледникового максимума. Возраст ее предка, пре-V, согласно последним оценкам, составляет 26 400 лет, то есть, другими словами, он появился на свет задолго до ПЛМ. Пре-V в наши дни все еще встречается на Балканах и в Закавказье, что свидетельствует о восточном происхождении этой подгруппы. Данные о расселении V в послеледниковый период (16 300 лет на западе Европы) вполне согласуются с подобным сценарием событий. Точно такую же картину мы наблюдали у Y-хромосомного маркера Руслана, который, как мы знаем, проник в Европу с востока и обосновался в Северной и Западной Европе (глава 3). Данные последних исследований говорят о том, что наиболее высокую (около 90%) встречаемость Руслан имеет в Стране Басков (Испания); за ней следуют очень высокие показатели встречаемости, отмечаемые в Западной и Северной Европе[328]. С другой стороны, Италия, отгороженная хребтами Альп, в меньшей мере играла роль спасительного прибежища для жителей северных регионов, чем зоны умеренного климата на побережье Средиземного моря, где человек жил постоянно еще с эпохи, предшествовавшей ПЛМ. Этот факт получает подтверждение благодаря отмечаемой в том районе весьма высокой (более 1/3) доле линий мтДНК, сохранившейся с доледникового периода. Эти примеры показывают, что для таких зон-прибежищ было характерно широкое распространение линий, сложившихся в них в эпоху Последнего ледникового максимума, а также высокий уровень более архаических линий, сохранявшихся в них еще с доледниковой эпохи. Последняя картина особенно характерна для Украины, где, по данным археологических находок, сохранился как минимум 31% ранних материнских линий, восходящих к доледниковой эпохе. В странах Юго-Восточной Европы и на Балканах, где также сохранилось немало материнских линий доледниковой эпохи, эти показатели несколько ниже — 24—26% [329]. Мне остается сказать, что хотя от 20 до 34% линий мтДНК у современных европейцев восходят к доледниковой эпохе, это отнюдь не означает, что остальные линии, выявленные у них, проникли в Европу извне и непременно после Последнего ледникового максимума. Нет и еще раз нет. Они по большей части имеют местное происхождение. Из всей совокупности генетических линий современных европейцев примерно 55% возникли и сформировались в эпоху сразу же по окончании Ледникового периода (Поздний Верхний палеолит), но они, как и гаплогруппа V, по-видимому, произошли от уже существовавших в Европе линий и просто-напросто отражают волну расселения народов в послеледниковый период, то есть, другими словами, являются новыми ветвями от старого ствола. На долю мигрантов с Ближнего Востока, действительно проникших в Европу в эпоху неолита (начиная с 8 тысяч лет тому назад), приходится всего лишь 15% от общего числа генетических линий, присутствующих сегодня в Европе[330]. Одним из интереснейших открытий в области выявления генетической структуры населения Юго-Восточной Европы явилась локализация так называемого маркера Адама в Румынии. Дело в том, что в эпоху Последнего ледникового максимума Карпатские горы были покрыты ледниками и создавали практически непреодолимую преграду между Юго-Восточной Европой и прибрежными районами Черного моря. Так вот, ледяная стена Карпат как бы поделила Румынию пополам. Сегодня Карпаты, давно утратившие свои мощные ледяные плиты, уже не составляют сколько-нибудь серьезной преграды для человека, однако они остаются четко выраженной генетической границей. Это ясно показывают Y-хромосомные генные маркеры, характерные для северо-восточной Европы и Украины и обладающие очень высокой встречаемостью к востоку от Карпат, тогда как для западных предгорий Карпат более характерны маркеры, распространенные в Западной Европе. Однако эта микрорегиональная граница выглядит весьма размытой из-за доминирующего присутствия линии M17, главной Y-хромосомной линии Восточной Европы. M17 можно считать следствием первоначального проникновения носителей восточной ветви граветтийской культуры откуда-то с востока, имевшего место в доледниковую эпоху. Эта линия очень широко представлена во всей Восточной Европе — от Польши и Словакии до Венгрии и Украины. Кроме того, линия М17 и сегодня часто встречается среди славянского населения Балкан, что можно считать свидетельством существования на Балканах в эпоху ледникового максимума еще одного прибежища для беженцев из Северной Европы[331].
Северная Азия в Ледниковый период
В эпоху Последнего ледникового максимума на Гималаях тоже возникли ледяные шапки, а по обе стороны 40-й параллели и даже далеко к югу от нее, в Центральной Азии, появилась обширная полоса пустынь. Население южных районов Центральной Азии резко сократилось, хотя, по всей вероятности, не исчезло совсем, по крайней мере — на Тибете и в его окрестностях. Полоса пустынь и горные хребты, покрытые льдами, вполне могли стать преградой, отделившей Южную, Восточную и Юго-Восточную Азию от Центральной и Северной Евразии, но это отнюдь не означает, что жизнь в центральных и северных районах оказалась полностью уничтоженной. Во время ПЛМ граница распространения вечной мерзлоты опустилась вплоть до 50-й параллели. Вечная мерзлота не является препятствием для жизни человека в приполярной тундре. Мы располагаем материальными доказательствами того, что люди жили значительно севернее от границы вечной мерзлоты еще в доледниковую эпоху, в частности на Русской равнине и далее на северо-востоке, в районе Талицкого озера в Сибири. Охотники и собиратели Южной Сибири постоянно жили в целом ряде древних стоянок, особенно таких, как Афонтова Гора на берегу Енисея, где и создали свою собственную, уникальную южносибирскую культуру. Полосы приполярной тундры и лесостепи, богатой всевозможными видами травянистой растительности, до сих пор простираются от Русской равнины на западе до северовостока Сибири (см. рис. 6.3). Несмотря на резкие перепады температур и колебания климата, здесь существовали вполне благоприятные условия для жизни охотников, которые смогли бы адаптироваться к климату. И они действительно адаптировались к нему, о чем недвусмысленно говорят находки на целом ряде древних археологических стоянок в этой полосе, датируемых как раз этим периодом[332]. И вновь генетические данные говорят нам о том, что эти суровые охотники, жившие на вечной мерзлоте, сохранили некоторые из древних материнских линий, восходящих еще к доледниковому периоду, и что после потепления произошло активное расселение целого ряда подгрупп. Как я уже сказал в главе 5, линии А, С и Z являются характерными для Северной Азии, тогда как D простирается далее к югу, в Китай. Из числа этих линий А сохранилась еще с доледникового периода, то же самое можно сказать и о различных вариантах линий Манью и Насрин. В эпоху между 10 и 17 тысячами лет тому назад в расселении этих линий возникла продолжительная пауза, а затем началось новое распространение вариантов таких линий, как D, А, С и, наконец, Z[333]. (См. рис 6.3.) Генетические данные по Северной и Центральной Азии просто-напросто подтверждают те же самые выводы, которые мы вправе ожидать от археологии. Когда наступало очередное похолодание, охотники, жившие в Мамонтовой степи, уходили на юг, к ее окраинам, а их численность сокращалась. Когда же возвращалось тепло, люди опять возвращались в степи и начинали быстро обживать их. Для нас особенно интересен вопрос о том, как выглядели эти люди и какие народы сегодня являются их ближайшими потомками. Почему и когда именно монголоиды стали доминирующим типом на необъятных пространствах Азии и Северной и Южной Америки? Не может ли генетическая история поведать нам нечто такое, что ускользает от сухих свидетельств археологии?
 Рис.6.3
Рис.6.3
Изгнанники холодных степей
В последней главе я уже затронул некоторые из этих вопросов, в частности — вопрос о генетических корнях происхождения монголоидов. Я попытался показать, что целый ряд имеющихся свидетельств указывают на Южную Сибирь, верховья реки Енисей и район озера Байкал как возможные локусы дальнейшей физической специализации северных монголоидов в эпоху палеолита. Древнейшими останками скелетов евразийцев, относящихся к типу внешности монголоидов, является фрагмент черепа, найденный на стоянке Афонтова Гора в Южной Сибири, датируемый временем около 21 тысячи лет тому назад, то есть задолго до Последнего ледникового максимума (ПЛМ) (см. главу 5). Однако эта находка, оставаясь единственной, является не слишком убедительной, ибо ни в каком другом регионе Старого Света, включая Восточную Азию, пока не обнаружено аналогичных типов, относящихся к доледниковому периоду или даже к послеледниковой эпохе. К тому же останки скелетов, относящиеся к эпохе до 11 тысяч лет тому назад, являются исключительной редкостью во всем Дальневосточном регионе. Подобная пауза в находках создает впечатление — которое иной раз воспринимается как догма, — что в Восточной Азии вплоть до 7—10 тысяч лет тому назад вообще не было монголоидных типов, а в Юго-Восточной Азии — и гораздо позже этого времени[334]. Важная проблема, связанная с этой паузой в находках ископаемых останков, — как объяснить тот факт, что на долю монголоидного типа внешности приходится более 99% людей, живущих сегодня в этом регионе. Более того, именно этот, монголоидный, тип является наиболее распространенным среди всего населения Земли. Действительно, поскольку большинство коренных жителей Северной и Южной Америки относятся к монголоидам и, за редкими исключениями, являются выраженными представителями синодонтного типа, они в географическом отношении были наиболее распространенным типом внешности задолго до того, как Колумб совершил свое знаменитое плавание в Америку, необратимо изменившее ход ее истории. Право, не надо обладать особым даром воображения, чтобы понять, что подобный тип внешности возник у северных монголоидов в качестве адаптации к суровому, ветреному и холодному климату географически изолированного региона Южной Сибири. Главными занятиями человека в ту эпоху оставались охота и собирательство. Как же могло случиться, что эти бедные охотники и собиратели, ведущие отчаянную борьбу за выживание, расселились по доброй половине мира и быстро стали доминирующим типом человека в послеледниковую эпоху? Эта загадочная история хранит свои тайны, запечатленные в наших генах. Другая проблема, с которой нам приходится сталкиваться перед лицом позднего и быстрого расселения монголоидов, — это датировка времени заселения монголоидами Америки. В следующей главе мы рассмотрим археологические и генетические свидетельства, позволяющие определить дату их первого проникновения на американский континент, и попытаемся показать, что такая дата свидетельствует о том, что все основные генетические линииродоначальницы проникли в Северную Америку задолго до Последнего ледникового максимума. Храня в памяти образы легендарных пионеров[335] Америки, я думаю, что концепция о том, что расселение монголоидов в Восточной и Юго-Восточной Азии имело место лишь в последние несколько тысяч лет, является совершенно ошибочной, поскольку подавляющее большинство генетических наследственных линий, характерных для типа южных монголоидов, берут свое начало именно в этом регионе. Однако невозможно назвать какие-то очевидные причины, по которым обширная и наделенная специфическими чертами внешнего облика группа охотников, обитавшая в Центральной Азии, могла расселиться настолько быстро, чтобы стать доминирующей во всей Восточной Азии в самом начале эпохи неолита, менее 10 тысяч лет тому назад. На мой взгляд, возникновение, миграции и расселение монголоидов относятся к куда более ранним временам. Более того, они продолжались вплоть до сравнительно недавнего времени. Но прежде чем перейти к более детальному рассмотрению этого вопроса, позвольте мне вкратце изложить в последующих двух параграфах свое мнение о том, как это могло происходить. Из-за отсутствия бесспорных археологических свидетельств моя гипотеза по большей части базируется на генетических данных. В эпоху Верхнего палеолита, то есть от 40 тысяч лет тому назад до Последнего ледникового максимума, между Тибетским плато и Южной Сибирью простирались огромные степи — заповедные места для охоты. В них обитали умелые охотники, которые вели свою генетическую родословную из самых разных и часто весьма удаленных друг от друга районов Азии (см. главу 5). Прежде всего, я думаю, что Дэйл Гатри во многом прав, утверждая, что северные монголоиды сумели выработать особый вариант монголоидного типа внешности, более приспособленный к крайне холодному и ветреному климату высокогорья в южной и восточной части Великой степи. В то же время далее на западе, в более равнинных районах, охотники по своему внешнему облику более походили на европейцев. Когда же ледниковые плиты начали смыкать свои смертоносные объятия, охотники, жившие на степных плато, оказались в ледяном плену. Те же, кто жили к югу от этой прародины, в юго-восточном Тибете, были вынуждены покинуть высокогорное, холодное и крайне засушливое Тибетское плато и отправиться вниз по течению великих рек Азии в Китай и Индокитай. Те, кто охотились на Цзиньхайском плато, раскинувшемся к северо-востоку от Тибета, двинулись вниз по течению Хуанхе в Северный Китай. Наконец, те, кто обитали к северу от 40-й параллели на нагорьях Синьцзяня и Монголии, были оттеснены еще дальше к северу быстро распространявшимися песчаными пустынями Такламакан и Гоби, которые простирались от южных районов Центральной Азии до обширных низменностей Южной Сибири, в частности окрестностей стоянки Афонтова Гора на берегах Енисея (рис. 6.4). Главная проблема, с которой столкнулись охотники, стремившиеся найти более теплые и пригодные для жизни земли, естественно, заключалась в том, что чем ближе они приближались к Полярному кругу, тем выше были их шансы оказаться в еще более холодных и ветреных краях. Из этой ледяной ловушки было только одно спасение — миграция на северо-восток. Люди могли двинуться туда по узкой полосе тундры, протянувшейся от р. Лены до Якутска, а затем — на восток, к берегам Охотского моря, где климат на побережье был куда более мягким. Поскольку уровень Мирового океана, установившийся в ту эпоху, был очень низким, широкая прибрежная полоса низменных земель, далеко выдававшаяся в Охотское море, находилась выше уровня воды. Другой спасительный путь из ледяного плена, пролегавший от окрестностей озера Байкал до побережья Тихого океана, проходил чуть южнее, вдоль по течению великой сибирской реки Амур, к берегам Тихого океана. А прибрежная полоса Тихого океана, на которой установился весьма умеренный климат, была удобным коридором в Америку. Такая миграция охотников из Центральной Азии сразу по трем потенциальным направлениям могла стать источником возвращения монголоидов в те самые прибрежные районы Восточной Азии, из которых их далекие предки в свое время, за 20 тысяч лет или даже раньше, отправились на север.
 Рис 6.4
Южный Китай вполне мог быть зоной активного расселения монголоидного населения после их массового исхода со степных плоскогорий, но, судя по генетическим данным, большинство линий мтДНК в Юго-Восточной Азии имеют локальный характер и всегда присутствовали в том регионе. Такие группы, как В и F, которые считались типичными для монголоидного населения, имеют на юге очень глубокую древность[336]. Что же касается внешнего облика обитателей прибрежной полосы, то они выглядят совершенно иначе.
Рис 6.4
Южный Китай вполне мог быть зоной активного расселения монголоидного населения после их массового исхода со степных плоскогорий, но, судя по генетическим данным, большинство линий мтДНК в Юго-Восточной Азии имеют локальный характер и всегда присутствовали в том регионе. Такие группы, как В и F, которые считались типичными для монголоидного населения, имеют на юге очень глубокую древность[336]. Что же касается внешнего облика обитателей прибрежной полосы, то они выглядят совершенно иначе.
Прибрежная полоса была более обширной
Эпоха наканунеПоследнего ледникового максимума и во время него была не только временем быстрого сокращений территорий, пригодных для жизни человека, и активного поиска спасительных прибежищ в новых местах. Как это ни парадоксально звучит, человек получил возможность обживать новые обширные земли, появившиеся над водой в результате резкого понижения уровня океана, вызванного оледенением и концентрацией громадных масс воды в ледниковых плитах. Огромные прибрежные пространства по всему побережью Азии от Индии до Сибири расширились на многие сотни километров благодаря отступлению океана практически к краю континентального шельфа. Сибирь соединялась с Северной Америкой сухопутным мостом, роль которого выполнял новый обширный материк Берингия. Восточно-Китайское море и Желтое море превратились в сухопутные массивы. Японские острова оказались соединенными с азиатским континентом через остров Сахалин. Климат на этих островных территориях был куда более мягким и теплым, чем в степях Центральной Азии. Австралия и Новая Гвинея были связаны сушей и составляли единый огромный континент Сахул. Индийский субконтинент значительно расширил свою площадь и соединился просторным сухопутным перешейком с нынешним островом Шри-Ланка (Цейлон). Регионом, который более всего расширил свои сухопутные территории, стала Юго-Восточная Азия, где Южно-Китайское море, Бангкокский залив и Яванское море быстро высохли и стали сушей, соединив Индокитай, Малайзию и крупнейшие острова Индонезии в гигантский сухопутный массив, покрытый густыми лесами и саваннами. Этот материк, по площади вдвое превосходивший Индийский субконтинент, принято называть Сундалендом (рис. 6.4)[337]. Однако с наступлением ледниковых плит внимание человека стали все больше привлекать не только обширные прибрежные пространства, освободившиеся от воды после отступления моря. Сухопутный мост, образовавшийся между Сибирью и Аляской, дал человечеству реальный шанс проникнуть на необозримые пространства Северной и Южной Америки. И люди не упустили этот шанс на последнем этапе ПЛМ. Это хорошо согласуется с точкой зрения о том, что миграции людей через Берингию (ныне — Берингов пролив) в Америку в тот холодный и засушливый период были вызваны резким ухудшением климатических условий в Азии. Центробежное расселение монголоидных охотников и собирателей во всех направлениях по великим рекам Азии не обязательно повлекло за собой быстрое вытеснение и замещение туземных жителей — потомков собирателей на прибрежной полосе, обитавших там ранее. Более чем вероятно, что там возникла широкая сеть селений и стоянок пришельцев, которая просуществовала достаточно долгое время, в результате чего генофонд новых мигрантов во многом перемешался с генофондом коренных жителей. По всему побережью Северного Китая, Кореи, Японии и Сундаленда возникли огромные прибрежные массивы суши, покрытые лесами умеренной полосы и субтропическими низменностями, на которых быстро освоились и начали расселяться мигранты из глубинных районов и степей Центральной Азии. Таким образом, первоначальные поселения мигрантов могли возникать как на месте древних заброшенных стоянок на пути собирателей на прибрежной полосе, так и новых землях, чтобы не вступать в конфликт с коренными жителями этих районов. Когда же по окончании Ледникового периода море, словно гигантская приливная волна, вернулось к своим прежним границам, подавляющее большинство материальных археологических свидетельств обитания человека в прибрежных районах было смыто волнами и ушло на дно морское. Поэтому мы можем надеяться найти археологические свидетельства Ледникового периода только в тех районах, где береговая линия достаточна крута, да на континентальном шельфе, например, в Японии и на некоторых островах Индонезии. Останки человека доледниковой эпохи можно обнаружить только в тех районах, которые оказались изолированными в силу географических причин или повышения уровня океана, например в Японии. Здесь мне хотелось бы особо отметить, что гипотеза о расселении монголоидов, которую я излагаю здесь, существенно отличается от официально принятой археологической реконструкции не только с точки зрения хронологии, но и с точки зрения направлений миграции древнего человека. Некоторые специалисты по доисторическому периоду развития Юго-Восточной Азии, такие, как австралиец Питер Беллвуд, высказываются в поддержку версии о недавнем вытеснении мигрантами-монголоидами древних охотников и собирателей австрало-меланезийского типа, обитавших на островах Юго-Восточной Азии, причем эти мигранты прибыли с острова Тайвань, находящегося на самой кромке Тихоокеанского побережья. Согласно этой гипотезе, древние охотники и собиратели австрало-меланезийского типа в этническом плане были наиболее близки к «негритосам» племени семанг (см. главу 5). Более того, Беллвуд утверждает, что одной из ветвей монголоидного населения следует считать рисоводов эпохи неолита из Южного Китая, которые прибыли сюда морским путем через о. Тайвань и Филиппинские острова около 4000 лет тому назад. Ранее мне уже приходилось выказывать аргументы против этой гипотезы[338]. Главное, о чем мне хотелось бы сказать в первую очередь, — это то, что первичная и вторичная экспансия монголоидов в Восточной и Юго-Восточной Азии началась значительно раньше, еще на пороге Последнего ледникового максимума, около 18 тысяч лет тому назад. Мигранты предпочитали по большей части сухопутные маршруты, которыми они пользовались не раз и не два. По сути, едва ли не самое крупное расселение человека по островам Юго-Восточной Азии, имевшее место еще до нашей эры, совпало по времени с началом века металлов на этих островах, что произошло как минимум за несколько веков до Р.Х., когда имело место широкое распространение культуры возделывания риса[339]. Второй момент, который мне хотелось бы отметить, сводится к тому, что речь идет не просто о двух группах — монголоидах, мигрировавших с севера, и охотниках-собирателях, этнически близких к племени семанг, которые заселили обширные пространства Юго-Восточной Азии. Кроме них, существовала и другая туземная группа, южные монголоиды, которые могли с древнейших времен населять острова и земли вдоль континентального шельфа Юго-Восточной Азии и большей части западного побережья Тихого океана и которых вполне можно считать истинными предками самих монголоидов. Эта туземная этническая группа южно-монголоидного типа лучше всего идентифицируется по характерным для нее южным генетическим маркерам, доминирующим в том регионе[340]. По внешнему виду они, скорее всего, были похожи на представителей некоторых национальных меньшинств, живущих сегодня на юго-западе Китая, и, в частности, на одну из групп аборигенов Малайзии, обитающих на юге Малайского полуострова и известных под названием малайских аборигенов. Последние напоминают большинство жителей Малай в том смысле, что тоже имеют ряд черт монголоидного типа. В этом отношении некоторые аспекты внешнего облика так называемых южных монголоидов могли сформироваться в гораздо более позднее время. Как мы увидим, предков этих людей можно идентифицировать отнюдь не только по генам и зубам. Малайские аборигены, как и громадное большинство жителей Юго-Восточной Азии, относятся к сундадонтному типу. С точки зрения строения зубов, малайские аборигены очень близки к жителям западного побережья Тихого океана, в том числе полинезийцам, и по морфологии зубов могут считаться предками синодонтов, расселившихся к северу (см. главу 5 и рис. 5.3)[341]. Короче говоря, южные монголоиды уже обитали на юге Юго-Восточной Азии задолго до Последнего ледникового максимума.
Проникновение выходцев из Азии на острова Юго-Восточной Азии и Океании в послеледниковую эпоху
Как я уже отмечал, предлагаемая мною реконструкция основана в большей мере на генетических данных, нежели на крайне редких находках останков скелетов и изучении строения зубов. Действительно, интерес к генетической истории доисторического периода впервые пробудился у меня еще в начале 1980-х гг., когда я работал на островах в юго-западной части акватории Тихого океана. Мои находки позволили установить, что очень ранние миграции локальных генетических типов из Юго-Восточной Азии в Новую Гвинею и на острова Меланезии представляются куда более вероятным сценарием развития событий, чем массовые миграции монголоидов из Китая, заместивших аборигенное население австрало-меланезийского типа путем переселения через Австралазию и Тихий океан, как считалось ранее[342]. И я приступил к сбору генетических свидетельств и данных о расселении монголоидов, проникших в Юго-Восточную Азию и Океанию. В главе 3 мы уже говорили о том, как оптимальное сочетание археологических, физических и генетических данных по раннему маршруту исхода из Африки было выявлено именно в конечных точках ветвей этого пути: Малайзии, Австралии и Новой Гвинее. Я познакомил читателей и с тем, насколько глубоки различия между ветвями генетических линий для Австралии и Новой Гвинеи, что свидетельствует о древности их заселения. Уникальная генетическая идентичность подобных ранних миграций позволяет нам выявить и идентифицировать всех позднейших пришельцев и установить, откуда именно они прибыли. Не менее ценна для нас и возможность датировки и оценки масштабов таких позднейших миграций. С помощью этого подхода известному английскому генетику Мартину Ричардсу и мне удалось доказать, что, хотя, согласно современным представлениям, предки полинезийцев — этих великих пионеров Тихоокеанского бассейна, избороздивших его около 3500 лет тому назад, — по большей части сильно отличались от древнейших туземных обитателей Новой Гвинеи и прилегающих островов, они прибыли туда отнюдь не с современного о. Тайвань, как считают большинство ученых, а непосредственно из Юго-Восточной Азии. Более того, хотя генетические линии полинезийцев и были пришельцами в Тихоокеанском регионе, их общие прапредки, по всей видимости, прибыли в Восточную Индонезию из Индокитая около 17 тысяч лет тому назад[343]. Линии, принесенные полинезийцами, были ветвями линий, обнаруженных в континентальных районах Юго-Восточной Азии, а одна древняя линия (принадлежащая к гаплогруппе В4 мтДНК) оказалась даже общей у них с мигрантами, направившимися в Америку по прибрежной полосе на крайнем западе Тихого океана Помимо генетической связи предков туземцев Америки с мигрантами в Юго-Восточную Азию, этот факт свидетельствует о том, что наиболее вероятным временем миграции на острова из континентальных районов Юго-Восточной Азии следует считать предледниковый период. Мы с Ричардсом продолжили изучение других генетических маркеров миграций в Юго-Восточной Азии, итогом которых явилось исследование коренных этнических групп Малайского полуострова, изложенное в этой книге. Ряд других генетиков также предложили свою датировку локальных скоплений азиатских генетических линий, как мужских, так и женских, которые некогда проникли из континентальных районов Восточной Азии на острова Юго-Восточной Азии и Тихоокеанского бассейна. Кембриджский генетик Питер Форстер попытался датировать время образования таких скоплений генетических линий, которые могут отражать местное увеличение численности населения на юго-западе Тихоокеанского бассейна. Одной из них стал локальный вариант восточно-азиатской гаплогруппы Е, который встречается сегодня у жителей Малайзии и Сабаха (северо-восточное Борнео). Форстер датировал возраст этого скопления линий в Азии (Е) 12 100 лет тому назад. Другим центром явился субклан крупнейшей в Юго-Восточной Азии гаплогруппы F, выявленный во Вьетнаме и Малайзии, который ученый датировал временем около 9100 лет тому назад. Форстер также установил, что один из вариантов гаплогруппы В4 прибыл в Новую Гвинею из Азии около 12 500 лет тому назад. Все эти результаты говорят в пользу моей гипотезы о том, что подобное расселение жителей в Восточной Азии и на островах юго-западной части бассейна Тихого океана носило постоянный характер, начиная с Последнего ледникового максимума, то есть задолго до начала революции эпохи неолита. Другими словами, экспансия типично восточно-азиатских линий наблюдалась в обширном регионе Юго-Восточной Азии и дальше к востоку, в Меланезии, задолго до того времени, как развитие земледелия могло дать импульс активной колонизации этого региона. Аналогичные датировки расселения в послеледниковую эпоху были получены при изучении проникновения Y-хромосом из Восточной Азии[344]. Эти данные сами по себе не могут показать уровень притока генов из Индокитая через острова Юго-Восточной Азии в Меланезию после окончания Ледникового периода. В 1994 г. итальянский генетик Антонио Торрони со своими коллегами совершил заметный прорыв в области идентификации семи материнских кланов Юго-Восточной Азии, которые получили название A-G. Исследователи установили, что все эти кланы присутствуют на Тибете и в меньшей мере — в других районах азиатского континента (более подробно об A-G см. главу 5). Вместе с недавно идентифицированной восточно-азиатской линией М7 эти семь генетических групп Восточной Азии были выявлены у 85% обследованных корейцев, 79% китайцев-ханьцев юга Китая, 75% вьетнамцев, 25% малайцев, 44% жителей Сабаха и 20% коренных жителей Новой Гвинеи[345]. Такое устойчивое снижение присутствия материнских линий Юго-Восточной Азии по мере продвижения от Восточной Азии к Новой Гвинее отражает постоянно увеличивающуюся долю местных туземных линий при удалении от континентальных районов Азии. В исследовании Торрони и его коллег те группы, в которых присутствовало менее всего восточноазиатских линий, поддающихся идентификации, были сконцентрированы в континентальных районах Юго-Восточной Азии. К ним относятся линии оранг асли, туземных жителей джунглей Малайского полуострова. Нам с Мартином Ричардсом и нашими малайскими коллегами недавно удалось получить подтверждение этой картины распространения линий в гораздо более обстоятельном исследовании оранг асли (см. также главу 5). Двумя ключевыми аборигенными группами, менее всего затронутыми последующими волнами иммиграции, были народность семанг и малайские аборигены. В генетическом фонде первых выявлено всего 22% типичных восточноазиатских генетических линий. Это хорошо согласуется с точкой зрения о том, что негритосы народности семанг представляют собой весьма изолированную реликтовую этническую общность. В то же время столь же низкий уровень континентальных линий у малайских аборигенов, принадлежащих к южным монголоидам, можно считать неожиданным. Лишь у четверти из них были выявлены типичные восточноазиатские генетические линии, тогда как у половины преобладали две основные азиатские ветви. Одна из них (включавшая в себя четверть типичных восточноазиатских линий) возникла чуть ранее, но на той же самой ветви, что и евразийская генетическая подгруппа Насрин. Другими словами, она была двоюродной сестрой Насрин, одной из двух дочерей Евы выходцев из Африки. Эта древняя наследственная линия пре-Насрин ранее никогда не встречалась. Другим неожиданным фактором явилось то, что последняя четверть линий принадлежала к ветви npe-F. А поскольку F считается крупнейшей линией-родоначальницей для всей Восточной и Юго-Восточной Азии, ее находка — аргумент в пользу высказанной в главе 5 версии о том, что такую группу южных монголоидов можно считать представителями географической прародины монголоидов[346]. Возвращаясь к эпохе Последнего ледникового максимума, следует отметить, что эти неожиданные находки в области генетики хорошо согласуются с современными представлениями о том, что изолированные этнические группы малайских аборигенов и народности семанг обладают наименьшей долей притока генов извне и, следовательно, наиболее близки к типам коренного населения Малайского полуострова, обитавшего здесь в глубокой древности до массового переселения монголоидов с севера. Теперь нам остается рассмотреть вопрос о том, какова доля материнских линий северных монголоидов у представителей коренного населения Юго-Восточной Азии. Демонстрируя тенденцию к более высокому уровню проникновения мужских линий через межнациональные границы, в Индокитае и на островах Юго-Восточной Азии доминируют типичные континентальные Y-хромосомные линии, характерные для Восточной Азии и определяемые единым генетическим маркером Хо (см. главу 5). Доля этих линий в разных районах данного региона колеблется от 54 до 97%, причем на линии Уоллеса отмечается резкий спад (линия Уоллеса служит юго-восточной границей континента Сунда, отделяя его и всю остальную континентальную Азию от островов Восточной Индонезии, Новой Гвинеи и Австралии — см. рис. 5.6). Однако, так как линия Хо, по всей вероятности, возникла в Юго-Восточной Азии, нет оснований полагать, что подобное доминирование отражает собственно проникновение северных монголоидов, а не простое расселение жителей по Юго-Восточной Азии в эпоху после Последнего ледникового максимума[347]. Таким образом, имеющиеся генетические свидетельства поддерживают версию о том, что расселение восточноазиатских линий, которое было направлено на юг, в Юго-Восточную Азию, началось около 18 тысяч лет тому назад (в эпоху Последнего ледникового максимума) и продолжается вплоть до сегодняшнего дня. Объяснение всех сложных особенностей смешения монголоидных и немонголоидных черт внешнего облика у доминирующих в наши дни представителей южно-монголоидного типа в Юго-Восточной Азии вплоть до линии Уоллеса займет слишком много времени (см. рис. 5.6). Негритосы немонголоидного типа с характерным строением зубов, не поддающимся идентификации (народность семанг — см. главу 5), обитающие в джунглях Центральной Малайзии, представляют собой изолированную, относительно мало ассимилированную реликтовую этническую общность, происходящую от древнейших собирателей на прибрежной полосе, обосновавшуюся в этих местах задолго до появления монголоидов. После прихода мигрантов с севера народность семанг сохранилась в глухих джунглях в качестве этаких генетических островков. Изучение малайских аборигенов, обитающих в джунглях Малайского полуострова, по всей вероятности, может дать важные генетические и дентологические ключи к выявлению наследственных преемственных связей между сундадонтными типами Юго-Восточной Азии и синодонтным населением более северных районов.
Археологические свидетельства древнейшего расселения монголоидов в Юго-Восточной Азии
В числе наиболее значительных археологических находок последнего времени следует выделить открытие останков скелетов людей монголоидного типа в Сонг Кеплек, пещере в Гунунг Севу на о. Ява, датируемых около 7000 лет тому назад. Останки эти восходят к эпохе задолго до начала возделывания риса в Индонезии, а это уже ставит под сомнение базовую гипотезу о том, что вторжение монголоидов на острова Юго-Восточной Азии совпало по времени с активным распространением культуры риса. Начиная с Последнего ледникового максимума и вплоть до 10 тысяч лет тому назад для монголоидов не существовало морского барьера до того момента, как они достигли линии Уоллеса в районе острова Бали. Когда же они, проделав посуху долгий путь из Азии, достигли берегов современных островов Бали и Борнео, то увидели перед собой море, которое им предстояло преодолеть. Как мы знаем благодаря исследованиям ранних аборигенов Австралии, море не являлось для человека непреодолимой преградой, однако существенно замедлило приток мигрантов и новых генов. Соответственно, Восточная Индонезия, лежащая по другую сторону линии Уоллеса, представляет собой своего рода разграничительную линию, по одну сторону которой живут представители негритосов, а по другую — монголоиды. Если же направиться в Новую Гвинею, лежащую по другую сторону пограничной зоны, образуемой Молуккскими и прочими мелкими островами Восточной Индонезии, нетрудно заметить, что для подавляющего большинства местных жителей характерны типично меланезийские курчавые волосы и темная кожа. Жители Новой Гвинеи по целому ряду антропологических параметров близки к негритосам Малайского полуострова, живущим в нескольких тысячах километров к западу отсюда[348]. Несмотря на нарисованную мной колоритную картину, до сих пор бытует мнение, что в Юго-Восточной Азии вплоть до недавнего времени обитали по большей части охотники и собиратели австрало-меланезийского типа. При практически полном отсутствии на обширных массивах суши останков человека, которые можно было бы отнести к эпохе подъема уровня океана в послеледниковый период, это мнение получило достаточно широкое распространение. Однако факты, изложенные здесь, противоречат гипотезе об австрало-меланезийском этническом составе. На сегодняшний день наиболее древними документально зафиксированными останками человека анатомически современного типа, найденными в Юго-Восточной Азии, является знаменитый «глубокий череп» из пещер Ниах на о. Борнео. Этот череп, возраст которого, согласно радиоуглеродной датировке, составляет 42 тысячи лет, прежде относили к «австрало-меланезийскому тонкокостному типу», точнее — к вымершим сегодня аборигенам Тасмании. Однако, как подчеркивает видный австралийский антрополог и археолог Дэвид Балбек, относительно тонкокостные черепа тасманцев нельзя считать типично австралоидными или меланезийским, и найденный в пещере Ниах череп, как и черепа аборигенов Тасмании, скорее близок к айнам — коренным жителям Японских островов. К такому же выводу Балбек пришел и в результате изучения другого черепа, найденного в пещере Табон на Филиппинских островах. Его возраст — около 20 тысяч лет[349]. Два других знаменитых черепа, найденных в конце XIX в на стоянке Ваджак на о. Ява, вызвали большой интерес и послужили основой для целого ряда попыток воссоздать историю жизни человека в этих местах в доисторическую эпоху. На протяжении более чем ста лет эти черепа считались относящимися к эпохе до Последнего ледникового максимума и, следовательно, ранними образцами протоавстралоидного типа. Однако, вопреки концепции об австраломеланезийском типе этих останков, целый ряд антропологов высказывают предположение, что важдакские черепа не имеют ничего общего с останками австралоидов аналогичной древности. Вместо прежней гипотезы было высказано мнение, что эти черепа относятся к ранним южным монголоидам или даже идентичным, но более толстокостным, чем черепа современных аборигенов Явы. Дэвид Балбек высказывает и еще более убедительное предположение о том, что эти останки также весьма похожи на черепа айнов Японии и представляют собой останки жителей обширного континуума, в состав которого входили земли вдоль побережья Тихого океана и Сундаленд. И хотя прежние оценки возраста ваджакских черепов настаивали на том, что они относятся к эпохе плейстоцена, то есть они старше (или даже гораздо старше) 10 тысяч лет, две последние датировки их по радиоуглеродному методу показывают, что их возраст составляет 10 560 или 6560 лет, что позволяет отнести их к началу голоценового периода, времени, предшествующему развитию земледелия[350]. Естественно, сама возможность того, что черепа из пещер Ниах и Табон, а также со стоянки Ваджак могут считаться свидетельствами раннего присутствия первых монголоидов или премонголоидных типов на островах Юго-Восточной Азии, делает актуальным вопрос о времени их появления там. Вполне вероятно, что какую-то общую дату установить просто невозможно, если миграции южных монголоидов в этот регион носили характер длительной эволюции. А это может стать вызовом общепринятой точке зрения на позднейшее появление монголоидов в регионе. И хотя это еще не свидетельствует о том, что монголоидные черты внешнего облика появились у жителей островов Юго-Восточной Азии еще во времена Последнего ледникового максимума, подобные находки дают основания полагать, что монголоиды могли поселиться на о. Ява задолго до неолита, который обычно считается общепризнанной датой их появления в этом регионе. Таким образом, и археологические, и генетические свидетельства говорят о том, что малайские аборигены вполне могли быть потомками древнейших южных монголоидов, или протомалайцев, как звучит их научное название[351].
Монголоиды как замещающий этнос в Китае?
А теперь давайте обратим взгляд на север. Аналогичная картина локального генетического расселения ветвей мтДНК в послеледниковую эпоху наблюдается и вдоль побережья Тихого океана от Южного Китая до Северной Азии. Она отражает поток мигрантов на восток, хлынувший с равнин Центральной Азии, и их последующее расселение по всему Тихоокеанскому побережью. Среди этих генетических миграций наиболее близкой по времени к Последнему ледниковому максимуму является миграция части общеазиатской гаплогруппы D, возраст которой — около 16 800 лет. Время появления одной из характерных линий подгруппы D, свойственной чукчам — обитателям крайнего северо-востока Сибири, можно датировать 14 900 лет. В числе других ветвей — типы А2 (11 200 лет) и С (10 800 лет)[352]. В Китае и на побережье Тихого океана мы вновь встречаем реликтовые туземные группы, поселившиеся здесь еще задолго до прихода монголоидов и изолированные от внешнего мира не только джунглями, но и морем. Одна из таких групп — уже знакомые нам айны, издревле живущие на севере Японии. Айны являются прямыми потомками йомонов — коренных жителей Японских островов, которые около 12 500 лет создали одни из первых в мире гончарные изделия. Позднейшая иммиграция с континента народа йайои из Кореи привела к замещению коренного населения, и поэтому у современных айнов отмечаются больший или меньший уровень примеси инородной крови. Между доисторическими черепами йомонов, японским черепом Минатогава 1 времени Ледникового периода с острова Окинава (датируемым временем между 16 600 и 18 250 лет тому назад) и черепами современных айнов очень много общих, близкородственных черт, позволяющих отнести их к одной и той же наследственной линии. Как отмечалось выше, эти группы, обитавшие на западном побережье Тихого океана, могут составлять остатки домонголоидного этнического субстрата[353]. В результате изоляции Японии последние иммигранты-монголоиды с материка, народность йайои, прибыли на Японские острова около 2300 лет тому назад. Это событие тоже оставило генетический след в истории Японии. Исследования, проводившиеся в разных районах Японских островов, показали, что для йомонов был характерен маркер YAP+, или Авель, одной из древнейших линий собирателей на прибрежной полосе — выходцев из Африки. Так, на о. Окинава этот редкий для Азии маркер YAP+ имел встречаемость порядка 55%. Другой архаический Y-маркер собирателей, Каин, также выявлен у японцев, но встречаемость его гораздо ниже — 10%[354]. Череп Минатогава 1 имеет целый ряд общих черт с двумя другими знаменитыми древнейшими черепами из Китая: черепом из Верхней пещеры 101 и люйцзяньским черепом. Первый бы найден в Чжоукудянь (Холм Драконьих Костей), в Северном Китае, и хотя он с трудом поддается датировке, его возраст, несомненно, превышает 10 тысяч лет, и вполне возможно, что он относится к доледниковому периоду. Люйцзяньский череп был найден в Южном Китае (см. главу 4). Австралийский палеоантрополог Питер Браун выступает против мнения о том, что эти три толстокостных черепа имеют протомонголоидные черты. Согласно его схеме, они скорее ближе к черепам аборигенов Австралии, чем современных жителей Восточной Азии, хотя и с первыми их сходство не слишком заметно[355]. Эти факты вполне согласуются с гипотезой о том, что в доледниковую эпоху население Китая и Тихоокеанского побережья Восточной Азии до прихода монголоидов могло в значительной степени состоять из потомков первых собирателей на прибрежной полосе, генетическое наследие которых еще вполне могло сохраниться в некоторых районах. Другой архаический Y-маркер собирателей на прибрежной полосе, Каин (см. главу 5), также выявлен в Северо-Восточной Азии, на этот раз — в континентальных районах и на берегах Амура, в районе Охотска, в Монголии и Центральной и Южной Сибири. В этих районах Каин, обычно представленный весьма незначительно, имеет встречаемость порядка 85—90% [356]. Тот факт, что этот древнейший североазиатский маркер собирателей на прибрежной полосе присутствует в этих местах, вполне согласуется с данными исследования черепов, свидетельствующими, что в этом регионе древнее немонголоидное население сохранялось как минимум до Последнего ледникового максимума.
Культурно-археологические следы расселения в Азии
Существуют ли какие-нибудь археологические материалы, помимо ваджакских черепов, свидетельствующие о присутствии в эпоху палеолита в Северной и Восточной Азии древнейших сообществ, которые впоследствии были ассимилированы или замещены в результате расселения монголоидов из степей Центральной Азии около 20 тысяч лет тому назад? Да, существуют, но к ним следует относиться с осторожностью. Интерпретацию археологических материалов, найденных в Северо-Восточной Азии, Китае и Корее (а также в Юго-Восточной Азии) и относящихся ко времени между 5500 и 25 тысячами лет тому назад, весьма затрудняет отсутствие достоверных археологических находок на прибрежной полосе. Это объясняется тем, что современный высокий уровень океана привел к затоплению громадных прибрежных территорий континентального шельфа, на которых в древности селились переселенцы (см. рис. 6.3). Все, что нам осталось, — это несколько материковых и островных пещерных стоянок, которые вполне могли принадлежать скорее беженцам из числа коренного населения, чем агрессорам-мигрантам. А теперь мне хотелось бы изложить весьма упрощенную трактовку археологических находок в Восточной Азии, относящихся к Ледниковому периоду. Эта трактовка способна послужить своего рода фоном для анализа изменений, начавшихся с эпохи Последнего ледникового максимума. Как мы уже знаем (см. главу 3), в Европе происходил процесс замены архаической техники орудий Среднего палеолита, созданной неандертальцами, на более сложные орудия Верхнего палеолита, созданные первыми людьми анатомически современного типа. Мы видели также, что техника эпохи Среднего палеолита, разработанная неандертальцами, ни в коей мере не была примитивной, а представляла собой как бы параллельную традицию стилей создания орудий Среднего Каменного века человеком анатомически современного типа в Африке около 50 тысяч лет тому назад. Таким образом, можно считать странным и даже удивительным низкий технический уровень каменных орудий, использовавшихся людьми современного типа в Восточной и Юго-Восточной Азии в эпоху решающей борьбы неандертальцев с кроманьонцами. Каменные орудия, созданные человеком анатомически современного типа в Юго-Восточной Азии и Австралии, выглядят куда более примитивными, чем орудия, которыми пользовались неандертальцы[357]. Контраст между ними настолько велик, что в некоторых случаях археологи, привыкшие иметь дело с африканской и европейской палеолитическими техниками, просто не могут поверить, что грубые куски камня, которые их азиатские коллеги уверенно атрибутируют людям современного типа, вообще являются артефактами и делом рук человеческих. Первым этот разительный контраст в стилях и «качестве» орудий заметил в 1948 г. американский археолог Холлэм Мовиус. Он обнаружил как бы линию, получившую впоследствии имя линии Мовиуса, которая отделяет весь Дальний Восток, включая Юго-Восточную Азию, от остального мира, так что Восток предстает зоной культурнотехнической «отсталости» и создания «устаревших» орудий вплоть до последнего Ледникового периода. В довершение путаницы эпоха существования восточноазиатской палеолитической техники создания каменных орудий, так называемых ножей и скребков, была отодвинута во времени от людей современного типа до Среднего палеолита, то есть как минимум на миллион лет назад. В те времена, задолго до появления человека современного типа, в Восточной Азии жили представители вида Homo erectus. Существуют разные мнения в вопросе о том, когда именно на островах Юго-Восточной Азии впервые появились ножи, скребки и рубящие орудия, и некоторые археологи не без оснований утверждают, что они были созданы человеком современного типа примерно около 70 тысяч лет тому назад[358]. Некоторые археологи указывают, что низкий уровень сложности рубящих и режущих орудий той эпохи является отражением плохого качества камня, из которого они делались, в частности кварцита или базальта. Было высказано мнение, что люди в те времена (как, впрочем, и сейчас) основную массу орудий делали из недолговечных материалов: твердых пород дерева, бамбука и растительных волокон[359]. Тот факт, что жители островов Океании, построив лодки, сумели колонизовать и захватить Северные Соломоновы острова еще 30 тысяч лет тому назад, за несколько десятков тысяч лет до начала кораблестроительства и мореплавания в Европе, можно считать неопровержимым доказательством — если таковые вообще нужны — высокого технического уровня жителей Дальнего Востока.
Легенда двух рек
Каковы бы ни были причины существования примитивных техник создания рубящих и режущих орудий на Дальнем Востоке, они являются неким материальным фоном, позволяющим судить о проникновении носителей других культур в Китай из Центральной Азии в эпоху Последнего ледникового максимума. В центре этого фона находится географическая и хронологическая структура, которую мы можем назвать «легендой двух рек». Палеолитические культуры Северного Китая географически и культурологически были сконцентрированы вокруг Желтой реки (Хуанхэ) и задолго до Последнего ледникового максимума испытали на себе сильное влияние инновационных техник Верхнего палеолита, проникавших в этот регион из Центральной Азии. Первые орудия-отщепы, затем ножи и, наконец, характерные обоюдоострые наконечники, эта визитная карточка диуктайских культур Восточной Сибири в эпоху Последнего ледникового максимума, были впоследствии занесены в куда более отдаленные районы Азии, вплоть до Японии, где они начали активно вытеснять более архаичные виды оружия и бытовых орудий. С другой стороны, Южный Китай, культуры которого были сосредоточены вокруг Янцзы и защищены как в географическом, так и в культурно-историческом отношении от проникновения новаторских влияний из степей Центральной Азии, развивались во многом иначе, и их эволюция носила более медленный характер[360]. Другим потенциальным фактором влияния западных культур на Китай и Юго-Восточную Азию в эпоху Последнего ледникового максимума, который мог развиваться параллельно с сибирскими культурами Верхнего палеолита, было производство орудий из дробленого камня в бассейне реки Тарим у южных окраин пустыни Такламакан в провинции Синьцзян, а также производство микролитических орудий на Цзиньхай-Тибетском плато (микролиты — это небольшие призматические отщепы с параллельными кромками, делавшиеся из заранее обработанной заготовки)[361]. Некоторые аспекты западного технического влияния в эпоху Верхнего палеолита появились в Северном Китае и южных районах Внутренней Монголии задолго до наступления Ледникового периода. Пожалуй, наиболее ранние следы таких влияний, относящиеся к эпохе между 35 и 50 тысячами лет тому назад, были обнаружены у излучины в среднем течении Хуанхэ, известной как плато Ордос. В знаменитой Верхней пещере, находящейся неподалеку от Пекина, были найдены артефакты из кости, раковин и камня, относящиеся к Последнему ледниковому максимуму, в том числе — иголка с ушком и сложное и изящное ожерелье из кости, раковин и цветного камня, датируемые 18 тысячами лет тому назад. Еще более сложные культуры Верхнего палеолита, мастера которых создавали, в частности, иглы с тремя ушками и сложные орудия из кости и рога, продолжали развиваться и после Последнего ледникового максимума на территориях в нижнем течении Хуанхэ вплоть до самого устья[362]. Но большая часть восточного побережья Китая, которая была наиболее населенным районом в эпоху ПЛМ, сегодня находится глубоко под водой и совершенно недоступна для археологов. Еще далее на северо-восток, на Дальнем Востоке России, в Японии и Корее, наблюдаются еще более радикальные изменения в техническом уровне орудий, созданных в эпоху ПЛМ. Так, на Дальнем Востоке России наиболее ранней стоянкой, где были найдены самые древние, согласно датировке по радиоуглеродному методу, макролезвия, является так называемая пещера Географического общества на берегу реки Амур. (Кстати, о макролезвиях. Любое каменное орудие длиной более 50 мм классифицируется как макролезвие.) Макролезвия появились в этом регионе также в эпоху Последнего ледникового максимума (около 19 350 лет назад), но — ближе к побережью Японского моря, в Приморье (Россия)[363]. В Японии получены археологические данные, позволяющие реконструировать самую подробную во всем мире хронологию эпохи Последнего ледникового периода. В период до 20 тысяч лет тому назад в Японии преобладала общая для всей Восточной Азии техника создания режущих и рубящих орудий. Между 18 и 20 тысячами лет назад наступил новый этап развития техники создания каменных орудий, во многом сходный с техниками Верхнего палеолита на Западе, появились орудия из дробленого камня и лезвия (последние получили широкое распространение в ту эпоху) и целый ряд других типов орудий Наконец, между 12 и 14 тысячами лет тому назад появились микролиты и наконечники с двойными кромками. Что касается Кореи, то там проведено значительно меньше исследований, хотя, в общем, наблюдается та же тенденция: сперва появились орудия из цельного камня, затем — из дробленого (отщепы и сколы) и, наконец, в эпоху около ПЛМ — настоящие лезвия. И в том и в другом случае мы видим, что эпоха Последнего ледникового максимума была временем активных технико-технологических изменений, когда устаревшие традиционные восточноазиатские техники уходили в прошлое, по всей видимости — в результате появления новых навыков и притока беженцев-охотников с запада, из ледяных степей Центральной Азии[364]. В то же время районы по берегам Янцзы в Южном Китае, по всей видимости, были в значительной мере защищены от проникновения с запада технических инноваций эпохи Верхнего палеолита, однако технологии доледникового периода, базировавшиеся на использовании кварцита для производства режущих и рубящих орудий, претерпели существенные изменения (хотя и не исчезли совсем) в эпоху Последнего ледникового максимума благодаря появлению новых орудий и материалов. Эти новые орудия были значительно мельче и отличались широким разнообразием, включая отщепы, и сделаны они были из самых разных материалов, в том числе сланца, кости и рога. Как и на севере, прибрежные земли Южного Китая, густо заселенные в эпоху Последнего ледникового максимума, в наши дни находятся глубоко под водой, навсегда скрыв от исследователей реальные археологические свидетельства технического прогресса во времена ПЛМ[365].
Юго-Восточная Азия: пауза в заселении или всего лишь последствие повышения уровня океана?
В отличие от тех важных технико-технологических перемен, которые имели место в эпоху Последнего ледникового максимума в северных районах Азии, в Юго-Восточной Азии сохранилось очень мало свидетельств, относящихся к апогею Ледникового периода. Действительно, до нас дошло крайне мало следов материальной культуры того времени. И независимо от того, отражает ли эта лакуна реальное отсутствие населения или объясняется тем, что следы древних обитателей ищут не там, где следует, сам факт их отсутствия не может быть использован в качестве аргумента в пользу идеи об активной иммиграции с севера. Дело в том, что маскирующий эффект низкого уровня океана в эпоху ПЛМ нигде не был выражен столь явно, как в Юго-Восточной Азии, где современные участки суши, возвышающиеся над водой, представляют собой лишь жалкие остатки некогда огромного континента Сундаленд (см. рис. 6.4). Уже упоминавшийся нами Дэвид Балбек указывает, что одна из главных причин того, почему до нас дошло так мало материальных свидетельств деятельности человека в эпоху Последнего ледникового максимума, может заключаться в том, что люди в те времена жили на землях, лежавших на высоте гораздо менее 100 метров над уровнем океана. Древние люди просто-напросто следовали за морем, отступавшим от берегов, обнажая обширные пространства суши, которые сегодня вновь ушли глубоко под воду. Когда же уровень океана в послеледниковый период начал быстро повышаться, люди были вынуждены отойти подальше от берегов. Именно с этого времени началось создание новой материальной культуры, артефакты которой и находят археологи[366]. Итак, какова бы ни была истинная причина, налицо явная пуаза в заселении глубинных материковых земель Индокитая и Малайского полуострова, продолжавшаяся от времени Последнего ледникового максимума до 14 400 лет тому назад, когда пещеры в глубинных районах начали вновь активно заселяться людьми, которые создавали орудия из дробленого камня, а также режущие и рубящие орудия. В некоторых случаях архаические техники обработки камня сохранялись вплоть до эпохи неолита, около 3000 или даже 2000 лет тому назад. Поскольку все эти районы, несомненно, были обитаемы в эпоху Последнего ледникового максимума, вполне вероятно, что их жители по мере повышения уровняокеана просто перебирались на новые земли, подальше от берега[367]. Весьма интересен и вопрос о том, кто были эти обитатели пещер, создававшие орудия из дробленого камня. Принято считать, что они были предками кочевых племен негритосов, кормившихся охотой и собирательством в лесах, тех самых племен, которые и сегодня живут на тех же землях. Археолог Зурайна Маджид утверждает, что послеледниковая техника обработки камня, возникшая в долине Ленггонг в Малайзии, была на самом деле развитием традиции, которая сложилась в той же долине на стоянке Кота Тампан, возраст которой датируется временем около 74 тысяч лет тому назад (см. главу 4). Существует ряд генетических свидетельств, показывающих, что предки малайских аборигенов могли жить в ту же эпоху в прибрежных районах Индокитая. В наши дни эти районы также находятся глубоко под водой[368]. На островах Юго-Восточной Азии, где архаические техники создания орудий и лезвий из дробленого камня просуществовали вплоть до 2000 лет тому назад, панорама изменений в технологии в эпоху ПЛМ выглядит несколько более выразительной. Находки, сделанные здесь, служат ключами к истории расселения жителей на островах ЮгоВосточной Азии во время Ледникового периода, как о том свидетельствуют первые следы колонизации Филиппинских островов еще 17 тысяч лет тому назад. Американский археолог Вильгельм Сольхейм, патриарх в области изучения доисторического этапа развития Юго-Восточной Азии, считает, что техника создания орудий из дробленого камня и отщепов была занесена в эпоху позднего плейстоцена на острова Юго-Восточной Азии из Южного Китая, а не явилась результатом местной эволюции[369]. Пожалуй, единственной «звездочкой», свидетельствующей о проникновении действительно новаторской технологии, можно сказать, с другой планеты, является стоянка в восточном Сабахе, на берегах высохшего озера Тингкайю. Там были найдены изящные обоюдоострые ланцетообразные сланцевые ножи, датируемые периодом между 18 и 28 тысячами лет тому назад. (Термин «ланцетообразные» означает «имеющие форму листа или наконечника копья».) Вторая из этих дат говорит о том, что эта технология могла появиться там скорее до Последнего ледникового максимума, нежели после него. По словам Питера Беллвуда, эти ножи «являются уникальными для всей Юго-Восточной Азии, если не считать одного-единственного ланцетообразного ножа, найденного... в оловянном руднике в Кедахе на Малайском полуострове». Если ножи тингкайюского типа на о. Борнео представляют собой артефакты, занесенные извне, то замечание Беллвуда может стать ключом к маршруту, по которому эта техника проникла сюда из Китая, так как он, кроме того, упоминает еще о двух стоянках доледникового периода, найденных на том же полуострове, но — в тайской его части. Эти стоянки — Мо Хиеу и Ланг Лонгриен. На каждой из этих стоянок найдены следы технологии создания обоюдоострых орудий, но они тонут в массе более «грубых» изделий хоабинийского типа[370]. Более того, Беллвуд отыскал технические «отзвуки» ланцетообразных наконечников тингкайюского типа в Северо-Восточной Азии. Это — орудия, найденные, в частности, в районе Диуктая на северо-востоке Сибири (датируемые около 18 тысяч лет тому назад) и в нескольких районах Японии, также относящиеся к тому самому периоду[371]. И хотя Беллвуд склонен полагать, что культура Тингкайю была изолированным локальным явлением, ее техникокультурные отзвуки, о которых он упоминает, хорошо согласуются с данными генетической модели, предлагаемой мною. Другими словами, расселение охотников и собирателей из степей Центральной Азии в эпоху Последнего ледникового максимума повлекло за собой распространение передовых охотничьих орудий на восток и юго-восток, в Китай и Сундаленд, где существовали травянистые степи с умеренным климатом. Любопытно, что другой технический аналог орудий диуктайского типа найден в Новом Свете, которому посвящена следующая глава и который стал крайней точкой на долгом пути человека к самым отдаленным уголкам земного шара. Чрезвычайно специфический, региональный характер генетических древ материнских и отцовских линий, существующих в наши дни, позволяет проследить маршруты древних миграций. Это говорит о том, что, когда люди заселили весь Старый Свет и обосновались на приглянувшихся территориях, они на протяжении как минимум пяти столетий с успехом отражали натиск мигрантов из других краев. Конец подобному консерватизму положило наступление Последнего ледникового максимума. В Северном полушарии громадные территории Старого и Нового Света стали необитаемыми и были покинуты человеком из-за появления там льдов, ледниковых озер и приполярных пустынь. И перед охотниками и собирателями, еще недавно процветавшими на просторах степей Центральной Евразии, возникла проблема выбора путей выживания, причем выбор этот был очень невелик и, как всегда, продиктован географическими и климатическими факторами. В полуостровной Европе, изолированной от внешнего мира морями, горами и пустынями, единственный шанс выжить для беженцев заключался в том, чтобы попытаться найти прибежище в более южных и теплых климатических районах, граничащих со Средиземным и Черным морями. После ПЛМ беженцы вернулись, их численность быстро увеличилась, и они начали заселять обширные территории, по большей части — те же, с которых некогда ушли их предки. В Центральной и Северной Азии, некогда покрытой необъятными травянистыми степями, где паслись огромные стада травоядных, резкое похолодание и наступление сильной засухи вынудило охотников и собирателей Верхнего палеолита покинуть степи на горных плато и двинуться во все стороны света в поисках более теплых и пригодных для жизни краев. К числу таких мест, по всей вероятности, относились Украина на западе, Китай на юге и востоке, а также Япония, Корея и Северо-Восточная Сибирь. Как всегда, великие реки Азии играли в древности роль транспортных магистралей, но на этот раз направление миграции было обратным — вниз по течению. Археологические свидетельства подобной миграции культур охотников Верхнего палеолита к побережью Тихого океана в эпоху Последнего ледникового максимума особенно многочисленны в Японии, но они встречаются практически повсюду. В результате понижения уровня Мирового океана громадные территории континентального шельфа в Южной и Юго-Восточной Азии стали доступными для обитания людей. Вопрос о том, какую долю в заселении Сундаленда (крупнейшего сухопутного массива Юго-Восточной Азии) составляли местные жители и какую — мигранты-беженцы с далекого севера, не вполне ясен, однако генетические и дентологические данные говорят в пользу преобладания первых. Понижение уровня океана привело к появлению на карте мира нового континента, так называемой Берингии, возникшей на крайнем северо-востоке Азии. Этому континенту предстояло стать сухопутным мостом в Америку, где и разворачивается действие последней главы нашей истории.
ГЛАВА СЕДЬМАЯ ЗАСЕЛЕНИЕ АМЕРИКИ
Тот факт, что американские индейцы, перебравшись по некогда существовавшему континенту Берингия из Азии в Америку, является естественным объяснением сходства, которым эти народы, очевидно, обязаны наличию у них общего предка. Как писал в 1784 г. Томас Джефферсон, «между индейцами Америки и жителями Восточной Азии наблюдается поразительное сходство, которое наводит нас на мысль, что либо первые являются потомками последних, либо последние — первых...» Надо отметить, что еще за 200 лет до Джефферсона эту справедливую, хотя несколько двусмысленную идею высказал иезуит Хосе де Акоста, известный ученый и путешественник. Именно он выдвинул гипотезу, что жители Азии приплыли в Америку за 2000 лет до испанцев. Еще недавно у этой гипотезы не было соперников[372]. За исключением колонизации Полинезии, Америка, по всей вероятности, была тем континентом, на неисследованные земли которого был направлен вектор последнего переселения народов. Если учесть, что за последние годы наука и техника сделали значительный шаг вперед, позволив исследователям заглянуть в доисторическое прошлое, и что у американцев, считающихся самой богатой в мире нацией, располагающей к тому же неслыханными ресурсами, собственное прошлое вызывает столь жгучий интерес, можно предположить, что не за горами то время, когда в изучении истории заселения Америки не останется белых пятен. Нам хотелось бы знать, когда примерно первопроходцы вторглись на земли Америки и сколько было волн миграции, что это были за народы и откуда, как и когда они переселились на эти земли. Мы надеемся получить своего рода лингвистический «ключ» к этой тайне и выяснить, на каком языке они говорили. Но, несмотря на все достижения в области науки и техники, мы, кажется, ничуть не приблизились к пониманию доисторической действительности. Ученые не могут прийти к единому мнению даже относительно того, когда именно совершилось первое вторжение переселенцев на земли Америки. По разным оценкам, это произошло от 11,5 до 50 тысяч лет тому назад. Остается неразрешенным и другой вопрос: сколько волн миграции было. Некоторые ученые считают — одна, другие — несколько. Неясно, к скольким языковым группам принадлежали говоры или диалекты коренных жителей Америки, не говоря уже о том, представители скольких языковых групп впоследствии вторглись в Америку. Многие проблемы и противоречия коренятся в стремлении ответить на поставленные вопросы, опираясь на кажущуюся самоочевидность некоторых гипотез, ставших предрассудками. В результате академики высказывают противоположные мнения и не могут прийти к разумному согласию. Тем временем обозреватели, специализирующиеся на доисторическом прошлом Америки, любят подогревать читательский интерес к прошлому сенсационными сообщениями, основанными на фактах и выводах «свободных предпринимателей» от науки, но подробнее об этом мы поговорим позже[373]. На основе формального научного метода можно выдвинуть лишь расплывчатые теории, неопределенность и способность теории видоизменяться является гарантом ее строгой научности. Грубо говоря, задача теории (или «модели») состоит в логическом обосновании возможности тех путей доисторического развития, которые кажутся отнюдь не самоочевидными и нуждаются в научном подтверждении.
Базовая концепция «Первый Кловис»
Какое же отношение имеет все вышесказанное к заселению Америки? Отвечу: громадное. Изо всех учений, сложившихся в американской археологии, самым вздорным является так называемая ортодоксальная базовая концепция «Первый Кловис», или теория, основанная на находках в первом культурном слое Кловиса. Историю этого ортодоксального учения можно проследить с конца XIX в., ранее же оно воспринималось как ересь. В конце 1890-х гг. Вильям Генри Холмс с кафедры американской этнологии Смитсоновского института и Томас Чемберлен из Комитета геологии Соединенных Штатов начали травлю на смутные и еще не вполне сложившиеся представления о заселении Нового Света в плейстоцене (ледниковом периоде). В 1920-х гг. роль защитника этой теории перешла к антропологу Алесу Хрдличке, авторитетному ученому из того же Смитсоновского института. Значительно позже, в 1995 г., американский писатель Вайн Делориа в своих книгах «Красная земля, белые ландшафты» указал на Хрдличку как на рьяного деспотичного защитника академического status quo, всячески препятствовавшего реализации исследовательских программ, которые ставили перед собой задачу оценить альтернативные теории и сопоставить их с реальными фактами[374]. В 1926 г. Джесси Хиггинс из Музея естественной истории, штат Колорадо, во время раскопок в окрестностях Фолсома, штат Нью-Мексико, нашел остроконечный окаменевший артефакт, сделанный из скелетной кости вымершего бизона. Поскольку археологи не пришли к единому мнению относительно того, действительно ли окаменелость сделана из кости, Хрдличка до тех пор отказывался признать эту находку в качестве свидетельства присутствия человека в этих местах в плейстоцене, пока наконечник не был доставлен с места раскопок. Следующий наконечник был найден в 1927 г. Независимые эксперты подтвердили подлинность найденного артефакта, его внимательно изучили, сфотографировали, а затем решили оставить на исконном месте. Хиггинс нашел наконечники, которые были больше и весомее прежних, причем насечки (или рифление — то есть удаление с основания кости чешуек — позволяло, видимо, упростить операцию прикрепления рукоятки, или древка, к наконечнику) во всех случаях были нанесены в едином стиле. Первый подобный артефакт, сделанный, вероятно, из скелетных костей мамонта, была найден в 1932 г. в штате Колорадо. Следующий, также вырезанный из скелетной кости мамонта, — пять лет спустя, в Кловисе, штат Нью-Мексико. Эти более крупные наконечники, известные как наконечники из Кловиса, залегали в более глубоких культурных слоях, поверх которых были обнаружены представители другого типа артефактов, известные как наконечники из Фолсома, которые, очевидно, сделаны из костей скелета бизона[375]. Что же касается времен и сроков, можно считать «доказанным» тот факт, что впервые нога человека ступила в Новый Свет более чем 10 тысяч лет тому назад. Прежняя теория — относительно недавней колонизации Америки — потерпела крах. «Остальное — дело истории», как впрочем и преданных ей американских археологов. Но, в отличие от художественных произведений, наша драма вовсе не клонилась к счастливой развязке. Колесо Фортуны завершило свой очередной круг, и старые теории оказались на высоте: претерпев сокрушительные удары судьбы и поругание, авторитетное догматическое учение проявило чудеса выносливости и, словно ориентируясь по звездам и солнцу, вновь вернулось на «круги своя». Жизнь идет своим чередом, и колесо Фортуны совершает оборот за оборотом. В результате археологических раскопок наконечники, типичные для культуры Кловиса, были обнаружены по всей континентальной части Соединенных Штатов. Американские ученые укреплялись во мнении, что эти каменные орудия принадлежали первым колонизаторам Америки. Кроме того, все меньше сомнений оставалось в том, что первопроходцы появились здесь на исходе последнего крупного оледенения. Это были охотники за крупной дичью эпохи Верхнего палеолита, которые, двигаясь вслед за мамонтами, покинули Азию и переправились через Берингию. В 1964 г., основываясь на данных радиоуглеродного метода, американский геохронолог Вэнс Хейнс собрал воедино и сопоставил между собой даты стоянок человека, на которых были найдены наконечники, аналогичные обнаруженным в Кловисе Эти даты ограничивались рамками временного отрезка между 11 — 11,5 тысячами лет тому назад (такова датировка древнейших наконечников, характерных для эпохи или культурного периода, также получившего название Кловис) и 12 тысяч лет тому назад. Ни один из наконечников невозможно отнести к более раннему периоду. Последняя дата очень важна для геологов, поскольку они соотносят ее с событиями, происшедшими непосредственно вслед за образованием между двумя тающими ледниковыми плитами Северной Америки коридора, по которому, вероятно, двигались первопроходцы, пришедшие в Канаду с Аляски и расселившиеся впоследствии по всей Америке (примерно 12—13 тысяч лет тому назад). Эти две ледниковые плиты были поистине громадны. Одна из них — Канадская, или Лаврентийская, — располагалась поверх плато Канадский щит, перекрывала Гудзонов залив и простиралась далее к востоку. Другая, получившая название по горной системе Кордильеры, спускалась с горных массивов на запад. На основании данных, полученных еще в 60-е гг., можно, по видимости, утверждать, что эти гигантские ледяные плиты охватывали весь северо-американский континент[376] (рис. 7.1.).
 Puc.7.1
Базовая концепция «Первый Кловис» превратилась в наши дни во всесторонне разработанное академическое учение. Она исходит из того, что люди не могли прийти в Америку раньше эпохи Кловис, ибо в более глубокой древности путь мигрантам из Азии преграждал ледник. Все связанные с концепцией «Первый Кловис» даты, указывающие на начальный период заселения Америки, совпадают с другим важным событием, произошедшим примерно 13 тысяч лет тому назад, — формированием во льдах гигантского коридора, протянувшегося с северо-запада на юговосток. Прежде чем сформировался этот коридор, ни один человек не смог бы пересечь ледовый панцирь — на этом факте базируются доказательства теории «Первый Кловис». Причем никому и в голову не приходит, что, когда коридор сформировался, он представлял собой протянувшуюся на многие тысячи километров пустыню, которую невозможно было бы преодолеть, не запасшись заблаговременно продовольствием, типа тех обедов или сухих пайков, которые выдают отправляющимся на продолжительную экскурсию туристам. Между тем в столь грандиозной теории должно быть продумано все, вплоть до мелочей. Архитекторы этой теории стали теперь маститыми учеными, кардиналами от науки. Теория вполне готова к очередному витку эскалации вооружений, к защите и обороне своего статуса в научном мире и борьбе за сохранение status quo. Новые атаки и решительная оборона длятся уже более тридцати лет.
Силой, но одновременно и главной слабостью концепции «Первый Кловис» является необходимая взаимосвязь датировки самых древних наконечников типа Кловис периодом, наступившим сразу после формирования коридора — и, наоборот, коридор между ледниковыми плитами должен был открыться непосредственно перед эпохой Кловис. Эта гипотеза — своего рода несущая конструкция. Чуть толкнешь ее — и вся теория рассыплется, как карточный домик. Конструктивной опорой теории является жесткая взаимосвязь даты появления первопроходцев на американском континенте и предшествовавшей ей даты формирования коридора между ледниковыми плитами. Если заселение Северной или Южной Америки окажется возможным отнести к более раннему периоду истории — всего на какие-нибудь тысячи лет назад, ко времени, когда коридора не существовало, — то теория обрушится под собственным весом. В таком случае первое вторжение на континент произошло до Последнего ледникового максимума (ПЛМ), не позже 22 тысяч лет тому назад.
Puc.7.1
Базовая концепция «Первый Кловис» превратилась в наши дни во всесторонне разработанное академическое учение. Она исходит из того, что люди не могли прийти в Америку раньше эпохи Кловис, ибо в более глубокой древности путь мигрантам из Азии преграждал ледник. Все связанные с концепцией «Первый Кловис» даты, указывающие на начальный период заселения Америки, совпадают с другим важным событием, произошедшим примерно 13 тысяч лет тому назад, — формированием во льдах гигантского коридора, протянувшегося с северо-запада на юговосток. Прежде чем сформировался этот коридор, ни один человек не смог бы пересечь ледовый панцирь — на этом факте базируются доказательства теории «Первый Кловис». Причем никому и в голову не приходит, что, когда коридор сформировался, он представлял собой протянувшуюся на многие тысячи километров пустыню, которую невозможно было бы преодолеть, не запасшись заблаговременно продовольствием, типа тех обедов или сухих пайков, которые выдают отправляющимся на продолжительную экскурсию туристам. Между тем в столь грандиозной теории должно быть продумано все, вплоть до мелочей. Архитекторы этой теории стали теперь маститыми учеными, кардиналами от науки. Теория вполне готова к очередному витку эскалации вооружений, к защите и обороне своего статуса в научном мире и борьбе за сохранение status quo. Новые атаки и решительная оборона длятся уже более тридцати лет.
Силой, но одновременно и главной слабостью концепции «Первый Кловис» является необходимая взаимосвязь датировки самых древних наконечников типа Кловис периодом, наступившим сразу после формирования коридора — и, наоборот, коридор между ледниковыми плитами должен был открыться непосредственно перед эпохой Кловис. Эта гипотеза — своего рода несущая конструкция. Чуть толкнешь ее — и вся теория рассыплется, как карточный домик. Конструктивной опорой теории является жесткая взаимосвязь даты появления первопроходцев на американском континенте и предшествовавшей ей даты формирования коридора между ледниковыми плитами. Если заселение Северной или Южной Америки окажется возможным отнести к более раннему периоду истории — всего на какие-нибудь тысячи лет назад, ко времени, когда коридора не существовало, — то теория обрушится под собственным весом. В таком случае первое вторжение на континент произошло до Последнего ледникового максимума (ПЛМ), не позже 22 тысяч лет тому назад.
Новаторы, еретики или ученые?
История предстает в новом свете, если согласиться с концепцией новаторов, полагающих, что заселение Америки в действительности началось до, а не после Ледникового периода — эта радикально новая теория относит дату вторжения мигрантов к периоду, отстоящему от эпохи Кловис на 15 тысяч лет назад. Хотя эта теория кажется на первый взгляд менее убедительной, но данные, на которых она основана, показывают, насколько маловероятно, что наконечники типа Кловис относятся к периоду начального заселения американского континента, ибо факты и различные новые технологии свидетельствуют о более раннем заселении Южной Америки. В течение последних десятилетий возросло число археологов-новаторов, обнаруживших новые стоянки человека и артефакты, относящиеся к эпохе, предшествующей эре наконечников типа Кловис, возросло и число сторонников этой теории. В одной научно-популярной статье, опубликованной еще в 1990-е гг., были множественные ссылки на результаты раскопок в местах древнейших стоянок человека — я насчитал их восемнадцать, — и все они оспаривают у стоянок эпохи Кловис право называться старейшими. Однако ученые-консерваторы по крайней мере половине из этих стоянок отказывают в праве именоваться древнейшими и в целом не видят оснований оспаривать устоявшиеся взгляды, и не понимают, из-за чего, собственно, поднялась вся эта шумиха в последние два десятилетия. Большинство новаторов вступили в ожесточенную схватку со сторонниками базовой концепции «Первый Кловис», оспаривающими корректность научной методологии своих соперников, ставящими под сомнение правильность раскрытия культурного слоя и беспристрастность его изучения. Под перекрестным огнем защитников официальной концепции уцелели совсем немногие стоянки, зато их сторонники до сих пор держат круговую оборону. Наиболее упорные бои идут сейчас вокруг стоянки в Монте-Верде на севере Чили и в Медоукрофт Рокшелтер[377] на юго-западе Пенсильвании (см. рис. 7.1). В число новобранцев добровольно вступили Кактус-Хилл в Виргинии и Топпер/Биг Пайн в Южной Каролине[378]. Теперь кажется совершенно безосновательным a priori утверждать, что первые племена мигрантов начали обживать Америку после Ледникового периода, а не до него, ведь многие окруженные со всех сторон морями и океанами земли были заселены задолго до Последнего ледникового максимума, среди них и Австралия, и Новая Гвинея, и даже архипелаг Бисмарка, и Северные Соломоновы острова. С другой стороны, есть все основания предполагать, что глобальные процессы Великого Оледенения уничтожили свидетельства пребывания в Северной Америке первобытных людей, зато относительно недавние стоянки сохранились очень хорошо. А следовательно, тот факт, что найдено множество наконечников типа Кловис, которые сделаны спустя несколько тысячелетий после Последнего ледникового максимума и дошли до нас в прекрасно сохранившемся культурном слое, еще не является доказательством того, что они принадлежат эпохе первопроходцев. До нас дошли материальные следы доледниковых поселений человека, хотя и не в столь хорошей сохранности. Все это доказывает лишь то, что последняя официальная доктрина, сложившаяся еще в XIX в. и утверждающая, что ранее 10 тысяч лет тому назад Америка оставалась необитаема, — несправедлива. В результате следовало бы предположить, что (любые) следы доледниковых поселений человека ставят под вопрос концепцию «Первый Кловис», доказывая ее слабость, что, впрочем, не бросает тени на достоверность и важность для науки самих наконечников типа Кловис.
Монте-Верде
Остановимся на некоторых ключевых моментах спора вокруг Монте-Верде. Том Диллхей из университета штата Кентукки с 1977 г. участвовал в раскопках древних стоянок человека в Монте-Верде на юге Чили. Он и его коллеги добыли богатый материал, который мог бы уничтожить последние цитадели сторонников концепции «Первый Кловис», — но не уничтожил. Ибо хотя в Монте-Верде и были обнаружены следы пребывания человека в самой глубокой древности, — например, возраст орудий из мелких дробленых камней оценивается примерно в 33 тысячи лет, — «лучшие» артефакты относятся к значительно более позднему периоду. Место древней стоянки превратилось со временем в торфяное болото, которое и сохранило до наших дней немалое число органических и неорганических свидетельств пребывания здесь первобытного человека. Среди них — и отпечаток ступни; и деревянные артефакты; предполагаемые остатки жилых сооружений; очаги; останки лам и мастодонтов эпохи палеозоя, в частности — предметы, вырезанные из костей этих животных; а также зерно, орехи, фрукты, ягоды и клубни. Датировка при помощи радиоуглеродного метода позволила установить возраст органических останков, он составляет от 11 790 до 13 565 (средний возраст — 12 500) лет. Были также найдены и самые обычные каменные орудия: мелкие дробленые камни и крупные булыжники[379]. Монте-Верде находится в 12 км (7500 милях) к югу от того места, где примерно 12 500 лет тому назад сформировался коридор между ледниковыми плитами Аляски. Там найдены следы стоянки человека эпохи Кловис, которая впоследствии была названа Золотым веком первобытной цивилизации. В связи с этим археологи ставят следующий вопрос: как 13 тысяч лет тому назад хоть одно племя смогло преодолеть гигантскую пустыню ледникового коридора, за счет чего у людей было столько свободного времени, что они могли себе позволить дальнее путешествие на юг, и почему изменились их культурные навыки[380]. Все эти факты и соображения содержат в себе огромный потенциал, способный разрушить концепцию «Первый Кловис». И нетрудно предсказать, что «Первый Кловис» вынужден будет сдаться. Чтобы выйти из замкнутого круга «результативной ничьей», необходимо, чтобы коллегия состояла из независимых арбитров. В 1997 г. в Кентукки, чтобы ознакомиться с докладом Диллхея, посетить Чили и осмотреть место стоянки человека, была приглашена группа выдающихся специалистов по палеоиндейской культуре, в которую входил также известный скептик Вэнс Хейнс. В программе были и другие мероприятия. Каждому члену группы вручили изданный Смитсоновским институтом подробный отчет о работах на месте стоянки древнего человека. Ученые пришли к единодушному мнению относительно того, что археологические раскопки ведутся действительно на месте древней стоянки человека, возраст которой составляет приблизительно 12 500 лет. Эти данные содержатся в отчете, опубликованном в том же 1997 году академическим журналом «American Antiquity». Впоследствии в этом же журнале вышла статья, подписанная группой специалистов в области древней хронологии, включая Вэнса Хейнса. Ученые подтвердили приданные гласности даты стоянки в Чили и уничтожили даже тень подозрения в том, что эти даты появились под влиянием данных, полученных в ходе радиоуглеродного анализа[381]. Затем в результате некоторых не слишком важных событий дебаты ученых предстали перед нами в своем истинном свете. Оказалось, что это вовсе и не дебаты, а грязные академические дрязги, в коих не остается места для непредвзятости, объективности и благоразумия, которого поневоле ждешь от ученых пэров. Диллхей предпочел удалиться со сцены, и вскоре глухая стена предвзятости, неприязни и скептицизма рухнула: незаурядная древность Монте-Верде, в конце концов, была признана официальной наукой. Независимый археолог-консультант Стюарт Фидель опубликовал внушительных размеров критическую статью, в коей язвительно нападал на предыдущий отчет о раскопках на месте древней стоянки в Монте-Верде. Драма, в которую были вовлечены mass media, принимала все больший размах. Фидель поступил весьма неординарно: он предпочел атаковать не официальный журнал академиков и ученых пэров, оказавшихся столь предвзятыми арбитрами, но научно-популярный журнал «Discovering Arhaeology»[382]. Здесь регулярно печатались полемические статьи, авторами которых были коллеги и сторонники Диллхея, а в дальнейшем — скептические комментарии Хейнса, привыкшего с пристрастием отвергать неугодные ему данные. К этому времени выступить с полемическими статьями успели не только сторонники Диллхея, но и представители официальной науки — в частности ученые Археологического института Америки, печатным органом которого является весьма представительное издание «Arhaeology», отличающееся высоким уровнем культуры и строгим тематическим отбором статей в соответствии с профилем издания. С каким пафосом выступали полемисты, будет видно из следующих фрагментов. Два первых извлечены из статей, опубликованных в «Discovering Arhaeology»:
«В том случае, если новая идея или теория кажется убедительной, старомодная концепция с миром умирает, и пусть земля ее научных изысканий будет ей пухом. Что же касается «архаико-археологии», то факты упорно говорят об обратном — все согласны, что реальность далека от идеала и события так редко развиваются в согласии с ним. Зато своенравию и гордости видных ученых, кажется, не будет конца. В результате желчных и самонадеянных нападок в стиле доводов ad hominem складывается атмосфера нескончаемых дебатов, голос объективности тонет в шуме личной полемики. Поэтому концепции, особенно старомодные, сопротивляются смерти еще усерднее знаменитого Брюса Виллиса. Именно в таком положении оказалась теперь концепция «Первый Кловис», называемая также «Кловис-прима», которой уже перевалило за пятьдесят»(Джеймс Адовазио «Смерть концепции и смертельные перестрелки»[383].)
«[Фидель], с ног до головы покрытый коростой почти конспираторской подозрительности и сутяжничества, просто погряз в удручающих по своей частоте и язвительности замечаниях (примером чему служит и «Диллхей, с его гамлетовскими страданиями»). При этом в своей критике первый не жалует ни правых, ни виноватых. Кстати сказать, самым полезным, продуктивным и конструктивным поступком (и, конечно, самым здравым и прекраснодушным) было бы его решение послать Диллхею таблицу критики на критику, в которой он мог бы тщательно рассортировать всевозможные проблемы: какие из них можно отнести к разряду тривиальных, а какие — передовых, незначительные же вопросы можно было бы распределить между глобальными проблемами, дериватами которых они являются, — а затем представить сей опус на рассмотрение в академический журнал въедливых пэров-арбитров. Но Фидель не сделал даже этого».А следующие два фрагмента взяты из статей, опубликованных в «Arhaeology»:(Дэвид Мелътцер «На Монте-Верде»[384].)
«Тон полемических выступлений Фиделя крайне тенденциозен и недоброжелателен. Ученый попросту игнорирует фактический материал, не работающий на его критические взгляды. А когда дело доходит до дискуссий, то альтернативные концепции в его изложении предстают как нечто до того несуразное, что нечего и думать подвести под них хоть сколько-нибудь научную платформу».(Мишель Коллинз, «Древняя стоянка в Монте-Верде»[385].)
«Меня утомил въедливый тон замечаний Фиделя, его звериное чутье на малейшие противоречия в интерпретации, которые были допущены в отчете о Монте-Верде более двадцати лет назад. Фидель стал жертвой своих собственных убеждений, которые, на мой взгляд, ни на йоту не соответствуют действительности. Держу пари на крупную сумму, что ученые мужи из группы исследователей Монте-Верде достойны всяческих похвал; хватит относиться к ним как к подкаблучникам: они достойны уважения уже за то, что с готовностью откликнулись на предложение опубликовать подробнейший отчет о находках, хотя их и преследовали дурные предчувствия относительно предложенной ими датировки... Идея моя состоит в том [sic], что тот хочет пожить за чужой счет, кто выкапывает из архивов предварительные оценки и, основываясь на них, обкладывает двойным налогом критики каждый постулат, считая себя вправе использовать любое слово отчета о Монте-Верде против самого Монте-Верде и против запроектированных там научно-исследовательских работ. А еще моя идея состоит в том [sic], что нужно сохранить способность пересматривать свою точку зрения (пусть эта способность не покидает вас до тех пор, пока вы будете сю гордиться). Поэтому решение Фиделя отдать в печать рукопись, которую тот не решился предложить Диллхею или кому-нибудь из его коллег для предварительного чтения и комментариев, — вызвало у меня массу вопросов. Поясню столь дорогую моему сердцу мысль о необходимости самокритики на примере Фердинанда, сознание которого, очевидно, давно превратилась в пороховой склад критических замечаний; но и в его голове здравые мысли можно отсеять от плевел, для этого нужно только пойти на доверительный контакт с ведущими специалистами, работающими на раскопках в Монте-Верде. Если подобный обмен мнениями произошел бы, мы смогли бы получить лаконичный отчет о действительном положении дел, тогда плевела насмешек и предвзятых мнений рассеются, как облака... Ученые, работающие в Монте-Верде, видимо, давно уже затаили в глубине сердца вопрос: «Какие, собственно, задачи ставит перед собой Фидель?» Действительно, преследует ли его критика цель выяснить, были ли в Монте-Верде поселения во времена, предшествующие эпохе наконечников типа Кловис? Или он, как уличный воришка, желающий пожить за чужой счет, был выслежен полицией и пойман с поличным, отчего и попал в заголовки криминальных хроник. Кто знает, возможно ли вообще преследовать какую-то цель в этой жизни, или жизнь преследует нас?»Другие отзывы на критические статьи Фиделя были одобрительными. Его концепция вызвала фурор, и теперь все прежние выводы, касающиеся Монте-Верде, поставлены под огромный, парящий в воздухе вопрос. Что же касается глубоко взволнованных этим вопросом лежебок, то есть людей, привыкших усваивать информацию в положении лежа, или даже тех «архаико-археологов», которые не были на месте раскопок в Монте-Верде, то все они подобны больным, пытающимся оценить ситуацию и понять, как же все-таки выбрать одного из массы врачей, которые наперебой обвиняют друг друга в профессиональной некомпетентности. В этой гипотетической ситуации наиболее разумным будет усомниться в непредвзятости высказываемых врачами мнений относительно квалификации друг друга. Неистовость воплей, которые доносятся из стана сторонников концепции «Первый Кловис», пропорциональна их ужасу перед Монте-Верде, измеренному в децибелах. К тому же когда встает вопрос, не древнее ли те или иные первобытные стоянки эпохи Кловис, то по подобному шумовому эффекту можно даже определить, насколько заслуживают уважения те или иные возражения. Если эти бурные возражения исходят из стана защитников концепции «Первый Кловис» — усомнитесь в них. Если возражения (или подтверждения) исходят от нейтральных экспертов или слышатся со всех сторон от профессионалов, представляющих различные школы и направления в археологии, то отнеситесь к ним со вниманием.(Давид Томас «Взгляд археолога на проблему противоречий в датировке Монте-Верде»[386].)
Медоукрофт
Есть и еще одна стоянка первобытного человека, которая давно уже находится на осадном положении в окружении ученых и археологов — это Медоукрофт Рокшелтер. Джеймс Адовазио, археолог из Пенсильвании, уже в течение тридцати лет возглавляет научно-исследовательскую работу в этом месте (см. Фото 20). Он и его коллеги углубились в землю на одиннадцать культурных слоев, в результате было извлечено 20 тысяч орудий из дробленого камня и другие артефакты, а также огромное количество животных и растительных останков. На основе радиоуглеродного анализа был определен возраст пятидесяти двух объектов, найденных на стоянке в Медоукрофт. Старейшим оказался артефакт, извлеченный с самого «дна», из так называемого «стерильного» пласта глины, в котором нет ни малейших признаков животной или растительной жизни, — возраст этого артефакта составляет 31 тысячу лет. Самым же поздним оказался артефакт, извлеченный из верхнего культурного слоя, возраст этого объекта — 1000 лет. Некоторые артефакты, бесспорно связанные с палеоиндейским населением, можно датировать 16 225 годами, а возраст самого раннего культурного слоя достигает, по мнению некоторых исследователей, 19 тысяч лет[387].
 Фото 20. Археолог Джеймс Адовазио осматривает многочисленные культурные слои в Медоукрофт Рокшелтер, одной из наиболее важных стоянок эпохи пре-Кловис в Америке.
Фото 20. Археолог Джеймс Адовазио осматривает многочисленные культурные слои в Медоукрофт Рокшелтер, одной из наиболее важных стоянок эпохи пре-Кловис в Америке.
Публикация материалов, содержащих столь древние датировки, немедленно вызвала шквал протестов. И неудивительно, что среди гула критики выделялся один полный гнева голос — голос Вэнса Хейнса. Кажется, на добрый километр растянулись критические замечания, год за годом вращающиеся вокруг одних и тех же тем и разбирающие детали и подробности данных, предоставленных стратиграфией[388]; документальные свидетельства; датировки тканей животного происхождения и возможные неточности, связанные с радиоуглеродным анализом. Адовазио, видимо, относился к подобной критике как к патологическому расстройству на почве скептицизма. Одни и те же въедливые вопросы требовали все новых ответов, коими исписана не одна тысяча страниц. Стараниями независимого специалиста в области геоморфологии радиоуглеродный метод со всеми его неточностями был признан несостоятельным и погребен в 1999 г., но Хейнс не пожелал предать его забвению, ему все-таки хотелось получить основанные на этом методе датировки некоторых остававшихся у него археологических находок: скорлупы ореха и нескольких семян. Эта капля переполнила чашу терпения Адовазио. На состоявшейся три года назад конференции в Монте-Верде Адовазио, как известно, ответил Хейнсу: «Сколько бы вы меня не спрашивали и не переспрашивали, я никогда не откажусь от принятых мной дат, поскольку начиная с 1974 года не было такого критического замечания, на которое мы не обратили бы внимания и не дали бы ему достойный отпор. Этому я отдал полжизни»[389]. Если столь известные скептики, как Хейнс и Фидель, правы, им стоит поздравить друг друга с тем, что их союз перед лицом массы вымышленных свидетельств и сфабрикованных фактов оказался столь прочным. Если же они, наоборот, не правы или же их научный метод тенденциозен, то их можно было бы поздравить с успешной борьбой против прогресса в американской археологии, с тридцатилетней стагнацией и с тем, что концепции «Первый Кловис» была искусственно продлена жизнь — хотя у бедняги атрофировались все жизненно важные органы, она дотянула-таки до семидесяти. Что ни говори, это — выдающееся достижение даже для современной науки, которая семимильными шагами идет от открытия к открытию. В начале же прошлого, двадцатого, столетия даже Хрдличка не добился подобного результата в искусстве продлевать жизнь идеям, паразитирующим на науке. В Северной Америке были обнаружены еще две стоянки первобытного человека: Кактус Хилл и Топпер — они древнее, хотя и не намного, стоянок типа «Первый Кловис» и словно состязаются друг с другом в том, кто из них привлечет к себе больше внимания. Стоянка в Кактус Хилл — на восточном побережье близ Ричмонда, штат Виргиния, — всесторонне описана в отчете, составленном археологами двух соперничающих друг с другом частных экспедиционных групп, которые поджидают всякого неосторожного скептика, чтобы наброситься на него[390]. Одну из этих групп возглавляют Джозеф и Линн Макэвой из отделения Института исторического наследия штата Виргиния, другую — Мишель Джонсон из Археологического общества Виргинии. Кактус Хилл — это древняя песчаная дюна, склоны которой летом сплошь покрыты опунцией — кактусом в виде груши с шипами. Кактус Хилл, как это часто случается, был случайно найден наблюдательным фермером при весьма примечательных обстоятельствах. Он заметил каменный наконечник, который сполз с вершины холма и лежал неподалеку от него в грудах песка, при этом оставшаяся за ним борозда указывала на место, откуда тот скатился. Закапываясь сквозь песок в глубину времен, экспедиционные группы нашли каменные наконечники, которые значительно древнее тех, что принадлежат к рифленому типу Кловис. В самом архаичном культурном слое археологи обнаружили орудия из дробленого камня, мотыги, кварцитовые стержни и несколько небольших режущих орудий. На основе радиоуглеродного метода был определен возраст этих древнейших артефактов, они были выполнены 15— 16 тысяч лет тому назад, то есть во времена, предшествующие формированию ледового коридора. В одном из наиболее древних культурных слоев ученые обнаружили необычные каменные наконечники, в потенции содержащие все те стилистические особенности, которые впоследствии воплотились в типе Кловис, предшественниками которого эти наконечники являются. Нет нужды говорить, что Хейнс и Фидель бывали в этих местах — правда, цель их визита сводилась к стремлению усомниться в правильности определения возраста найденных здесь артефактов, которым не удалось, конечно, постоять за себя и доказать, что они отнюдь не молоды, но гораздо старше, чем того хотелось бы некоторым ученым[391]. Выше было упомянуто название еще одной стоянки первобытного человека, Топпер, штат Южная Каролина, носящей имя Дэвида Топпера — лесничего, который ее обнаружил. Раскопками руководит американский археолог, аккредитованный Университетом штата Каролина — Эл Гудъе. В результате наводнения, затопившего три года назад одну из древних стоянок, Гудъе и его экспедиционной группе пришлось покинуть ее и заняться повторными раскопками в Топпер. На этот раз они вели археологические работы на большой глубине, ниже культурного слоя эпохи Кловис. Гудъе первоначально был убежденным членом одиозной партии сторонников концепции «Первый Кловис». Но то, что он теперь нашел, не могло его не шокировать — Гудъе обратился в новую веру. Ниже культурного слоя эпохи Кловис залегали небольшие сланцевые режущие орудия, точеные резцы, мотыги и микролезвия. По технологии изготовления эти орудия более напоминали верхнепалеолитические памятники Сибири, нежели какой-либо из известных артефактов юго-восточной части Америки. Датировка на основе люминесцентного анализа позволила установить, что возраст найденных здесь артефактов составляет 13 тысяч лет[392]. Что же скептики — вновь принялись бить в набат? Конечно. Но теперь их усилия были направлены не против дат, а против самих орудий. Возможно ли, чтобы они были делом рук человеческих? Или такое возражение: большинство стоянок эпохи Кловис не таит под собой никаких более древних орудий. При этом Вэнс Хейнс, как свидетельствует журнал «Science», отказывается учитывать роль случайных «совпадений». Однако с тем же успехом можно отрицать эпоху римского владычества на том основании, что далеко не каждый дом современной Англии хранит где-нибудь под собственным фундаментом остатки древнеримских построек. На что Хейнс отвечал: «Вот уже сорок лет, как они стали постоянно попадаться мне на глаза»[393]. На юго-востоке штата Висконсин обнаружены еще две стоянки первобытных людей: Скифер и Гебиор — будучи совсем немногим древнее эпохи Кловис, они оказались вовлечены во всеобщий круговорот вещей, где взлеты, связанные с признанием их возраста, чередуются с падениями под натиском скепсиса и критики. Датировка на основе радиоуглеродного анализапозволила определить приблизительный возраст этих стоянок — 12 500 лет. На одной из них, Гебиор, расположенной в районе города Кеноша, найдены орудия из дробленого камня, топорики, а среди костей убитого мамонта были обнаружены два заточенных с обеих сторон камня[394]. Археологам, таким, как Диллхей, Адовазио, Джозеф и Линн Макэвой, Джонсон, Гудъе и их коллегам, пришлось составить документальные отчеты невиданной длины о всех находках и артефактах, без прикрас описать все и вся, ответить на все критические замечания, обычно начинающиеся со слов «а что, если...». Благодаря стараниям этих ученых дискуссии, постоянно разгорающиеся вокруг стоянок, предшествующих эпохе Кловис, приняли совершенно иное направление. Наконец-то корректные споры и обсуждения в академических кругах стали более заметным явлением общественной жизни, чем критические замечания, которые, в свою очередь, играли значительно более важную роль в обсуждениях, чем вопли тех, кто «предается возмущению сверх меры». Партия приверженцев теории «Первый Кловис» наконец почувствовала свою слабость. На новом витке развития науки решительно возобладала новая, или альтернативная, теория, ставящая своей целью объяснить, как заселялся Новый Свет в глубокой древности. Дебаты развивались по следующему сценарию: сперва обсуждался вопрос, действительно ли люди проникли в Америку и расселились по ее территории во времена, предшествующие эпохе Кловис; а затем — какие экзотические маршруты были ими выбраны и сколько их было. Вот эти «великие стрелы, направленные в Америку»[395], которыми, если все действительно происходило именно так, символически обозначают многочисленные пути продвижения в глубь континента переселенцев со всех концов света, — это маршрут, пролегающий по западному побережью; морской путь вдоль западного берега; северно-атлантический путь из Европы; маршрут из Австралии, пролегающий по южным водам Тихого океана; а также южноамериканский путь вторичного заселения после Ледникового периода. Однако все эти доисторические сценарии не столь полно и досконально подтверждены данными археологии, как те, которые освещают события в Монте-Верде и Медоукрофт.
Сравнительно-историческое языкознание
Прежде чем приступить к обсуждению возможных маршрутов и того, что могли бы рассказать о них гены, я хотел бы переключить внимание и как бы перенестись в область иной научной дисциплины, которая всегда манит обещаниями приоткрыть завесу тайны перед взглядом, устремленным в далекое прошлое. Наука эта — сравнительно-историческое языкознание. К сожалению, академики, со всем присущим им темпераментом, и здесь навели беспорядок, лишив науку ее природной ясности и здравомыслия. Сравнительно-историческое языкознание играет весьма почтенную — и даже ничуть не менее важную, чем археология, — роль в процессе выдвижения гипотез относительно того, кем же были первые жители Америки. Свой заметный вклад в каждую из этих областей науки внес повсеместно уважаемый отец-основатель и достойнейший ученый-энциклопедист Томас Джефферсон. В 1784 г. он руководил работами, которые велись на древнем кургане в Виргинии — это были первые в истории Америки научные археологические раскопки. А четырьмя годами раньше Томас Джефферсон приступил к поистине новаторским исследованиям языка аборигенов, составив пробные словари наречий коренных жителей. Посредством сравнительного языкознания он хотел выяснить, где их прародина — проследить, куда тянутся своими корнями их языки. К 1809 г. он заполнил словарными статьями многие десятки страниц, однако во время транспортировки материалы были украдены и почти полностью испорчены. Джефферсон набело переписал уцелевшие фрагменты и послал их в Философское общество Америки. Таким образом, сквозь это, пока еще очень мутное, но обещающее стать прозрачным, окно в прошлое Америка впервые могла созерцать национальные судьбы аборигенов[396]. Как же, собственно, Джефферсон намеревался использовать лингвистические данные? Его идея была очень проста на словах, но вряд ли осуществима на практике. Суть ее состояла в следующем: со временем языки дробятся и разветвляются. Если же лингвистические данные отражают процесс распространения народов по континенту и дробления их на группы, то мы могли бы определить, где и когда произошло подобное разделение[397]. В результате можно получить представление о родовом древе миграций. Например, европейские языки, такие, как немецкий, французский, испанский и английский[398], являются ветвями на могучем древе индоевропейских языков. Сопоставив, как действуют в этих языках некоторые правила и законы, лингвисты доказали, что эти языки нельзя считать непосредственными ответвлениями от центрального ствола, ибо они соединены с индоевропейским целым посредством таких мертвых языков-предков, как латынь и протогерманский, наследниками которых они и являются. Если когда-нибудь удастся полностью реконструировать родовое древо европейской группы языков — хотя, возможно, на нем и останутся некоторые белые пятна, — тогда мы сможем проследить географические аспекты дробления народов. Если мы перенесем на карту данные генеалогического древа языков, то окажется, что французы и испанцы стали наследниками великой латыни, которая, в свою очередь, произошла от протоиталийского языка. А английский и немецкий уходят корнями в западный диалект протогерманского языка. Все представители европейской языковой семьи в конечном счете восходят к протоиндоевропейскому корню, от которого они отделились многие тысячи лет тому назад. Теперь можно приступить к нанесению на карту больших стрел: от пункта А — к пункту В и С, от С — к D и так далее, — и написать историю заселения Европы, как о ней рассказали сами языки. Проблема заключается в том, что жизнь всегда оказывается сложней теории. Новые языки начинают формироваться не только в результате разделения народов и накопления случайных изменений. Например, примерно 15% английской лексики восходит не к германским, а к французским корням. Дело в том, что после норманнского завоевания, произошедшего примерно тысячу лет тому назад, небольшая группа норманнской знати пополнила свой словарный запас за счет лексики французского языка — этот феномен принято называть языковым заимствованием. А затем в результате господства в Англии немногочисленной норманнской элиты — заметьте: массового вторжения норманнов в Англию не было — эти же заимствования из французского языка проникают в английский. Французский относится к романским языкам, но никому и в голову не придет утверждать, что он возник в результате непосредственного развития языка римских завоевателей, хотя Римская империя и оказала на французский столь значительное влияние, что язык порвал родственные связи с кельтской семьей языков и перешел в романскую группу. Этот феномен называют языковым сдвигом. Языковые заимствования и сдвиги изначально проникают в язык небольшой группы людей, которая вдруг с жадностью набрасывается на упрощенные модели чужого языка, а затем продолжает двигаться вместе со своим народом по предназначенному им пути миграций. Однако серьезных оснований сравнивать историю развития языков Европы и Америки, скорее всего, нет и быть не может. Америка была заселена предками современных аборигенов как минимум 45 тысяч лет тому назад и пережила не одно переселение народов, когда племена, жившие в ее пределах, начинали перемещаться по весьма сложным траекториям. Можно, кстати, привести несколько выразительных примеров сохранения собственного языка народами, вовлеченными в процесс переселения. Укоренившееся на новой почве языковое древо позволяет нам проследить, куда тянутся его корни, то есть выяснить путь миграций этого народа, и понять, где была его прародина. Народ сохраняет свой язык в том случае, если переселяется на никем не занятые территории. Вспомним, например, процесс распространения полинезийской ветви малайско-индонезийской семьи языков по прежде необитаемым островам Тихого океана. Неудивительно, что это маленькое «подсемейство» полинезийских языков кажется самостоятельным языковым древом, которое не устает повторять историю своего появления в этих местах, и рассказ его полностью подтверждается наблюдениями археологов. Более того, выводам лингвистики полностью соответствует и генетическая картина заселения Полинезии, полученная на основе данных современной генетики[399]. Очевидно, что у процесса завоевания Америки больше типологического сходства с покорением тихоокеанских просторов, ибо народы переселялись на просторы, где не ступала нога человека — в Новый Свет, — и подобно звездным лучам расходились в разные стороны. Однако Америка была заселена задолго до Полинезии, на которой первые жители появились лишь 8000—3500 лет тому назад — поэтому провести однозначные параллели между Америкой и Полинезией невозможно. Под прессом времен язык претерпел столь разительные изменения, что вряд ли кто-то сможет его реконструировать. Но не только страх перед океаном столетий мешает выяснить, когда же язык американских аборигенов откололся от праязыка, — лингвисты знают, что в языке неизбежно идут процессы распада, поэтому родственные узы, соединявшие некогда слово со словом, а язык с языком уже невозможно ни пронаблюдать, ни восстановить, особенно в том случае, если со времени разделения языков прошло более 6—8 тысяч лет[400]. А это немногим более половины того временного отрезка, который отделяет нас от эпохи Кловис. Поэтому реконструировать единое генеалогическое древо языков Нового Света и проследить, куда оно уходит своими корнями — задача непосильной сложности. Если история развития языкового древа американских аборигенов может быть прослежена только на протяжении последних 7000 лет, то мы увидим лишь его вершину — придется смириться с тем, что его корни и нижние ветки давно отмерли. Мы увидим лишь полусгнивший валежник, у нас будет мало шансов приладить одну ветку к другой и представить, каким некогда было дерево. Именно так обстоит дело с Северной, Центральной и Южной Америкой, где сосуществует множество языковых семей. В целом их насчитывается более сотни. Они включают в себя около 1200 языков. По мнению многих лингвистов, все эти языки невозможно собрать воедино и представить в виде целостного древа. Остается лишь мечтать, что когда-нибудь их удастся объединить хотя бы в несколько языковых семейств. И уже по количеству семейств можно будет судить о том, каким числом ветвей проникло некогда в Америку неведомое нам языковое древо — это все, чего можно ожидать от научной интуиции. Однако все эти Доводы рассудка не останавливают энтузиастов от попыток реконструировать праязык американских аборигенов. По оценкам австралийского лингвиста Роберта Диксона, в промежуток времени, отстоящий от нас на 12—20 тысяч лет, в Америку проникло около дюжины отдельных этнических групп, говорящих на различных языках. Американский лингвист Джоанна Николc, известный специалист в области анализа изменений, которые претерпевали в древнейшие времена отдельные слова и словоформы языка, считает, что для того, чтобы объяснить существующее разнообразие языковых семейств в Америке, нужно предположить, что с начала заселения Нового Света прошло приблизительно 35 тысяч лет, причем среди переселенцев из Азии было множество самобытных племен. Английский лингвист Дэниэл Неттл, наоборот, считает, что великое разнообразие языковых семейств Северной и Южной Америки сложилось значительно позже, примерно 12 тысяч лет тому назад[401]. Ученый, отважившийся судить о времени заселения Америки по количеству семейств, представленных в говорах американских аборигенов, столкнется с серьезными трудностями. Его задача осложняется не только тем, что единой общепризнанной хронологической таблицы просто не существует, но и тем, что до сих пор у специалистов в области сравнительно-исторического языкознания нет четкого ответа на вопрос, по каким критериям определяются границы языковых семейств. Вследствие этого ученые, представляющие различные направления и страны, безрезультатно спорят о том, сколько же их, то есть языковых семейств, существует. Всех историков языка можно условно разделить на «дровосеков» и «портовых грузчиков», сваливающих весь товар в одно место. Первые даже несколько языков умудряются разделить на множество различных семейств. А вторые в небольшую группу суперсемейств объединяют великое множество языков. Сторонником одного из этих полярных направлений является маститый американский лингвист Джозеф Гринберг, который, как известно, значительно превзошел своих коллег из числа соотечественников в умении видеть общее: все языки коренных жителей Америки были объединены им в три огромные группы. На другом полюсе мы обнаружим классификации, насчитывающие свыше 160 семейств. Большинство американских специалистов склонны скорее преувеличивать, чем преуменьшать число языковых групп. Занимаясь вычислением среднего возраста американских языков (речь об этом уже шла выше), Джоанна Николс, опираясь на четко сформулированные правила сравнительной лингвистики, сделала вывод, что в Америке насчитывается 167 языковых «стволов» (т.е. лингвистических групп, для которых может быть реконструирован базовый праязык, или конкретная область генеалогического древа, от которой они начали во все стороны ветвиться). Подсчитывая количество языковых стволов, Николс ставила перед собой и еще одну задачу — сравнить Америку с другими странами на основании разнообразия бытующих там языков. При этом она стремится доказать, что в регионах (на континентах или субконтинентах), изолированных от внешних влияний, количество «стволов» представляет собой простую функцию времени[402]. Немало трудностей подстерегает ученого, положившего в основу своего аналитического метода принцип разделения языков на подклассы. Во-первых, всеми признанного метода сведения множества языков к общему стволу не существует, и даже ясно сформулированные Николс правила вычленения «стволов» оказывается возможным применять по собственному вкусу и разумению, то есть весьма субъективно. Во-вторых, в разных странах и на разных континентах языковые стволы проявляют различную продуктивность, производя на свет неодинаковое количество языков. По этой причине само существование «ствола» оказывается спорным[403]. И все же, хотя ученые определяют границы языков и «стволов» по-разному и на основании совершенно различных признаков, можно с уверенностью сказать, что Южная Америка значительно богаче Северной с точки зрения языкового разнообразия (см. рис. 7.2). После обнаружения ранее неизвестных математических закономерностей и соотношений были получены статистические данные, позволяющие использовать в аналитических целях скорее результаты самого простого подсчета, нежели сложные величины, полученные в результате научных обобщений, которые могут увести нас в дебри неведомого. Под результатами простого подсчета я разумею здесь данные о количестве языков. Речь здесь пойдет именно о них, а не о количестве языковых стволов. Я позволю себе воспользоваться материалами публикаций, подготовленными Николс и Неттл, и расположить собранные ими данные вдоль простых координат, указывающих на количество языков и время (рис. 7.2). В результате получился график, выявляющий взаимосвязь между числом языков, распределенных по регионам (т.е. по субконтинентам/континентам — как это было сделано в оригинальном исследовании Николс), и данными о времени заселения этих регионов. При этом я воспользовался средним арифметическим возрастом цивилизаций Северной, Центральной и Южной Америки, который составляет 16 тысяч лет. Полученный график говорит сам за себя: перед нами — просто прямая линия. Впрочем, эту закономерность обнаружила еще Николс количество языков в каждом регионе прямо пропорционально времени его заселения. Исключение составляет только Австралия. Уже стало общепризнанным фактом, что в Австралии нет того языкового богатства и разнообразия, которое можно было бы ожидать исходя из величественности сроков ее заселения. В Австралии до сих пор господствует одна языковая семья. Вышеописанная закономерность столь очевидна, что мы можем на ее основании проделать обратную операцию и вывести формулу для вычисления времени заселения Северной, Центральной и Южной Америки, а также предсказать, когда приблизительно в каждом из этих регионов появились люди. Результат получился интересный, я бы сказал — почти карикатурный, поскольку приблизительный лингвистический возраст заселения Северной Америки совпадает с эпохой Кловис — 10 500 лет; Центральной Америки — с Ледниковым периодом — 18 400 лет, а Южной Америки — с Ранним Верхним палеолитом — 30 700 лет тому назад.
 Рис. 7.2
Невозможно гарантировать абсолютную точность этих дат, но они дают ясное представление о последовательности заселения территорий Америки, которое, видимо, происходило с юга на север. Подобная хронология согласуется с представлениями о том, что Америка была заселена в период, предшествующий последнему Ледниковому периоду, во время которого Северная Америка в значительной степени обезлюдела — с чем связан эффект ее вторичного заселения, произошедшего уже после Последнего ледникового максимума (ПЛМ), а именно — в эпоху Кловис, и повлекшего за собой проникновение значительно меньшего количества языков, чем при первом заселении. В Южной Америке, которая почти не пострадала от оледенения, процесс формирования новых языков происходил непрерывно, что и отразилось на лингвистическом возрасте этого континента. Центральная Америка, численность населения которой увеличилась во время так называемого вторичного заселения за счет мигрантов с севера, занимает промежуточное положение между Северной и Южной Америкой по численности представленных в ней языков[404].
Хотя Южная Америка могла быть заселена людьми, переправившимися через Панамский перешеек из Северной Америки (см. ниже), но скорее всего южный континент представляет собой совсем иной Новый Свет. Короче говоря, нога человека впервые ступила на земли Южной Америки еще до Ледникового периода. Этот факт и дает нам наиболее простое и ясное объяснение того, что в лингвистическом отношении юг оказывается значительно старше севера. В Северной Америке, особенно на Крайнем Севере, о разнообразии языков говорить и вовсе не приходится, ибо во время ПЛМ эти земли обезлюдели. И лишь в период великого таяния ледников они вновь были заселены людьми, которые принесли с собой совсем небольшое число языков. Как мы далее увидим, концепция вторичного заселения Америки подтверждается также данными генетики.
Рис. 7.2
Невозможно гарантировать абсолютную точность этих дат, но они дают ясное представление о последовательности заселения территорий Америки, которое, видимо, происходило с юга на север. Подобная хронология согласуется с представлениями о том, что Америка была заселена в период, предшествующий последнему Ледниковому периоду, во время которого Северная Америка в значительной степени обезлюдела — с чем связан эффект ее вторичного заселения, произошедшего уже после Последнего ледникового максимума (ПЛМ), а именно — в эпоху Кловис, и повлекшего за собой проникновение значительно меньшего количества языков, чем при первом заселении. В Южной Америке, которая почти не пострадала от оледенения, процесс формирования новых языков происходил непрерывно, что и отразилось на лингвистическом возрасте этого континента. Центральная Америка, численность населения которой увеличилась во время так называемого вторичного заселения за счет мигрантов с севера, занимает промежуточное положение между Северной и Южной Америкой по численности представленных в ней языков[404].
Хотя Южная Америка могла быть заселена людьми, переправившимися через Панамский перешеек из Северной Америки (см. ниже), но скорее всего южный континент представляет собой совсем иной Новый Свет. Короче говоря, нога человека впервые ступила на земли Южной Америки еще до Ледникового периода. Этот факт и дает нам наиболее простое и ясное объяснение того, что в лингвистическом отношении юг оказывается значительно старше севера. В Северной Америке, особенно на Крайнем Севере, о разнообразии языков говорить и вовсе не приходится, ибо во время ПЛМ эти земли обезлюдели. И лишь в период великого таяния ледников они вновь были заселены людьми, которые принесли с собой совсем небольшое число языков. Как мы далее увидим, концепция вторичного заселения Америки подтверждается также данными генетики.
Северный анклав
Хотя мало кто из американских лингвистов признал крупнейшую американскую языковую семью Гринберга — Америнд, которая, по его словам, включает в себя 97% всех языков американских индейцев, почти никто не пытается оспаривать тот факт, что на Крайнем Севере Америки распространены языки, относящиеся лишь к двум семьям — эскимосско(инуитско)-алеутской[405] и на-дене[406]. И хотя к этому можно добавить еще много интересных фактов, пора сменить тему. В 1986 г. Гринберг решил объединить свои усилия с генетиками и дентологами — специалистами по изменению строения зубов. В публикации, подготовленной тремя членами этого новоявленного союза, была изложена теория, ставшая впоследствии известной как «гипотеза Гринберга»[407]. В ней органически соединились выводы, сделанные представителями трех дисциплин и свидетельствующие о трех независимых волнах миграции в Новый Свет. Эти волны ясно вычленяются на основании строения зубов, данных генетики и лингвистики. Потомками первой волны миграции являются те коренные жители Америки, которые говорят на многочисленных языках группы, значащейся в классификации Гринберга под именем Америнд. Вторая волна, на-дене, принесла с собой конгломерат языков, распространенных ныне в северо-западном регионе страны, включая побережье. К языкам этой семьи относятся такие, как атапаскский, хайда и тлингит. Третья волна миграции, в свою очередь, принесла с собой языки, распространенные ныне на Алеутских островах и в Арктике — то есть эскимосско(инуитско)-алеутскую группу языков. Когда же встает вопрос о хронологии, гипотеза Гринберга проявляет свою консервативность, полагая, что три языковые группы были привнесены на территорию Америки соответственно 11, 9 и 4 тысячи лет тому назад. Ясная, но упрощенная концепция Гринберга, основанная на сведении сложных и многообразным форм к простейшим, построена на научных выводах генетиков, работающих над проблемами заселения Америки. Что же касается американских лингвистов, то в большинстве своем они почти единодушно отвергают метод Гринберга, основанный на «инстинкте овчарки», привыкшей собирать овец в стадо. Некоторые языковеды отстаивают концепцию множественности независимых волн миграции, в результате которых в Америке возникло существующее разнообразие языков и «стволов». Надо сказать и о ученых, которые проявляют осторожность и не идут дальше обсуждения целесообразности объединения языковых групп Америки в большие единства и не делают определенных выводов относительно того, насколько эти гипотезы соответствуют действительности[408] .
Сколько генетических линий-родоначальниц существует? Сколько было волн миграции?
В конце 80-х гг., когда генетические коды, занесенные в Америку волнами мигрантов, были в значительной степени изучены, разногласия между лингвистами и генетиками, вместо того чтобы смягчиться, разрослись и превратились в пропасть непонимания. Отвергая концепцию множественности миграций, генетики полны решимости разрушить гипотезу Гринберга, основанную на магической власти числа «три», и уменьшить количество миграционных волн до двух, а затем и до одной волны. Генетики стремятся отмежеваться не только от Гринберга, но и от археологов, строя свою концепцию на предположении, что возраст первой волны миграции равен 50 тысячам лет. Столь значительный срок может заставить застыть от изумления как археологов (даже тех, что относят заселение Америки к периоду, предшествующему эпохе Кловис), так и лингвистов (в том числе специализирующихся на изучении глубокой древности). Согласовать данные разных дисциплин в пределах единой концепции оказалось задачей не из простых, тем более что каждая из наук заводит свою песнь. Если бы только можно было исправить положение, предоставив всем по концертному залу. Поскольку мы хотим выяснить, почему голоса поющих разошлись и перестали звучать в унисон, нам необходимо понять, в каком направлении начиная с 1990 г. происходило исследование генетического кода американских аборигенов. В 1991 г. новозеландский генетик Рик Уорд выявил небольшое число типов митохондриевой ДНК у представителей племени нуу-чах-нулт (племя, говорящее на одном из языков американских индейцев и обитающее на северо-западе Америки) и идентифицировал 4 комплекса, или линии. По подсчетам Уорда, эти линии превратились в одну общую примерно 41—78 тысяч лет тому назад. С присущей ему проницательностью Уорд правильно определил, что это — первоначальная ветвь, по всей видимости, существовала в Азии задолго до прихода в Америку. Это означает, что в Америку проникли уже множество отдельных линий. Предложенная Уордом трактовка четырех линий-родоначальниц положила начало спорам и дискуссиям среди генетиков о количестве миграций, и споры эти не умолкают и по сей день[409]. В 1993 г. японский генетик Сантоши Хораи высказал предположение, что четыре эти родословные линии могут означать четыре миграции, имевшие место в древности[410]. Подобная симметричная точка зрения — «одна линия соответствует одной миграции» — рассматривалась многими как излишне упрощенная интерпретация эффектов линий-родоначальниц и генетического многообразия американских этнических групп, однако подобную трактовку не следует считать последним словом в этой области. Сегодня другие генетики уже высказывают мнение, что даже три волны миграции — это слишком много.
Генетическая география Северной и Южной Америки
Американские и итальянские генетики Дуглас Уоллэс, Антонио Торрони и их коллеги на основании исследований ранних мтДНК (митохондриевых ДНК) пришли к выводу, что американские индейцы заселили Америку 20 тысяч лет тому назад, в то время как мигранты, говорящие на языках группы на-дене, пришли на северо-западное побережье Аляски значительно позже — примерно 6—10 тысяч лет тому назад. В следующем (1993) году Торрони и Уоллэс продолжили работу над выяснением типологии мтДНК у первобытных жителей Америки. Они сделали тщательный анализ ДНК 527 американских аборигенов, представляющих 24 этнические группы (то есть все американские группы, за исключением эскимосской (инуитской); 404 жителей Сибири, представляющих 10 этнических групп, а также 106 жителей Восточной Азии[411]. Так была сделана первая попытка сформулировать правила «генной географии», позволяющие выявить исходные «генетические типы кланов-родоначальников». Торрони и Уоллэс заявляют, что: 1. Каждый из исходных типов мтДНК, характеризующих кланы родоначальников, должен стать корнем, который произведет на свет собственную ветвь; все типы, являющиеся производными от этой ветви, в конечном счете произошли от ДНК кланов родоначальников. 2. Должна сохраняться возможность обнаружить типы мтДНК кланов родоначальников на прародине мигрантов, то есть в Восточной Азии. 3. Дочерние или производные мтДНК кланов-родоначальников, ставшие основополагающими для американских этнических групп, должны встречаться только в Америке, в Азии их не может быть. Торрони и Уоллэс идентифицировали четыре основные клана родоначальников Америки. По всей вероятности, каждый из этих кланов возник на основе единственного типа мтДНК. Каждый клан (представлявший собой строго гомогенную группу), обозначим их буквами от А до D, скрещивался с тремя другими кланами. На севере, в зоне распространения языков группы на-дене, представлен лишь один клан — А. Кроме того, занимаясь идентификацией мутационных типов кланов родоначальников, нужно иметь в виду, что кланы-сестры могут быть обнаружены и по другую сторону Берингова пролива, и в Восточной Азии. (В 4 и 5 главах мы уже упоминали четыре клана Азии, в дальнейшем об этом будет рассказано более подробно.) Эти четыре генетические группы-родоначальницы встречаются как в Америке, так и в Восточной Азии, однако американские дочерние типы, сложившиеся на их основе, в Азии не встречаются. По всей вероятности, они сформировались уже в Америке. Итак, поскольку эти находки прекрасно согласуются с тремя сформулированными выше правилами, можно предположить, что четыре клана, переправившиеся из Азии и ставшие родоначальниками жителей Америки, уже в те древние времена обладали идентификационными признаками групп А, В, С и D[412]. (Рис. 7.3.) Торрони и Уоллэс считают также, что конкретные типы мтДНК определяют специфику и типические особенности конкретных групп американских аборигенов. Иначе говоря, уже в глубокой древности племена существовали в изоляции друг от друга, и перетекание генной информации между ними было незначительным. За исключением группы В, все другие группы-родоначальницы представлены также на просторах Сибири, что наводит на мысль, не является ли этот регион источником миграционных волн, или из этого следует то, что у населения Сибири и Америки была общая прародина. Когда была разработана типология групп-родоначальников, ученые получили возможность использовать молекулярные часы для определения даты возникновения каждой из этих групп. Результаты были просто ошеломляющие: группы А, С и D оказались очень древними (период их пребывания в Америке составляет 20—41 тысячу лет), словом, они появились задолго до эпохи Кловис, в то время как появление группы В как раз и можно отнести к названной эпохе. Получив эти данные, Торрони и Уоллэс удостоверились в том, что заселение Америки произошло задолго до эпохи Кловис. Лишь вторжение группы В относится к более позднему периоду. Ученые предположили, что группы на-дене и эскимосско(инуитско)-алеутская появились в Америке независимо друг от друга. И хотя ученые были весьма осторожны в определении времен и сроков и не ставили перед собой задачи уточнить количество волн миграций, у них сложилось впечатление, что волн было три[413]. Научные изыскания Торрони и Уоллэса, кажется, хорошо согласуются с концепцией трех волн Гринберга. Осталось лишь согласовать хронологические таблицы. Группа В — самая молодая. На просторах Северной Америки она появилась независимо от других групп и распространилась лишь в послеледниковый период, то есть в эпоху Кловис[414]. Названный на основании показаний молекулярных часов возраст колонизации Америки просто громаден, он подтверждается лишь минимальным количеством артефактов, и ничтожно малое число археологов станет на его защиту, но даже самые «молодые» даты, вычисленные по этой методике, значительно старше эпохи Кловис. В целом, полученные генетиками новые данные едва ли могут быть согласованы с научными выводами археологов и лингвистов, взгляды ученых расходятся все дальше.
 Рис.7.3
При более серьезном и тщательном исследовании на основании генетических датировок были определены еще более грандиозные сроки. Среди современных американских индейцев удалось выявить представителей генетической группы Б, в результате последовательной переоценки ее возраста было установлено, что даже эта группа появилась скорее в период, предшествующий эпохе Кловис, нежели в последующие за ней времена. Эти научные выводы были опубликованы уже после того, как Торрони и Уоллэс в 1993 г. получили генетические датировки, возводящие к столь же глубокой древности формирование трех других групп: А, С и D. Итак, в перспективе генетика будет все более настойчиво заявлять о том, что основные линии мтДНК проникли в Америку еще перед Последним ледниковым максимумом. Те из ученых, которые относят период заселения Америки к наиболее древним временам, а именно к периоду предпоследнего ледникового периода, находят возможным оценить генетический возраст Америки в 30 или даже в 40 тысяч лет[415].
На вопрос, что же активнее всего повлияло на наши представления о заселении Америки, можно ответить так более точные представления о том, каковы же были генетические ответвления от линий-родоначальниц, а также развитие методологии определения возраста этих ответвлений. В результате обширных и дорогостоящих исследований генов и закодированных в генах процессов, которые шли в доисторические времена в Америке, на свет появились сравнимые с откровением научные теории и новаторские методики, которые открыли перед учеными новые перспективы в исследовании Африки, Азии и Европы. Разворачивавшиеся на этих континентах доисторические события оказались значительно более сложными для научного анализа. Корпус научных знаний, касающийся обсуждаемых в этой книге древнейших миграционных процессов, шедших в этих регионах, весьма велик.
Рис.7.3
При более серьезном и тщательном исследовании на основании генетических датировок были определены еще более грандиозные сроки. Среди современных американских индейцев удалось выявить представителей генетической группы Б, в результате последовательной переоценки ее возраста было установлено, что даже эта группа появилась скорее в период, предшествующий эпохе Кловис, нежели в последующие за ней времена. Эти научные выводы были опубликованы уже после того, как Торрони и Уоллэс в 1993 г. получили генетические датировки, возводящие к столь же глубокой древности формирование трех других групп: А, С и D. Итак, в перспективе генетика будет все более настойчиво заявлять о том, что основные линии мтДНК проникли в Америку еще перед Последним ледниковым максимумом. Те из ученых, которые относят период заселения Америки к наиболее древним временам, а именно к периоду предпоследнего ледникового периода, находят возможным оценить генетический возраст Америки в 30 или даже в 40 тысяч лет[415].
На вопрос, что же активнее всего повлияло на наши представления о заселении Америки, можно ответить так более точные представления о том, каковы же были генетические ответвления от линий-родоначальниц, а также развитие методологии определения возраста этих ответвлений. В результате обширных и дорогостоящих исследований генов и закодированных в генах процессов, которые шли в доисторические времена в Америке, на свет появились сравнимые с откровением научные теории и новаторские методики, которые открыли перед учеными новые перспективы в исследовании Африки, Азии и Европы. Разворачивавшиеся на этих континентах доисторические события оказались значительно более сложными для научного анализа. Корпус научных знаний, касающийся обсуждаемых в этой книге древнейших миграционных процессов, шедших в этих регионах, весьма велик.
Оказалось, что три выделенные Гринбергом группы связаны друг с другом в пределах Америки родственными узами
Когда ученые привлекли к исследованиям большее число генетических маркеров, выяснилось, что внутренняя структура распространенных в Америке ветвей мтДНК свидетельствует о существующих между носителями языков на-дене, эскимосами (инуитами) и американскими индейцами генетических связях, которые не были обнаружены Торрони и Уоллэсом во время прежних научных изысканий. Ученые сперва полагали, что для носителей языковых групп на-дене и эскимосско(инуитско)-алеутской характерны только маркеры группы А, что было принято объяснять общностью предков, распространившихся на берегах Америки под влиянием единой миграционной волны. Однако вскоре стало очевидно, что для эскимосско(инуитско)-алеутской группы свойственны как маркеры группы А, так и D. Эта группа D стала восприниматься как новая подгруппа азиатского типа D, которая прежде не была зафиксирована ни в Азии, ни в Америке, теперь ее принято обозначать — D2. D1 — наиболее распространенный в Америке тип линии-родоначальницы D — отчасти связан с типом D2, но встречается только среди американских индейцев[416]. Существуют и значительно более убедительные доказательства существования общих предков у носителей языков, относящихся к эскимосско(инуитско)-алеутской группе и к на-дене, с американскими индейцами. В основе этих доказательств лежит следующее открытие: в основе американской группы А, оказывается, лежат уникальные линии-родоначальницы А1 и ее дочерний тип А2 (а также и другие дочерние типы). А1 принадлежит к первому дочернему поколению коренного азиатского типа А. Тип А2 является дочерним по отношению к А1 и отличается от последнего единственной мутацией. А1 характерен только для группы, названной Гринбергом «Америнд», в то время как А2 обнаружен у представителей всех трех языковых семей, но при этом он является единственно возможным для носителей языков группы на-дене и эскимосско(инуитско)-алеутской. Эти сложные взаимосвязи, характеризующиеся включением или исключением отдельных линий-родоначальниц, доносят до нас одну очень простую весть: они соединяют три языковые группы Гринберга в единую семью, в основании которой лежит один общий генетический источник — американская линия-родоначальница А. При этом ни А1, ни А2 не были найдены в Азии, но их можно встретить в Сибири у местных эскимосов (инуитов) и чукчей. Язык чукчей относится к чукотско-камчатской семье языков. (Эту изолированную языковую семью объединяют с некоторыми другими семьями под общим названием палеосибирских (палеоазиатских) языков, которые распространены на дальневосточной оконечности Сибири и на полуострове Камчатка)[417] Эти два сибирских народа входят в уникальную подгруппу А2, к которой также относятся все прочие эскимосы (инуиты), живущие в Америке и Гренландии; таким образом, их языковая группа хранит свои семейные узы, подобно другим родственникам из клана А. Эти данные указывают на то, что Новый Свет был заселен в результате единой волны миграции, которая прокатилась по обеим Америкам. Подобные семейные узы связывают и другую, распространенную по всей Америке подгруппу, которая сохранила преемственность от линии-родоначальницы D D2 является отличительным признаком эскимосско(инуитско)-алеутской группы А такой ее редкий родственник, как D1, встречается почти исключительно среди американских индейцев, которые составляют большинство американских аборигенов[418]. Эти сделанные генетиками открытия укрепили ученых во мнении, что все три группы американских аборигенов (арктическая, субарктическая и более южная группа, названная Гринбергом Америнд) связаны между собой прочными родственными узами. Кажется, есть все основания вернуться к высказанной некогда идее о том, что была лишь одна волна миграции, но и эта концепция оставляет без ответа многие каверзные вопросы. Например, следующий: наиболее убедительным доказательством существования родственных уз, соединяющих представителей трех языковых групп, является тот факт, что они унаследовали общие для их семьи генетические типы А1 и А2, которые, хотя и возникли уже после исхода из Азии, свидетельствуют о событиях глубочайшей древности, к которой только и можно отнести разделение этих народов. В результате произошедшего раскола народы Арктики и Субарктики отделились от той части американцев, которые заселили более южные земли. Но как же тогда объяснить тот факт, что у двух подгрупп — на-дене и эскимосско(инуитско)алеутской — тип А2 значительно моложе, чем у остальной части американцев?
Почему север генетически однообразен? Должно быть, в этом виноват Ледниковый период?
Ученые, стремящиеся связать колонизацию американской Арктики и Субарктики с общим процессом заселения обеих Америк, которое они считают результатом единственной волны миграции из Азии, постоянно сталкиваются с таким трудно объяснимым феноменом, как неравномерное распределение кланов родоначальников. На основе закономерностей, выявленных в результате анализа этих кланов и эволюционных изменений большого пальца, можно вывести следующее правило: чем ближе мы находимся к региону, ставшему источником миграции, тем больше у нас шансов обнаружить представителей всех кланов родоначальников. И наоборот — чем дальше мы уйдем от стартовой точки миграции, тем беднее в этническом отношении будет регион, в котором мы оказались. В высоких широтах, на северо-востоке Аляски, там, откуда началось заселение Америки, можно найти лишь одну линию-родоначальницу — А, представленную ее дериватом — типом А2. На всей территории Арктики и Субарктики линии В и С полностью утрачены. Напротив, в прочих регионах Америки широко распространены все четыре клана-родоначальника, от А до D, и каждый из них представлен большим количеством разнообразных дериватов. Все это кажется нагромождением нелепиц. Как могло так случиться, что все типы — А, В, С и D — широко представлены в большинстве регионов Северной и Южной Америки, в то время как на Аляске, служившей вратами миграции, и на прилегающих к ней землях Канады выявлен лишь более молодой тип клана-родоначальника А2. Генетическое разнообразие, характеризующее южную часть Северной Америки, и еще более ярко выраженное разнообразие Южной Америки, и при этом генетическая бедность Крайнего Севера (Канады и Аляски) соответствует такому же перевернутому принципу распределения языков. Если бы концепция «Кловис Первый» соответствовала действительности и заселение Америки относилось бы ко временам после ПЛМ, то картина была бы обратной. Но если Америка была первоначально заселена во времена, предшествующие Ледниковому периоду, то мы имеем дело с феноменом, о котором я уже говорил, когда касался проблем языка: во время ПЛМ обширные пространства Северной Америки обезлюдели. Если вторичное заселение Америки произошло в период «великого таяния», то неудивительно, что народы севера характеризуются меньшим лингвистическим и генетическим разнообразием и что, как следствие этого, мы можем утверждать, что срок их жизни на этих территориях не столь велик, чем у более южных народов. Высказанная еще в 1993 г. идея заселения Северной Америки после Ледникового периода прекрасно объясняет перевернутый принцип распределения генетического материала[419]. Уроженец Аляски генетик Джералд Шилд и его коллеги сделали следующее открытие: оказалось, что в отличие от населения, распространенного на юге Азии и обеих Америк, народы, живущие за Северным полярным кругом этих континентов, с точки зрения генетического возраста можно признать ровесниками, причем одинаково молодыми. О чем свидетельствует то, что севернее 55-й параллели группа В, в частности, не представлена совершенно. Ученые доказали, что за Северным полярным кругом население появилось в результате относительно недавнего вторжения на эти территории одного северного народа, бедного в генетическом отношении. Им также удалось аргументированно обосновать, что это вторжение произошло уже после того, как оба американских континента были заселены людьми, одаренными большим генетическим разнообразием. Подобная точка зрения позволяет убедительно объяснить генетическую и лингвистическую загадку этих регионов, связанную с тем, что после Ледникового периода некогда уже обжитые земли Америки, точнее, Канада и Аляска, были заселены людьми, обладавшими бедным генофондом.
Приют на погибшем континенте Берингия
Однако остается еще неясным, откуда пришел этот народ, приспособившийся к жизни на Крайнем Севере. Был ли он выходцем из Азии — или из Америки? Современные генетические данные, свидетельствующие о взаимосвязи между А1 и А2, указывают скорее на Америку, в то время как присутствие в генах эскимосов и чукчей Сибири типа А2 наводит на мысль о том, что исходный пункт миграции мог располагаться в Азии. Но как бы там ни было, неясным остается вопрос о том, как возникло различие между генетическим возрастом носителей языков на-дене и эскимосско(инуитско)-алеутской группы, с одной стороны, и всеми прочими жителями Америки. С другой стороны, если предположить, что все они происходят от единого генетического ствола, получается, что генетический, физический и лингвистический раскол между этими народами относится к глубокой древности. В 1996 г. генетик Питер Форстер с интернациональной группой своих коллег прояснил проблему и распрямил все хитрые завитки ее противоречий. Проблема была сродни шахматной задаче: большое число людей билось над ее решением, но стоило догадаться, в чем суть, — решение оказалось на удивление простым. Среди сотрудничавших с Форстером ученых были и Антонио Торрони, и немецкий математик и энциклопедист Ханс-Юрген Бандельт, который был создателем и вдохновителем исследовательского проекта, ставившего перед собой цель построить генеалогическое древо генов, — которым так часто мне приходилось пользоваться в процессе написания этой книги. Разрешение американской загадки оказалось весьма простым: родиной северного народа, покинувшего в Ледниковый период ее просторы, была не Азия и не Америка, но совсем иной континент — Берингия[420]. (Рис. 7.4.) Между 11 и 25 тысячами лет тому назад уровень моря был настолько низок, что на месте Берингова пролива была суша, служившая мостом между Азией и Северной Америкой. Но Берингия занимала большепространства, чем Берингов пролив: это был огромный континент в полном смысле этого слова, его максимальная площадь достигала 1,3 млн. кв. километров (500 тысяч кв. миль). На нем не только не было ледяного покрова — но его просторы были заняты травянистой тундрой, способной прокормить стада травоядных млекопитающих. Летом, конечно, бывало холоднее, чем сейчас, зато зимы, как это ни парадоксально, были мягче. Почти в течение всего периода существования континентального моста ледниковый покров сковывал земли, лежащие значительно южней. И, как мы уже говорили, между 15 и 22 тысячами лет тому назад ледовый коридор был закрыт, что, видимо, препятствовало контакту между Крайним Севером и остальной частью Америки. В те времена охотничьи угодья Сибири превратились в ледяную пустыню и едва ли могли привлечь больше интереса, чем Северная Америка. Таким образом, Берингия и западная часть Аляски, будучи отрезанными от Азии и Америки, стали для многих народов прибежищем, которое помогло пережить Ледниковый период. Поскольку в Берингии уцелели лишь некоторые характерные для Америки типы генов-родоначальников, в группах на-дене и эскимосско(инуитско)-алеутской мы наблюдаем малое генетическое разнообразие. И хотя эти народы были связаны с Америкой посредством таких групп генного древа, как А1 и А2, нас не может не поражать то, как сильно на генетическом уровне они отличаются от остальной Америки[421].
 Рис. 7.4
Рис. 7.4
Согласуются ли новые датировки с предположением, что заселение континента произошло в доледниковый период?
Дальше — больше. Авторитетная комиссия признала, что анализ мтДНК свидетельствует о том, что известный набор митохондриевых линий-родоначальниц появился на территории Берингии перед Последним ледниковым максимумом и затем распространился по обеим Америкам. Затем начался Ледниковый период, отделивший Канаду и Аляску от остальной части Америки, которая осталась заселена лишь в южных областях. В это время голод, мучительные лишения и почти полное вымирание многих родов серьезно подорвали генетическое разнообразие народов Крайнего Севера, из последних сил цеплявшихся за жизнь, теплившуюся на землях Берингии во время Ледникового периода. Итак, о чем же нам поведали генетические датировки? Согласуются ли они с теорией первой волны заселения Америки перед Ледниковым периодом или же второй — после него. Ответим: согласуются, и на удивление хорошо. Форстер и его коллеги вычислили возраст линий A1, A2, В и С у народов, населяющих территории, лежавшие к югу от ледников, а также возраст линии D1 — для американских индейцев из Северной, Центральной и Южной Америки. Анализ показал, что генетический возраст десяти из двенадцати групп старше эпохи Кловис. Стремясь добиться большей точности расчетов, ученые вычислили средний арифметический возраст для всех четырех групп (А — D), распределив их по трем регионам (рис. 7.5). Оказалось, что генетический возраст индейцев Северной Америки составляет 23 тысячи лет, Центральной Америки —16 тысяч лет, а Южной Америки — 21 тысячу лет. Следовательно, возраст линий-родоначальниц A1, A2, В, С и D1 свидетельствует о том, что они появились в Америке за 21—22 тысячи лет тому назад, то есть во времена, когда ледниковый коридор был еще закрыт. Когда же Форстер и его коллеги приступили к исследованию дочернего типа А2 у группы на-дене и эскимосско(инуитско)-алеутской, то оказалось, что ее возраст, соответствующий периоду заселения этих территорий, составляет 11 300 лет, что свидетельствует о вторичном заселении этих земель в эпоху Кловис. Но результаты этих исследований можно интерпретировать поиному: дело не только в том, что культура, да и сам народ эскимосов (инуитов) является относительно молодым феноменом, но и в том, что свои гены эскимосы преимущественно унаследовали от древних родоначальников Америки[422]. Бьюсь об заклад, что результаты, полученные Форстером и его коллегами, имеют просто ни с чем не сравнимое значение для выяснения датировок, касающихся запутанной истории заселения Америки. Их данные, полученные в том числе и в результате генного анализа, впервые со всей ясностью подтвердили то, что Северная Америка была вторично заселена племенами, вышедшими с континента Берингия, — надо сказать, что археологи высказали эту идею еще раньше.
 Рис. 7.5
Итак, подведем итоги.
Во-первых, получено подтверждение того, что основные распространенные в Америке четыре родоначальных типа мтДНК произошли от четырех азиатских групп А — D, идентифицированных в 1993 г. Антонио Торрони и группой его коллег: А1 (и ее производная А2), В, С и D1.
Во-вторых, четыре родоначальных типа появились в Америке, по всей вероятности, до эпохи Кловис и даже до Последнего ледникового максимума, случившегося 18 тысяч лет тому назад.
В-третьих, родиной и исходным пунктом, давшим импульс для движения и распространения этих четырех кланов-родоначальников по Америке, является отнюдь не Азия, а Берингия.
Последний пункт может у кого-то вызывать ассоциации с дровосеком, стремящимся расщепить волоски (или континенты), — что ни говори, а народы Берингии в конечном счете могли быть выходцами из Азии, — но это не самая правильная точка зрения. Как в результате доказали Форстер и его коллеги, Берингия откололась от Азии и перестала подпитываться ее генетическим материалом еще до начала Ледникового периода — может быть, за целое тысячелетие до него. Вскоре станет понятно, почему мы придаем такое значение разграничению этих континентов. Берингия служила для Америки источником линий-родоначальниц как перед Последним ледниковым максимумом, так и во время вторичного заселения Арктики и субарктических областей народами на-дене и эскимосско(инуитско)-алеутской группой. Итак, после Ледникового периода произошло вторичное заселение Америки выходцами из Берингии, которые, вероятно, были носителями единственной линии — А2, поскольку Ледниковый период поставил обитателей севера на грань выживания и привел к формированию так называемого генетического сужения, выразившегося в том, что выжить смогла лишь одна генетическая линия
Остальное, в сущности, происходило по прежнему сценарию, впрочем — с некоторыми вариациями на старую, как мир, тему миграций, в результате которых на обоих американских континентах перед последним Ледниковым периодом появились если не все, то большинство линийродоначальниц[423].
Рис. 7.5
Итак, подведем итоги.
Во-первых, получено подтверждение того, что основные распространенные в Америке четыре родоначальных типа мтДНК произошли от четырех азиатских групп А — D, идентифицированных в 1993 г. Антонио Торрони и группой его коллег: А1 (и ее производная А2), В, С и D1.
Во-вторых, четыре родоначальных типа появились в Америке, по всей вероятности, до эпохи Кловис и даже до Последнего ледникового максимума, случившегося 18 тысяч лет тому назад.
В-третьих, родиной и исходным пунктом, давшим импульс для движения и распространения этих четырех кланов-родоначальников по Америке, является отнюдь не Азия, а Берингия.
Последний пункт может у кого-то вызывать ассоциации с дровосеком, стремящимся расщепить волоски (или континенты), — что ни говори, а народы Берингии в конечном счете могли быть выходцами из Азии, — но это не самая правильная точка зрения. Как в результате доказали Форстер и его коллеги, Берингия откололась от Азии и перестала подпитываться ее генетическим материалом еще до начала Ледникового периода — может быть, за целое тысячелетие до него. Вскоре станет понятно, почему мы придаем такое значение разграничению этих континентов. Берингия служила для Америки источником линий-родоначальниц как перед Последним ледниковым максимумом, так и во время вторичного заселения Арктики и субарктических областей народами на-дене и эскимосско(инуитско)-алеутской группой. Итак, после Ледникового периода произошло вторичное заселение Америки выходцами из Берингии, которые, вероятно, были носителями единственной линии — А2, поскольку Ледниковый период поставил обитателей севера на грань выживания и привел к формированию так называемого генетического сужения, выразившегося в том, что выжить смогла лишь одна генетическая линия
Остальное, в сущности, происходило по прежнему сценарию, впрочем — с некоторыми вариациями на старую, как мир, тему миграций, в результате которых на обоих американских континентах перед последним Ледниковым периодом появились если не все, то большинство линийродоначальниц[423].
Линия X
Прежде чем обратиться к фактам, позволяющим разрешить вопрос, сколько же всего было в доледниковый период миграционных волн — одна или несколько, — постараемся повнимательнее изучить еще одну материнскую линию-родоначальницу. Пятая линия-родоначальница — персона весьма загадочная. Она лишь в последние годы предстала взорам ученых. Поэтому и название для этой «сверхштатной» линии-родоначальницы было выбрано соответствующее — линия X. В 1991 г. Рик Уорд выдвинул идею четырех генных кластеров. Через два года и независимо от него Торрони и его коллеги самостоятельно идентифицировали группы А, В и С, то есть подтвердили существование трех из четырех открытых Уордом кластеров и частично свели оставшиеся типы в новую группу D, а остатки четвертого генного кластера Уорда попросту сбросили со счета: им отвели место в корзине для хлама с невразумительным названием «остальное». Спустя еще пару лет многие исследовательские группы обратились к этому «остальному», стремясь выяснить, чем же оно все-таки является. Уорд сразу предположил, что обнаружен еще один кластер генов, объединенных единством происхождения, — иными словами, пятая американская линия-родоначальница. Среди ученых, открывших эту линию, была аргентинский генетик Грациела Байльет с группой сотрудников, а также математик Ханс Бандельт с коллегами. Именно они, опираясь на свой новый метод построения генеалогического древа генов, заговорили о возможном существовании пятой линии-родоначальницы. В результате проведенного в 1996 г. повторного анализа американских линий-родоначальниц Форстер выявил новый американский клан и дал ему новое название — европейская группа X. В результате было убедительно доказано, что пятая американская линия-родоначальница и обнаруженная Торрони в том же году европейская группа X восходят к общему предку — Х[424]. Американская группа X распространена по преимуществу в двух отдельных культурных регионах американских индейцев, живущих компактно на севере страны — примерно по 50-й параллели. Первая из этих культурных групп проживает на северо-западном побережье Тихого океана в г. Йакима, штат Вашингтон. К этой группе относится также рыболовецкая культура племени нуу-чах-нулт, обосновавшемся на западном побережье острова Ванкувер (а также на полуострове Олимпия, штат Вашингтон). Возраст последней культуры — 4000 лет. В этих культурных группах на тип X приходится соответственно 5 и 11 — 13%. В другую северную группу, носительницу линии X, входят племена сиузов и оджибва, живущих в восточных лесах на западной границе Великих озер. В последнем из названных племен линия X представлена у 25% населения[425]. Мог ли тип X проникнуть в гены американских аборигенов из набора материнских линий-родоначальниц европейцев, которые еще совсем недавно вторглись своей многонациональной массой на эти территории. Вероятность этого крайне ничтожна. Дело в том, что тип X был представлен в генах народов, живущих в этих регионах, еще в доколумбовскую эпоху. Уорд провел оригинальные исследования, в результате которых возможность сколько-нибудь значительного проникновения генного материала европейцев в гены племени нуу-чах-нулт была практически исключена. Что же касается американских аборигенов, живущих в восточных лесах, непосредственное свидетельство присутствия в их генах линии X в доколумбовскую эпоху было получено в результате анализа древних мтДНК у доисторического народа онеота из долины реки Иллинойс[426].
Кенневикский человек: связи с доколумбовской Европой
Оказалось, что проверка на трансатлантическую совместимость ДНК европейцев и американских аборигенов подлила масла в огонь и породила бесконечные и бессмысленные споры о том, кто первым достиг Америки. Вопрос о сходстве скелетов древних американцев и европейцев застрял как кость в горле спорящих. Ученые уже не одно десятилетие изучают черепа древних обитателей Америки, датируемые 5—11 тысячами лет тому назад. Однако в результате столь длительного изучения стало известным лишь то, что древние черепа совсем не похожи на те, что принадлежали американцам последующих эпох, но почему эти черепа так сильно отличаются друг от друга, остается до сих пор неясным. В 1996 г. (в том самом, когда ученые дали линии-родоначальнице X ее настоящее имя и установили ее родственные связи с Европой) на реке Колумбия в Кенневике, штат Вашингтон, два молодых человека нашли череп. Коронер[427] пригласил Джеймса Чаттерса, американского антрополога, специалиста в области реконструкции облика человека, предлагая ему по возможности восстановить скелет. Распространенный в этих местах 4500—8500 лет тому назад каменный наконечник копья застрял в костях глаза. Очевидная древность каменного острия, имеющего форму зубчатого листа, навела коронера на мысль подвергнуть находку радиоуглеродному и ДНК-анализ[428]. С самого начала было очевидно, что найденный череп не типичен для современных аборигенов Америки. От этого человека, умершего за сорок — сорок пять веков до нас, остался длинный узкий череп. Видимо, и лицо его было узким, а подбородок — выступающим вперед. Вопреки нашим ожиданиям, он выглядел скорее как европеец и вовсе не напоминал людей с широкими лицами и головами. Выполненная Джеймсом Чаттерсом реконструкция головы этого человека (см. Фото 21) имеет поразительное сходство с Патриком Стюартом из знаменитого фильма «Звездный путь». Впрочем, возможно, подобное впечатление возникает благодаря смелому подходу к реконструкции.
 Фото 21. Джеймс Чаттерс со своей реконструкцией и реальным черепом Кенневикского человека. Ученый подчеркивает загадочный, выдающийся вперед подбородок и, возможно, кавказоидные черты лица.
Фото 21. Джеймс Чаттерс со своей реконструкцией и реальным черепом Кенневикского человека. Ученый подчеркивает загадочный, выдающийся вперед подбородок и, возможно, кавказоидные черты лица.
Проблема заключается в том, что кенневикский человек не принадлежит к кавказоидной расе. Есть, конечно, такие черты его внешности, которые придают ему сходство с кавказоидами, но есть и другие черты, свидетельствующие о его азиатском происхождении. Кенневикский человек выглядит как европеец благодаря особым пропорциям черепа — кажется, его сплющили сбоку прессом, отчего черты лица стали нарочито вытянутыми. Расисты склонны интерпретировать эту находку как доказательство того, что заселение Америки представителями доминирующей в мире белой расы произошло в очень глубокой древности. Зубы кенневикского человека относились к сундадонтному типу, что указывает на место происхождения подобной разновидности человека: это — юго-восточный регион Азии. Его глазные впадины не типичны ни для европейцев, ни для современных аборигенов Америки. В результате тщательного изучения всевозможных пропорций и соотношений между различными частями черепа ученые пришли к выводу, что кенневикский человек более всего похож на людей племени айну и на народы, населяющие южную часть Тихого океана, включая полинезийцев[429]. Но поразительнее всего было то, до какой степени кенневикский человек не похож ни на один из народов, заселявших Землю в течение последних 5 тысяч лет. Теперь мы представляем его жизнь во многих подробностях. Он многократно страдал от различных ранений и переломов костей. А рацион его питания изобиловал протеинами морского происхождения, например лососем. Радиоуглеродный анализ позволил установить возраст скелета, он составляет 8400 лет[430]. Корпус военных инженеров Соединенных Штатов объявил скелет своей собственностью. С тех пор различные коллегии адвокатов ведут из-за него тяжбу. Несмотря на всю эту юридическую неразбериху, уже многие лаборатории, специализирующиеся на анализе ДНК, предпринимали попытки извлечь ДНК из костных тканей. К сожалению, их деятельность не увенчалась успехом, отчасти это связано с техническими трудностями, а отчасти — с контаминацией древних структур современными молекулами ДНК. Если бы извлечение и анализ ДНК увенчались успехом, мы получили бы результаты, которые открыли бы нам много нового, но и тогда едва ли было бы возможным ответить на все поставленные вопросы. Хотя анализ митохондриевых ДНК позволяет нам проследить миграционные пути народов, не надо забывать, что мтДНК — это лишь фрагмент генетической памяти, передаваемой по наследству. Он не поведает нам ничего о внешности человека. Но часто он оказывается способным рассказать нам о его прародине, то есть о том, откуда родом предки человека. Предположим, что произошло нежелательное для нас открытие и незагрязненные инородными генными вторжениями молекулы мтДНК кенневикского человека указали бы на его принадлежность к одному из европейских кланов и генетическую близость к современным европейцам, тогда мы расценили бы полученные данные как свидетельство о непосредственном вторжении европейцев в Северную Америку. Но результаты анализа, скорее всего, подтвердили бы в общих чертах то, что перед нами коренной американец. Однако это, в свою очередь, не послужило бы доказательством отсутствия в Америке выходцев из Европы, которые могли бы прийти сюда более 9 тысяч лет тому назад. Пока удалось извлечь слишком малое количество ДНК, не содержащих инородных генных вторжений и контаминаций, поэтому так много проблем встает перед учеными, исследующими древние ДНК. Если неконтаминированное ДНК будет извлечено, то и в этом случае мы будем обладать лишь единственным образчиком, который едва ли сможет нам многое рассказать о целом народе и его истории. По этим причинам более информативными указываются крупномасштабные исследования мтДНК современных людей, нежели погоня же за единственным образчиком древней ДНК. Если предположить, что связь между кенневикским человеком и европейцами существовала, то окажется весьма показательным, что место, где были найдены его останки — на реке Колумбия, штат Вашингтон, — расположено поблизости от северо-западного побережья Америки, рядом с одним из двух регионов, где была обнаружена группа X, в частности — недалеко от народности, локализованной в городе Йакима.
Солютрийская гипотеза
Мысль о том, что европейцы обладали достаточными техническими знаниями для того, чтобы переправиться через Атлантику еще в доисторические времена, — не нова и широко обсуждалась задолго до того, как был найден кенневикский человек. Одна из наиболее известных теорий такого типа получила название Солютрийской гипотезы. Денис Стэнфорд, антрополог из Смитсоновского университета, полностью солидарен с ней[431]. Исторические свидетельства указывают, что, когда последний Ледниковый период достиг своего максимума или, возможно, чуть раньше, некоторые охотники, нашедшие убежище на юго-западе Европы, в области, граничащей с Францией и Испанией (см. главу 6), сели на лодки и покинули родину. Они отправлялись в плавание с европейского побережья Атлантического океана, затем огибали его с севера и направляли лодки в сторону восточного побережья Америки, где бывшие европейцы превращались в американцев эпохи Кловис. Культура людей, укрывшихся на юго-западе Европы от множества невзгод, на кои их обрек Последний ледниковый максимум, именуется солютрийской. Она более всего прославилась изысканными обоюдоострыми наконечниками. Солютрийские наконечники, как оказывается, относятся к числу довольно редких изобретений, они имеют поразительное сходство лишь с наконечниками эпохи Кловис и с их предшественниками, найденными в районе Кактус Хилл. Солютрийская теория находит свое подтверждение и в том, что для древних людей, создавших культуру по обеим сторонам Атлантики, характерен обычай загонять стада диких животных на самые вершины отвесных скал. Однако подобное сходство едва ли можно считать достаточным научным доказательством теории, тем более что аналогичная практика была известна даже в Китае людям типа, предшествующего современному. Считается также, что самая большая концентрация древнейших наконечников эпохи Кловис, которые чаще всего находят на юго-востоке США, указывает на иммигрантов, перебравшихся в Америку с другой стороны Атлантики. Но и в этом пункте теория сталкивается со множеством трудноразрешимых проблем, самой важной из которых, конечно, является вопрос о том, каким образом трансатлантическое плавание могло быть осуществлено на практике. Солютрийская культура пришла к берегам Америки раньше, чем успела сформироваться культура эпохи Кловис, а культура эпохи Кловис, как это убедительно доказал Вэнс Хейнс, могла сложиться лишь по окончании Последнего ледникового максимума. Это заставляет нас предположить, что наконечники солютрийского типа обогнули Атлантику с севера непосредственно во время Последнего ледникового максимума. При этом они должны были бы пройти вдоль окруженного ледниками побережья, в центре которого в наши дни расположен Нью-Йорк[432]. Какой бы заманчивой не выглядела теория, позволяющая называть европейцев древнейшей опорой обеих Америк, а вовсе не насильниками, осквернившими красоты Нового Света, как это принято считать в наше время, — приходится с сожалением признать, что историческая генетика не располагает данными, которые могли бы подтвердить истинность этой теории. Мишель Браун, генетик с кафедры медицины Эморийского университета в Атланте, приводит прямо противоположные данные. Доказав, что американская линия X может быть с полным правом признана самостоятельным пятым генным кластером Америки, Браун с группой своих коллег вычислил возраст этой линии-родоначальницы, которая должна была появиться в Новом Свете 23—36 тысяч лет тому назад, то есть задолго до событий, которые описывает Солютрийская гипотеза, а также значительно раньше любого из возможных вторжений послеледникового периода. Вероятно, линия X появилась в Америке ничуть не позже четырех других линий-родоначальниц. Дочерние типы пятого генного кластера X распространились по различным регионам Северной Америки: на Великих озерах и на западном побережье, — приблизительно в одно и то же время и в едином направлении, примерно вдоль 50-й параллели[433]. Нет оснований сомневаться в том, что у американской европейской линии X были общие предки. Но они могли отделиться от народа-прародителя уже 30 тысяч лет тому назад. Если бы линия X являлась относительно недавним, хотя и доколумбовским присадком европейского генного материала, то ученые обнаружили бы в генах американских аборигенов типичные для европейцев подгруппы кластера X или другие наиболее распространенные в современной Европе линии, тем более что X довольно редко встречается в Европе. Однако другие, типичные для европейцев кластеры в Америке обнаружены не были. Следовательно, американский тип X, скорее всего, был занесен из Азии тем же способом, что и четыре другие родоначальные мтДНК (см. рис. 7.4). На основании тех открытий, которым посвящена эта книга, был сделан документальный фильм «Истинная Ева», во время работы над которым с особой ясностью была явлена мощная способность мтДНК прослеживать древнейшие связи между народами и континентами. Продюсеры из множества предложенных выбрали наугад несколько образцов ДНК, извлеченных из тканей добровольцев, представляющих разнообразные этнические группы Америки. В своей лаборатории в Хаддерсфилде, Западный Йоркшир, Мартин Ричардс извлек ДНК и сделал их анализ. Ближе к концу фильма были вставлены кадры, на которых запечатлен рассказ участников эксперимента (совершенно добровольный) о результатах анализа своих генов. Редкое совпадение: выяснилось, что по типу мтДНК двоих участников — американского грека и коренного американца-аборигена из племени Кри — можно отнести к клану X. Узнав об этом, они на какой-то миг замерли, пристально вглядываясь в черты друг друга через всю лабораторию. Казалось, они потеряли дар речи от удивления, зато затем они смогли отдаться нахлынувшим чувствам, вызванным мыслью о существующей между ними древней связи.
Сколько было волн миграции и откуда они?
Перед генетиками стоит весьма трудный вопрос: откуда родом американская линия-родоначальница X, ведь в Азии она встречается весьма редко. Второй, связанный с первым, вопрос звучит так: какая часть Евразии является прародиной линии X? Эти вопросы — часть более крупной проблемы, а именно: проблемы формирования американского этноса. Ученым предстоит решить, в результате скольких волн миграции — одной или множества — была заселена Америка. Этот вопрос и должен привлечь наше внимание в первую очередь. Итак, перед тем как мы приступим к разрешению проблемы, каким образом линия X появилась в Европе и на обоих континентах Америки, мы должны вновь обратиться к фактам, которые помогут нам выяснить, сколько всего было волн миграции. Однако (даже если учесть, что генетиками были получены неопровержимые свидетельства о вторжении всех пяти линий-родоначальниц в Америку еще до начала Ледникового периода) разве можно считать, что я уже решил эту проблему на основании так называемой гипотезы «одна волна — одно вторичное заселение»? Конечно, нет. Мы обязаны учитывать, что погрешность, допустимая при вычислении генетического возраста, может исчисляться тысячами лет. И если использовать данные генетики для опровержения концепции «Первый Кловис» еще возможно, то доказать на основании тех же данных одновременность вторжения в Америку всех пяти линий-родоначальниц мы не можем. Пробегая мысленно историю генетических исследований в области заселения Америки, мы обнаруживаем отчетливую тенденцию к редукционизму, наметившуюся еще в 1990-е гг.: от четырех волн миграции мы пришли к трем, от трех — к двум и, наконец, к идее о единичном акте колонизации Америки и последовавшим за ним вторичным заселением теми же родоначальными типами. Этот минималистический подход не получил отражения в рационализации общего числа материнских линий-родоначальниц Америки: десять лет назад ученые наблюдали четыре линии, теперь же их насчитывается как минимум пять. Если обратиться к простой аналогии, то окажется, что пяти мтДНК цепочек, которые колонизировали оба континента Америки, так же много, как и пяти волн для единственной африканской линии мтДНК, которая покорила мир за пределами Африки. Итак, в прошлом появился либо большой клан-родоначальник, для которого был не свойствен генный дрейф, либо произошло множество вторжений. Мы не можем с полной уверенностью утверждать, что Америка была колонизирована каким-то одним кланом. И для этого есть свои причины. Во второй главе я доказывал, что исход из Африки был единым последовательным процессом, в основании которого лежали единственная отцовская и материнская африканские линии, от которых и произошло население всего остального мира Чтобы существующее генное разнообразие можно было свести к единой линии, нужно доказать, что клан-родоначальник переживал длительный период изоляции и генного дрейфа. Однако эта подходящая на все случаи жизни гипотеза не может с достаточной ясностью описать ситуацию в Америке. Возможно, конечно, что Северная и Южная Америка были колонизированы одной большой группой охотников-собирателей, но это не подтверждается никакими данными генетики, даже свидетельствами хромосомы Y. Хотя еще совсем недавно генетики Энн Стоун и Алан Стоункинг сами были сторонниками гипотезы «единственного» вторжения, теперь они по умолчанию признали слабость существующих свидетельств. Тремя строками выше того места, где постулируется «единая волна заселения», эти ученые обезоруживающе утверждают: «Вероятнее всего, люди, преследуя дичь, медленно брели мелкими группами через всю Берингию...»[434]. Но не все ли равно, что представляла собой единственная волна миграции: была ли она этнически единой или состояла из множества мелких групп? Думаю, что не все равно, ибо если клан переселенцев был однороден, то логично предположить существование лишь одного-единственного географического пункта его исхода. Идея единой волны заселения Америки в качестве рабочей гипотезы принята многими генетическими лабораториями. В поисках географического пункта исхода предков американских аборигенов генетики сделали один шаг вспять и обратили свое внимание на Азию. Будучи убеждены в том, что прародина должна находиться в Азии, они ищут азиатский регион, в котором сосредоточены пять американских линий-родоначальниц[435]. Как мы вскоре увидим, предположение о том, что все необходимые генные линии и даже технологии изготовления палеолитических орудий могли произойти из какой-то одной части Азии, кажется не только наивным, но и противоречащим другим фактам и наблюдениям. Итак, основываясь на данных генетики, Питер Форстер и его коллеги представили свою концепцию истории, и она была вовсе не о том, что прародину стоит искать в Азии. Они рассказали нам скорее об иной прародине — о частично затопленной в наши дни Берингии, которая, будучи своего рода естественным оазисом посреди льдов, простиравшимся между двумя великими континентами, оставалась заселенной в течение всего Ледникового периода. Именно Берингия приютила людей, принесенных к ее берегам миграционной волной, докатившейся сюда, видимо, с севера Азии, а затем сыграла роль подмостков, с которых отправились в путь мигранты следующих поколений. Климатические условия, окончательно сложившиеся на пике Последнего ледникового максимума, сделали Берингию заповедным уголком, который в результате природных катаклизмов оказался отрезанным сперва от Азии (арктической пустыней), а затем и от Америки (ледниками). Прежде чем были заселены оба американских континента и прежде чем ледниковые плиты окончательно преградили путь в Канаду, характерные для поселений Берингии материнские линии-родоначальницы претерпели изменения (генетический дрейф), в результате линий осталось пять. Именно в таком составе линии-родоначальницы отправились покорять Америку. Затем, во время Ледникового периода, но уже после заселения Америки, произошло дальнейшее сокращение числа линий-родоначальниц, в результате из пяти мтДНК осталась лишь одна группа А. В поисках прародины нам придется вернуться вспять — в Азию или в другие регионы, которые могли послужить источниками этих пяти линий. Однако прежде всего необходимо выяснить, существуют ли какие-либо определенные свидетельства того, сколько было миграций из Азии в Берингию: одна или много? Мы привыкли смотреть на проблему односторонне и считать первых жителей Америки не чем иным, как — да извинят меня читатели за такое термин — клонами единственного азиатского племени Причиной подобной односторонности является нежелание обращать внимание на какие-либо иные археологические находки, кроме кенневикского человека. Его немонголоидная внешность сокрушила многие наши стереотипы. Его облик характерен для южных островов Тихого океана и для племени айну. Мы, очевидно, никогда ничего не узнаем о цвете его кожи А волосы — какими они были темными, вьющимися или прямыми или какими-нибудь еще? Но его облик в корне отличался от внешности современных американских аборигенов. При этом среди древних жителей Северной Америки он был не единственным человеком, чей облик для нас столь непривычен. В наши дни стал широко известен и еще один древний американец, живший примерно 9400 лет тому назад — человек из пещеры Спирит Кейв с берегов озера Грейт-СолтЛейк Внешне он серьезно отличался от всех современных народов и американских аборигенов в том числе. Его тело было мумифицировано и волосы оставались черными до тех пор, пока эту уникальную находку не извлекли на свет, спровоцировавший их обесцвечивание и покраснение. Ученые полагают, что ближайшими родственниками этого человека являются люди племени айну, а также полинезийцы и австралийцы. В северных штатах обнаружены еще три атипичных палеоиндейца эпохи палеолита, женщина с речного порога Пеликан Рэпидз, штат Миннесота (возраст — 7800 лет, найдена в 1938 г), мужчина из долины Браунз-Вэлли (также штат Миннесота, возраст которого — 8900 лет) и бульская женщина из штата Айдахо (ее возраст — 10 800 лет). Однако стоит помнить, что палеоиндейцев Северной Америки, сравнимых с раритетными винами, не всегда возможно по внешнему виду отличить от современных аборигенов Америки. В качестве примера приведем человека из Уайзард Бич, жившего 9200—9500 лет тому назад. Его трудно отличить от американских аборигенов наших дней[436]. У Южной Америки есть все основания требовать, чтобы и ее признали континентом, заселение которого относится к глубокой древности, и едва ли она намерена промолчать в ответ на заявление о «нелепости» предположения, что еще в эпоху палеолита на этих землях жили предки современных южноамериканцев. Хотя прошло немало времени с тех пор, как ученые получили свидетельства об их существовании, широкие круги общественности совсем недавно узнали о дошедших до нас из глубины времен скелетах жителей Южной Америки. Как и в случае с кенневикским человеком, средства массовой информации не упустили возможности распространить сенсационные слухи и сомнительные комментарии, посвященные этому интереснейшему открытию. В августе 1999 г. Би-биси документально подтвердила, что Вальтер Невес, бразильский специалист в области эволюции человека, занят изучением так называемого негроидного человека из вымершего южноамериканского племени. В центре всеобщего внимания оказался череп двадцатидвухлетней женщины, родившейся более 11 500 лет тому назад. Ее условно назвали Люзия. Первоначально француз-археолог, обнаруживший ее останки в семидесятые годы XX в. в Минас Жераис, Бразилия, решил избавиться от нее как от ненужного хлама и подарил Национальному музею Бразилии. Судя по черепу, прародиной женщины могли быть Австралия, Меланезия или даже Африка. В 1999 г. антрополог и судмедэксперт Ричард Нев из Манчестерского университета реконструировал внешность Люзии и сделал ее скульптурный портрет[437]. Как бы рожденная заново, женщина оказалась удивительно похожей на современных представительниц коренного населения Америки (см. Фото 22). Признаемся, однако, что каждый из нас считает себя знатоком, способным помнить и узнавать множество лиц. Могут быть различные мнения относительно того, к какому типу внешности относится эта женщина — негроидному, австралийскому, меланезийскому или люйцзяньскому, но, с моей точки зрения, она очень похожа на одну из голов великого племени ольмеков, живших 3000 лет тому назад в пещерах Центральной Америки (см. Фото 23).
 Фото 22. Современная реконструкция Лузии, двадцатидвухлетней женщины, череп которой был найден в Минас Жерайс, Бразилия. Его возраст — свыше 11 500 лет.
Фото 22. Современная реконструкция Лузии, двадцатидвухлетней женщины, череп которой был найден в Минас Жерайс, Бразилия. Его возраст — свыше 11 500 лет.
 Фото 23. Резная каменная голова ольмеков из Мексики. Быть может, ее заметное сходство с Лузией — намек на давние генетические связи?
Фото 23. Резная каменная голова ольмеков из Мексики. Быть может, ее заметное сходство с Лузией — намек на давние генетические связи?
Кем является для нас Люзия: драгоценным звеном, соединяющим нас с неизвестным прошлым, или она лишь одна из множества? Люди часто повторяют, что первая ласточка весны не делает, так и в данном случае единственная находка еще не свидетельствует о массовой миграции. Нам необходимо большее количество находок. Исследования множества древних черепов из Серра да Капивара на севере Бразилии позволили Невесу сделать вывод о том, что примерно 9 тысяч лет тому назад форма черепов изменилась: толстокостные черепа уступили место монголоидным современного типа. Перед ученым предстали свидетельства о ранней миграции людей с толстокостными черепами немонголоидного типа, к которому относились собиратели на прибрежной полосе, чьи маршруты я описывал в главах 5 и 6. Во внешности собирателей на прибрежной полосе отразились все последние изменения, которые произошли в первичных типах людей уже за пределами Африки. Все современные австралийцы, меланезийцы, айну и полинезийцы (и даже европейцы) по типу скорее ближе к собирателям на прибрежной полосе, нежели к монголоидам Восточной Азии и Америки (см. главы 4 и 5). Поэтому, чтобы убедиться, что миграционный поток в Америку мог увлечь за собой часть людей немонголоидного типа, нам предстоит провести соответствующий сравнительный анализ. Однако среди обнаруженных в Азии останков собирателей на прибрежной полосе, живших более 12 тысяч лет тому назад, лучшей сохранностью отличаются толстокостные черепа восточноазиатского типа, которые более всего сходны с найденными в Китае. При этом они не имеют ничего общего с современными реликтовыми группами смешанного типа. Невес доказал связь между черепами палеоиндейцев, жителей Северного Китая, и черепами островитян южной части Тихого океана[438]. В отличие от сотрудников Би-би-си, создавших в 1999 году документальный фильм «Пути миграции первых жителей Америки», Невес, по-видимому, не был согласен с теорией, согласно которой люди, обладавшие такой же, как и Люзия, внешностью, переправились через Тихий океан в южной его части и приплыли в Южную Америку из Австралии. С его точки зрения, народ, для которого были типичны толстокостные черепа, переправился через континент Берингия или проплыл вдоль его берегов, то есть проделал тот же самый путь, что и все прочие народы. Интуиция подсказывала ученому, что этот тип людей впоследствии мог быть вытеснен или изгнан с обжитых земель под натиском мигрантов монголоидного типа, принесенных в Северную и Южную Америку новой, отдельной волной миграции. А может быть, останки собирателей на прибрежной полосе принадлежали американским аборигенам современного типа? Причем они могли принадлежать людям, жившим здесь еще в самом недавнем прошлом. Первопроходцы из Европы, высадившиеся на богатый остров на юге Америки, обнаружили здесь множество огней, разведенных на земле. С тех пор этот архипелаг на окраине континента называется Tierra del Fuego, или Огненная Земля. Люди, разводившие костры в надежде защититься от пронизывающих порывов холодного ветра, и были теми охотниками-собирателями, которые разительно отличались от своих соседей и всех известных этнических групп Америки по своему биологическому и культурному типу. На юго-западе, в лабиринте водных путей меж островами вокруг мыса Горн, жили так называемые «индейцы на каноэ». Немного севернее расположились поселения так называемых «пеших индейцев». К ним относилось племя теуэльче из Патагонии и селькнам, жившие на северо-востоке Огненной Земли. Проплывая по водам пролива, который ныне называется Магеллановым, сам Фернандо Магеллан и все члены его корабельной команды видели крупные фигуры охотников-собирателей с толстокостными черепами и группу костров, разведенных этими людьми. Черепа многих из них хранятся теперь в музеях, потому что англичане, захватившие впоследствии эти территории и обзаведшиеся здесь собственными ранчо, решили устроить свое благополучие на головах теуэльче и селькнам и получили от правительства щедрое денежное вознаграждение за каждую голову, ибо эти туземные народы стали нападать на овец, пасшихся на отобранных под овцеводческие хозяйства землях, которые служили некогда «раздольем для гуанако». Возможно, среди массы живущих здесь сегодня народностей осталось лишь две, которые могут по праву считаться потомками тех племен (см. Фото 24).
 Фото 24. Сегодня почти вымершая народность техуэльче в Терра дель Фуэго (фото датируется 1905 г.) — единственная современная этническая группа с чертами лица, близкими к австралоидам и горцам Новой Гвинеи.
Фото 24. Сегодня почти вымершая народность техуэльче в Терра дель Фуэго (фото датируется 1905 г.) — единственная современная этническая группа с чертами лица, близкими к австралоидам и горцам Новой Гвинеи.
Работающий в Кембриджском университете бразильский биолог и антрополог Марта Лар провела классическое исследование различий в строении черепа человека, в котором интересующий нас исторический период представлен группой черепов (двадцатью девятью, включая селькнам и теуэльче)[439]. Оказалось, что у этой группы черепов есть много общего со строением черепа австралийцев, в частности — толщина кости. Однако, если оставить в стороне толщину кости и связанные с ней особенности и параметры, то окажется, что других общих черт в строении и морфологии черепа (например, внеметрических параметров) у австралийцев и людей племени селькнам и теуэльче попросту не существует. В частности, у американских аборигенов головы были значительно крупней, в их облике сохранились черты, характерные для людей эпохи плейстоцена, а может быть, даже и для представителей еще более древних эпох. Лар придерживалась мнения, что эти охотники и собиратели не походили на современных аборигенов Америки, в их чертах было больше сходства с жителями Юго-Восточной Азии и островитянами южной части Тихого океана. Она пришла к выводу, что эти различия между американцами указывают на независимые волны миграции, которые увлекли за собой в Америку разные этнические группы. Недавно два самых известных в своей области ученых — Вальтер Невес, представляющий университет в Сан-Пауло, и Джозеф Пауэлл из университета в Нью-Мексико — объединили свои усилия в крупномасштабном сравнительно-аналитическом исследовании. Они стремились выяснить, не свидетельствуют ли накопленные факты о том, что прародиной палеоиндейцев служили сразу несколько регионов, что отразилось на большем этническом разнообразии древних жителей Америки, в коем они значительно превосходят американских аборигенов современности[440]. Сделанные исследователями открытия подтвердили, что в прошлом Америка пережила множество волн миграции, которые увлекли за собой многочисленные народы, ставшие родоначальниками местных аборигенов (альтернативная теория утверждает, что этническая дифференциация народов, по-видимому, произошла уже в самой Америке). Палеоиндейцы обладали значительным сходством с недифференцированным типом собирателей на прибрежной полосе Индийского и Тихого океанов. Современные же американские аборигены похожи скорее на жителей Северо-Восточной Азии; есть у них и черты, общие с европейцами. Далее ученые доказали, что палеоиндейцы Северной и Южной Америки сильно отличались друг от друга. Первые обладали поразительным сходством с полинезийцами, последние были более похожи на австралийцев. Причем существующие свидетельства о миграции из Австралии эти ученые не восприняли как прямое доказательство того, что где-то в южных широтах древние народы пересекли весь Тихий океан. Вместо этого они, соглашаясь с Лар, установили, что и у австралийцев, и палеоиндейцев юга была общая прародина — материковая часть Азии. Перед нами предстает картина множественных и независимых друг от друга взаимовлияний одного этноса на другой. Этот процесс, сказавшийся на особенностях строения черепа и скелета, шел между различными американскими группами (древними и появившимися совсем недавно), а также разнообразными племенами, типичными для Азии и Океании. Подобный взгляд на прошлое хорошо согласуется с предположением о как минимум двух, а возможно, трех и более отдельных волнах миграции из различных регионов Азии. В конце концов, самым распространенным этносом оказался монголоидный тип, который и вытеснил народы палеоиндейских типов[441]. Мы можем окончательно убедиться в этом, если вспомним, что некогда все американские генетические кланы-родоначальники мирно уживались на континенте Берингия, то есть на прародине, сыгравшей роль прибежища, сохранившего генетический материал для внутренней миграции, в результате которой различные народы разошлись по разным частям Восточной Азии. Согласуются ли взгляды антропологов с представлениями специализирующихся в области историигенетиков относительно того, что первые американцы были выходцами из различных географических зон? Столь сложные задачи по силам решать лишь мтДНК и Y-хромосоме, так предоставим им возможность ответить на этот вопрос. Теперь мы можем взглянуть на все американские линии-родоначальницы с позиций теории, которой была посвящена глава 6, и определить для каждой из линий, в какой части Азии находится ее генетическая прародина.
Два или три региона Азии можно назвать прародиной?
В последней главе я привожу развернутую повесть о Великом Оледенении, охватившем восток, юго-восток и северо-восток Азии. Вне досягаемости ледников осталась лишь чрезвычайно неблагоприятная для обитания Мамонтовая степь, простершаяся по высокогорному плато Центральной Азии к северу от Гималаев. Я также стремился доказать, что в Восточной Азии остались обитаемыми как минимум три различных региона, отличавшиеся друг от друга генным набором и техникой изготовления орудий. Первый регион был заселен потомками древних собирателей на прибрежной полосе, которые обжили восточное побережье Азии и отчасти Японию. Второй регион, по всей вероятности, был заселен монголоидами. Южная ветвь монголоидов расположилась на Юго-Востоке Азии, северная ветвь занимала земли к северу от Янцзы и, весьма возможно, центральную часть Цзиньхай-Тибетского плато. И, наконец, третья группа, по всей вероятности, расселилась на более северных территориях Евразии, которые лежат за горной системой Алтай, входящей в состав современной России, и включала в себя те же генные типы, которые характерны для жителей Северной и Восточной Европы. Возможной прародиной мигрантов в Америку следует считать как минимум три региона. Два из них, по всей видимости, находились в Центральной Азии. В доледниковый период эти регионы повсеместно соприкасались друг с другом и пересекались. В их состав входили неравномерно заселенные южные степи Сибири, земли, простиравшиеся на юг вплоть до Цзиньхай-Тибетского плато и на север — до горной системы Алтай в современной России. Итак, две интересующие нас этнические группы различались генным набором, культурным типом и внешним обликом: во-первых, это были охотники на мамонтов Верхнего палеолита, жившие на западе и северо-западе Евразии, а во-вторых, обживавшие более южные земли изготовители микролитов, которые принадлежали к только что начавшей складываться монголоидной расе. Как я уже упоминал в главе 6, первая группа, по всей видимости, ушла на восток, спустившись вниз по течению реки Уда к Охотскому морю и далее — вниз по Амуру к Сахалину и северной части Японии. Вторая из упомянутых этнических групп, двигаясь вниз по течению двух великих рек Китая — Янцзы и Хуанхэ, называемой также Желтой рекой, устремлялась к морскому побережью. Евразию в древности заселяли три народа со своеобразным набором генов, неповторимым обликом и техникой изготовления каменных орудий. Перед Последним ледниковым максимумом эти народы то смешивались друг с другом, то расходились. Они жили на азиатском побережье, простершемся от Японии и Кореи на северо-востоке до Охотского моря, на землях с относительно умеренным климатом. Каждый из этих народов мог свободно перебраться на Берингию, а затем в Америку. И возможность эта сохранялась вплоть до того времени, когда ледниковые шапки не преградили вход. Хотелось бы только знать, подтверждают ли результаты генетических экспериментов то, что народы переходили с континента на континент именно в этой последовательности. Первое интервью мы могли бы взять у мужской линии-родоначальницы, ибо ее оборванный рассказ донес до нас относительно ясный фрагмент истории.
Не была ли мужская линия-родоначальница Америки выходцем с севера Евразии?
Подтверждает ли историческая память Y-хромосомы то, что генный материал обязан своим происхождением сразу нескольким различным регионам? Полученный ответ звучит так: и нет, и да. Ответ «нет» связан с тем, что Y-xpoмосома является единственной доминирующей линией на территории всей Америки. А ответ «да» объясняется тем, что, если мы присмотримся повнимательнее к той картине, которую представляют параллельные мтДНК, мы обнаружим, что в них проникли элементы, разительно отличающиеся друг от друга и обязанные своим происхождением разным регионам Азии (см. рис. 7.6). Более того, мы столкнемся с забавными совмещениями линий. Наиболее поразительные факты связаны с мужской линией-родоначальницей. Дело в том, что более чем у 90% современных аборигенов Америки Y-хромосомы происходят от линии, распространенной в Северной Евразии. Поло (см. рис. 7.7), или так называемая ветвь Поло, одновременно встречается и у европейцев, среди которых в целом насчитывается 50% ее носителей, и у жителей Сибири, где она также достаточно представлена. Но ее не встретишь ни в Восточной Азии, ни в каком-либо ином регионе[442].
 Рис. 7.6
Рис. 7.6
 Рис. 7.7
Если учесть, сколько времени утекло, у нас не вызовет удивления тот факт, что в большинстве своем представленные в Европе подтипы линии Поло — это совсем не те подтипы, которые встречаются в Америке. Характерные для этих подтипов различия аналогичны тем, что обнаружены между вариантами митохондриевой Х-линии. И объясняются они тем, что в глубокой древности, возможно 25 тысяч лет тому назад, народ, который был когда-то единым, разделился. Став независимыми, две линии-родоначальницы начали развиваться каждая в своем направлении. И чем больше времени проходило со времени распада единого народа, тем больше накапливалось отличий. Эта история далеко не полностью описывает положение, в котором оказалась линия X. Что же касается линии Поло, то ее типы, встречающиеся в Северной Азии, характерные для основной линии-родоначальницы Америки и обнаруженные, хотя и в малом количестве, в Европе, — все эти типы восходят к единому корню. Данные научные изыскания подтверждают гипотезу о том, что у мужской Y-линии европейцев и американцев была общая прародина — какой-нибудь из регионов Северной или Центральной Азии, а также гипотезу о вторжении народов, заселивших Америку, с севера.
Проанализировав расселение племен по Сибири, российский генетик Татьяна Карафет со своими коллегами пришла к выводу, что регион, прилегающий к озеру Байкал, является прародиной древних мигрантов в Америку, унаследовавших местный ген-маркер (хотя высокая степень распространения здесь гена-маркера сама по себе еще не указывает однозначно на родину мигрантов). Европейско-американские связи этим не ограничиваются. Доминирующая в Европе подгруппа линии Поло, линия Руслан, встречается у 30 % европейцев; она обнаружена также у 12% аборигенов Америки. Столь высокий процент нельзя объяснить недавней примесью генного материала европейцев. Родоначальный тип Руслан встречается также в Северной Азии, где могла располагаться прародина американской линии-родоначальницы Руслан[443].
Значительно чаще, чем эти две линии (Поло и Руслан), в Америке встречается дериват линии Поло — Q, который я буду называть Quetzalcoatl (Кецалькоатль)[444], по имени мифического пернатого змея. На Кецалькоатль и его сыновей приходится 64% Y-хромосом у американцев, а среди американских аборигенов он встречается лишь в исключительных случаях. Исключений из этого правила совсем немного: это три эскимоса в далекой Сибири, по одному уникальному человеку найдено в соседней этнической группе Сибири — среди чукчей; среди эвенков, живущих изолированной группой в Сибири на дальнем южном побережье Тихого океана; в народности маньчжуров (в Маньчжурии) и среди узбеков в Центральной Азии. Для всех этих исключений характерен специфический американский тип Y-хромосомы — линия Кецалькоатль. С тех пор как линия Кецалькоатль была обнаружена у представителей группы на-дене и эскимосско(инуитско)-алеаутской группы, стало очевидным, что американский генмаркер Y появился здесь в глубокой древности, возможно, еще до Ледникового периода, во времена, когда кланы-родоначальники еще не вторглись в Северную Америку. Иначе говоря, Кецалькоатль — исконная линия-родоначальница, а не производный тип-дериват первого поколения. Такая точка зрения подтверждается данными генетики. Представители этой науки оценивают возраст американского гена-маркера Кецалькоатль в 22 тысячи лет[445].
Есть и еще один тип Y-линии, который распространен в Северной Евразии, но крайне редко встречается среди аборигенов Америки. Это — тип ТАТ. Не исключено, что он появился в результате послеколумбовского влияния генного материала европейцев, но, с точки зрения статистики, предположение, что этот тип-маркер распространился из Северной Азии, является более обоснованным[446].
Доминирование в Америке характерной для Северной Евразии линии Y кажется удивительным фактом, который, по всей вероятности, указывает на Южную Сибирь как на единственно возможный регион, который мог послужить исходным пунктом для мигрантов в Америку. Концепцию о многочисленных волнах миграции можно забыть. Однако следует учитывать, что по сравнению с мтДНК Y-хромосомы значительно чаще подвержены дрейфу. Столь поразительное доминирование одной Y-линии можно объяснить еще и следующим образом: в традиционных обществах некоторая группа мужчин стремится обзавестись большим количеством детей, в то время как у женщин детей бывает приблизительно поровну. В результате эффект дрейфа увеличивается, и становится ясно, почему американский тип Y-линии столь явно преобладает.
Однако остатки американской линии-родоначальницы Y скорее всего связаны с Дальним Востоком, а не с Южной Сибирью, и унаследованы они, вероятно, от собирателей на прибрежной полосе, а не от охотников Центральной Азии. Линия-родоначальница собирателей на прибрежной полосе получила название Каин. Покинув Африку, это племя поселилось вдоль побережья Тихого и Индийского океанов. Генетическую преемственность этих племен можно установить благодаря особой мутации М217. Хотя линия Каин широко распространена в Северной и Восточной Азии, но с максимальной плотностью она представлена на побережье Тихого океана, в Маньчжурии и у Охотского моря, а также вдоль нижнего течения реки Амур. Если спуститься вниз, на юго-восток Азии, то окажется, что линия Каин встречается как минимум у 10% населения (см. таблицу). Учитывая прошлое линии Каин, которая бытовала в среде собирателей на прибрежной полосе, можно предположить, что она была тем типом Y, который на своей прародине, то есть в более южных областях Азии, сочетался с маркером мтДНК — В (см. ниже)[447].
Рис. 7.7
Если учесть, сколько времени утекло, у нас не вызовет удивления тот факт, что в большинстве своем представленные в Европе подтипы линии Поло — это совсем не те подтипы, которые встречаются в Америке. Характерные для этих подтипов различия аналогичны тем, что обнаружены между вариантами митохондриевой Х-линии. И объясняются они тем, что в глубокой древности, возможно 25 тысяч лет тому назад, народ, который был когда-то единым, разделился. Став независимыми, две линии-родоначальницы начали развиваться каждая в своем направлении. И чем больше времени проходило со времени распада единого народа, тем больше накапливалось отличий. Эта история далеко не полностью описывает положение, в котором оказалась линия X. Что же касается линии Поло, то ее типы, встречающиеся в Северной Азии, характерные для основной линии-родоначальницы Америки и обнаруженные, хотя и в малом количестве, в Европе, — все эти типы восходят к единому корню. Данные научные изыскания подтверждают гипотезу о том, что у мужской Y-линии европейцев и американцев была общая прародина — какой-нибудь из регионов Северной или Центральной Азии, а также гипотезу о вторжении народов, заселивших Америку, с севера.
Проанализировав расселение племен по Сибири, российский генетик Татьяна Карафет со своими коллегами пришла к выводу, что регион, прилегающий к озеру Байкал, является прародиной древних мигрантов в Америку, унаследовавших местный ген-маркер (хотя высокая степень распространения здесь гена-маркера сама по себе еще не указывает однозначно на родину мигрантов). Европейско-американские связи этим не ограничиваются. Доминирующая в Европе подгруппа линии Поло, линия Руслан, встречается у 30 % европейцев; она обнаружена также у 12% аборигенов Америки. Столь высокий процент нельзя объяснить недавней примесью генного материала европейцев. Родоначальный тип Руслан встречается также в Северной Азии, где могла располагаться прародина американской линии-родоначальницы Руслан[443].
Значительно чаще, чем эти две линии (Поло и Руслан), в Америке встречается дериват линии Поло — Q, который я буду называть Quetzalcoatl (Кецалькоатль)[444], по имени мифического пернатого змея. На Кецалькоатль и его сыновей приходится 64% Y-хромосом у американцев, а среди американских аборигенов он встречается лишь в исключительных случаях. Исключений из этого правила совсем немного: это три эскимоса в далекой Сибири, по одному уникальному человеку найдено в соседней этнической группе Сибири — среди чукчей; среди эвенков, живущих изолированной группой в Сибири на дальнем южном побережье Тихого океана; в народности маньчжуров (в Маньчжурии) и среди узбеков в Центральной Азии. Для всех этих исключений характерен специфический американский тип Y-хромосомы — линия Кецалькоатль. С тех пор как линия Кецалькоатль была обнаружена у представителей группы на-дене и эскимосско(инуитско)-алеаутской группы, стало очевидным, что американский генмаркер Y появился здесь в глубокой древности, возможно, еще до Ледникового периода, во времена, когда кланы-родоначальники еще не вторглись в Северную Америку. Иначе говоря, Кецалькоатль — исконная линия-родоначальница, а не производный тип-дериват первого поколения. Такая точка зрения подтверждается данными генетики. Представители этой науки оценивают возраст американского гена-маркера Кецалькоатль в 22 тысячи лет[445].
Есть и еще один тип Y-линии, который распространен в Северной Евразии, но крайне редко встречается среди аборигенов Америки. Это — тип ТАТ. Не исключено, что он появился в результате послеколумбовского влияния генного материала европейцев, но, с точки зрения статистики, предположение, что этот тип-маркер распространился из Северной Азии, является более обоснованным[446].
Доминирование в Америке характерной для Северной Евразии линии Y кажется удивительным фактом, который, по всей вероятности, указывает на Южную Сибирь как на единственно возможный регион, который мог послужить исходным пунктом для мигрантов в Америку. Концепцию о многочисленных волнах миграции можно забыть. Однако следует учитывать, что по сравнению с мтДНК Y-хромосомы значительно чаще подвержены дрейфу. Столь поразительное доминирование одной Y-линии можно объяснить еще и следующим образом: в традиционных обществах некоторая группа мужчин стремится обзавестись большим количеством детей, в то время как у женщин детей бывает приблизительно поровну. В результате эффект дрейфа увеличивается, и становится ясно, почему американский тип Y-линии столь явно преобладает.
Однако остатки американской линии-родоначальницы Y скорее всего связаны с Дальним Востоком, а не с Южной Сибирью, и унаследованы они, вероятно, от собирателей на прибрежной полосе, а не от охотников Центральной Азии. Линия-родоначальница собирателей на прибрежной полосе получила название Каин. Покинув Африку, это племя поселилось вдоль побережья Тихого и Индийского океанов. Генетическую преемственность этих племен можно установить благодаря особой мутации М217. Хотя линия Каин широко распространена в Северной и Восточной Азии, но с максимальной плотностью она представлена на побережье Тихого океана, в Маньчжурии и у Охотского моря, а также вдоль нижнего течения реки Амур. Если спуститься вниз, на юго-восток Азии, то окажется, что линия Каин встречается как минимум у 10% населения (см. таблицу). Учитывая прошлое линии Каин, которая бытовала в среде собирателей на прибрежной полосе, можно предположить, что она была тем типом Y, который на своей прародине, то есть в более южных областях Азии, сочетался с маркером мтДНК — В (см. ниже)[447].
Материнские линии из других регионов Азии
В отличие от мужской линии, у материнских линий-родоначальниц Америки было значительно большее число исходных зон распространения, и это были иные географические районы, нежели те, что признаны прародиной мужской линии. Обратимся к примеру группы мтДНК — В, которая произошла из Юго-Восточной Азии и отсутствует в северных регионах этого континента. Но именно из Северной Азии, по всей видимости, произошла американская линия Y. В соответствии с гипотезой о множественности первоисточников линий-родоначальниц Нового Света мы должны расширить географию поисков прародины мигрантов. В последней главе я обозначил на карте те районы Азии, которые соответствуют пяти генетическим материнским линиям-родоначальницам Америки. Группа С обнаружена в Азии, на севере Гималаев. Линия А значительно чаще, чем в более южных районах, встречается на северовостоке Азии. Итак, нам следует искать прародину этих двух линий-родоначальниц, принявших участие в заселении Америки, на севере Гималаев. Группа С широко распространена на обширной территории от Тибета и почти до Урала. Максимальной плотности она достигает среди селькупов. Так называется племя охотников и рыболовов, живущее за Северным Полярным кругом, в нижнем течении Енисея. На азиатском побережье Тихого океана и далее на юг линия С встречается значительно реже. Как уже говорилось в главе 6, во времена последнего Ледникового периода свободным ото льдов оставался лишь узкий перешеек, в результате возникла необходимость вторичного заселения территорий. Поэтому частотность, с которой встречается тот или иной ген, сама по себе не всегда указывает на какой-либо регион как на прародину этого гена. Приняв к сведению эту информацию, мы вынуждены признать, что поскольку единственным местом, где во время последнего Ледникового периода продолжали жить люди, оставался северный регион Центральной Азии, включающий в себя озеро Байкал, верховья Енисея и земли на юге Сибири, то оснований называться прародиной американской группы С у этого региона не больше, чем у любого другого[448]. Итак, группа А пришла в Америку с юга Китая, скорее всего, из области, простершейся от верховьев Янцзы до плато Цзиньхай. За исключением крайней северовосточной оконечности Сибири, где эта группа явно доминирует, она остается нетипичной для Центральной и Северной Азии. Учитывая то, что северо-восточная часть Сибири входила в состав Берингии и, следовательно, оказалась в непосредственной близости от Северной Америки, а также то, что после Последнего ледникового максимума линия А была вовлечена в процесс вторичного заселения Берингии, можно утверждать, что северо-восточная область Сибири более других регионов Азии соответствует нашим представлениям о прародине американской линии-родоначальницы А. В главе 5 уже было высказано предположение о том, что линия С в конечном счете произошла из Индии и Пакистана, ее путь лежал через окраинные области Гималаев на западе. Для людей Верхнего палеолита, заселявших перед последним Ледниковым периодом территории от Франции до Маньчжурии, ключевым геном-маркером была родоначальница Запада — материнская линия С. Весьма возможно, что охотники той поры передали свои навыки жившим в доледниковый период предкам охотников за крупным зверем, которые впоследствии создали в Северной Америке культуру Кловис, известную своими наконечниками и получившую широкое распространение уже в послеледниковый период. Подобно линии С, американская линия X тоже, по всей вероятности, происходит из расположенных ближе к северу регионов Евразии (глава 5), но поскольку между Америкой и Азией есть обширная зона, в которой эта линия отсутствует, остается загадкой, какой именно регион послужил исходным пунктом распространения этой линии по Америке. Недавно были получены неопровержимые свидетельства распространения линии X на севере Азии, среди алтайцев юга Сибири. Может быть, этот регион и является прародиной линии X? Существовавшая 30 тысяч лет тому назад сухопутная связь с Европой не позволяет сомневаться в том, что линия X пересекла азиатские степи: она двигалась вместе с охотниками Верхнего палеолита. Редко встречавшаяся на Кавказе, линия X участвовала в заселении Америки, а затем в Азии она почти полностью вымерла. К пяти американским линиям-родоначальницам относится также группа D, которая и в наши дни часто встречается на северо-востоке Азии. Перебравшись через Южные отроги Гималаев, прапредки группы D могли проникнуть на север Азии с юго-востока этого континента. В наши дни эта группа распространена в Азии вдоль всего восточного побережья Тихого океана. Тот факт, что ареал, в котором представлена эта группа, простирается далеко на юг, захватывая даже Южный Китай, наводит на мысль о магистральных прибрежных маршрутах передвижения с севера[449]. Прародиной материнской линии-родоначальницы В, очевидно, являются южные регионы, в противоположность более северным территориям, где в наши дни распространены четыре уже названные материнские линииродоначальницы. Группа В доминирует в Индокитае, на Юго-Востоке Азии, в Тихом океане. Ареал распространения этой группы значительно отличается от областей, где представлены северные линии. Линия В найдена в Японии, Китае и Монголии, но она отсутствует в более северных регионах, начиная с субарктических поясов Евразии и Северной Америки. Этот факт позволяет предположить, что источником распространения американской линии-родоначальницы В являются иные регионы, чем те, что были прародиной для групп С и X. В результате мы можем сузить область поиска прародины группы В. В Азии группа В представлена двумя основными ветвями — В4 и В5, которые уживаются бок о бок друг с другом в большинстве регионов за исключением Америки и южной части Тихого океана, где представлена только линия В4. Это единственная ветвь, которая достигла Америки. Кроме того, она доминирует в южной части Тихого океана. На востоке Азии ветвь В4 настолько окрепла и разрослась, получила такое широкое распространение и разделилась на столь обильное многообразие вариантов, что обследовать Азию в надежде найти точный аналог родоначального типа Америки — это все равно что искать иголку в стоге сена. Однако в результате случайного стечения обстоятельств двум исследовательским группам, специализирующимся в области изучения цепочек мтДНК, удалось независимо друг от друга обнаружить приблизительные аналоги американских вариантов. Одну из этих групп возглавляли японские генетики, а другую — генетики из Швеции и Германии. Этот трудоемкий метод, напоминающий скорее миниатюрный вариант проекта «Геном человека», примененный лишь к одному отрезку ДНК, позволяет понять последовательность всей спирали молекулы мтДНК. Полученный в результате исследований своего рода дактилоскопический отпечаток пальца, точнее, отрезка материнского генетического кода оказался четким и передающим всю его специфику. Когда я сравнивал данные, полученные этими двумя исследовательскими группами, оказалось, что искомый аналог американской группы-родоначальницы представляет собой нечто среднее между пимской индийской группой В4 и японской группой В4, что указывает на узкую полосу азиатского побережья Тихого океана как на прародину как минимум одного американского генетического типа, а именно В. Найденные аналоги обладают поразительным сходством, хотя, как того и следовало ожидать, они не похожи друг на друга как две капли воды. Однако они настойчиво свидетельствуют о связи между Японией и Америкой, исключая из области наших поисков все остальные древние ответвления азиатской линии В4[450].
Коалиция мигрантов: пестрая смесь
Оказалось, что факты, добытые в ходе научной работы по выявлению приблизительных соответствий между конкретными материнскими типами Азии и Америки, не позволяют точно установить, какие регионы Азии могли послужить прародиной для каждой из пяти американских линий-родоначальниц. Эта задача кажется еще более трудной, нежели поиск региона типа Монголии, под небом которого можно встретить все типы: А, В, С и D, — что, возможно, позволит установить единственную область Азии, которая явилась прародиной американских линий-родоначальниц. Накопленные данные, однако, учат нас более непредвзято оценивать череду событий далекого прошлого, приведших к заселению Америки, и учитывать возможность того, что мигранты, колонизировавшие Америку, вышли из различных регионов Азии. В главе 6 мы уже обращались к данным археологии, которая свидетельствует, что на азиатском побережье Тихого океана сохранились как относящиеся к Верхнему палеолиту следы техники обработки каменных орудий, принесенной сюда из степей Западной Евразии, так и следы, по-видимому, культуры микролезвий, зародившейся во время похолодания, предшествовавшего последнему Ледниковому периоду, на Тибете и Цзиньхайском плато. Возможно, эти артефакты появились в этих местах в результате миграции населения с Кавказа — на запад, а из Монголии — дальше на юг. Достигнув северо-востока Азии, Кореи и Японии, племена мигрантов стали жить бок о бок с потомками собирателей на прибрежной полосе (их кровь течет ныне в жилах людей народности айну (айнов), раньше других народов облюбовавших восточное и юго-восточное побережье Азии). Итак, возможно, как минимум три различные этнические группы с ярко выраженными отличительными признаками во внешнем облике двинулись вдоль побережья на северо-восток. Жившие за счет моря, изобиловавшего различными формами жизни, и охотившиеся на дичь, эти народы в конечном счете достигли Берингии и принесли на этот континент различные виды культуры, генное разнообразие и неповторимые типы внешности.
Путешествуя в одиночку
Как могли выглядеть народы Берингии 22—25 тысяч лет тому назад накануне вторжения на Аляску? Может быть, они были похожи на обитателей звездной базы № 9 из фильма «Звездный путь», населенной полиглотами, наделенными способностью к полиморфизму (многообразию облика) и полихромии (многообразию цвета кожи, волос и т.д.)? Другими словами, представляли ли они собой общество, в котором завершился процесс смешения народов в единую однородную массу? Я так не думаю. Берингия была огромным, удобным для жизни континентом, богатым разнообразными ресурсами. Она могла предоставить народам различных культурных типов территории с подходящими для них условиями жизни. Несмотря на то что процессы ассимиляции были неизбежны, есть все основания полагать, что этнические группы существовали независимо друг от друга, что позволило сохранить культурное и генетическое многообразие. Во времена, когда этнические группы мигрантов только что вступили на земли Северной и Южной Америки и, разойдясь веерообразно, стали плодиться и размножаться, они вовсе не были похожи на многонациональное объединение или своеобразную Лигу наций. Народы, только что пришедшие из разных регионов Азии, пустили корни в Новом Свете. Условия жизни не только способствовали сохранению каждым народом своего физического, генетического и культурного своеобразия, но они благоприятствовали расцвету культур. Однако подтверждают ли подобную картину данные археологии? Думаю, что несомненно подтверждают. В результате археологических работ по изысканию артефактов, оставшихся от времен, предшествующих эпохе Кловис, ученые обнаружили под культурным слоем Кловис и в самых ранних культурных слоях не относящихся к типу Кловис стоянок в Северной и Южной Америке орудия труда, которые были сделаны представителями совершенно различных культурных традиций, причем в Азии найдены аналоги для каждого из типов орудий. Приняв гипотезу о параллельной колонизации Америки народами, принадлежащими к разным культурным типам, мы вправе были бы ожидать именно таких археологических находок. Мы хотели бы надеяться, что нам удастся выявить четкие генетические отличия между этими народами, поскольку внешние особенности, отличавшие одну первобытную общину Америки от другой, очевидно, были весьма многообразны. Внешние отличия народов могут быть выявлены на основании выводов, сделанных Торрони и Уоллэсом (см. выше), — эти ученые обнаружили, что для различных племен характерны различные подтипы мтДНК. Итак, Америка была заселена в результате единой и единственной волны миграции, об этом свидетельствует также тот факт, что по всей Америке — в Северной, Центральной и Южной ее части — обнаружены крупные этнические группы, каждая из которых относится к определенному генному типу: А, В, С или D. Просторы Южной и Северной Америки поистине огромны, однако среди множества населяющих их народов пять американских родоначальных типов мтДНК встречаются с различной частотой. В наши дни принадлежность к той или иной этнической группе определяется на основании специфических особенностей типов мтДНК, характерных для конкретного клана. За редким исключением почти не существует таких типов мтДНК, которые являлись бы отличительной чертой определенной этнической группы, а вместе с тем встречались бы и среди других групп. Кроме того, оказалось, что области преимущественного распространения основных родоначальных генных кластеров распределены на американском континенте весьма неравномерно и встречаются с различной частотой. По всей вероятности, из этого можно сделать вывод, что единый родоначальный клан мигрантов очень быстро разделился на группы, которые стали жить независимо друг от друга. Причем внутри каждой из групп начались процессы генного дрейфа, которые и привели к тому, что родоначальные генные кластеры стали встречаться с различной частотой. Однако мы можем с равным успехом объяснить то, как сложилась подобная картина, на основании альтернативной гипотезы, в соответствии с которой первоначальная волна миграции содержала множество разрозненных генных примесей. Их путь хорошо прослеживается, и мы можем мысленно пройти по нему в обратном направлении: через Берингию — к различным регионам Азии, в которые уходят корнями эти генные типы. Мы уже видели, что в Сибири сложилась чрезвычайная ситуация: жители Аляски и северо-западного побережья, говорящие на языках группы на-дене, сохранили единственный американский материнский тип — группу А2. Мы уже объясняли также, почему среди носителей языков групп на-дене и эскимосско(инуитско)-алеутской оказалась распространена лишь одна американская линия-родоначальница — А2. Все это является результатом крайне трудных климатических условий, поставивших предков этих народов на грань выживания и вызвавших генный дрейф, вследствие которого количество линий-родоначальниц сократилось до одной[451]. Генный кластер А явно преобладает на территориях Северной Америки, простершихся к северу от 50-й параллели и включающих не только области распространения языковых групп на-дене и эскимосско(инуитско)-алеутской, но и америнд, и ареал северного племени оджибва, живущего в Канаде и в районе Великих озер, язык которого относится к алгонкинской[452] семье. Здесь группа А представлена тремя уникальными и нигде более не обнаруженными типами. Кроме того, среди представителей племени оджибва отмечен самый высокий процент носителей редкого типа X, он составляют 25%. Кроме того, на долю этого же племени выпала удивительная честь — в их генах представлен только доминирующий родоначальный тип Y, а не его дериваты, которые характерны для подавляющего большинства аборигенов Америки. Хотя язык оджибва и относится к макросемье америнд (американских индейцев), у представителей этого племени много общего с носителями языков группы на-дене: это и субарктический регион распространения и даже некоторые культурные черты, характерные для народов Севера. Генный набор оджибва уникален, а вместе с тем достаточно разнообразен. В его состав преимущественно входят исконные линии-родоначальницы. Это наводит на мысль, что в отличие от народов группы на-дене, которые во время Ледникового периода были поставлены на грань выживания, генетические коды оджибва и родственных ему этнических групп сохранились в неизменном виде, что делает их достойными особого внимания. Возможно, благодаря изолированной жизни в северных регионах эти народы удержали генные коды, являющиеся непосредственными дериватами того генного материала, источником которого была Берингия, а в конечном счете — Северо-Восточная Азия[453]. Во время работы над фильмом «Истинная Ева» была набрана команда добровольцев, согласных предоставить для генного анализа образцы своих тканей, в ее состав вошли два американских аборигена, родившихся в районе Великих озер. Благодаря удивительному совпадению оба они были представителями этих уникальных, редко встречающихся локальных типов. Участник эксперимента из племени оджибва оказался обладателем редкого типа А1, в то время как у члена племени кри (язык народа кри также относится к алгонкинской семье языков) был выявлен тип X, обнаруженный ранее лишь у северян из племени оджибва[454]. На карте распространения материнских линий-родоначальниц из группы А отражена отчетливая тенденция к снижению уровня частотности в направлении с севера, точнее — с субарктического пояса Северной Америки, где на долю этой группы приходится почти 100%, на юг, то есть в области Южной Америки, где, если говорить в общем, группа А встречается крайне редко. Однако при более внимательном рассмотрении карты окажется, что в других, несубарктических регионах Северной Америки уровень линии А претерпевает значительные колебания: он опускается до нулевых отметок и поднимается до очень высоких значений, соответствующих столь же высокому проценту встречаемости среди местного населения. В результате небольшого числа исследований ДНК древних жителей Северной и Южной Америки были обнаружены народы, у которых группа А полностью отсутствовала, из чего можно сделать вывод, что современная картина распространения группы А существенно отличается от той, что была в древности. В качестве примера народов, в генах которых совершенно отсутствует группа А, можно назвать северные фремонтские культуры, сложившиеся в районе озера Грейт-Солт-Лейк, а также исчезнувшие племена фуэга, жившие некогда на мысе Южной Америки (об этом см. выше), у которых были лишь линии С и D[455]. Выше уже было сказано, что если сравнить ареалы распространения среди американских индейцев групп А и X, то ареал группы А покажется чрезмерно большим, в то время как распространение линии X ограничено двумя этническими группами, живущими на Дальнем Севере Америки. С другой стороны, в Северной Америке группа D встречается крайне редко, зато она широко распространена в Южной Америке, особенно в экваториальном поясе. Короче говоря, то удивительное разнообразие, которым характеризуются закономерности распределения по Америке пяти линий-родоначальниц, свидетельствует, возможно, о независимых этнических примесях к первоначальной волне миграции (хотя возможны и другие объяснения).
Служило ли западное побережье коридором в Америку?
Напоследок я приберег самую интересную информацию — об особенностях распространения группы В. Как было сказано выше, группа В полностью отсутствует к северу от 55-й параллели, хотя и представлена в Северной, Центральной и Южной Америке. Часто на основании того факта, что на юге существуют разрозненные ареалы распространения линии В, высказываются предположения о том, что линия В появилась в Америке одновременно с другими линиями, то есть еще до Ледникового периода. Хотя я и согласен с этим утверждением, но вопрос о точном возрасте линии В остается открытым. Торрони полагает, что линия В значительно моложе А, С или D, что согласуется с данными повторного анализа Форстера, который подтвердил, что группа В, представленная в Центральной Америке, моложе других линий. В Южной Америке группа В оказалась достаточно старой, а диапазон, в котором варьируется уровень ее встречаемости, здесь очень широк: он составляет от 0 до 70%. Хотя указанные противоречия, связанные с оценкой возраста и ареала распространения линии В, могли сложиться вследствие межледникового перешейка и генного дрейфа или появиться в результате неадекватных выводов на основании некорректно проведенных анализов, но есть и другие, весьма интригующие объяснения названного эффекта. Начнем с того, что в Восточной Азии группа В4 широко распространена и представлена множеством вариантов. Так что вторжение типа В4, точнее, типов-близнецов линии В4 в Америку произошло как минимум дважды и в различное время, хотя мы пока не располагаем на этот счет неопровержимыми доказательствами, которые могут быть обнаружены при более детальном анализе последовательности оснований в ДНК жителей Америки[456]. С другой стороны, линия В4 могла проникнуть независимо от других линий и появиться в последний момент, уже по окончании эпохи заселения Америки линиями А, С, D и X. Именно к таким выводам пришла российский генетик Елена Стариовская. Как бы оставив на время в стороне факты, свидетельствующие о молодости типа В, она положила в основу своей гипотезы удивительные наблюдения: оказывается, группа В часто встречается в регионах Центральной Америки, где находят орудия эпохи Кловис, но при этом отсутствует на крайнем юге Америки. Линия В не обнаружена также на западном побережье Субарктики (см. выше) среди носителей языков группы на-дене, однако представлена в более южных регионах, там, где распространены языки группы америнд. Кроме того, Стариовская выдвигает гипотезу независимого вторжения линии В в Америку, носители которой, по ее мнению, могли проникнуть на американский континент, двигаясь вдоль западного побережья Северной Америки, — при помощи этой гипотезы исследовательница, видимо, стремится объяснить отсутствие этой линии в Субарктике, а также противоречивые следствия ледникового барьера. В согласии с этой гипотезой существовал как минимум один тип линии В4, который был близким аналогом японской разновидности индийского пимского типа В4. Чтобы выяснить, насколько справедливо данное предположение, необходимы дальнейшие исследования последовательности оснований ДНК людей, живущих по обеим сторонам Тихого океана[457]. Возможно, миграция происходила по ломаной траектории: сперва быстрое скачкообразное движение по береговой линии вперед — от Берингии, а затем резкий поворот вниз, к западному побережью Северной и Южной Америки. Эта идея созрела в головах ученых уже очень давно, давно отбродила и встала на полку рядом со старинными винами, но в наши дни она вновь вошла в моду. Впервые эту идею выдвинул Кнут Фладмарк из университета Симона Фразера в Британской Колумбии. С тех пор прошло уже тридцать лет. Эта идея до сих пор притягивает умы ученых, так как позволяет объяснить феномен крайне быстрого заселения Южной Америки. Однако из-за недостатка фактов и научных свидетельств доказать эту теорию оказывается сложно. Большая часть прибрежной полосы, по которой могли пройти гипотетические мигранты, в наши дни скрыта в глубине морской. Только крутые берега могли противостоять колебаниям уровня моря. Именно на таких берегах и расположены две стоянки древнего человека, найденные на южном побережье Перу: Квебрада Джаквай и Квебрада Такахуай. Здесь были обнаружены кости толстоногих птиц и фрагменты морских раковин (двустворчатых ракушек и мидий), ракообразных и анчоусов. Все эти находки рассказывают нам о жизни искусных рыболовов и собирателей на прибрежной полосе, осваивавших побережье Южной Америки 11 тысяч лет тому назад, то есть в то самое время, когда охотники эпохи Кловис предпринимали героические попытки истребить всех крупных животных севера. К этой же эпохе относятся найденные на западном побережье Северной Америки материальные свидетельства пребывания здесь людей, занимавшихся в те давние времена собирательством на прибрежной полосе[458]. Некоторые археологические находки подтверждают, что вскоре после того, как Последний ледниковый максимум остался позади, на землях, скрытых ныне под толщей воды, жизнь кипела — здесь пролегали древнейшие маршруты переселения людей, о чем свидетельствует совсем другое всеядное млекопитающее — медведь. Дженнифер Леонард, биолог из Калифорнийского университета в Лос-Анджелесе, и Алан Купер, работающий в Оксфордском университете, новозеландский специалист по древним ДНК, опираясь на данные, записанные в древних и современных мтДНК североамериканского бурого медведя, попытались реконструировать пути расселения этого животного. Ученых интересовало, какая судьба постигла медведей, живших на Аляске и северо-западном побережье Америки, во время Великого Оледенения[459]. Оказалось, что мтДНК бурых медведей, распространенных ныне в некоторых регионах Восточной Евразии и в США, очень похожи друг на друга, что наводит на мысль о том, что мтДНК бурых медведей Евразии восходит к линиям, которые были распространены на Аляске, а затем проникли в Америку по существовавшему перед Ледниковым периодом в Берингии коридору. Хотя, как и в случае с людьми, во время ПЛМ разнообразие типов медведей значительно сократилось, они, как известно, выжили и не только в Берингии, но и на острове Принца Уэльского, расположенного за пределами юго-восточного региона Аляски. В Южной Канаде перед Последним ледниковым максимумом бурые медведи не водились, но они обитали здесь 13 тысяч лет тому назад. Группы митохондриевых ДНК медведей свидетельствуют о том, что животные не были выходцами с Аляски, покинувшими ее в эпоху ледникового коридора или уже после таяния льдов в процессе вторичного заселения Америки. Но медведи были выходцами с островов, расположенных у западного побережья и послуживших животным укрытием во время ПЛМ. Когда борьба за жизнь стала невыносимой, некоторые кланы медведей, должно быть, ушли с Аляски — они двинулись вниз, в сторону западного побережья, где им удалось выжить и сохраниться до наших дней. К каким же выводам пришли ученые относительно бурых медведей, живущих ныне в Канаде и в горах Роки-Маунт в США? Оказалось, что для этих животных характерны типы мтДНК, которые восходят к линиям-родоначальницам, получившим распространение на Аляске 35—45 тысяч лет тому назад, но к настоящему моменту давно на ней вымершим. Другими словами, медведи Америки произошли от древних обитателей Берингии. Вполне возможно, что история распространения медведей укажет нам на те маршруты, которыми могли воспользоваться люди, жившие 12—15 тысяч лет тому назад, тем более что и медведи, и жившие на прибрежной полосе люди были всеядны, и рационы их питания в значительной степени совпадали. Возраст артефактов и человеческих останков, найденных на острове Принца Уэльского, составляет 9300 лет, причем эти археологические находки, возможно, и еще старше. Если же мы перенесемся на 3200 км к югу от этого острова, то окажемся в пещере Дейзи Кейв (или в пещере Маргариток), расположенной на острове Сан-Мигель в проливе Санта-Барбара на юге Калифорнии — именно здесь Джон Эрландсон извлек из земли археологические свидетельства пребывания в этих местах собирателей на прибрежной полосе, живших приблизительно 11 600 лет тому назад. Совершенно очевидно, что преодолеть 40 км, отделяющие материк от острова, не обладая специальными умениями и навыками, было бы невозможно. Радиоуглеродный анализ фрагментов костей женщины, найденных поблизости от острова Санта-Роза, позволил определить возраст ее останков, который составляет 13 тысяч лет[460]. Насколько давно прибрежные земли освободились от ледяных оков и появилась возможность проложить маршруты для свободного передвижения вдоль береговой линии? Кажется, это произошло 14 тысяч лет тому назад. Дарил Феджи из Управления национальных парков Канады, расположенном в провинции Британская Колумбия, и Хейнер Джозенханс из Геологической службы Канады при помощи гидролокатора, обладающего высокой разрешающей способностью, составили подробную карту океанического дна у северо-западного побережья Канады[461]. Ученые выбрали для исследований область вокруг островов Королевы Шарлотты, лежащих к югу от Аляски. Карта открывает нам новый мир, в котором текли когда-то извилистые реки, спускаясь к затопленным ныне равнинам и древним озерам. Период, во время которого эти равнины лежали выше уровня моря и были свободными ото льда, начался 14 тысяч лет тому назад и продолжался несколько тысячелетий, до тех пор, пока неуклонный процесс повышения уровня моря не завершился тем, что прибрежные земли вновь оказались скрытыми под толщей воды. Вооружившись новой картой затопленной части береговой линии, исследователи предприняли попытку составить коллекцию древесных пород затопленных лесов. Они подняли со дна океана сосновый пень и частицы деревьев других пород. Установленный на основе радиоуглеродного метода возраст обнаруженных фрагментов составляет 12 200 лет. К этому же периоду относятся найденные учеными остатки съедобных моллюсков и ракообразных. Береговой шельф, расположенный на глубине 60 метров (200 футов) ниже уровня моря, оказался несколько моложе — ему 10 тысяч лет, но именно на нем исследователи обнаружили самое древнее каменное орудие из когда-либо найденных на северо-западном побережье Америки. Обобщив все эти факты и свидетельства, можно с достаточной степенью уверенности утверждать, что период, когда люди получили возможность покинуть Берингию и Аляску и, беспрепятственно миновав покрытые ледниками земли, спуститься к западному побережью Америки, начался 14 тысяч лет тому назад. Археологические находки свидетельствуют о том, что люди продолжали жить на затопленном ныне побережье по крайней мере еще 10 тысяч лет тому назад. Более того, археологические находки, сделанные на западном побережье Южной Америки, свидетельствуют, что 11 тысяч лет тому назад народ, известный нам как собиратели на прибрежной полосе, мог проделать путь из Берингии в более южные широты на судах, предназначенных для каботажного плавания. Елена Стариовская убежденно доказывает, что прибрежные западные маршруты позволяют дать географическое объяснение факту позднейшего распространения группы В. Примерно 17 тысяч лет тому назад американская линия-родоначальница В4 широко распространилась на островах Восточной Индонезии, куда, предположительно, попала вместе с причалившими на лодках мореходами, и далеко на юге — на юго-западных островах Тихого океана (см. главу 6). Если маршруты каботажного плавания позволяли огибать ледники, то это достаточное основание для того, чтобы сделать одно неожиданное предположение — даты возникновения ледового барьера и коридора никак не могли повлиять на историю эпохи Кловис. Когда я навестил Кнута Фладмарка из Университета Симона Фразера, штат Ванкувер, я был удивлен, обнаружив, что он остается сторонником консервативной концепции «Первый Кловис», которая почувствовала себя несколько моложе, когда получила возможность опереться на костыль в виде теории каботажного плавания.
Заселение Америки могло предшествовать эпохе Кловис
Можем ли мы восстановить генетическую картину заселения Америки? Думаю, можем. Хотя минувшее оставило нам лишь разрозненные и отрывочные свидетельства, я считаю, что альтернативная гипотеза каботажных плаваний вдоль западного побережьяреальна и неопровержима, однако она описывает лишь параллельный маршрут заселения Америки, который не являлся единственным и основным. Группа Б4, например, была распространена на западном побережье Канады уже 12—15 тысяч лет тому назад, и скорее всего — подобно предкам бурых медведей, живущих на западном побережье, — предки древних канадцев уже перед Последним ледниковым максимумом заселили просторы Берингии, где жили бок о бок с представителями других линий (А, С, D и X). Существует и альтернативная гипотеза, однако она кажется мне менее убедительной, ибо основана на предположении, что кластер В4 лишь по окончании ПЛМ покинул Азию и отправился в плаванье вдоль побережья. Дело в том, что во время плавания из Восточной Азии к берегам Британской Колумбии людям пришлось бы преодолевать чудовищные преграды, которые воздвиг на их пути Последний ледниковый максимум, которые еще долго служили напоминанием о нем, эти преграды, например Алеутский ледниковый панцирь, представляли собой гигантские ледниковые плиты. Итак, я остаюсь сторонником гипотезы, в соответствии с которой вторжение предков американских аборигенов в Берингию и Америку произошло до ПЛМ, а следовательно, и до эпохи Кловис. Генетическая картина заселения Америки линиями-родоначальницами А, В, С, D и X представляет собой множество параллельных маршрутов, коими перед ПЛМ воспользовались группы первопроходцев, прародиной которых, в конечном счете, следует считать северо-восточные степи Евразии и восточное побережье Азии. Кланы переселенцев значительно отличались друг от друга. Одни из них, возможно, были похожи на европейцев, другие на людей племени айну или на жителей некоторых островов Тихого океана. Кроме того, среди древних американцев были и предки человека из Уайзард-Бич, в чьем облике прослеживается сходство с северной монголоидной расой, а также с американскими аборигенами недавних времен. Возможно, передвигаясь вдоль побережья и занимаясь собирательством на прибрежной полосе, этот тип человека смог завоевать просторы Южной и Северной Америки быстрее других групп аборигенов. Линия В4 (или одна, заслуживающая особого внимания разновидность линии В4), по всей вероятности, проникла на американский континент на заре его заселения и устремилась вниз, в Южную Америку, где в отличие от относительно молодых североамериканских вариантов она столь же стара, как и другие линии-родоначальницы. Затем начался Ледниковый период. Люди, запертые среди ледников на севере Берингии, столкнулись с невыносимыми лишениями и нуждой. Однако предки народов, говорящих на языках на-дене и эскимосско(инуитско)-алеутской групп, то есть представители генетического кластера А, выдержали схватку с лютой стужей и заявили о себе как о первоклассных рыболовах. Группа В4, возможно, пережила Ледниковый период на западном побережье, а затем в процессе вторичного заселения вместе с бурыми медведями проникла в центр континента — а может быть, линия В4 получала свежие гены из Азии: следуя сперва вдоль береговой линии вверх, она обогнула Тихий океан и направилась вниз — к Америке. Однако, если вести отсчет от Последнего ледникового максимума, то, насколько нам известно, ни Америка, ни Аляска (а это все, что осталось от погибшего континента Берингия) не получали сколько-нибудь значительных генных вливаний из Азии, во всяком случае, так было до тех пор, пока не появился Лейф Эрикссон и новая приливная волна, которую всколыхнул Христофор Колумб и его «открытие» Нового Света, не обрушилась со всей силой на берега Америки и не наводнила ее новыми людьми и новым генным материалом.
э п и л о г
Несколько миллионов лет тому назад холодные и засушливые климатические условия, установившиеся в ту эпоху, практически уничтожили прежнюю среду обитания приматов, живших в лесах. Некоторое время спустя эволюция сделала первые шаги в сторону появления того двуногого существа, обладающего крупным мозгом, которое мы сегодня называем Homo sapiens. И хотя палеонтологи все еще спорят о том, к какому именно времени следует отнести первые свидетельства бипедализма, обнаруженные в ископаемых останках этих существ, и когда — 5 или 7 млн. лет тому назад — предки этих существ разошлись по пути эволюции с предками шимпанзе, самое главное заключается в том, что этот шаг был сделан. И пусть у первого прямоходящего вида приматов Australopithecus мозг был лишь немногим больше, чем у шимпанзе, он начал быстро увеличиваться у новых видов. Вместе с интенсификацией холодной и засушливой фазы климата, имевшей место чуть более 2 млн. лет тому назад, началось активное развитие и увеличение объема мозга, свойственное только человеку и представителям его близкородственного вида — так называемого Paranthropus. Невиданно быстрые темпы этих исходных изменений никогда более не были повторены.
Биология и культура: проблема коэволюции
Новые возможности, которые обрели представители этих двух видов гоминидов, обеспечили им массу преимуществ в борьбе за выживание в тот необычайно суровый период. Новые навыки и черты поведения, по всей вероятности, не были обусловлены некими особенностями питания, поскольку рационы питания у этих двух видов были весьма различными, но представители и того и другого в равной мере использовали преимущества, предоставляемые крупным мозгом. На том этапе эволюции человека объем нашего мозга увеличивался исключительно быстрыми темпами, тогда как тело менялось малосущественно. Скорость увеличения объема мозга достигла своего максимума примерно во время появления вида Homo, что, наряду с данными изучения Paranthropus, свидетельствует о том, что наши общие предки-гоминиды уже обладали теми уникальными особенностями, которые впоследствии и повлекли за собой активную эволюцию. Наиболее вероятным претендентом на роль «уникальных особенностей», обусловленных крупным мозгом, является тот самый дар, который выделяет нас, людей, среди всех прочих живых существ, а именно — дар речи. Однако, поскольку в доисторическую эпоху не было кассетных манитофонов, наиболее реальным материальным свидетельством культурных изменений следует считать тот факт, что человек с самого начала стал создавать всевозможные каменные орудия. Несмотря на быстрое увеличение объема мозга, прогресс в области создания каменных орудий был мучительно медленным, растянувшись на добрый миллион лет, и новые технические достижения отнюдь не были автоматическим следствием появления очередного вида человека. Так, каменные орудия ашолийского типа были изобретены представителями африканского вида Homo erectus еще 1,4 млн. лет тому назад, но это произошло много тысячелетий спустя после того, как их родичи, предки азиатского вида erectus, покинули свой родной континент. Таким образом, ашолийская технология не присутствовала в Евразии вплоть до времени последнего исхода.
Люди как вид млекопитающих
История человечества за последние 2,5 млн. лет характеризуется резкими скачками и прорывами в области технологии и освоения мира, за которыми следовали длительные периоды спада и стагнации. Обычная точка зрения на сей счет сводится к тому, что наши предки постоянно поднимались все выше и выше по ступеням лестницы эволюции и развития, каждый новый этап которой был недоступен и недостижим для их непосредственных предков. Эта точка зрения исходит из предположения о том, что на всем протяжении нашей истории мы, люди, были хозяевами собственной судьбы и что пределы заселения новых территорий определялись лишь интеллектуальным и созидательным потенциалом того вида, представители которого доминировали на планете в ту или иную эпоху. Подобный оптимистический взгляд на возможности нашего интеллекта и самоутверждения явно преувеличен. Так, один из первых видов человека, Homo erectus, достаточно быстро совершил исход из Африки, отправившись заселять всю Евразию. Разумеется, он был далеко не первым видом приматов, проделавшим подобную миграцию, как показывает пример тех же орангутангов и гиббонов, расселившихся по всей Юго-Восточной Азии. Точно так же представители вида erectus не были и последними гоминидами, совершившими подобный исход прежде нас, людей. По сравнению с более ранними видами человека наши предки были всего лишь одними из мигрантов-землепроходцев, которые достигли Америки и островов Океании. Расселение человека было как бы зеркальным отражением миграции других видов млекопитающих, будучи обусловлено в основном климатическими и географическими факторами. Оно происходило по двум давно проторенным маршрутам. Мы, люди, отличаемся от других крупных млекопитающих, но не слишком отличаемся от грызунов с точки зрения обширности и многообразия районов обитания, освоенных нами, а также высокой численности населения. В этом контексте мы отличаемся от грызунов разве что тем, что, поскольку мы гораздо крупнее их, нам требуется гораздо больше пищи в расчете на каждую особь, то бишь душу. Массовые миграции человека из Африки по северному и южному маршрутам всегда определялись климатическими циклами и наличием продовольственных ресурсов. Наступление и окончание циклов оледенения не только открывало и закрывало врата исхода из Африки, но и вызывало периодические сокращения численности населения, ставя один вид на грань вымирания и одновременно побуждая новый, обладающий большим объемом мозга вид остаться на своей древней прародине или попытать счастья в дальних краях.
Увеличение объема нашего мозга давно остановилось
Около 1,2 млн. лет тому назад мозг представителей вида Homo rhodesiense, обитавшего в Африке, составлял всего 6% от объема мозга современного человека. Около 300 тысяч лет назад «механизм» по увеличению объема мозга, действовавший под влиянием климатических факторов, достиг уровня, когда мозг тогдашнего вида человека на целых 11% превышал объем мозга современного человека. С тех пор наши мозг и тело несколько уменьшились в объеме и пропорциях. Циклы оледенения продолжали сменять друг друга, совершая свою глобальную работу, но, за исключением косметических изменений в пропорциях тех или иных членов, очертаниях надбровных дуг и форме черепа, эволюция внешнего облика рода человеческого в наши дни продвигается вперед черепашьим шагом. Быть может, здесь, как и в области автомобилестроения, действует закон эволюционного возврата к малым формам, когда становится экономически нецелесообразным создавать модели со все более и более крупными двигателями. Как настоятельно утверждают антропологи Салли Мак-Брирти и Алисон Брукс, именно в ту эпоху был достигнут порог эволюции внешнего облика вида Homo sapiens. С этой точки зрения люди анатомически современного типа представляют собой особую позднейшую расу, которая развилась в Африке на основе более древнего, так называемого архаического Homo sapiens после окончания очередного цикла оледенения, имевшего место около 150 тысяч лет тому назад. Существуют доказательства того, что архаические типы Homo sapiens также покинули Африку и задолго до нас отправились заселять просторы Евразии.
Эволюция культуры — решающий фактор
В рамках картины, которую нарисовали Мак-Брирти и Брукс, все отличительные особенности поведения современного человека можно проследить в ретроспективе вплоть до эпохи Среднего Каменного века в Африке. Это отнюдь не означает, что тогда, около 300 тысяч лет тому назад, произошло нечто вроде Большого взрыва в области техники и технологии, создававшейся человеком поначалу медленно, а затем все более и более быстрыми темпами. Первые признаки прогресса были едва заметными и появились достаточно поздно, но по мере того, как с каждым последующим поколением происходило накапливание знаний и возникал устойчивый интерес к ним, эволюция культуры начала развиваться опережающими темпами, оставив далеко позади биологическую эволюцию. Посмотрим на эту тему с несколько иной точки зрения. Если эволюция культуры действительно началась около 300 тысяч лет тому назад вследствие активной генетической эволюции, это означает, что различия между нами и представителями архаического вида Homo sapiens обусловлены всего лишь культурными факторами, а интеллектуальный потенциал вполне мог бы позволить этим людям совершить полет на Луну, живи они среди нас.
Генетическая история: что она нам дает
Итак, что могут сказать нам о нас самих и наших далеких предках новые генетические методы, в отношении которых многие палеоантропологи и археологи настроены, мягко говоря, скептически? Отвечаю: очень и очень многое, как это, смею надеяться, показала моя книга. Генетическая палеонтология вносит ясность в область, в которой царит почти средневековая неразбериха. Обмеры черепов и анализ их форм, а также обстоятельное изучение и оценки возраста каменных орудий создают неадекватную картину древнейшей эпохи истории человечества. Помимо крайней малочисленности останков человеческих скелетов эпохи палеолита, существует громадное множество вариантов форм человеческих черепов. Использование формы черепов в качестве системы маркеров для определения этнической принадлежности вызывало еще большую путаницу в результате неадекватного питания и малорослости у представителей традиционных сельскохозяйственных общин, а также в результате присутствия инородных генетических примесей у различных групп населения. Все это позволяет без особых трудностей выявить слабые места в реконструкции миграций человека, базирующейся на подобных замерах. Поскольку каменных орудий сохранилось несравненно больше, чем человеческих останков, картина, получаемая в итоге, всегда грешит односторонностью и может поведать нам разве что об уровне технической культуры создателей таких орудий, а не о том, кем были эти люди, где пролегали маршруты их миграций и каково их биологическое происхождение. Двумя наиболее выразительными примерами противоречий во взглядах антропологов, обусловленных применением неадекватных средств анализа, являются, во-первых, гипотезы мультирегионалистов и теории сторонников исхода из Африки, и, во-вторых, вопрос о том, существовали ли межвидовые браки между неандертальцами и представителями первых европейцев современного типа — кроманьонцами. Классическая генетика, основанная на использовании простейших протеиновых маркеров, дает картину столь же неясную, как и обычные краниометрические обмеры черепа. И лишь выявление прямой, без всяких межвидовых «адьюльтеров», передачи генов Адама и Евы из поколения в поколение в корне изменило картину происходящего. Достоверные генетические древа для Y-хромосом и мтДНК современного человека позволяют проследить его эволюцию вплоть до общего предка всех людей современного типа, жившего как минимум 200 тысяч лет тому назад, и выявить его миграцию из Африки, происшедшую менее 100 тысяч лет тому назад. Представители этой новой линии достаточно быстро вытеснили все предыдущие генетические линии человека, в том числе и неандертальцев. Разумеется, сама возможность межвидовых браков между представителями архаических ветвей и современными Homo sapiens в принципе не исключена, однако никаких убедительных следов подобных контактов в мужских и женских генетических линиях современных людей не обнаружено. Таким образом, если такие межвидовые связи и имели место, они носили малосущественный характер и все архаические линии давно вымерли. Некоторые ученые полагают, что следы таких межвидовых контактов могут таиться в глубине нашего необъятного нуклеарного генома, но ключевыми все-таки остаются слова «не обнаружено». Принимая во внимание тенденцию к перекомбинированию и созданию в каждом поколении все новых и новых сочетаний, очень трудно построить абсолютно точные деревья для нуклеарных генов, так что некоторые из их ветвей всегда будут вызывать вопросы.
Первый исход в Южную Азию
В этой книге, опираясь на всевозможные генетические свидетельства, подтверждаемые различными археологическими материалами из целого ряда древних стоянок, я отстаиваю точку зрения о том, что успешный исход из Африки был всего один, что он имел место около 80 тысяч лет тому назад и проходил по южному маршруту. Его участники-мигранты около 74 тысяч лет тому назад достигли Малайского полуострова, а около 70 тысяч лет тому назад — берегов Австралии. Участники более раннего исхода на Левант по северному маршруту около 90 тысяч лет тому назад к тому времени давно вымерли. И хотя сохраняется некая вероятность позднейшей колонизации Австралии из Восточной Индонезии людьми архаического вида, наиболее ранними останками человека являются останки людей современного вида, и на сегодняшний день нет никаких сколько-нибудь убедительных свидетельств присутствия генетических линий Адама и Евы архаического вида у ныне живущих аборигенов Австралии. Во всех этих данных обращает на себя внимание ключевая роль Южной Азии (в частности — Индии, Пакистана и района Персидского залива), служившей как бы источником и отправной точкой многих путей расселения выходцев из Африки. Датировка времени первой волны колонизации Восточной и Центральной Азии людьми современного типа носит весьма проблематичный и спорный характер, что объясняется неточностью методов датировки черепов и наличием фактов о позднейшем замещении монголоидами, но если уточненная датировка люйцзяньского черепа (около 68 тысяч лет тому назад) будет признана достоверной, можно будет говорить о том, что Южный Китай был заселен в то же время, что и Юго-Восточная Азия. А если люди современного типа проникли в Юго-Восточную Азию еще до катастрофического извержения вулкана Тоба, то объяснением резкой генетической паузы между Индией и Дальним Востоком вполне можно считать колоссальное облако вулканического пепла, накрывшее Индию около 74 тысяч лет тому назад.
Путь на север
Европа и Западная Азия были заселены значительно позже, около 50 тысяч лет тому назад, когда в мире наступило глобальное потепление, открывшее сухопутный коридор от района побережья Персидского залива к землям нынешнего Восточного Средиземноморья. Археологические данные свидетельствуют о том, что Центральная Азия была заселена практически в то же время, что и Западная Евразия, то есть около 40—50 тысяч лет тому назад. Целая группа генетических линий, и сегодня присутствующих в Северной и Центральной Азии, являются аргументами в поддержку этой гипотезы и свидетельствуют о прямом пути продвижения на север с территории Индийского субконтинента до западных склонов Гималаев. Некоторые из этих северо- и центральноазиатских линий, по всей видимости, восходят к генофонду древнейших собирателей на прибрежной полосе в Юго-Восточной Азии, которые предположительно могли проделать долгий кружной путь и обойти Гималаи с запада. Прародина так называемого монголоидного типа внешности, если вообще можно говорить о каком-то едином комплексе признаков, остается неизвестной и спорной, хотя большинство имеющихся дентологических данных указывают на южные регионы. Эта простейшая точка зрения на генетические и географические свидетельства хорошо согласуется с мнением о том, что где-то в Центральной Азии (скорее всего — к северу от Гималаев) существовал особый регион, где в результате миграций, вызванных резким похолоданием и ухудшением климата, у коренных жителей Юго-Восточной Азии начали формироваться характерно «монголоидные» черты внешнего облика.
Последний ледниковый период: беженцы и пионеры
Наиболее драматичным после извержения вулкана Тоба событием глобального масштаба, повлиявшим на судьбы человечества, явился так называемый Последний ледниковый максимум (ПЛМ), наступивший около 18 тысяч лет тому назад. Для Африки это означало быстрое расширение районов пустынь и очередное резкое сокращение численности населения. Для Европы, региона, ситуация в котором изучена наиболее хорошо, наступление ледника повлекло за собой массовое перемещение населения с севера в южные районы, служившие прибежищами для беженцев. Это были в первую очередь Страна Басков, Италия и Балканы, а также Украина в Восточной Европе. Генетическая картина подтверждает археологические данные о резком сокращении численности населения в Западной и Центральной Европе. Согласно одному археологическому исследованию, остатки охотников и собирателей эпохи палеолита были в последние 10 тысяч лет в большей или меньшей степени вытеснены и замещены новым этническим материалом — земледельцами и скотоводами из Анатолии и Леванта. В отличие от этих данных, материалы генетических исследований показывают, что большинство материнских и отцовских генетических линий у жителей современной Европы происходят от предков, живших в Европе еще в эпоху палеолита, которые впоследствии расселились из своих прибежищ на юге до наступления периода неолита. Гораздо меньше нам известно о влиянии Последнего ледникового максимума на население Азии. Некоторые ученые говорят даже о полном исчезновении населения огромных степных пространств Центральной Азии, заселенных некогда охотниками на мамонтов эпохи Верхнего палеолита. Археологические данные показывают, что там, в Центральной Азии, тоже было как минимум одно прибежище, находившееся на юге Сибири, где люди жили даже в эпоху Великого Оледенения. Уцелевшие генетические линии показывают нам, что немногочисленные группы населения сохранялись там даже в эпоху наиболее жестокого холода. Одно можно утверждать с полной уверенностью: даже если некоторые жители решили остаться на прежних местах, большинство все же наверняка воспользовалось любым шансом на спасение от усиливающейся стужи в степях, как только он им предоставился. В отличие от Европы, где зона спасения на юге была ограничена побережьем Средиземного моря и Сирийской пустыней, в Центральной и Северной Азии существовал целый ряд маршрутов исхода в более теплые регионы с умеренным климатом: на запад, на крайнем западе Евразии, на северо-восток, в Берингию и далее в Америку, на восток, в Корею и Японию и, наконец, на юго-восток, в Южный Китай и Юго-Восточную Азию. Последний из этих путей выглядел особенно заманчивым, поскольку, в отличие от всех прочих маршрутов, площади земель, пригодных для обитания, в Юго-Восточной Азии постоянно увеличивались в результате неуклонного понижения уровня Мирового океана, а прибрежные территории не были ограничены панцирем льдов. В главе 6 я уже рассматривал это расселение в доледниковый период в свете имеющихся генетических и археологических свидетельств. Заселение Северной и Южной Америки всегда было крайне болезненной темой для американских археологов. Верные приверженцы старой школы готовы стоять насмерть, отстаивая ортодоксальную гипотезу о Первом Кловисе, тогда как другие на основании недавних находок убеждены в том, что миграции происходили еще в доледниковый период, а бедные лингвисты не в силах поддержать ни ту ни другую сторону в этой академической битве. Что же касается генетиков, то они, напротив, сходятся во мнении, что большинство, если не все, генетические линии-родоначальницы проникли в Северную Америку еще до Последнего ледникового максимума, между 22 и 30 тысячами лет тому назад. Первая из этих датировок и в хронологическом, и в генетическом отношении хорошо согласуется с исходом мигрантов в доледниковую эпоху, о котором речь шла в главе 5. Однако далеко не все линии-родоначальницы американских туземцев проникли на континент из северных районов Центральной Азии: как минимум одна из них является выходцем из Японии, и вполне вероятно, что существовали и другие источники. Разнобой в данных замеров и датировках также свидетельствует о том, что далеко не все первые обитатели Америки были выходцами с общей азиатской прародины; они представляли собой скорее пеструю смесь монголоидов и кавказоидов, а также толстокостных типов, характерных для западного побережья Тихого океана. Как и в Европе, в Америке в послеледниковую эпоху тоже происходили повторные расселения первоначальных палеолитических линий-родоначальниц, затронувшие в первую очередь Берингию и Крайний Север Америки. От них произошли своеобразные этнические группы жителей Арктики и субарктических районов и западного побережья Канады.
Считаю ли я, что читатели мне поверят, а если да, то насколько?
Сама попытка описать генетическую историю рода человеческого в эпоху около 10 тысяч лет тому назад в книге, число глав в которой даже меньше, чем в известном труде Джулиана Бэрнса «Мировая история в 10 1/2 главах», — задача весьма амбициозная. Ведь неизбежно существуют регионы, такие, например, как Восточная и Северо-Восточная Азия, где следы миграций человека — кости, камни и даже гены — неуловимы и практически незаметны. Перед лицом множества трудов по реконструкции доисторического этапа истории человечества в эпоху палеолита я скорее могу заслужить упрек в излишней смелости домыслов, чем в недостаточной информированности и верхоглядстве. И тем не менее я оказался в весьма почтенной компании. Целый ряд работ, созданных авторскими коллективами и опубликованных за последние два-три года, предлагают подробные генетические карты мира, испещренные всевозможными стрелками и указателями. Работы эти написаны специалистами по генетике, археологии и палеонтологии, и карты, представленные в них, весьма существенно отличаются друг от друга, ибо генетическая история человечества и сегодня по большей части остается малоизведанной областью. Помимо смелости и даже дерзости, тенденция к мультидисциплинарному синтезу является здесь вполне естественным подходом, поскольку ни одна из дисциплин не охватывает в полной мере всего многообразия истории. В некоторых разделах реконструкции, в частности — в области датировки, синтез опирается на взаимодополняющие данные из разных дисциплин, которые объединяются в логически единое целое, представляющее собой нечто большее, чем простая сумма его составляющих. Естественно, при этом возникает опасность путаницы и блуждания по замкнутом кругу, поскольку некоторые аспекты могут характеризоваться случайными или ложными совпадениями. В качестве альтернативы и одного из способов защиты моих гипотез (хотя подобная апология уже явно запоздала) я хотел бы сказать несколько слов о достоверности самих свидетельств: о том, где именно достигнут генетический консенсус, где такого консенсуса пока нет, но, насколько мне известно, шансы достичь его весьма высоки, и, наконец, где доказательства слишком шатки и способны вызвать разве что недоумение. В противном случае есть все шансы, что даже самые убедительные доказательства, представленные мной, будут отметены прочь той же метлой, что и ничего не стоящие пустяки. Итак, исход из Африки — вопрос практически решенный. Сегодня найдется очень мало генетиков, работающих в данной области, которые еще сомневаются в том, что позднейшие расселения людей современного типа, выходцев из Африки, привели к вытеснению и замещению архаических подвидов Homo sapiens на территории всего остального мира. Да, возможность существования нескольких генетических следов древнейших линий, не связанных с исходом из Африки, сохраняется, но она носит чисто теоретический характер. Генетическая логичность гипотезы об одном-единственном исходе из Африки практически неопровержима, хотя большинству генетиков и археологов еще только предстоит признать ее правоту. Южный маршрут исхода из Африки через устье Красного моря уже давно был признан зоологами как наиболее удобный сухопутный маршрут миграций многих видов млекопитающих в эпоху плиоцена, но лишь недавно он стал всерьез рассматриваться как один из параллельных маршрутов исхода человека современного типа. Дело в том, что главным маршрутом исхода предков европейцев по-прежнему считается северный путь, через Суэц. В этой книге я предлагаю синтез генетических и других данных. И все они подтверждают, что в древности имел место один-единственный исход по южному маршруту из Эритреи в Йемен и что все неафриканские мужские и женские генные линии произошли от одних и тех же общих линий-родоначальниц, возникших в Южной Азии (или, по крайней мере, неподалеку от южного маршрута). Я считаю генетическую логичность этих материалов серьезным основанием подобного синтеза, и именно на них базируется все построение моей реконструкции диаспоры[462] древнего человека. Понятно, что «выбор» исходной точки определил все последующие маршруты и пути расселения наших предков и ближайших сородичей. Отслеживание подобных путей возможно только в результате выраженной специфичности регионального распределения генетических ветвей. Географическая конкретность распределения ветвей генного древа мужских и женских линий, представленного в этой книге, являет собой разительный контраст по сравнению с крайне запутанной картиной межрегионального распределения ветвей генетических линий, наблюдаемой в более ранних генетических исследованиях. Уровень распределения линий по странам и континентам сам по себе является убедительным свидетельством того, что некогда все они находились в неком общем «доме», но затем первопроходцы-пионеры, по крайней мере, — в эпоху до Последнего ледникового максимума, способствовали активному продвижению и распространению некоторых из них. Консервативный аспект нашей генетической предыстории также предлагает частичное объяснение того факта, что, когда мы видим человека, мы обычно можем сказать, откуда он и на каком континенте жили его предки, и можем перечислить некоторые характерные признаки, которые некоторые из нас по-прежнему называют «расовыми отличиями». По мере продвижения далее по этому пути, на север и восток из Индии, генетическая реконструкция маршрутов приобретает все более и более гипотетический характер, причем одни из этих маршрутов прослеживаются более точно, чем остальные. Особенно легко проследить путь мигрантов в Австралию. Древнейшая дата колонизации Австралии, если ее достоверность подтвердится, также позволяет считать продвижение собирателей на прибрежной полосе по всему побережью Индийского океана через Юго-Восточную Азию единственно возможным маршрутом. Между тем глубокие генетические различия между аборигенами Новой Гвинеи и Австралии со всей определенностью свидетельствуют о том, что в древности там имели место как минимум две разные волны колонизации этих территорий. Позднейшее заселение Европы и Леванта из района Персидского залива, а не из Северной Африки — это моя основная гипотеза, базирующаяся на самой структуре генетического древа. Она объясняет, почему в Западной Азии и Европе, единственных неафриканских регионах мира, практически отсутствует азиатский суперклан М. Позднейшая дата колонизации Леванта объясняется засушливым климатом, существовавшим на планете до 50 000 г. до н.э., в результате чего коридор между районом Персидского залива и Левантом оказался закрытым. Однако затем заметное улучшение климата, начавшееся после 50 000 г. до н.э., вновь открыло путь, ведущий в район Плодородного Полумесяца, сделав возможной миграцию на северо-запад — на Левант и Кавказ и далее, в Европу. Остальные версии о заселении Евразии носят гипотетический характер. Описания колонизации Восточной Азии по пути миграции собирателей на прибрежной полосе, а также в результате пунктирных исходов из Центральной и Северной Азии по обе стороны Гималаев — это всего лишь наиболее вероятные с географической точки зрения объяснения имеющихся генетических данных. Концепция заселения прибрежных низменностей Восточной Азии, на которые в эпоху Последнего ледникового максимума хлынул поток переселенцев из степей Центральной Азии — это тоже моя гипотеза, способная кое у кого вызвать удивление. Но дело в том, что пока что не существует другого убедительного объяснения существующего распределения генетических линий и типов внешности, отмечаемого в Восточной Азии, и подтверждаемые археологическими свидетельствами резкие изменения в Японии и Корее, имевшие место на пике Последнего ледникового максимума, являются весьма показательными для проникновения на эти территории культур Верхнего палеолита из Центральной Азии. Что касается заселения Северной и Южной Америки, то я предпочитаю согласиться со сложившимся консенсусом генетиков о проникновении всех генетических линий на американский континент во время Последнего ледникового максимума или непосредственно перед ним, рассматривая некоторые особенности поведения человека с точки зрения господствующей археологической концепции Кловиса.
Несколько слов о датировке доисторического периода
Точность датировок в решающей мере зависит от того, какой метод используется для их получения. Дело в том, что даже радиоуглеродный метод датировки заключает в себе системные погрешности. В некоторых ситуациях датировки, полученные путем расчетов с помощью молекулярных часов, для генетических линий очень близко согласуются с датировками, основанными на археологических свидетельствах. Митохондриевые молекулярные часы пока что находятся в стадии настройки, а датировки по Y-хромосомам остаются весьма приближенными. Более того, поскольку сам принцип действия молекулярных часов зависит от нечастых случайных мутаций, для получения более или менее надежных данных требуется провести большое число замеров (т.е. множество анализов у представителей разных генетических типов). Это означает, что многие из недавних датировок археологических объектов, полученных при использовании радиоуглеродного метода, являются гораздо более точными. В то же время в датировках по радиоуглеродному методу археологических объектов, относящихся к периоду Последнего ледникового максимума, присутствует множество погрешностей, а для эпохи старше 40 тысяч лет тому назад радиоуглеродный метод и вовсе представляется ненадежным. Между тем для генетических часов пресловутого потолка в 40 тысяч лет просто не существует. Единственными по-настоящему надежными датировками, точность которых связана с колебаниями климата, являются недавно разработанные методы датировки по древесным кольцам и аналогичному варьированию толщины слоев в образцах льда, полученных при бурении скважин в ледниковых шапках Гренландии, позволяющих охватить период 100 тысяч лет и более. Эти пробы дают точные и детальные датировки для колебаний климата. Так, например, теперь мы можем установить даты частых потеплений или промежуточных циклов, которые влияли на растительный мир и миграции крупных масс населения. Наиболее достоверно датируемым природным катаклизмом в обозримом прошлом человечества до Последнего ледникового периода является катастрофическое извержение вулкана Тоба на острове Суматра, происшедшее около 74 тысяч лет тому назад. Вулканический пепел от извержения Тоба обнаружен и в пробах льда, полученных на Гренландии, и в пробах грунта морского дна в Индийском океане, взятых с подводной лодки; таким образом, он является точным маркером для датировки. Могу представить, какая ужасная участь постигла жителей Малайзии, живших в ту эпоху в районе стоянки Кота Тампан, где были обнаружены каменные орудия, засыпанные толстым слоем вулканического пепла от извержения вулкана Тоба. Эти орудия, бесспорно, представляли собой древние артефакты, и вулканический пепел обрушился на них прямо с неба около 74 тысяч лет назад. Однако, несмотря на бытующее мнение о том, что орудия, найденные на стоянке Кота Тампан, представляют собой изделия людей современного типа, они вполне могли быть созданы и представителями более архаических видов человека, поскольку на самой стоянке не было обнаружено человеческих останков, позволяющих определить, кем были создатели этих артефактов и когда они жили. Единственными надежно датируемыми останками человека современного типа, найденными в этом регионе и имеющими такую древность, являются знаменитый люйцзяньский череп и фрагменты скелета из Южного Китая. Материалы по датировке древнейших останков человека современного типа, найденных в Восточной Индонезии (Флорес), пока еще не опубликованы. У меня есть целый ряд причин, дающих основание полагаться на столь шаткие аргументы, как датировка каменных орудий. Это прежде всего сама логика колонизации Австралии водным путем в эпоху около 70 тысяч лет тому назад, а во-вторых, заметное увеличение генетических датировок линий за пределами Африки, указывающих именно на это время. Следующий период понижения уровня океана, открывавший путь для колонизации Австралии, мог иметь место лишь около 50 тысяч лет тому назад, но эта дата никак не согласуется с другими имеющимися свидетельствами. Главная проблема здесь связана с тем, что крайне трудно убедить большинство австралийских археологов в том, что Австралия была заселена человеком около 60 тысяч лет тому назад, не говоря уж о еще более древней дате — 70 тысяч лет тому назад. Ключевая проблема получения надежной датировки для расселения человека современного типа за пределами Африки в эпоху старше 45 тысяч лет тому назад обусловлена пресловутым радиоуглеродным «потолком». Если полагаться исключительно на датировку по радиоуглеродному методу, получается, что человечество начало свой путь на Земле немногим ранее 40 тысяч лет тому назад. Однако с помощью других методов были получены датировки различных памятников в Австралии, восходящие к значительно более раннему времени (примерно в районе 60 тысяч лет тому назад), но они сразу же вызвали столько проблем, что оказалось проще вообще отбросить эти даты. Мы тоже можем допустить, что дата активной колонизации Австралии совпадает с «потолком» радиоуглеродного метода, но это было бы заведомо ошибочным постулатом. Безопаснее всего, разумеется, сказать, что более или менее точная дата первой волны колонизации вообще неизвестна. В рамках своего гипотетического синтеза я отдал предпочтение наиболее ранней дате, которая согласуется с датировкой артефактов из Кота Тампан и периодом минимального уровня океана, нежели дате, которая на сегодняшний день представляется более точной, но вскоре практически наверняка окажется ошибочной. По мнению некоторых ученых, извержение вулкана Тоба повлекло за собой массовое вымирание жителей в глобальном масштабе в результате так называемого эффекта «ядерной зимы», наступившей вслед за ним. Я учел этот фактор в своей реконструкции. Основная масса вулканического пепла обрушилась на Индию, и именно там имела место массовая гибель всего живого, поскольку колоссальное облако пепла, взметнувшееся в небо после извержения, было подхвачено ветром и направилось с Суматры на северо-запад, прямо над водами Индийского океана. Эта природная катастрофа может служить объяснением того, почему большинство индийских подгрупп линии М не встречаются более нигде в Азии, а датировки времени повторного расселения клана М парадоксальным образом являются в Индии более поздними, чем в любом другом районе Азии или в Австралии.
Что же нас ждет в будущем?
На мой взгляд, наши перспективы выглядят далеко не столь радужными, как это представляется некоторым. Мы, как все прочие виды живых существ, которые испытывают на себе законы эволюции и вынуждены участвовать в борьбе за выживание, не только оказываем воздействие на окружающую среду и стараемся контролировать происходящие в ней изменения, но и сами подвержены их влиянию — независимо от того, чем они вызваны: цикличностью ли ледниковых периодов или нашей собственной расточительностью и хищническим отношением к ресурсам планеты. Если соотнести себя с большой шкалой времени, то одним из уроков прошлого следует признать периодически повторяющийся ледниковый максимум. Придет время, и снова наступит Великое Оледенение. Достигнув своей кульминации, оно скует холодом все живое, оставив для жизни лишь узкий перешеек, на котором будут бороться за место под солнцем оставшиеся роды и виды. Нам вряд ли удастся в полной мере оценить, насколько велико влияние ледниковых периодов на судьбу нашей планеты. Углубляясь мысленно в далекое прошлое, мы стремимся оценить также следствия глобального потепления, которое, как нам кажется, оказывает на земную жизнь более существенное влияние, чем следующий за ним в перспективе ледниковый максимум. Однако пришла пора заключительных аккордов и лаконичных фраз. Генетическое разнообразие — вот то ключевое понятие, рассмотрению которого посвящена эта книга. Нашими проводниками в глубину времен стали медленно эволюционирующие, но многообразные линии митохондриевых ДНК и Y-хромосом. Однако мы еще не осветили должным образом того факта, что восстановительный период после последнего периода оледенения, ограничившего сферу обитания узким перешейком, все еще не завершен, а совокупное генное разнообразие представлено узким спектром вариаций. Лишившись генных богатств и расового многообразия, мы стали беззащитными перед лицом инфекций, которые грозят нам новыми пандемиями и готовы поглотить каждого, кто посмел затеряться среди прохожих на улице или вступить в общение с группой людей.
Многообразие, многообразие, многообразие
Однажды со мной произошел весьма забавный случай. Когда я работал в Гонконге, моей коллегой и старшим лектором оказалась одна почтенная и очаровательно искренняя дама — кардиолог-педиатр с Тайваня. Она как-то спросила меня, вижу ли я разницу между убийством тигра ради его шкуры, костей и мяса и забоем скота. Этот вопрос не показался мне вздорным. Моя коллега подкрепляла свои убеждения тем, что отказалась от лекарственных препаратов, в состав которых входят кости тигра, ибо испытывала жалость к этим зверям, и сердце ее наполнялось скорбью при мысли, что убито такое красивое животное. Короче говоря, она была против промышленного убийства. Ей была чужда навязываемая Западом философия, внушающая, что акт убийства может рассматриваться как нечто недозволенное и аморальное в отношении одного крупного животного и не вызывать протеста в отношении другого. Она не могла осознать, какими законами логики и здравого разума руководствуются те, кто считают, что охранять крупных животных важнее, чем, скажем, грызунов, и почему лишь красивые млекопитающие признаются полезными для человека. В поисках достойного и философски взвешенного ответа напоставивший меня в тупик вопрос я почесал затылок: мне не хотелось пускаться в обсуждение проблемы, вправе ли мы вообще распоряжаться жизнью животных, кроме того, сводить объяснение к эстетическому восприятию столь внешне привлекательных диких кошек мне тоже не хотелось. И вот, наконец, я подумал, не удастся ли мне убедить ее в том, что существует один важный объективный критерий, которым мы руководствуемся при выборе, какое крупное животное нам забить, отправив его мясо на консервный завод: дикое или домашнее, — и критерий этот — биологическое многообразие и вариативность, которые столь важно сохранить и поддерживать в природе. В целом домашние животные сохранили очень немногое из унаследованного ими от предков (например, от зубров) разнообразия, которое было заложено в банке генных данных, а то немногое, что им удалось все-таки сохранить, существует теперь в миллионах копий. Одомашнивание животных приводит к утрате исходного многообразия форм, хотя и в подобных условиях может произойти мутация, в результате которой появится новая разновидность животных, наделенных каким-нибудь отличительным качеством — например, желательным для этого вида размером. Наоборот, генный набор каждого тигра — уникальная сокровищница, в которой хранятся генные богатства определенного вида. Поэтому один тигр представляет большую ценность, чем одна корова. На сегодняшней день в дикой природе сохранилось несколько тысяч тигров, и генный код каждого из них в значительной степени неповторим. Разнообразие типов («рас») тигров представлено в дикой природе разнообразием особей. Итак, даже если мы оставим на время в стороне тот факт, что над тиграми нависает непосредственная угроза исчезновения — чего не скажешь о коровах, — то и тогда убийство одного тигра трагичней убийства одной коровы, ибо наносит более серьезный ущерб видовому разнообразию жизни. К семейству кошачьих относится еще один вид крупных кошек — гепарды, угроза исчезновения подступила к ним еще ближе, чем к тиграм. И это определяется не только малочисленностью популяции гепардов, но и тем, что в дикой природе не сохранилось естественного разнообразия видов и форм — все гепарды — близкие родственники, произошедшие, вероятно, от единственной пары особей, которой удалось выжить во время последнего Ледникового периода. Удивительно, но факт: если не считать Африки, все животные Земли по своему генетическому богатству и разнообразию ближе к гепардам и одомашненному крупному рогатому скоту, нежели к тиграм. Можно проследить родоначальные линии всех этих животных и выяснить, что все они произошли от нескольких мужских и женских особей, которые покинули Африку только 80 тысяч лет тому назад. Итак, многообразие... Оставим в стороне эстетический аспект этого явления и подумаем, что заставляет нас придавать ему такую важность. Ответ на волнующий нас вопрос таков — выживание. Своеобразным источником энергии для эволюции и самого существования природы является хаотическое многообразие. Без основанного на случайности генного многообразия виды утрачивают необходимую для выживания гибкость и способность адаптироваться к жизни в условиях стресса. Прежде чем потомство единственной пары особей достигнет хаотического многообразия, сменится множество поколений. Итак, видам животных, которые прошли сквозь ледниковый перешеек, не удалось сохранить свое генетическое многообразие, и теперь настал срок собирать камни и восполнять утраты.
Смертоносные болезни
Возможно, вы будете удивлены, узнав, что живете в условиях постоянного эволюционного стресса. Самым ярким примером эволюционного стресса являются вездесущие и изменчивые инфекции. Бактерии и вирусы эволюционируют значительно быстрее нас. В организм каждого из нас встроена иммунная система, позволяющая распознавать множество новых вариантов вирусов, которыми кишит наша плоть, бороться с ними и воздвигать на их пути специфическую защиту. Способность организма распознавать многообразных возбудителей различных инфекционных заболеваний и бороться с ними определяется генетическим кодом человека. Реакции иммунной системы весьма разнообразны, но ее возможности ограничены и отчасти определяются опытом борьбы с теми возбудителями, которые в прошлом были распространены в обществе. Генном человека определяет многообразные механизмы сопротивления заболеваниям, которые приводятся в действие при помощи адаптивной иммунной системы. Некоторые народы обладают стойким иммунитетом к заболеваниям, которые перенесли их предки. В качестве примера приведу тот факт, с которым я столкнулся, когда работал в Гонконге, где дети этнических китайцев почти никогда не страдают менингококковыми заболеваниями (менингитом и/или септицемией). У них выявлена хорошо развитая уникальная особенность иммунной системы — способность сопротивляться менингококкам в крови, но в отличие от европейцев и других этнических групп китайцам удается избегать этих заболеваний и даже не становиться носителями их возбудителей. Наоборот, наиболее распространенным возбудителем менингита у китайцев, живущих в Гонконге, являются бациллы туберкулеза, которые крайне редко вызывают менингит у других развитых народов. Этот пример свидетельствует, что между разными народами существуют серьезные различия в эффективности приобретенного иммунитета к специфическим заболеваниям. Что касается других видов инфекционных заболеваний, таких, как малярия, то наша врожденная сопротивляемость им носит не только иммунологический характер. Так, некоторые генетические нарушения, распространенные в тропиках и являющиеся результатом эволюционной селекции, прямо препятствуют успешному размножению малярийных паразитов. Этот механизм генетической защиты против болезни, в конкретном случае малярии — одного из самых грозных убийц человека, — обусловлен наследственными расстройствами в структуре красных кровяных телец, к которым паразиты проявляют особый интерес. Такие генетические нарушения, большая часть которых определяется общим названием «талассемия»[463], являются общераспространенными в тех районах, в которых малярия широко распространена сейчас или была распространена в прошлом. Название этой болезни — талассемия — происходит от греческого слова talassa, что означает «море», поскольку на таких островах Средиземного моря, как Кипр, отмечается высокий уровень заболеваемости этими недугами. Кроме того, эти болезни встречаются и в тропических и субтропических регионах. Так, у носителей развивается защита от паразитов, однако паразиты обладают способностью приспосабливаться к ней и преодолевать новые средства защиты. Главная беда здесь заключается в том, что их эволюция протекает куда более быстрыми темпами, чем наша. Лучшие шансы выжить имеют те паразиты, которые не убивают своих хозяев. К сожалению, далеко не все насекомые знают это. Эволюционная гонка вооружений между инфекционными заболеваниями и животными— носителями их часто принимает для носителя дурной оборот, когда паразиты перебираются с одного вида на другой. Некоторые из наиболее острых вирусных и бактериальных болезней человека, включая бубонную чуму, мы получаем от животных, живущих в девственных районах, куда эпизодически вторгается человек. Другой грозный хакер (взломщик) нашего иммунитета — знаменитый вирус иммуннодефицита человека (ВИЧ), более известный как СПИД, который в наши дни обогнал по уровню смертности такие традиционные бичи человека, как туберкулез и малярия. Сегодня можно слышать утешительный миф о том, будто СПИД, эта чума и роковая кара, является суперсмертоносным, ибо поражает иммунную систему, и когда ее носители вымрут, на Земле более не возникнет аналогичных болезней. Это не так: СПИД — грозное предупреждение о комплексной приспосабливаемости инфекционной болезни, которую мы встречаем во все более широких масштабах по мере проникновения в самые отдаленные уголки нашей планеты, которая становится все более и более тесной. В любом случае два варианта этого вируса, ВИЧ1 и ВИЧ2, могли развиться независимо друг от друга и произойти от двух различных видов африканских приматов. Наша усиливающаяся тяга к межконтинентальным путешествиям и поездкам в дальние края способствует переносу и распространению болезней, которые в прошлом носили исключительно локальный характер. Пока я писал эту книгу, появились сообщения о возникновении в Южном Китае новой неизвестной болезни, обозначаемой аббревиатурой SARS (атипичная пневмония). Она просто пополнила обширный список экзотических и очень серьезных инфекций, которые могут возникать у животных. Впоследствии эта болезнь благодаря авиаперелетам была занесена во многие другие страны.
Генетические вмешательства
Одним из ответов на увеличивающуюся угрозу возникновения пандемических заболеваний можно считать генетические вмешательства. Пресса и прочие средства массовой информации чуть ли не каждую неделю сообщают о новых успехах в области генетического прогнозирования и вмешательства, которые способны коренным образом изменить и нашу жизнь, и жизнь последующих поколений. Нас уверяют, что с генетическими заболеваниями будет покончено, а самые «продвинутые» представители нашего общества смогут заказывать детей с заданными свойствами или собственные клоны. Пренатальная диагностика и службы генетических консультаций, позволяющие выявить носителей неизлечимых недугов и серьезных генетических нарушений, появившиеся не так давно, уже внесли громадные перемены в жизнь многих людей. Так, эти службы сыграли огромную позитивную роль в таких странах, как Кипр, где отмечается очень высокий уровень заболеваемости мутационной бета-талассемией, обусловленный малярией, свирепствовавшей здесь в прошлом. Подобные вмешательства, мотивированные этическими соображениями, и в будущем будут продолжать бороться с ростом болезней и нищеты. Однако подобным генетическим вмешательствам не по силам расширить диапазон нашего генного многообразия. Не касаясь проблем этического и технического характера, надо признать, что генетическое вмешательство способно лишь сузить генное многообразие. Это в равной степени относится и к бесчеловечной концепции отбраковывания субъективно оцениваемых «нежелательных» генетических элементов, практиковавшейся нацистами в отношении многих пациентов, страдавших умственными и прочими недугами, а также в отношении евреев и цыган, и к концепции «здорового нового мира», пропагандируемой иными специалистами по генной инженерии. Даже если генетикам нового поколения удастся сконструировать особенно «удачную модель», которая займет видное место на рынке среди потенциальных родителей, подобный опыт может оказаться пагубным и самоубийственным. Бесконечное воспроизведение клонов такой «супермодели» приведет к резкому сокращению генного многообразия нашего вида и тем самым повысит подверженность новым инфекционным болезням. И самые новейшие технологии не смогут найти спасительную брешь в стене той тюрьмы нищеты, которой станет наша планета для подавляющего большинства своих жителей. Любопытно, что две основные особенности нашего вида, интересующие нас более всего, а именно объем мозга и продолжительность жизни, потенциально допускают возможность генетического вмешательства. Причем к первой из них это относится в большей мере, чем ко второй. Представляется вполне возможным, что, если ученым будет позволено, они, опираясь на имеющиеся знания, через несколько десятилетий смогут «создать» человека с еще более крупным объемом коры головного мозга. Это может быть достигнуто либо путем манипуляций с единичными генами (генами, которые контролируют формирование эмбриона и развитие органов тела), либо — более грубо — посредством впрыскивания «продуктов» такого гена в эмбрион на соответствующем этапе его развития. Не берусь судить, станет ли такой «увеличенный мозг», созданный посредством генной инженерии, более мощным и интеллектуально развитым. Мне лично не хотелось бы дожить до времени, когда это станет реальностью. Что же касается долголетия для богатых, то в этой сфере существует много чисто коммерческих возможностей... и одновременно опасностей. У Джорджа Гершвина есть одно изящное, хотя политически и не вполне корректное высказывание о пресловутом долголетии Мафусаила, запечатленное в песенке «Это не обязательно так и было» из оперы «Порги и Бесс»: «Это жизнью не назовется, // Ведь девчонка не отдается // Старику в девятьсот лет?» Разумеется, здесь возникает угроза перенаселенности. Согласно одному из вьетнамских преданий, люди первоначально достигали бессмертия, сжигая своих усопших под кроной древа жизни. Но однажды ящерица, которая хотела накормить толпу голодных, отделив от тела собственный хвост, посоветовала не предавать усопших огню, а хоронить их под тем же древом жизни. В мир пришла смерть, и жить на свете стало легче.
Эволюция остановилась?
Некоторые генетики утверждают, что в наше время естественная эволюция человека уже остановилась, и теперь благодаря невиданному прогрессу в области борьбы с болезнями и генетическим вмешательствам, таким, как консультации и пренатальная диагностика, роль гаранта существования человека взяла на себя медицина. Это представляется абсурдным. Для подавляющего большинства жителей планеты подобная роскошь абсолютно недоступна, а ее влияние на многообразие типов рода человеческого в количественном отношении в целом ничтожно. В частности, пренатальная диагностика в любом случае направлена на коррекцию «сбоев» единичных генов, вызывающих серьезные заболевания, наследуемые от обоих родителей. Люди, страдающие подобными недугами, даже если им удастся выжить, обычно имеют серьезные проблемы с репродуктивной передачей этих болезней. Подобная практика отнюдь не является евгеникой. До тех пор, пока мы будем умирать от болезней, так или иначе связанных с нашими генами, эволюционный отбор будет продолжать работать. Помимо инфекционных болезней не следует сбрасывать со счета и другие смертельные недуги, обусловленные генетической предрасположенностью, такие, например, как рак. Постоянно снижается количество сперматозоидов в мужской сперме. И хотя виновниками этого в известной мере является загрязнение нашей среды обитания и пищи всевозможными химикатами, наша подверженность им колеблется и имеет генетическую обусловленность. Что же касается биологической эволюции, то она не прекратилась и не остановилась, а лишь немного замедлилась. В конце концов, люди — создание тех же самых эволюционных сил, что и другие животные, и они будут оставаться таковыми и дальше. Хотелось бы надеяться, что мы сумеем осознать это, пока не поздно. Быть может, мы наконец избавимся от заносчивости, присущей нашему виду, и признаем, что являемся крошечным пятнышком на поверхности нашей планеты и зависим от своих соседей — животных и растений. Только тогда мы сможем помочь окружающему миру восстановиться после страшных ран, нанесенных ему нашим прогрессом. Как-то раз мой сын спросил меня, могут ли в будущем появиться — или быть искусственно созданными — новые виды человека. Что ж, мне не оставалось ничего другого, как дать стандартный родительский ответ — «это зависит от многого». Я верю, что в первую очередь это будет зависеть от того, как многообразные культуры человечества повлияют на нас самих и окружающую среду. Наши безрассудные и агрессивные действия, пагубно влияющие на окружающую среду и продиктованные быстрым ростом численности населения, могут возыметь нежелательные последствия и привести к катастрофе или даже исчезновению человека как вида. То, как мы обустроим наше родное гнездо — нашу планету, и сумеем ли избежать ее дальнейшего разрушения, опять-таки зависит от направления развития человеческой культуры. Если мы сумеем выжить, оказавшись на грани вымирания, самоистребления и т.д., наши потомки могут стать несколько иными в биологическом отношении, но нет никакого сомнения, что они будут иными в отношении культуры.
ПРИЛОЖЕНИЕ 1 ИСТИННЫЕ ДОЧЕРИ ЕВЫ


Последние комментарии
5 часов 55 минут назад
5 часов 58 минут назад
2 дней 12 часов назад
2 дней 16 часов назад
2 дней 18 часов назад
2 дней 19 часов назад