Расовые различия в интеллекте [Ричард Линн] (fb2) читать онлайн
Книга 241226 устарела и заменена на исправленную
[Настройки текста] [Cбросить фильтры]
Энцефалометр Д. Н. Зернова. Производство фирмы Ф. Б. Шваве, Поставщика Двора бго Императорского Величества, Россия, 1909 г.
Сканировал и создал книгу - vmakhankov

Race Differences in Intelligence
An Evolutionary Analysis
Richard Lynn
Washington Summit Publishers Augusta, GA
A National Policy Institute Book 2006

Ричард Линн
РАСОВЫЕ РАЗЛИЧИЯ В ИНТЕЛЛЕКТЕ
ЭВОЛЮЦИОННЫЙ АНАЛИЗ

Москва
2010
ББК 87.3 Л 59
Идея проекта: В. Б. Авдеев
Перевод с английского: Д. О. Румянцев
Научный консультант: доктор филологических наук А, А. Григорьев Ричард Линн
Л 59 Расовые различия в интеллекте. Эволюционный анализ. /Пер. с англ. Румянцев Д. О. — М.: Профит Стайл, 2010. — 304 с.
ISBN 5-98857-157-3
ISBN 1-593680-21-Х (Washington Summit Publishers, 2006)
Фактор интеллекта в жизни тех или иных народов и рас многократно освещался в преданиях и пословицах на протяжении всей истории. Примеров тому несметное множество. И речь вовсе не обязательно идет о том, что одни «лучше» или «хуже», чем другие. Сама специфика умственной деятельности с точки зрения стиля, а также динамики различных психологических актов отмечена во множестве письменных памятников с древнейших времен. В священных текстах мировых религий мы без труда обнаруживаем ту же взаимосвязь, а эпоха «великих географических открытий» стараниями первооткрывателей и миссионеров лишь подтвердила правило гласящее, что во всех частях света отношение к базовым этическим ценностям глубоко различно. Желания и интересы людей всюду кардинально отличаются. Одни народы и расы с поразительным упорством на протяжении веков являют нам чудеса практической смекалки и выживаемости, считая это высшим благом, другие, напротив, с нескрываемым презрением относятся к такой стратегии поведения, предпочитая создавать гигантские идеалистические концепции. Именно это многообразие проявлений высшей нервной деятельности и дает нам великолепную палитру величия рода человеческого.
Richard Lynn 4, Longwood House, Failand,
Bristol BS8 3TL, UK Tel.: 01275 392 092
E-mail: Lynnr540@aol.com
Охраняется Законом РФ об авторском праве. Воспроизведение всей книги или любой ее части воспрещается без письменного разрешения издателя. Любые попытки нарушения закона будут преследоваться в судебном порядке.
No part of this publication may be reproduced, distributed, or transmitted in any form or by any means, including photocopying, recording, or other electronic or mechanical methods, or by any information storage and retrieval system, without prior written permission from the publisher, except for brief quotations embodied in critical reviews and certain other noncommercial uses permitted by copyright law. For permission requests, write to the publisher at the address below.
EAN 978-5-98857-157-5
© Ричард Линн, 2010
© Профит Стайл, 2010
СОДЕРЖАНИЕ
Обращение Ричарда Линна к русским читателям 9
Эволюция, раса и интеллект — Предисловие к русскому изданию 10
Глава 1. Значение и измерение интеллекта 17
Определение интеллекта 19
Иерархическая модель интеллекта 20
Коэффициент интеллекта (IQ) 21
Эффект Флинна 22
Глава 2. Значение и формирование рас 24
Формирование рас, разновидностей и пород 24
Разновидности в животном мире 25
Таксономии рас 27
Расовые различия в заболеваемости 29
Существуют ли расы? 30
Глава 3. Европейцы 35
Интеллект автохтонных европейцев 36
Европейцы вне Европы 41
Европейцы — студенты университетов 43
Размер мозга 44
Наследуемость интеллекта у европейцев 45
Глава 4. Африканцы 47
Интеллект африканцев в Африке южнее Сахары 48
Студенты университетов в Африке 54
Африканцы островов Карибского бассейна и Латинской Америки 56
Афроамериканцы в Соединенных Штатах Америки 57
Африканцы в Великобритании 62
Африканцы в Нидерландах 66
Африканцы в Израиле 67
Кратковременная память и скорость восприятия у африканцев 68
Музыкальные способности 70
Время реакции 73
Размер мозга африканцев и европейцев 76
Показатели интеллекта африканско-европейских гибридов 79
Наследуемость интеллекта у афроамериканцев 81
Генетические и средовые объяснения низкого интеллекта африканцев 82
Оценка генотипного IQ африканцев 86
Глава 5. Бушмены и пигмеи 88
Интеллект бушменов 89
Размер мозга 92
Пигмеи 92
Глава 6. Аборигены Южной Азии и Северной Африки 94
Интеллект аборигенов Южной Азии и Северной Африки 95
Выходцы из Южной Азии и Северной Африки в Великобритании и Австралии 97
Выходцы из Южной Азии и Северной Африки в континентальной Европе 99
Индусы в Африке, на Фиджи, в Малайзии и на Маврикии 101
Учащиеся полной средней школы и студенты 103
Размер мозга аборигенов Южной Азии 105
6
Ричард Линн
Наследуемость интеллекта у аборигенов Южной Азии и Северной Африки 106
Генетические и средовые детерминанты интеллекта аборигенов Южной Азии и
Северной Африки 107
Интеллект в Израиле 108
Глава 7. Аборигены Юго-Восточной Азии 111
Интеллект аборигенов Юго-Восточной Азии 111
Аборигены Юго-Восточной Азии в Соединенных Штатах и Нидерландах 112
Размер мозга аборигенов Юго-Восточной Азии 113
Генетические и средовые детерминанты IQ аборигенов Юго-Восточной Азии 114
Глава 8. Австралийские аборигены 115
Интеллект австралийских аборигенов 116
Гибриды австралийских аборигенов с европейцами 118
Интеллект по Пиаже 119
Пространственная память 121
Размер мозга 123
Генотипный интеллект австралийских аборигенов 124
Интеллект новогвинейцев 126
Заключение 128
Глава 9. Аборигены островов Тихого океана 130
Интеллект новозеландских маори 131
Другие аборигены островов Тихого океана 132
Гибридное население Гавайских островов 133
Размер мозга аборигенов островов Тихого океана 134
Средовые и генетические детерминанты интеллекта аборигенов островов
Тихого океана 134
Глава 10. Аборигены Восточной Азии 136
Интеллект аборигенов Восточной Азии 136
Аборигены Восточной Азии в Соединенных Штатах 144
Другие исследования аборигенов Восточной Азии вне Северо-восточной Азии 150
Аборигены Восточной Азии, усыновлённые европейцами 152
Гибриды аборигенов Восточной Азии с европейцами 154
Время реакции 155
Зрительная память 156
Размер мозга 157
Наследуемость интеллекта у аборигенов Восточной Азии 159
Средовые и генетические объяснения IQ аборигенов Восточной Азии 159
Глава 11. Арктические народы 163
Интеллект арктических народов 163
Зрительная память 166
Размер мозга 167
Генотипный интеллект 167
Глава 12. Американские индейцы 169
Интеллект американских индейцев в Северной Америке 170
Коэффициент интеллекта по тесту «Нарисуй человека» 171
Латинская Америка 172
Зрительная память 173
Гибриды американских индейцев с европейцами 174
Музыкальные способности 174
Размер мозга 175
Испаноговорящие в Соединенных Штатах 176
Генотипный интеллект американских индейцев 178
Расовые различия в интеллекте
7
Глава 13. Надёжность и валидность расовых различий в интеллекте 180
Итоговый обзор расовых различий в интеллекте 180
Надёжность оценок IQ рас 183
Валидность оценок IQ рас: Понятия числа 184
Валидность оценок IQ рас: Успехи в образовании 185
Валидность оценок IQ рас: Доход на душу населения и экономический рост 190
Глава 14. Средовые и генетические детерминанты расовых различий
в интеллекте 193
Питание 194
Голландское исследование голодания в период Второй Мировой войны 198
Нейрофизиологический эффект недоедания 199
Образование 200
Генетические детерминанты расовых различий в интеллекте 202
Генотипно-средовая ковариация 206
Глава 15. Эволюция интеллекта 208
Общие принципы эволюции интеллекта 208
Млекопитающие 210
Птицы 211
Приматы 212
Гоминиды 1 215
Величины IQ обезьян, человекообразных обезьян и гоминид до Homo sapiens 217
Глава 16. Климат, раса, размер мозга и интеллект 219
Эволюция рас 219
Требования к когнитивной сфере в северных широтах 221
Расовые различия в размере мозга 223
Расовые различия: зимние температуры, размер мозга и IQ 225
Размер мозга и интеллект человека 226
Вклад расовых различий размера мозга в различия в интеллекте 228
Половые различия в интеллекте и размере мозга 230
Генетические процессы в эволюции расовых различий в IQ 234
Глава 17. Эволюция расовых различий в интеллекте 237
Африканцы 238
Бушмены 239
Аборигены Южной Азии и Северной Африки 240
Аборигены Юго-Восточной Азии 246
Аборигены островов Тихого океана 247
Австралийские аборигены 247
Европейцы 251
Аборигены Восточной Азии 253
Арктические народы 254
Американские индейцы 256
Заключение 259
Приложение. Тесты интеллекта 260
Список литературы 263
Экспертное заключение 297
Посвящается Джойс Sei il mio amor у tutta la mia vita (Ты моя любовь и вся моя жизнь) Мими — Дж. Пуччини, «Богема», Акт V
ОШЩЖ РИЧАРДА тм Л РУ((41ИИ ЧКТЛТШН
Дорогой русский читатель! Я рад, что моя книга издана в России и родственные нам русские люди смогут познакомиться с изложенными в ней взглядами на расовые различия в интеллекте. Я обобщил результаты нескольких сотен исследований, показывающих, что мы, европейцы, наряду с народами Восточной Азии (китайцами, японцами и корейцами), обладаем самым высоким интеллектом. Это можно считать очевидным. Почти все открытия в науке и технике сделали за последние две с половиной тысячи лет наши народы, народы Европы и Восточной Азии, они же создали почти все великие произведения изобразительного искусства, музыки и литературы. Во второй части книги я предлагаю этому объяснение. Я объясняю это тем, что нашим предкам пришлось жить в суровых условиях во время последнего ледникового периода, когда лишь обладавшие высоким интеллектом могли выжить. Теперь, однако, будущее наших народов в опасности. Количество рождающихся у нас детей недостаточно, чтобы сохранялась численность наших популяций. Как мы решим эту проблему? Сможем мы выжить? Или же наше место займут народы с более низким интеллектом? Предоставляю читателю судить об этих важных вопросах.
(Пер. с английского — А. А. Григорьев)
Ричард Линн Университет Ольстера
ШМЩМ. РАСА И ИНТШШ - ПР-ЕДИ((10^И^ К РУССКОМУ ИЭД1ШН40
Простота хуже воровства (русская народная пословица)
Фактор интеллекта в жизни тех или иных народов и рас многократно освещался в преданиях и пословицах на протяжении всей истории. Примеров тому несметное множество. И речь вовсе не обязательно идет о том, что одни «лучше» или «хуже», чем другие. Сама специфика умственной деятельности с точки зрения стиля, а также динамики различных психологических актов отмечена во множестве письменных памятников с древнейших времен. В священных текстах мировых религий мы без труда обнаруживаем ту же взаимосвязь, а эпоха «великих географических открытий» стараниями первооткрывателей и миссионеров лишь подтвердила правило гласящее, что во всех частях света отношение к базовым этическим ценностям глубоко различно. Желания и интересы людей всюду кардинально отличаются. Одни народы и расы с поразительным упорством на протяжении веков являют нам чудеса практической смекалки и выживаемости, считая это высшим благом, другие, напротив, с нескрываемым презрением относятся к такой стратегии поведения, предпочитая создавать гигантские идеалистические концепции. Именно это многообразие проявлений высшей нервной деятельности и дает нам великолепную палитру величия рода человеческого.
Неудивительно поэтому, что мудрецы всегда пытались соизмерить и объяснить умственные способности, как отдельных индивидуумов, так и целых племен. Уже в первых летописных источниках мы можем обнаружить попытки приведения в систему групповых физических и психических признаков отличий людей. Управление первыми крупными империями и поликультурными образованиями потребовало от администраторов перехода с образного мышления поэтов к сухой государственной статистике, так как каждый владыка желал знать, чего он вправе ожидать от нового неведомого народа, волею Богов попавшего под длань его управления. Дошедшие до нас законодательные акты Древнего Египта и Вавилона не дают нам никаких возможностей ошибиться в понимании этой естественноисторической закономерности. На заре развития человечества поначалу использовались различные косвенные показатели для определения умственных, а равно и моральных предпочтений народа: от количества выставляемых воинов, детей в семьях, величины городов и справед
Расовые различия в интеллекте
И
ливости законов, до запасов зерна и тяготения к тем или иным ремеслам. По мере развития цивилизации система критериев оценки усложнялась, но незыблемой оставалась одна и та же потребность, а именно: помимо эмоциональных качественных характеристик определить некие абсолютные параметры умственных способностей. Древнейшая как мир система экзаменования молодых людей при приеме на работу или в учебные заведения служила именно этим целям. Воинские звания, награды, научные степени и общественные привилегии также во все времена предназначались для активизации в гражданах витальных способностей, в том числе и интеллекта.
Таким образом, в связи с универсализацией бытия и принципов организации общежития в Новое время возникла потребность в унификации критериев оценки умственных способностей различных ветвей человеческого рода. Глобальные контакты автоматически потребовали от каждой расы понимания своего места в общем мироустройстве, и измерение интеллекта явилось закономерным шагом на пути к общей цивилизационной гармонии.
Выдающийся классик естествознания Карл Линней (1707-1778) основал первую научную расовую классификацию как раз на триединстве проявлений физических, психических и моральных качеств, чем и подготовил основы возникновения классической эволюционной теории. Ибо совершенно очевидно, что в процессе поступательного развития претерпевает изменения не только физическая оболочка человека, но также её ментальное и нравственное наполнение, которое вырабатывается как совокупность приспособительных реакций. Но абстрактный человек—это фикция, одно из величайших гуманитарных заблуждений Нового времени. Каждый человек от природы является носителем наследственных расовых признаков, которые неотъемлемо проявляются в каждом виде его деятельности, в том числе и интеллектуальной. Расовые признаки — это печать, которая легко узнаваема на каждом историческом деянии.
В этом плане книга британского профессора Ричарда Линна, впервые предлагаемая русскому читателю с любезного разрешения автора, является своего рода венцом магистральной линии классической эволюционной теории, в которой английская школа традиционно задавала тон.
Сегодня, когда как в средствах массовой информации, так и в научных кругах стало своего рода признаком хорошего тона критиковать Ч. Дарвина и вскрывать якобы несостоятельность его теории. Нужно четко ответить ангажированным критикам, что он нигде и никогда не писал, будто человек в прямом смысле этого слова произошел от обезьяны. Кроме того, в своей автобиографии мэтр многократно подчеркивал, что не сказал ничего нового, а лишь систематизировал многочисленные концепции естествознания, существовавшие до него.
12
Ричард Линн
Полное название книги современного автора «Расовые различия в интеллекте. Эволюционный анализ» четко показывает, в каком контексте мы должны воспринимать результаты его многолетнего труда.
Ричард Линн — профессор психологии Ольстерского университета в Дублине. Он является одним из старейших (родился в 1930 г.) и авторитетнейших представителей классической Британской школы психологии. Большая часть его работ посвящена изучению интеллекта. Среди основных открытий, сделанных Р. Линном, следует отметить установление факта, что коэффициент интеллекта (IQ) коренных жителей Восточной Азии в среднем на 5 баллов выше, чем у европейцев вне зависимости от места проживания испытуемых. Автором также впервые было показано, первоначально на японской популяции, что во второй половине XX столетия наблюдалось неуклонное повышение результатов оценок IQ посредством психологических тестов. Впоследствии была показана универсальность этого явления, получившего название «эффект Флинна» по фамилии исследователя, углублённо занимавшегося изучением данного феномена. Причиной считается улучшение условий жизни в экономически развитых странах в XX веке. Ричардом Линном впервые было показано (в 1994 г.), что интеллект взрослых мужчин в среднем на 4-5 баллов выше, чем у женщин, что он объясняет относительно меньшим размером мозга у женщин. Этот факт первоначально вызвал весьма бурную реакцию в научных и общественных кругах.
Среди крупных работ Р. Линна необходимо назвать монографии «Дис- геника» («Dysgenics»; Westport, СТ: Praeger, 1996) и «Евгеника» («Eugenics»; Westport, СТ: Praeger, 2001), в которых он приводит данные о наметившемся ухудшении физического и психического здоровья и интеллектуальных способностей современной человеческой популяции и оценивает возможные методы борьбы с этой угрозой. В книге «IQ и благосостояние наций» (в соавторстве с финским учёным Тату Ванханеном (Tatu Vanhanen) — «IQ and the Wealth of Nations»;Westport, CT: Praeger, 2002) P. Линн рассматривает проблему зависимости различий в показателях благосостояния и экономического роста наций от показателей интеллекта. По результатам анализа данных для 185 современных народов автор аргументировано показывает вклад коэффициента интеллекта каждой нации в показатели её экономического благосостояния.
С 1991 г. основным направлением работ Р. Линна является сравнительное изучение интеллекта народов разных рас. Настоящая книга «Расовые различия в интеллекте. Эволюционный анализ», изданная в 2006 г. («Race Differences in Intelligence: An Evolutionary Analysis»; Augusta, Georgia: Washington Summit Books) является результатом примерно 25летнего труда автора. Предисловие к книге написал замечательный психолог британской школы, работающий в Канаде, Джон Филипп Раштон,
Расовые различия в интеллекте
13
считающийся крупнейшим специалистом в области изучения расовых особенностей психологии. Он отмечает, что Р. Линн в своих исследованиях вступил на «минное поле», так как и первоначально, и сейчас сторонники гипертрофированной «политкорректности» не соглашаются принять результаты его исследований. Что неудивительно, так как, по словам другого выдающегося британского мыслителя Томаса Гоббса, «если бы геометрические аксиомы задевали интересы людей, их бы опровергали». Имя Р. Линна смело можно было бы поставить под словами, сказанными другим выдающимся учёным современности, нобелевским лауреатом, расшифровавшим структуру ДНК, Джеймсом Уотсоном: «Честность полезна этому миру, она заставляет мир работать эффективнее».
На материале собственных исследований и анализа более 500 работ других авторов, выполненных с начала вхождения психологических тестов в арсенал психологии до настоящего времени, Р. Линн демонстрирует особенности интеллекта представителей всех человеческих рас (более сотни наций и народностей) и оценивает вклад генетических и средовых факторов в эти различия. Автором предложена модель эволюции наблюдаемых расовых различий интеллекта. Несмотря на то, что труд Р. Линна представляет на сегодняшний день наиболее полный свод данных по проблеме, книга весьма компактна. Автор излагает свои мысли достаточно доступным языком, для их понимания в большинстве случаев не требуется углублённого предварительного изучения психологии или эволюционной биологии.
Его работа — это самый исчерпывающий и последний по времени (книга опубликована в 2006 г.) анализ данных исследований расовых особенностей интеллекта. Показатели интеллекта сопоставляются с другими психологическими и биометрическими характеристиками, предлагается ряд объяснений расовых различий интеллекта, включая влияние генетических факторов и среды обитания. Автор предлагает собственную теорию эволюции интеллекта человека.
В России книги по расовым различиям в интеллекте не публиковались, если не считать отдельных труднодоступных журнальных публикаций 20-х гг. XX столетия.
В исследованиях, анализирующихся Р. Линном, для измерения интеллекта используются разнообразные тесты IQ. Некоторые из этих тестов широко применяются в настоящее время в нашей стране для решения практических задач (например, прогрессивные матрицы Равена), в то время как другие являются совершенно неизвестными. Знакомство с книгой Р. Линна будет способствовать расширению инструментария отечественных психологов. Кроме того, практика работы с тестами IQ в нашей стране на сегодняшний день далека от совершенства. После длительного перерыва, вызванного разгромом тестологии в 1930-е гг., культура этой практики
14
Ричард Линн
была утеряна. В настоящее время используется огромное количество, мягко выражаясь, некачественных, психодиагностических тестов и психодиагностика не применяется для решения масштабных задач, например в сфере образования.
Чтобы разложить всё на свои места и облагоразумить активизировавшихся критиков эволюционизма, полагаем необходимым пролить некоторый свет на историю вопроса.
Именно англичане были теми первопроходцами, кто в эпоху «великих географических открытий» впервые сумел сделать сугубо прагматические выводы и привести в систему свод знаний об «экзотических» странах. Основанное в 1660 году в Англии Королевское общество служило целям пропаганды науки и её практического применения с учётом расовых различий представителей коренного населения новых континентов.
Перечислим имена гениев науки в данной области.
Джон Бульвер (John Bulwer; 1606-1656) заложил основы расологии, создав системное описание биотипов людей с их характерными физическими и психическими признаками. Томас Виллис (Thomas Willis; 1621-1675) первым описал анатомию и физиологию нервной системы человека и животных. Джон Рей (John Ray; 1627-1705) сформулировал принципы научной классификации рас. Эдвард Тайсон (Edward Tyson; 1650-1708) дал подробное сравнительное описание морфологического строения орангутанга, негра и европейца во всех деталях скелета, мышечных тканей и кожных покровов. Джон Хантер (John Hunter; 1728-1793) стоял у основ создания краниологии и, вместе с тем, развил философское понимание расовой проблемы. Чарльз Уайт (Charles White; 1728-1813) сформулировал принципы расовой хирургии. Сэр Уильям Лауренс (Sir William Lawrence; 1783— 1867) заложил базу сравнительной анатомии и медицинской этики в её современном понимании. С именем такого ученого, как Джеймс Коулз Причард (James Cowles Prichard; 1786-1848) связано становление новой самостоятельной науки — этнологии.
Джеймс Хант (James Hunt; 1833-1869) основал первое английское Антропологическое общество, а Роберт Нокс (Robert Knox; 1791-1862) создал учение о «трансцендентальной анатомии», согласно которому все живые существа структурированы сходным образом в силу универсального единства биологических функций. Именно эта концепция заменила христианскую идею замысла Божия и заложила анатомическую основу эволюционной теории.
Развивая идеи Чарльза Дарвина, Томас Генри Гекели (Thomas Henry Huxley; 1825-1895) придал современный вид классификации рас, а Альфред Рассел Уоллес (Alfred Russel Walles; 1823-1913) сформулировал глобальные принципы биогеографии. Наконец Джон Биддоу (John Beddoe; 1826-1911) привнёс в антропологию статистические методы и окончательно
Расовые различия в интеллекте
15
превратил её, таким образом, в точную науку. Эдвард Бёрнетт Тейлор (Edward Burnett Taylor; 1832-1917) в свою очередь обосновал доктрину о «шкале цивилизации» и предложил с её помощью измерять различные культуры. Сэр Фрэнсис Гальтон (Sir Francis Galton; 1822-1911) обессмертил своё имя созданием евгеники — науки об улучшении человеческого рода. Альфред Корт Хэддон (Alfred Cort Haddon; 1855-1940) сделал принципы английской физической антропологии универсальными, ибо именно в это время по всему земному шару начинается создание единой унифицированной методики расовых измерений. Карл Пирсон (Karl Pearson; 1857-1936) создал биометрику и заложил основы современной статистики, благодаря чему все отрасли естествознания, в том числе и гигантский корпус наук о человеке, получили мощный математический аппарат для обработки результатов исследований. Сэр Артур Кейт (Sir Arthur Keith; 1866-1955) создал теорию о наследственном гормональном происхождении расовых различий и, следовательно, о происхождении современных рас от различных первопредков в различных географических зонах и в разное время. Сэр Графтон Эллиот Смит (Sir Grafton Elliot Smith; 1871— 1937) на основе новейших методов изучал неврологические и психофизиологические особенности рас в контексте развития их культур. Уильям Хэле Риверс Риверс (William Halse Rivers Rivers; 1864—1922) создал шкалы цветовой и болевой чувствительности для представителей различных рас, чем окончательно подвёл биологическую базу под причины разнообразия культур. Джон Рэндал Бейкер (John Randal Baker; 1900-1984) оформил масштабную универсальную концепцию о биологических критериях оценки самостоятельности культур.
Этот перечень великих английских учёных-естествоиспытателей обозначает лишь некоторые вершины того гигантского мировоззренческого айсберга, в основе которого лежит глубинное архетипическое представление о том, что всё в этом мире обладает специфическими различиями, которые нужно уметь измерять. Таким образом, главная фундаментальная книга профессора Ричарда Линна на сегодняшний день венчает многовековую славную традицию развития английского естествознания в данной области, в чем и состоит ее непреходящая ценность.
Знание представляемого в книге материала необходимо, на наш взгляд, самым широким слоям читающей аудитории в России: от психологов, социологов, философов до политических деятелей, сотрудников спецслужб и правоохранительных органов. Полезна эта информация также для криминологов, врачей и педагогов. В условиях многонациональной и многорасовой России именно объективный анализ интеллекта граждан поможет гасить в зародыше любые конфликты и максимально рационально использовать генофонд страны в целях её процветания. Будучи по наивности опьянены иллюзиями равенства, мы уже потеряли нашу великую страну и потому не
16
Ричард Линн
имеем больше исторического права на повторение этой катастрофы, которая, как мы помним, сопровождалась войнами почти по всему периметру бывшего СССР, гибелью тысяч ни в чём не повинных граждан, и вынужденными миграциями миллионов соотечественников.
Войны порождаются забвением необходимости различий, когда одни пытаются навязать другим свои ценности. Напротив, цивилизованный взвешенный учёт многообразия генофонда планеты поможет нам сохранить мир в условиях глобализации.
В заключение хотелось бы отметить, еще один немаловажный факт, который подтолкнул нас на перевод и издание этой книги на русском языке. В средствах массовой информации, по телевидению и в газетах у нас в стране часто, пусть даже и неявно проводится мысль о многовековой неприязни англичан к России. Господин Ричард Линн не только подлинный ученый мирового масштаба, но еще и джентльмен классического образца, а также искренний и бескомпромиссный любитель русской культуры.
Всех перечисленных факторов вполне достаточно, по нашему мнению, чтобы эта книга заняла свое достойное место на полках книжных магазинов, библиотек и на письменных столах всех думающих граждан Российской Федерации.
Член Союза писателей РФ В. Б. Авдеев Доктор филологических наук А. А. Григорьев Кандидат биологических наук Д. О. Румянцев
Г(1Ш I
mmwiи нэнср«1н£ пшют
Определение интеллекта
Иерархическая модель интеллекта
Коэффициент интеллекта (IQ)
Эффект Флинна
Расовые различия в интеллекте начали анализироваться с научной точки зрения с середины девятнадцатого столетия. В 1830-х гг. Сэмюэль Мортон (Morton, 1849) в Соединенных Штатах собрал коллекцию черепов, измерил их объемы и вычислил, что европеоиды обладают наибольшим объёмом мозга, за ними следуют китайцы, малайцы, американские индейцы, а у африканцев и, наконец, у австралийских аборигенов объём мозга наименьший. Он заключил, что выявленные различия объёма мозга ответственны за расовые различия в интеллекте. Сходная точка зрения была представлена спустя несколько лет во Франции Полем Брока (Вгоса, 1861, стр. 304): «В целом, мозг больше у выдающихся людей, чем у обладающих посредственными способностями, больше у высших, чем у низших рас». Примерно в том же самое время Фрэнсис Гальтон (Galton, 1869) в Англии пришёл к тому же выводу другим путём. Он оценил интеллект рас по числу принадлежащих к этой расе гениев в отношении к размеру популяции. Он заключил, что греки классических Афин были самыми интеллектуально одарёнными людьми, за ними в убывающем порядке следуют равнинные шотландцы, англичане, африканцы и австралийские аборигены.
В двадцатом столетии этот вопрос продолжал обсуждаться. Альфред Бине во Франции в 1905 разработал тест интеллекта. Тест был переведен на английский язык в 1916 г. профессором психологии Стенфордского университета Льюисом Терменом (Terman, 1916). Позднее в XX столетии были разработаны многие другие тесты интеллекта. Всё это сделало возможным измерение и сравнение интеллекта разных рас, и к концу двадцатого столетия по этой проблеме были опубликованы сотни исследований. Большинство из них было посвящено различиям между чернокожими и белыми в Соединенных Штатах, но исследования интеллекта проводились практически среди всех народов мира. Что касается различий между чернокожими и белыми в Соединенных Штатах, то самыми авторитетными работами являются монография Шу (Shuey, 1966), подытожившего все исследования
18
Ричард Линн
от Первой мировой войны до 1965 г., работа Осборна и МакГурка (Osborne, McGurk, 1982), дополнивших обзор Shuey данными по 1980 г., книга Лоэ- лина, Линдзи и Спулера «Расовые различия в интеллекте» {«Race Differences in Intelligence»; Loehlin, Lindzey, Spuhler, 1975), книга «Кривая колокола» Хернстайна и Мюррея {«The Bell Curve»; Herrnstein, Murray, 1994), а также серия работ Дженсена, кульминационной среди которых является книга «Фактор g» {«The g Factor»', Jensen, 1998). Был проявлен некоторый интерес к изучению интеллекта китайцев и японцев, материал был обобщён Верноном в монографии « The Abilities and Achievements of Orientals in North America» (Vernon, 1982). Большое число исследований интеллекта африканцев, европеоидов и аборигенов Восточной Азии1 было суммировано Раштоном в книге «Раса, эволюция и поведение» {«Race, Evolution and Behavior»', Rushton, 2000). Все эти исследования касались двух проблем. Это свидетельство о существовании расовых различий в интеллекте и вопрос о степени обусловленности этих различий генетическими и средовы- ми факторами. Многие специалисты признают, что межрасовые различия в интеллекте существуют, но нет консенсуса в вопросе о том, имеется ли под ними какой-либо генетический фундамент. Все вышеназванные авторы утверждали, что существует некое генетическое основание под выявленными межрасовыми различиями. Но многие авторитетные специалисты утверждают, что убедительных свидетельств в пользу генетических факторов нет. Эту позицию занимают Флинн в его книге «Race, IQ and Jensen», (Flynn, 1980), Броди в «Intelligence»(Brody, 1992) и Макинтош в «IQ and Human Intelligence» (Mackintosh, 1998).
Настоящая книга отличается от предыдущих исследований в четырех отношениях. Во-первых, это самый всеобъемлющий обзор работ по выявлению расовых различий в интеллекте во всем мире. Во-вторых, рассматриваются десять, а не три большие расы (африканцы, европеоиды и аборигены Восточной Азии), анализировавшиеся Раштоном (Rushton, 2000). Нами рассматриваются следующие расы: европейцы, аборигены экваториальной Африки2, бушмены, аборигены Южной Азии и Северной Африки, аборигены Юго-Восточной Азии, австралийские аборигены, аборигены островов Тихого океана, аборигены Восточной Азии, арктические народы, американские индейцы. Данные представлены в главах 3-12; в главе 13 подводятся итоги исследования и обосновывается надёжность и
1 Автором используется принятые в настоящее время в англоязычной литературе названия больших рас. В отечественной литературе им соответствуют термины негроиды, европеоиды и монголоиды. По возможности, мы старались сохранять авторскую терминологию. — прим. перев.
2 У Р. Линна используется термин «Sub Saharan Africa» — Африка южнее Сахары. Хотя в отечественной литературе более распространены термины «экваториальная Африка» или «чёрная Африка», здесь мы также придерживаемся терминологии автора. — прим. перев.
Расовые различия в интеллекте
19
валидность оценок IQ рас. В-третьих, в главе 14 обсуждается, в какой степени межрасовые различия в интеллекте обусловлены средовыми и генетическими факторами. В-четвертых, в главах 15, 16 и 17 обсуждается, как расовые различия в интеллекте эволюционировали в течение последних приблизительно 100.000 лет. Этим обсуждениям предшествуют рассмотрение в настоящей главе природы интеллекта, измерения его расовых различий и концепции расы в главе 2.
Определение интеллекта
Широко распространено мнение, что интеллект — целостное образование, которое определяет эффективность решения задач, научения и запоминания. Плодотворное определение интеллекта было предоставлено комитетом под председательством Ульриха Найссера, учреждённым Американской психологической ассоциацией в 1995 г. и состоявшем из одиннадцати американских психологов, уполномоченных составить отчёт о том, что является известным и общепринятым относительно интеллекта. Определение интеллекта, предложенное целевой группой, состояло в том, что интеллект — способность «понимать сложные идеи, эффективно приспосабливаться к окружающей среде, учиться из опыта, использовать различные формы рассуждения, преодолевать препятствия посредством мышления» (Neisser, 1996, стр. 1). Это определение является в целом приемлемым, за исключением положения об эффективной адаптации к окружающей среде. Все виды живых существ эффективно приспособлены к своей среде обитания, иначе они не выжили бы, но многие виды, такие как змеи и другие рептилии, не могут считаться интеллектуальными. В экономически развитых странах низший слой с его культурой долгосрочной безработицы, преступности, зависимости от наркотиков и пособий матерям-одиноч- кам хорошо приспособлен к своей среде обитания, поскольку в состоянии жить на пособия и воспроизводиться, но при этом имеет низкий средний IQ, как было подробно показано Хернстайном и Мюрреем (Hernstein, Murray 1994), и не является интеллектуально-развитым как в любом разумном смысле слова, так и по тестам интеллекта.
Определение, которое свободно от этого неправильного положения, было предложено Готтфредсон и подтверждено 52 ведущими экспертами. Оно было опубликовано в «Wall Street Journal»в 1994 г.:
«Интеллект — очень общая умственная способность, которая, помимо прочего, включает способность рассуждать, планировать, решать проблемы, мыслить абстрактно, постигать сложные идеи, быстро учиться и учиться на основании опыта. Это не просто книжные знания, узкий академический навык или умение выполнять тесты. Скорее он отражает более широкую и глубокую способность к осмыслению
20
Ричард Линн
окружающего — «схватывание», «постижение смысла» вещей или «понимание» того, что делать» (Gottfredson, 1997, стр. 13).
Интеллект концептуализируется как единая сущность, которая может быть измерена тестами и численно выражена как IQ (коэффициент интеллекта). Теория интеллекта как преимущественно единой сущности впервые была выдвинута в первом десятилетии двадцатого столетия Чарльзом Спирменом (Spearman, 1904), который показал, что все познавательные способности положительно коррелируют друг с другом, так что люди, которые хорошо справляются с одними проблемами, обычно успешно справляться и со всеми другими. Спирмен разработал статистический метод факторного анализа, чтобы показать, что выполнение всех когнитивных задач частично детерминировано общим фактором. Он предложил обозначение gдля этого фактора «общего интеллекта». Чтобы объяснить существование этого единого фактора Спирмен предположил, что должна существовать некая общая умственная энергия, определяющая выполнение всех познавательных задач и ответственная за их положительную интеркорреляцию.
Иерархическая модель интеллекта
Спирмен предположил также, что помимо фактора gсуществует ряд частных способностей, которые определяют успешность решения специфических задач сверх действия g.В 1930-х гг. Терстоун (Thurstone, 1938) выдвинул альтернативную теорию, суть которой в том, что существует семь «первичных способностей», которые он определил как рассуждение, вербальное понимание, числовые способности, пространственные способности, беглость речи (способность за короткий промежуток времени назвать ряд слов, входящих в некоторое понятие), память искорость восприятия. Во второй половине двадцатого столетия было достигнуто общее согласие относительно того, что и модель Спирмена и модель Терстоуна правильны лишь отчасти, и что интеллект наилучшим образом концептуализируется как иерархическая структура, которая может быть представлена пирамидой, в основании которой лежит около семидесяти узких способностей (Спирменовские частные факторы), над ними — от восьми до десяти групповых факторов или факторов второго порядка (первичные способности по Терстоуну), а на вершине — единый общий фактор (фактор gСпирмена). В настоящее время доминируют конкретизации этой модели Хорна (Horn, 1991), Кэррола (Carroll, 1993), и МакГрю и Фланагана (McGrew, Flanagan, 1998). Их модели весьма сходны и предполагают, что от восьми до десяти факторов второго порядка включают: «флюид
Расовые различия в интеллекте
21
ную способность» (рассуждение), «кристаллизованную способность» (вербальное понимание), долговременную память, кратковременную память, визуализацию (визуальная и пространственная способности), числовую способность (арифметика), математическую способность, культурные познания, скорость обработки и время реакции. Эта иерархическая модель интеллекта признана многими современными авторитетами, такими как Американская целевая группа по интеллекту (Neisser, 1996), Дженсеном (Jensen, 1998), Макинтошем (Mackintosh, 1998), Дири (Deary, 2000) и многими другими. Исчерпывающее описание фактора g,его структуры, наследуемости, биологии, и его коррелятов было представлено Дженсеном (Jensen, 1998) в его книге «Фактор#». Он концептуализирует факторgкак конструкт или фактор, определяя его как «гипотетическую переменную, которая лежит в основе наблюдаемой или измеряемой переменной» (стр. 88). Невозможно измерить gнепосредственно, но данные невербальных тестов рассуждения, а также значения, полученные в тестах интеллекта и выражаемые как IQ (коэффициент интеллекта), являются приблизительными оценками g.
Коэффициент интеллекта (IQ)
Метрика, используемая для измерения интеллекта рас, состоит в том, что IQ европеоидов Великобритании, Соединенных Штатов, Австралии и Новой Зеландии со средним 100 баллов и стандартным отклонением в 15 баллов принята как стандарт, по отношению к которому могут быть вычислены величины IQ других рас. Средние величины IQ европеоидов в этих четырех странах практически идентичны, как показано в Главе 3 (Таблица 3.1), так что тесты, построенные и стандартизированные на европеоидах в этих странах, являются эквивалентными инструментами для меж- расовых сравнений. В Великобритании, Австралии и Новой Зеландии тесты интеллекта были стандартизированы на европеоидах, то же имело место в Соединенных Штатах в первой половине двадцатого столетия. Во второй половине двадцатого столетия американские тесты обычно стандартизировались на общей популяции, включавшей значительное количество афроамериканцев и выходцев из Латинской Америки. В этих выборках стандартизации средний IQ по всей популяции принимают за 100; средний показатель интеллекта у европеоидов составляет приблизительно 102, в то время как у афроамериканцев — 87, а у выходцев из Латинской Америки приблизительно 92 (см., например, Jensen, Reynolds, 1982). Это означает, что при оценке IQ других рас с помощью американского теста, стандартизированного для всей американской популяции с использованием шкалы
22
Ричард Линн
со средним 100, значение следует уменьшить на 2 балла, чтобы получить IQ относительно 100 для американских европеоидов. Этой проблемы не возникает только с одним британским тестом, используемым в кросс-куль- турных исследованиях интеллекта. Это — прогрессивные матрицы, которые был стандартизированы на британских европеоидах. Тесты, используемые в исследованиях интеллекта рас, обозначены в таблицах аббревиатурами. Полные названия тестов и описание способностей, которые они измеряют, даны в Приложении.
В таблицах, в которых приведены данные исследований расовых различий в интеллекте, представлены величины IQ для общего интеллекта и, где это возможно, для главных первичных способностей — рассуждения, вербального понимания и визуализации. Величины IQ для общего интеллекта получены или с помощью тестов общего интеллекта, содержащих задачи на оценку рассуждения, вербальных способностей, визуализации, перцептивных способностей, памяти и иногда другие задачи, или с помощью тестов на способность к невербальному рассуждению, таких как прогрессивные матрицы, которые дают результаты, близкие к получаемым в тестах общего интеллекта (Carroll, 1993; Jensen, 1998). Также представлены и суммированы результаты нескольких исследований расовых различий в кратковременной памяти и в музыкальных способностях.
Эффект Флинна
Проблема с количественной оценкой расовых различий в интеллекте состоит в том, что показатели интеллекта увеличивались с 1920-х гг. во многих регионах мира. Это долговременное увеличение впервые было отмечено Смитом (Smith, 1942) на Гавайях и нашло подтверждение в нескольких последующих исследованиях, например Кеттелла в Великобритании (Cattell, 1951). После описания выявленных различий Джеймсом Флинном (Flynn, 1984,1987) они стали известны как эффект Флинна. Когда представляются результаты по величинам IQ популяций, должна быть сделана поправка на эффект Флинна, поскольку иначе популяции получают иллюзорно высокие средние значения, будучи сравниваемыми со стандартными значениями, полученными у европейцев на много лет ранее. Величина эффекта Флинна варьирует в разных тестах. Средние величины IQ в тестах Векслера увеличивались в нескольких странах приблизительно на 3 балла за десятилетие с середины 1930-х до 1990-х гг., но вербальный IQ увеличивался примерно на 2 балла в десятилетие, а в тестах действия приблизительно на 4 балла за десятилетие (Flynn, 1984, 1998; Lynn, Pagliari, 1994). По стандартным прогрессивным матрицам средний пока
Расовые различия в интеллекте
23
затель IQ в Великобритании увеличивался со скоростью примерно 2 балла за десятилетие с 1938 г., когда тест был разработан, до 1979 г., когда была проведена его последняя британская стандартизация среди детей (Lynn, Hampson, 1986; Flynn, 1987). Показатели IQ в тесте Гудинаф «Нарисуй человека» увеличились в Соединенных Штатах на 3 балла за десятилетие между 1955 и 1968 гг., (вычислено по стандартизациям Харриса (Harris, 1963) и Министерства Здравоохранения, Образования и Социального Обеспечения Соединенных Штатов (1970)). Такая же скорость роста показателей в этом тесте была зафиксирована у чернокожих в Южной Африке с 1950 г. по 1988 г. (Richter, Griesel, Wortley, 1989). Поправки на эффект Флинна были сделаны для всех значений IQ популяций, представленных в последующих главах. В случае тестов, для которых не известна величина долговременного увеличения, предполагалось увеличение на 3 балла IQ за десятилетие.
Нет общепринятого представления относительно причин эффекта Флинна. Некоторые теории, выдвинутые ведущими экспертами, представлены в работе Найссера (Neisser, 1998). Ряд специалистов, включая самого Флинна (Flynn, 1987), полагают, что никакого существенного увеличения того, что можно назвать «реальным интеллектом» не было, и что увеличение обусловлено совершенствованием навыков работы с тестами. Другие, такие как Гринфилд (Greenfield, 1998), Макинтош (Mackintosh, 1998) и Вильямс (Williams, 1998) утверждают, что увеличение является действительным и, вероятно, обусловлено рядом факторов, таких как более стимулирующая познавательные способности окружающая среда, особенно телевидение, компьютерные игры, улучшение образования и повышение образованности родителей. Наличие эффекта Флинна в развитии младенцев, проявляющееся, например, в возрасте, в котором младенец в состоянии встать, делает сомнительным влияние перечисленных факторов. Вероятно, имел место подлинный рост интеллекта в результате улучшения питания, приводящего к увеличению роста, размера мозга и, вероятно, к ускорению неврологического развития мозга в течение двадцатого столетия (Lynn, 1990а, 1998b).
Г(1Ш 2
ттт и формиромм pjk
Формирование рас, разновидностей и пород
Разновидности в животном мире
Таксономии рас
Расовые различия в заболеваемости
Существуют ли расы?
Имея дело с расовыми различиями в интеллекте необходимо определить как интеллект, так и расу. В предыдущей главе был определён интеллект, в этой главе предлагается определение расы. Простое и прямое определение состоит в том, что раса является группой, которая заметно отличается от других групп. Более полное определение: раса — это воспроизводящаяся популяция, которая в определённой степени генетически отлична от соседних популяций, что возникает в результате географической изоляции, культурных факторов и эндогамии, демонстрирующая характерные наборы генотипических различий в частотах ряда интеркоррелированных, генетически детерминированных характеристик при сравнении с другими воспроизводящимися популяциями. В зонах географического контакта между расами обычно имеются межрасовые гибриды, для которых характерны промежуточные значения частот встречаемости гена в сравнении центральными распределениями в смешивающихся группах. Эти гибриды и популяции смешанных рас известны как переходные формы или клины.
Формирование рас, разновидностей и пород
Общим принципом эволюционной биологии является то, что когда популяции биологического вида оказываются изолированными друг от друга, они эволюционируют в два или более подвида. Эти подвиды обычно называют разновидностями, штаммами или породами. В случае человека разновидности называют расами. Эти разновидности развиваются в результате четырех процессов, а именно эффекта основателя, дрейфа генов, мутаций и адаптации. Эффект основателя заключается в том, что когда
Расовые различия в интеллекте
25
популяция разделяется и одна из групп мигрирует к новому местообитанию, то группа, которая мигрирует, не будет генетически идентична оставшейся на прежнем месте. Следовательно, эти две популяции будут отличаются генетически. Эффект дрейфа генов состоит в том, что частоты встречаемости генов изменяются с течением времени до некоторой степени в случайном порядке, и это приводит к различиям между популяциями. Дрейф продолжается, и со временем приводит к увеличению различий между расами. Влияние мутаций заключается в том, что новые аллели (аллели — альтернативные формы генов) появляются в популяциях в случайном порядке, и если они благоприятны для выживания и воспроизводства, то будут постепенно распространяться в популяции. Благоприятная новая аллель может появиться в результате мутации в одной расе, а в других нет. Эффект адаптации состоит в том, что после миграции популяции к новому местообитанию некоторые аллели, не являвшиеся благоприятными на старом местообитании, становятся благоприятными. Особи, обладающие благоприятными аллелями, на новом местообитании производят больше жизнеспособного потомства, так что их аллели будут отбираться и постепенно распространяться в популяции. Новые разновидности нескольких видов развились как адаптивные, когда их популяции мигрировали в арктическую зону. У некоторых из них, таких как лисы, медведи или зайцы, образовался белый мех, который маскирует их, и они не так легко могут быть замечены хищниками или добычей. Во всех этих случаях белый мех появился вследствие мутаций и распространился в популяции потому, что давал животным преимущество в процессе отбора. В конечном итоге, новые благоприятные аллели полностью заменяют менее благоприятные и, как тогда говорят, они становятся «закреплёнными».
Во многих случаях неясно, почему разные разы эволюционировали по тем или иным признакам. Например, мех у европейской белки рыжий, в то время как североамериканские белки серые. Возможно, один из этих цветов даёт преимущество в процессе отбора, и появился случайно в одной из популяций в результате генетической мутации.
Разновидности в животном мире
Было давно установлено, что большинство биологических видов имеют несколько разновидностей, которые в случае человека называют расами. В начале своей карьеры Чарльз Дарвин обратил внимание на различные варианты черепах на Галапагосских островах, и именно это заставило его задуматься о том, как они развились. Позже, в книге «Изменение домашних животных и культурных растений»(Darwin, 1868), он описал породы
26
Ричард Линн
многих биологических видов, в частности породы голубей, каждая из которых имеет свою особенную манеру полёта, движения и воркования.
Существует много разновидностей или рас среди обезьян. Имеется четыре расы шимпанзе. Это истинный шимпанзе (Pan satyrus verus),обитающий в западной Африке между Гвинеей и Нигерией, лысый шимпанзе (Pan satyrus satyrus) из Камеруна и Габона, карликовый шимпанзе (Pan satyrus paniscus) с севера центрального Заира, и шимпанзе Швейнфурта (Pan satyrus schweinfurthi) из северо-восточного Заира. Эти расы отличаются по физическому облику, по распределению групп крови и по крикам, которые они издают. Различные расы развились среди животных видов в соответствии с теми же принципами, что и у человека. Например, существует две расы горилл. Это горная горилла (Gorilla beringei),обитающая в горах вокруг озёр Эдвард и Киву в восточном Заире, Руанде и западной Уганде, и береговая горилла (Gorilla gorilla)из лесов Камеруна и Габона. Эти две расы географически отделены друг от друга расстоянием примерно в тысячу миль и у них развились различия в физическом облике и группе крови. У горной гориллы более узкий череп, более короткие руки, более длинные ноги, более толстые волосы и группа крови А, в то время как у береговой гориллы череп шире, руки длиннее, ноги короче, волосы тоньше и группа крови В (Baker, 1974). Некоторые из различий между двумя расами развились в ходе адаптации к особенностям среды обитания. Горная горилла населяет более холодную и открытую местность, в то время как береговая горилла населяет более теплую и густо покрытую лесом территорию. У горных горилл развились более толстые волосы, чем у горилл побережья, для защиты от холода. У береговых горилл развились более длинные руки, чтобы перепрыгивать с дерева на дерево. Нет очевидного объяснения тому, почему у горной гориллы более узкий череп, более длинные ноги и группа крови А. Эти различия возникли, возможно, вследствие эффекта основателя, дрейфа генов или случайных мутаций, или же они могут давать животным какие-то неизвестные нам преимущества.
Существует также множество разновидностей домашних животных. Они обычно называются породами и были выведены человеком в различных практических целях. Часто породы выводились с целью увеличения размера животных или, в случае рогатого скота, надоев молока. В некоторых случаях выводились породы, лучше приспособленные к определённой среде обитания. Например, для выпаса на горных пастбищах были выведены породы выносливых овец, отличающиеся от овец низменных местностей. Человеком выведено целых семьдесят девять пород собак с различными способностями, такие как ретриверы для игр, овчарки как помощники в выпасе овец, ротвейлеры для охраны жилища, кокер-спаниели как домашние любимцы и так далее. Эти породы отличаются по общему интеллекту, по специфическим способностям и по лёгкости дрессировки (Согеп, 1994).
Расовые различия в интеллекте
27
Таксономии рас
Биологи и антропологи начали анализировать и классифицировать расы в середине восемнадцатого столетия. Первая таксономия рас была выработана шведским биологом Карлом Линнеем в 1758 г. В своей «Системе природы» он описал четыре человеческие расы, которые он назвал Europaeus(европейцы), Afer(чёрные африканцы), Asiaticus(азиаты) и Americanus(американские индейцы). В 1776 г. немецкий врач Иоганн Фридрих Блюменбах добавил пятую расу и предложил классификацию, основанную преимущественно на цвете кожи. Он называл эти расы кавказская (белая), монгольская (жёлтая), эфиопская (чёрная), американская (красная) и малайская (коричневая). Эти таксономии были основаны на объединении морфологических особенностей и цвета кожи разных рас, как то белая кожа, прямые волосы и узкий нос у европейцев, чёрная кожа, вьющиеся волосы и широкий нос у африканцев, тёмные волосы, желтоватая кожа и уплощённый нос у монголов (аборигенов Восточной Азии), красноватая кожа и клювообразный нос у американских индейцев, и коричневая кожа у малазийцев. Мортон (Morton, 1849) использовал пяти-расовую классификацию Блюменбаха, когда он выполнил первый сравнительный анализ размера мозга у разных рас.
В начале двадцатого столетия были собраны данные о различиях в частотах групп крови человека в разных популяциях всего мира. Хирш- фельд и Хиршфельд (Hirszfeld, Hirszfeld, 1919) показали, что частоты некоторых групп крови связаны с расовыми различиями в окраске кожи и в морфологии. Например, группа крови А присутствует у 41-48 процентов европейцев, но только приблизительно у 28 процентов африканцев района Африки южнее Сахары, в то время как группа крови В наличествует у 10-20 процентов европейцев и приблизительно у 34 процентов африканцев. У американских индейцев практически не встречаются группы крови А или В, у них почти у всех группа крови 0.
Данные о распределении групп крови по резус-фактору (Rh) были использованы Бойдом (Boyd, 1950) для следующей пяти-расовой таксономии: (1) Европейцы с высокими частотами групп крови Rh cde и cde; (2) Африканцы с высокими частотами Rh cde; (3) Аборигены Восточной Азии с высокой частотой В и фактическим отсутствием cde; (4) Американские индейцы с очень высокой частотой 0, отсутствием В и редкой cde и (5) Австралоиды с высокой частотой А и крайне редкими В и cde. Этот анализ показал, что распределение групп крови согласуется с расовыми таксономиями классической антропологии по морфологии и окраске кожи.
28
Ричард Линн
Более детальная таксономия рас была разработана Куном, Гарном и Бёрдселлом (Coon, Gam & Birdsell, 1950), они предложили рассматривать семь основных рас, каждая из которых подразделяется на две или более подрас. По их классификации основными расами являются: (1) кавказои- ды, подразделяющиеся на нордиков Северо-Западной Европы, славян Северо-Восточной Европы, альпийцев центральной Европы, средиземноморцев Южной Европы, северной Африки и ближнего Востока, и индостанцев Индии и Пакистана; (2) аборигены восточной Азии, подразделяющиеся на тибетцев, северных китайцев, классических аборигенов Восточной Азии (корейцы, японцы, монголы) и эскимосов; (3) аборигены Юго-Восточной Азии, подразделенные на южных китайцев, тайцев, бирманцев, малайцев и индонезийцев; (4) американские индейцы, подразделенные на северных, центральных, южных и огнеземельцев; (5) африканцы, подразделенные на аборигенов Восточной Африки, суданцев, аборигенов Западной Африки, банту, бушменов и пигмеев; (6) аборигены островов Тихого океана, подразделенных на меланезийцев, микронезийцев, полинезийцев и негритосов и (7) австралийские аборигены, подразделенные на народы Murrayian Юго-Восточной Австралии и CarpentarianСеверной и Центральной Австралии. Бейкером (Baker, 1974) была предложена близкая таксономия с семью расами, включающая пять главных рас по Блюменбаху и Кой бушменов, представленных готтентотами и бушменами Юго-Западной Африки и пустыни Калахари, а также австралидов, представленных австралийскими аборигенами и меланезийцами.
В 1980-х и 1990-х гг. Ней и Райчаудхури (Nei, Roychoudhury, 1993), а также Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994) разработали новый метод классификации человеческих рас на основе ряда генетических полиморфизмов (под полиморфизмом подразумевается, что ген имеет более одной аллели, или альтернативной формы). Методика состоит в том, что данные о частотах аллелей большого числа полиморфных генов, определяющих группы крови, состав белков крови, лимфоцитарных антигенов и иммуноглобулинов в популяциях по всему миру сводятся в таблицы. Табулированые данные затем подвергаются факторному анализу, чтобы найти степень, в которой частоты аллелей связаны и образуют кластеры популяций, генетически сходных друг с другом. Данные Нея и Райчаудхури по 26 популяциям были подвергнуты факторному анализу Дженсеном (Jensen, 1998), выявившим существование в человечестве шести основных групп, которые близко соответствуют расам, выделяемым в классической антропологии. Используя традиционную терминологию это: (1) африканцы районов Африки южнее Сахары (пигмеи, нигерийцы, банту, бушмены); (2) европейцы (саамы, финны, немцы, англичане, итальянцы, иранцы, северные индусы); (3) аборигены Восточной Азии (японцы, китайцы, корейцы, тибетцы, монголы); (4) аборигены
Расовые различия в интеллекте
29
Юго-Восточной Азии (южные китайцы, тайцы, филиппинцы, индонезийцы, полинезийцы, микронезийцы); (5) америнды (индейцы Северной и Южной Америки и иннуиты3) и (6) австралийские аборигены (аборигены Австралии и Новый Гвинеи).
Такая же методика была использована Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994) для анализа большего набора данных по 120 аллелям в 42 популяциях. По этим данным вычислялись генетические различия каждой популяции со всеми остальными. По результатам вычислений было построено дерево генетических связей, группирующее популяции в то, что они назвали «кластерами». Исследователи выявили десять основных кластеров. Это: (1) бушмены и пигмеи; (2) африканцы районов Африки южнее Сахары; (3) аборигены Южной Азии и Северной Африки; (4) европейцы; (5) аборигены Восточной Азии; (6) арктические народы; (7) американские индейцы; (8) аборигены Юго-Восточной Азии; (9) аборигены островов Тихого океана; и (10) аборигены Австралии и Новой Гвинеи. Очевидно, что эта классификация близко соответствует расовым таксономиям классической антропологии, основанным на видимых особенностях окраски кожи, волос, глаз, пропорций тела, длины конечностей ит. п., но по какой-то причине Кавалли-Сфорца, Меноцци и Пиацци предпочли термин «кластеры».
Расовые различия в заболеваемости
Существуют межрасовые различия по заболеваемости многими болезнями, имеющими генетические корни, в число которых входят кистозный фиброз, фенилкетонурия (ФКИ), гипертония, инсульт, диабет, рак простаты, рак молочной железы, тучность, близорукость и шизофрения. Эти различия возникли вследствие эффекта основателя, дрейфа генов, мутаций и адаптации. Имеется огромный массив исследований на эту тему, изложение результатов которых составило бы целую книгу. В качестве иллюстрации мы приводим частоты встречаемости гена кистозного фиброза и ФКИ у европейцев, африканцев из Африки южнее Сахары и аборигенов Восточной Азии из работы Бодмера и Кавалли-Сфорца (Bodmer, Cavalli-Sforza, 1976). Данные приведены в Таблице 2.1. Числа обозначают частоты встречаемости гена (процент распространенности) в популяции. Как можно видеть, частоты встречаемости гена муковисцидоза среди европейцев в четыре-пять раз выше, чем среди африканцев районов Африки южнее Сахары и аборигенов Восточной Азии, а частоты встречаемости
3 Самоназвание эскимосов {прим. перев.)
30
Ричард Линн
гена ФКИ выше среди европейцев в два с небольшим раза, чем в двух других расах. В нижней части таблицы показано, что частоты встречаемости генов двух указанных болезней весьма сходны в различных европеоидных популяциях, столь удалённых друг от друга странах, как то: Австрии, Австралии, Канаде, Англии и Соединенных Штатах.
Таблица 2.1. Частоты встречаемости (проценты) гена кистозного фиброза и ФКИ у европейцев, африканцев районов Африки южнее Сахары и аборигенов Восточной Азии

Существуют ли расы?
С восемнадцатого столетия до середины двадцатого все антропологи, биологи и социологи признавали, что человеческий род состоит из ряда биологически различимых рас. Так, в 1920-х гг. британский антрополог сэр Артур Кейт писал:
«Человеческие типы различаются настолько очевидно, что из толпы людей, собранной из австралоидов, негроидов, жителей Восточной Азии и кавказоидов, антрополог смог бы отличить человека одного типа от другого без колебания или ошибки» (Keith, 1922 p. xviii).
Курьёзно, но это кажущееся бесспорным утверждение начало оспариваться с середины двадцатого столетия, когда многие антропологи стали заявлять, что рас не существует. Одним из первых занял эту позицию антрополог Эшли Монтагю (Montagu, 1945а) в своей книге «Самый опасный миф человечества: Заблуждение расы». Название предполагает, что понятие расы — миф, и, следовательно, никаких рас нет. Но из текста книги становится ясным, что для Монтагю расы всё же существуют. Он писал:
Расовые различия в интеллекте
31
«В биологии под расой понимают подразделение биологического вида, которое наследует физические особенности, отличающие его от других популяций вида. В генетическом смысле раса может быть определена как популяция, которая отличается по частоте определенных генов от других популяций, с одной или несколькими из которых она способна к обмену генами через барьеры (обычно географические), которые могут разделять их. Если нас спрашивают, существует ли в этом смысле определённое количество рас в человеческом роде, то ответ будет — да» (стр. 6).
Из этого ясно, что раса не «миф» и не «заблуждение». Принимая во внимание то, что Монтагю очевидно признал существование рас, кажется странным, что он дал своей книге такое вводящее в заблуждение название.
Позже, во второй половине двадцатого столетия, многие антропологи и генетики стали утверждать, что рас не существует. В 1962 г. антрополог Ф. Б. Ливингстон (Livingstone, 1962) опубликовал работу: «О несуществовании человеческих рас», в которых он заявил, что «нет никаких рас, есть только переходные формы» (стр. 279). Переходные формы — гибриды между двумя чистыми расами. Переходные формы неизменно появляются в зонах контакта подвергающихся интребридингу рас, производящих расо- вые-смешанные гибриды. Так, в Латинской Америке проживает большая популяция метисов, имеющих европейских и индейских предков, которая может считаться переходной формой. Аналогично, аборигены островов Тихого океана — переходная форма, возникшая в результате межрасового смешивания аборигенов Юго-Восточной и Восточной Азии. Часто утверждается, что существование промежуточных форм, градиентов признаков, или гибридов лишает правомочности понятие расы. Очевидно, что это не так. Переходные формы и гибриды собак называют полукровками, но существование полукровок не означает, что нет чистых пород.
Но в следующем десятилетии генетики Уолтер Бодмер и Луиджи Кавалли-Сфорца (Bodmer, Cavalli-Sforza, 1976, стр. 698) вынуждены были написать о «существовании многих расовых групп у человека» и что «расы можно назвать подвидами, если мы примем для человека критерий зоологической систематики. Критерием, позволяющим считать две или более групп подвидами, является наличие 75 или более процентов особей, составляющих группы, которые могут быть однозначно классифицированы как принадлежащие к определённой группе». Далее они пишут, что при более широком определении человеческих рас возможно идентифицировать расу при её более чем 75-процентном представительстве в популяции. Следовательно, человеческие расы безусловно существуют. Приблизительно двадцать лет спустя тот же Луиджи Кавалли-Сфорца предпочёл плыть по течению и мы читаем у него о «научной несостоятельности понятия человеческих рас» и что «концепция расы не получила признания» (Cavalli- Sforza, Menozzi, Piazza, 1994, стр. 19). Однако авторы пишут: «Мы можем идентифицировать «кластеры» популяций». Эти кластеры оказываются тем
32
Ричард Линн
же, что и расы классической антропологии, и далее в своей книге авторы используют классическую расовую терминологию. Например, они пишут, что Африка «населена двумя аборигенными группами, кавказоидами на Севере почти до южных границ Сахары и негроидами в районах Африки южнее Сахары» (стр. 167). Очевидно, они забыли свое предыдущее утверждение о том, что «понятие человеческих рас научно несостоятельно, человеческий род может быть разделен лишь на «кластеры»» (прозрачный эвфемизм для рас). Всего лишь шесть лет спустя тот же Луиджи Кавалли- Сфорца вновь явно передумал, так как заявил, что расы всё-таки существуют, и что раса может быть определена как «группа индивидуумов, которых мы можем распознать как биологически отличных от других» (Cavalli-Sforza, 2000, стр. 25). Возникает впечатление, что он решил отрицать существование рас, но время от времени он забывается, и слово р— проскальзывает.
К началу двадцать первого столетия отрицание существования рас становится все более и более частым. В 2004 г. Американская Антропологическая Ассоциация объявила на своём Интернет сайте, что «раса не является научно-обоснованной биологической категорией». «Нет никаких биологических рас», утверждает Джефферсон Фиш (Fish, 2002, стр. xii), профессор психологии из Университета Святого Иоанна в Нью-Йорке, но не поясняет, на каких основаниях он делает такое утверждение. Грейвс (Graves, 2002, стр. 2-5), биолог из университета штата Аризона, также утверждает, что «биологические расы не существуют», и пишет, что «термин раса подразумевает существование некоторых существенных основных наследственных особенностей, распространённых в одних группах людей и не представленных в других», и что это не верно для человеческих рас. Вопреки этому утверждению, существует ряд «наследственных особенностей», которые присутствуют в одних расах и отсутствуют в других. Например, гены, определяющие чёрную окраску кожи, присутствуют у африканцев и отсутствуют у европейцев, аборигенов Восточной Азии и американских индейцев, а гены, ответственные за развитие эпикантуса, присутствуют только у аборигенов Восточной Азии, у арктических народов и у некоторых американских индейцев. Более того, понятие расы не обязательно подразумевает, что существуют некоторые аллели (аллели — альтернативные формы генов), которые присутствуют только в одних расах и отсутствуют в других. Достаточно того, что существуют различия в частотах аллели между расами. Для ряда аллелей дело обстоит именно так. Например, аллель серповидно-клеточной анемии встречается намного чаще среди африканцев, чем среди других рас, а аллель кистозного фиброза намного шире распространена среди европейцев (Таблица 2.1).
Грейвс (Graves, 2002, стр. 5) пишет: «Большинство генетиков, эволюционных биологов и антропологов признают, что никаких биологичес
Расовые различия в интеллекте
33
ких рас в человеческом виде нет». Коэн (Cohen, 2002, стр. 211) также утверждает: «Почти все антропологи согласны в том, что расы в популярном смысле слова не существуют и никогда не существовали». Эти утверждения не верны. Обзор взглядов американских антропологов, выполненный в 1985 г., показал, что существование рас признавалось 59 процентами биологических и физических антропологов и приблизительно одной третью культурных антропологов (Lieberman и Reynolds, 1996).
Несмотря на отрицание существования рас многими американскими антропологами, реальность понятия расы широко признаётся среди остальной части общества. Медицинские журналы содержат многочисленные статьи на темы расовых различий в заболеваемости и инвалидизации, включая распространенность ВИЧ-инфекции. Издаётся журнал «Этническая принадлежность и здоровье», посвященный расовым различиям в распространенности болезней. В сфере общественных наук есть два журнала, посвященные расовым различиям («Раса и класс» и «Этнические и расовые исследования»). И в других журналах публикуются многочисленные статьи на темы расовых различий в интеллекте, достижениях в образовании, в доходах, социально-экономическом статусе, безработице, предубеждениях, дискриминации, потреблении алкоголя, употреблении табака, наркомании, сексуальном опыте, долголетии, преступности и задержке психического развития. Корпорации способствуют равным возможностям при приеме на работу для представителей разных рас. Служащие предъявляют иски к корпорациям за расовую дискриминацию и часто получают существенную компенсацию по решению судов, у которых нет проблем в понимании того, что такое раса. Многие университеты осуществляют положительную дискриминацию в пользу чернокожих и испано-говорящих соискателей. Судьи выносят решения о неконституционности расово-сег- регированных школ. Граждане многих стран заявляют свою расу в переписных листах, и эти листы анализируются социологами и демографами. В Великобритании есть Комиссия по расовым отношениям, задача которой состоит в том, чтобы способствовать равенству рас и преследовать по суду работодателей за расовую дискриминацию. Ни у людей, ответственных за эту работу, ни у широкой общественности нет трудностей в понимании того, что такое раса, и все они были бы поражены, узнав, что многие американские антропологи утверждают, что рас не существует.
Можно задаться вопросом, почему многие американские антропологи отвергают понятие расы. Ответ был дан двумя польскими антропологами (Kaszycka, Strkalj, 2002, стр. 334). Они пишут:
«Американцы стали чрезвычайно чувствительными к теме расы, и этот термин приобрел крайне щепетильную коннотацию. Многие американские ученые проголосовали за небытие человеческих рас. Кроме того, растущие требования «политической корректности» препятствуют использованию термина и внутри, и вне науки...
2 Ричард Линн
34
Ричард Линн
Немногие ученые отваживаются заниматься расовыми проблемами, опасаясь получить клеймо расиста только за то, что интересуются этой тематикой».
На причину отрицания понятия расы многими американскими антропологами указывает название книги Монтагю: «Самый опасный миф человечества». Очевидно Монтагю верил, что для людей осознание расы опасно, потому что оно имеет тенденцию питать расовые антагонизмы, которые могут перерасти в конфликт. Для предотвращения этого, было бы лучше понятие расы затушевать. В Европе большинство антропологов принимает правомочность понятия расы. Так, опрос польских антропологов, выполненный в 2001 г., показал что 75 процентов согласны с утверждением: «В пределах вида Homo sapiensимеются биологические расы» (Kaszycka, Strzalko, 2003). Преимущественно в Соединенных Штатах существование рас стало отрицаться многими антропологами и некоторыми биологами и социологами, принесшими свою научную честность в жертву политической корректности.
Г(1 т * шошнци
Интеллект автохтонных европейцев
Европейцы вне Европы
Европейцы — студенты университетов
Размер мозга
Наследуемость интеллекта у европейцев
Европейцы были признаны всеми классическими антропологами одной из основных рас. Карл Линней (Linnaeus, 1758) назвал её Europaeus. Европейцы часто назывались кавказцами или кавказоидами из-за поверья, что они происходят с Кавказа. Некоторые антропологи относили европейцев вместе с аборигенами Южной Азии и Северной Африки к единой кавказоидной группе. Однако европейцы отличаются от выходцев из Южной Азии и Северной Африки более светлым цветом кожи, а северные европейцы ещё светлыми волосами и голубыми глазами. Различие между европейцами и выходцами из Южной Азии и Северной Африки было подтверждено Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994), классифицировавшими человеческие расы по набору генетических маркеров. Этими авторами показано, что европейцы, представленные итальянцами, датчанами, англичанами и басками, образуют гомогенный «кластер», отделяясь от других рас. Кун, Гарн и Бёрдселл (Coon, Gam, Birdsell, 1950), Коул (Coul, 1965) и ряд других антропологов подразделили европейцев на семь подрас, а именно: Средиземноморские народы Испании, Италии и Юго-Восточной Европы; альпийские народы Франции, Центральной и Южной Германии; нордические народы Англии, востока Ирландии и Шотландии, Нидерландов, Бельгии, Северной Германии, Дании, Норвегии, Швеции и западной Финляндии; кельтские народы Уэльса, запада Ирландии, и западной горной части Шотландии; динарс- кие народы востока Центральной Европы; славянские народы северной Польши, государств Балтии и России к западу от Урала; баски северной Испании и Юго-Западной Франции. У нордических народов более светлый цвет кожи, светлые волосы, и голубые глаза, в то время как для европейцев Центра и Юга Европы типична более тёмная кожа, тёмные или чёрные волосы и тёмные глаза.
2*
36
Ричард Линн
Интеллект автохтонных европейцев
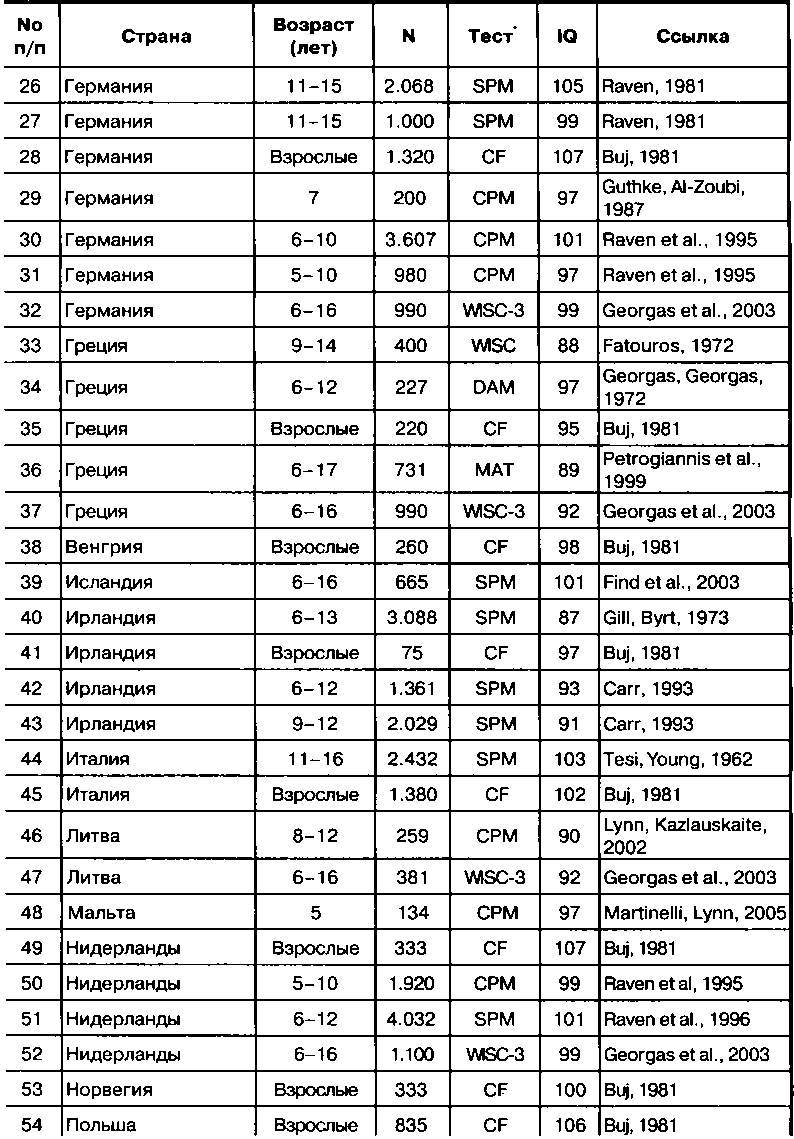
Результаты оценки коэффициента интеллекта IQ европейцев в Европе представлены в Таблице 3.1.
Таблица 3.1. Величины IQ автохтонных европейцев

*асовые различия в интеллекте
8
Ричард Лит

* Расшифровки аббревиатур использованных тестов приведены в приложена Тесты интеллекта».
Величины IQ были рассчитаны относительно среднего значения IQ бри- 'анцев, принятого за 100 (при стандартном отклонении в 15 единиц). В таб- шце представлены результаты двадцати одного исследования, выполненны? >ьюи (Buj, 1981) на выборках взрослых из больших городов. Большая часп >сгальных исследований была выполнена с использованием одной из трё> »ерсий прогрессивных матриц (СРМ, SPM и АРМ4). Представленный в стро- :е 61 коэффициент интеллекта для Сербии в 89 баллов надо, вероятно, счи- 'ать несколько заниженным, т. к. выборка, как она описана, была из «семей
4 СРМ — Цветные (Coloured) прогрессивные матрицы; SPM — Стандартные про рессивные матрицы; АРМ — Прогрессивные матрицы повышенной сложности (Advanced)
Расовые различия в интеллекте
39
преимущественно из низшего и нижней части среднего классов» из Белграда и его окрестностей (Moyles и Wolins, 1973, стр. 372). Диапазон величин IQ европейцев составляет от 87 баллов по данным одного из исследований в Ирландии и 88 баллов в одном исследовании в Греции, до 107 баллов по данным одного исследования в Германии и Нидерландах. Имеются некоторые противоречия результатов разных исследований, выполненных в одних и тех же странах, где показатели интеллекта обычно различаются на два-три балла IQ, а в случаях Португалии и Польши — на целых 13 и 14 единиц, соответственно. Эти различия частично вызваны ошибками выборок, но отчасти отражают действительные различия, обусловленные разницей в жизненном уровне и, возможно, неоднородностью расового состава европейских популяций. Ошибки выборок в исследованиях интеллекта в национальных популяциях возникают таким же образом, как при опросах о том, за кого намерены голосовать респондены, где обычно результаты нескольких опросов, выполненных в одно и то же время, дают результаты, различающиеся на несколько процентов. Нам не следует искать причины незначительных различий величин IQ в разных исследованиях, поскольку во многих случаях это просто ошибки выборки. Важно выявить основные паттерны.
Единственным значимым паттерном для коэффициента интеллекта в Европе является то, что величины IQ немного ниже на Юго-Востоке, чем в остальных частях Европы. На Балканах значения IQ следующие: 94 балла в Румынии, 92,5 (среднее по двум исследованиям) в Болгарии, 90 в Хорватии, 89в Сербии, и 92,5 (среднее значение четырех исследований) в Греции. Вероятное объяснение этого состоит в том, что балканские народы — гибридное население или переходная форма, являющаяся генетической смесью европейцев и аборигенов Южной Азии из Турции. Гибридные популяции или градиенты признаков возникают в пограничных областях между двумя расами в результате интербридинга. На Балканах такой градиент признаков развился из-за географической близости Юго-Восточной Европы и Турции и оккупации больших территорий Юго-Восточной Европы турками в течение многих столетий существования Османской империи. Это вызвало смешение турецких и европейских генов, так что в итоге современные турки и греки генетически весьма сходны. Это было показано Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994) на их дереве генетических связей, на котором греки расположены гораздо ближе к иранцам и другим народам Юго-Запада Азии, чем к итальянцам, датчанам и англичанам. Это генетическое сходство также очевидно и в случае интеллекта, коэффициент которого в Турции (90 баллов) весьма близок к таковому в Юго-Восточной Европе — в Греции, Румынии, Болгарии и Хорватии, где он варьирует от 90 до 94 баллов. Поскольку народы ЮгоВосточной Европы являются переходной формой, представляется уместным исключить их из оценки величины IQ европейцев. Медиана величины IQ
40
Ричард Линн
для остальных стран составляет 99 баллов и должна считается наилучшей оценкой величины IQ европейцев.
Помимо более низких величин IQ на Балканах, есть ещё три страны с величиной IQ несколько ниже среднеевропейской. Первая из них—Литва с IQ 90-92 балла. Эти низкие значения могут являться ошибками выборки, так как они заметно ниже, чем в соседних России (97), Польше (99) и Эстонии (99). Вторая страна — Ирландия, где среднее значение IQ по четырем исследованиям составило 92 балла. Наиболее вероятное объяснение этого — имеющая долгую историю эмиграция, когда люди с более высоким интеллектом имели тенденцию мигрировать, а менне интеллектуальные — оставаться. Это происходило также в Шотландии, где средний IQ равен 97, и на Корсике, где средний IQ ниже, чем в материковой Франции (Lynn, 1979, 1980). Третья страна с несколько сниженным IQ — Португалия, где результаты двух оценок величины IQ составили 101 и 88 балла, в среднем 94,5. Невысокий IQ в Португалии согласуется с тем, что там самый низкий доход на душу населения в Западной Европе и скромные интеллектуальные достижения. Португальцы получили всего одну Нобелевскую премию в области наук из 346, присуждённых за период 1901-2003 гг. Её удостоился в 1949 г. нейрохирург Антонио Мониц за внедрение операции префронтальной лейкотомии как метода лечения психических болезней, но ныне такая операция считается нежелательным лечением5. Возможно, интеллект в Португалии был снижен в результате примеси в популяцию выходцев из Африки. Португалия была единственной европейской страной, ввозившей с конца пятнадцатого столетия чернокожих рабов для сельскохозяйственных и домашних работ. Согласно Дю Буа (Du Bois, 1939, стр. 132-133), в шестнадцатом столетии число чернокожих превосходило численность белых в Лиссабоне и на плантациях Альграве на Юге страны. Может быть это преувеличение и, возможно, доля чернокожих уменьшилась в последующие столетия. Однако, если в нынешнем населении Португалии 20 процентов потомков выходцев из Африки, а средний IQ африканцев равен 70, можно ожидать, что величина IQ в популяции будет равна 94 баллам.
Может показаться удивительным, но, по-видимому, нет больших различий в величинах IQ между двенадцатью бывшими коммунистическими странами Восточной Европы, где медиана IQ равна 96 баллам, и четырнадцатью странами Западной Европы, где медиана IQ составляет 98,5 балла. Различие невелико и статистически незначимо. Так что, хотя в бывших коммунистических странах жизненный уровень был намного ниже в течение приблизительно шестидесяти лет с конца Второй Мировой Войны, это, по-видимому, не ослабило интеллект населения.
5 Использование этой операции в нашей стране было запрещено специальным приказом №1003 министра здравоохранения СССР Е. И. Смирнова от 9.12.1950 года, действующим до сих пор (прим. перев.)
Расовые различия в интеллекте
А'
Европейцы вне Европы
Европейцы мигрировали во многие части мира. Результаты исследо ваний интеллекта в европеоидных популяциях вне Европы представлень в Таблице 3.2.
Таблица 3.2. Коэффициенты интеллекта IQ европейцев вне Европы

42
Ричард Линн
В строках 1 и 2 приведены значения IQ в 93 и 98 баллов для Аргентины. В строке 3 представлен IQ в 97 баллов для Австралии, основанный на стандартизации американского теста Отиса. В строке 4 представлен IQ 100 для Австралии, измеренный у военнослужащих с помощью теста SPM (Величина IQ в этой выборке составляла 102 балла, но так как оценки мужчин по этому тесту в среднем на 5 баллов выше, чем у женщин (Lynn, Irwing, 2004), среднее значение было уменьшено до 100). В строке 5 приведён IQ 98 по выборке малолетних австралийских детей. Строка 6 дает IQ 95 для европеоидных детей в Бразилии из г. Сан-Паулу. В строке 7 представлен IQ 97 для Канады, полученной в выборке 7-12 летних детей. Строка 8 даёт IQ 100 для Канады, полученный из стандартизации теста WISC-3 на репрезентативной выборке в 2.200 детей возраста 6-16 лет.
В строке 9 представлен IQ в 99 балла для Чили, основанный на исследовании среди европеоидных студентов Католического университета г. Вальпараисо, показавшего, что их IQ не отличается от IQ австрийских студентов (п=320). В строке 10 представлен IQ в 95 баллов для европеоидных детей в Колумбии. В строке 11 приведён IQ 98 для европеоидных детей в Баха Калифорнии в Мексике. В строке 12 представлен IQ в 99 баллов для Новой Зеландии, полученный из стандартизации теста Отиса в 1930-х гг. Строка 13 дает величину IQ в 101 балл, полученную из стандартизации прогрессивных матриц. Строка 14 дает IQ 102, определённый в исследовании детского развития, выполненного христианской церковью. Строка 15 дает IQ 94 для 16-летних европеоидных детей в Натале в Южной Африке. В строках с 16 по 21 дано шесть значений величины IQ в диапазоне от 99 до 103 баллов для европеоидов в Соединённых Штатах, в сравнении с соответствующими значениями в Великобритании. Величина IQ в 100 баллов, приведённая в строке 20, выведена из стандартизации теста WAIS-3 в Великобритании. В строке 22 приведён IQ 96 из стандартизации прогрессивных матриц в Уругвае. В строка 23 дан IQ в 100 баллов для европеоидных 7-летних детей в Зимбабве.
Медиана приведённых величин IQ составила 99 баллов, так же как и у европеоидов в Европе. Результаты показывают, что даже в весьма бедных странах Латинской Америки (Аргентина, Бразилия, Колумбия, Мексика и Уругвай), в которых доходы на душу населения составляют приблизительно одну треть от доходов в Северной Америке и Западной Европе, величины IQ европеоидов лишь незначительно ниже, чем в богатых странах. Это подтверждают результаты по Европе, где в намного более бедных бывших коммунистических странах определяются примерно те же значения IQ, что и в богатых странах Запада.
Расовые различия в интеллекте
43
Европейцы — студенты университетов
Исследования интеллекта европеоидных студентов университетов подытожены в Таблице 3.3.
Таблица 3.3. Интеллект европейцев — студентов университетов

Как и можно было ожидать, во всех случаях величина IQ равнялась 100 баллам или выше, медиана составила 105 баллов. Главный интерес эти результаты представляют в плане их сопоставления с показателями, полученными среди аборигенных студентов университетов Африки и Южной Азии, где величины IQ обычно на 10-20 баллов ниже.
44
Ричард Линн
Размер мозга
В параграфе 1 настоящей главы мы отметили, что величины IQ в ЮгоВосточной Европе и на Пиренейском полуострове ниже, чем в остальной Европе. Мы можем ожидать, что подобные различия будут наблюдаться и в размере мозга вследствие того, что корреляции между размером мозга и интеллектом составляет 0,40 (Vernon, Wickett, Bazana, Stelmack, 2000). Рассмотрим различия между субпопуляциями европейцев, чтобы посмотреть, действительно ли это так. Данные о размерах мозга большого количества человеческих популяций, собранные Юргенсом, Ауне и Пипером (Jurgens, Aune, Pieper, 1990), представлены в Таблице 3.4 вместе с соответствующими значениями IQ.
Таблица 3.4. Размер мозга (смЗ) и интеллект европейцев и аборигенов Южной Азии

Из строки 1 видно, что у европеоидов в Северной Америке наибольшие размер мозга и IQ. Как видно в строке 2, за ними следуют европеоиды Северной, Центральной, и Восточной Европы. В строке 3 представлены немного более низкие значения размера мозга и величины IQ в Испании и Португалии. Как видно из строки 4, объём мозга и величина IQ в ЮгоВосточной Европе ещё меньше. Строка 5 показывает дальнейший нисходящий тренд размера мозга и величины IQ на Ближнем Востоке, измеренный на выборках выходцев из Южной Азии (из Турции и Ирака). В строке 6 приведены самые низкие значения размера мозга и IQ выходцев из Южной Азии (в Индии). Детально величины IQ аборигенов Южной Азии в Турции, Ираке и в Индии рассматриваются в Главе 6.
Расовые различия в интеллекте
45
Наследуемость интеллекта у европейцев
Наследуемость интеллекта — степень, в которой различия в интеллекте определены генетическими факторами. Здесь нас интересует вопрос о наследуемости расовых различий в интеллекте, но прежде чем обсуждать это, мы должны рассмотреть наследуемость индивидуальных различий в интеллекте в пределах стран. Имеются три источника данных по этой проблеме. Это исследования однояйцовых близнецов, воспитывающихся в разных семьях, сравнения однояйцовых и разнояйцовых близнецов, воспитанных в одной семье, и исследования не состоящих в родстве приёмных детей, воспитанных в одной семье. Результаты исследований всех трёх типов показывают, что наследуемость интеллекта у взрослых составляет приблизительно 0,80, или 80 процентов. Это означает, что если бы все люди воспитывались в одинаковых условиях, различия между индивидами уменьшились бы до 80 процентов от реально существующих.
Результаты работ по изучению наследуемости интеллекта среди взрослых и детей были подытожены Бушаром (Bouchard, 1993, стр. 58). По результатам пяти исследований среди взрослых однояйцовых близнецов, воспитанных порознь, средняя корреляция, взвешенная размером выборки, составила 0,75. Эту величину требуется скорректировать на надежность теста (коррекция на аттенюацию), приемлемое значение которого составляет около 0,9 (Bouchard, 1993, стр. 49; Mackintosh, 1998). Это поправка увеличивает корреляцию до 0,83, что и является мерой наследуемости. Из сравнения степени подобия между однояйцовыми близнецами и разнояйцовыми однополыми близнецами, воспитанными в одной семье, явствует, что корреляция для однояйцовых близнецов составляет 0,88, а для однополых разнояйцовых близнецов — 0,51. После коррекции на надежность тестов, принимая коэффициент надежности за 0,9, корреляции становятся 0,98 для однояйцовых и 0,56 для однополых разнояйцовых близнецов. Наследуемость может быть вычислена по формуле Фальконе (Falconer, 1960), заключающейся в удвоении разности между корреляциями однояйцовых и разнояйцовых близнецов одного пола. Разность между этими двумя корреляциями составляет 0,42, удвоение разности даёт наследуемость 0,84.
Третий метод оценки наследуемости интеллекта состоит в изучении корреляции между величинами IQ не состоящих в родстве детей, принятых на воспитание одной и той же семьёй. Величина влияния среды приёмной семьи («межсемейный эффект») выражается степенью корреляции между приёмными детьми. Обзор литературы, выполненный Бушаром (Bouchard, 1998), позволяет заключить, что среди взрослых корреляция
46
Ричард Линн
составляет 0,04, это свидетельствует о величине наследуемости 0,96. Однако, этот метод недооценивает влияние среды, так как не принимает во внимание воздействие на одного ребенка, но не на другого, таких факторов, как пренатальные и перинатальные воздействия. Два близнецовых метода, дающих оценки наследуемости интеллекта в 0,83 и 0,84, более точны. Эти значения очень близки к оценке примерно в 0,85, представленной Дженсеном (Jensen, 1998, стр. 179).
Наследуемость интеллекта у детей значительно ниже, приблизительно 0,42 у 4-6 летних и 0,55 для возрастной группы от 6 до 20 лет (Bouchard, 1993, стр. 58; Jensen, 1998, стр. 179). Причина этого вероятно в том, что родители оказывают на детей «средовые» влияния, которые ослабевают в подростковом возрасте. Именно из-за того, что некоторые исследователи объединяют более низкие показатели наследуемости интеллекта у детей с более высокими показателями у взрослых, они оценивают наследуемость интеллекта в пределах 0,40-0,80. Например, утверждение, сделанное Гот- тфердсон (Gottfredson, 1997, стр. 14) и поддержанное 52 экспертами, гласит: «Оценки наследуемости варьируют в диапазоне от 0,4 до 0,8, что скорее всего указывает на то, что генетика играет более значительную роль, чем среда, в формировании различий величины IQ между индивидами». Большинство исследований, в которых были получены эти высокие показатели наследуемости, проводились среди европейцев в богатых странах Запада. Однако, в исследовании 144 пар однояйцовых и разнояйцовых близнецов в России был получен показатель наследуемости 0,78, который после поправки на ненадежность теста возрос до 0,87 (Lipovechaja, Kantonistowa, Chamaganova, 1978).
Из вывода о высокой наследуемости интеллекта вытекает, что существуют гены, определяющие уровень интеллекта. Первый из таких генов был обнаружен в нормальных популяциях в конце 1990-х гг. Хорли и соавторами (Chorley et al., 1998). Он расположен в хромосоме 6, и обладание одной из аллелей этого гена привносит приблизительно 4 балла IQ в интеллект индивидума.
ГИШ 4 ДФРНШЦ41
Интеллект африканцев в Африке южнее Сахары
Студенты университетов в Африке
Африканцы островов Карибского моря и Латинской Америки
Афроамериканцы в Соединенных Штатах Америки
Африканцы в Великобритании
Африканцы в Нидерландах
Африканцы в Израиле
Кратковременная память и скорость восприятия у африканцев
Музыкальные способности
Время реакции
Размер мозга африканцев и европейцев
Африканско-европейские гибриды
Наследуемость интеллекта у афроамериканцев
Генетические средовые и объяснения различий IQ африканцев и европейцев
Оценка генотипного IQ у африканцев
Термин «африканцы» используется нами для обозначения основных аборигенных народов, обитающих в Африке южнее Сахары. Их следует отличать от северных африканцев — аборигенного населения районов Африки севернее Сахары, от пигмеев и от представителей другой расы африканцев, живущих южнее Сахары — бушменов или «койкоин»6, обитающих преимущественно в пустыне Калахари, а также от готтентотов Южной Африки; в настоящее время сохранились лишь нескольких десятков тысяч представителей этих народов. Использовалось множество терминов для обозначения африканских народов, в том числе «аферы» (Linnaeus, 1758), «эфиопы» (Blumenbach, 1776) и «негроиды» (Cole, 1965). Независимо от того, какое название использовалось, африканцы всегда расценивались как одна из больших рас в таксономиях классической антропологии, включая классификации Линнея (Linnaeus, 1758), Блюменбаха
6 Койкоин — самоназвание бушменов и готтентотов, означает «настоящие люди» — прим перев.
48
Ричард Линн
(Blumenbach, 1776), а также Куна, Гарна и Бёрдселла (Coon, Garn, Birdsell, 1950). Кавалли-Сфорца, Меноцци и Пиацци (Cavalli-Sforza, Menozzi, Piazza, 1994) подтвердили отличительные генетические особенности африканцев в своей классификации человечества на генетические «кластеры», в которой эти народы были представлены западными африканцами к Западу от Нигерии, нилотами верхнего Нила в Южном Судане, эфиопами и банту — большой группой, представленной в большей части Африки южнее Сахары от Нигерии на Западе до Кении. Главными отличительными особенностями африканцев являются: очень тёмная кожа, тёмные глаза, широкий нос, толстые вывернутые губы и густые курчавые волосы. Распределение групп крови у африканцев и европейцев различно. У африканцев реже встречается группа А (примерно 27 процентов против приблизительно 46 процентов у европейцев) и чаще группа В (около 34 процентов против примерно 14 процентов у европейцев).
Интеллект африканцев в Африке южнее Сахары
Первая попытка оценить интеллект африканцев была сделана Галь- тоном (Galton, 1869) на основе его собственных наблюдений во время путешествий по Юго-Западной Африке и записок других путешественников. Он построил шкалу уровней интеллекта, на которой одно деление было эквивалентно 10,425 баллам шкалы IQ. По его оценке африканцы стоят приблизительно на два деления ниже англичан, что равно 79 баллам по шкале IQ. Дальнейшими исследованиями интеллекта на выборках из всей популяции африканцев в районах Африки южнее Сахары было показано, что Гальтон переоценил IQ африканцев на одно с небольшим деление.
Результаты работ по определению коэффициента интеллекта африканцев в Африке южнее Сахары сведены в Таблицу 4.1. При необходимости приводятся пояснения результатов, изложенных в таблице.
В строке 1 приведён IQ в 64 балла для взрослых рабочих из Камеруна. В строке 2 дан IQ в 64 балла для молодых людей из Центральноафриканской республики, поступавших на технические курсы в колледже города Бангуи в 1951-1955 гг. В строках 3-5 даны IQ в 64 балла по результатам тестирований выборок из Конго-Браззавиля, проводившихся одновременно в городах Браззавиль и Пуэнт-Нуар. В строках 6, 7, 8,9 и 10 даны величины в 64,68,62,68 и 65 баллов IQ для Конго-Заира. В строке 11 приведён IQ 80 для взрослых в Гане. Такой IQ исключительно высок для Африки южнее Сахары. Возможно, столь высокое значение объясняется тем, что данные были получены в столице — в Аккре. Для столичных жителей типичны более высокие показатели IQ, чем в остальной части страны, вероятно
Таблицы 4.1. Показатели интеллекта африканцев в Африке южнее Сахары

Расовые различия в интеллекте

Расовые различия в интеллекте
51
вследствие того, что мигрировать в столицу склонны более интеллектуальные люди. Так, показатели интеллекта в Лондоне и Париже выше, чем в остальных частях Великобритании или Франции (Lynn, 1979, 1980). В строке 12 представлен IQ в 62 балла по репрезентативной выборке из всей Ганы. В строках 13 и 14 даны значения IQ 63 и 70 по результатам двух исследований в Гвинее. В строках 15-20 даны IQ в 69, 75, 69, 76, 89 и 63 балла для Кении. Величина IQ в 89 баллов в строке 19 была получена в группе 7-летних испытуемых в 1998 г. Это намного выше, чем IQ в 76 баллов, определённый теми же исследователями в их работе 1984 г. (строка 18) и чем любой другой IQ для популяций африканцев к югу от Сахары. Несоответствие указанного результата данным других исследований ставит под сомнение его достоверность. Так, в строке 16 приведён IQ 75, полученный в том же году в ходе стандартизации того же теста для всей Кении, а в строке 18 — IQ 76, тоже полученный в этом же году. Последние два показателя IQ типичны для Африки южнее Сахары и правдоподобны, но ставят под сомнение оценку в 89 баллов. Кроме того, увеличение IQ на 15 баллов от 76 до 89 баллов за 14-летний период уникально высоко для долговременного повышения IQ и не может считаться достоверным. Далее, с трудом верится, что у детей в Кении может быть более высокий IQ, чем у афроамериканцев в Соединенных Штатах, где IQ оставался постоянным на уровне приблизительно в 85 баллов с 1920-х гг., но где жизненный уровень и питание африканцев намного лучше, чем в Кении. По этим причинам мы считаем оценку IQ для Кении в 89 баллов недостоверной. Величина IQ в 63 балла, приведённая в строке 20, является средним значением между 65 баллами для 6-летних школьников и в 61 баллом для не посещавших школу, что даёт основание предположить, что обучение в школе повышает IQ на 4 балла.
В строке 21 приведён IQ в 82 балла для Мадагаскара. Хотя население острова обычно считается частью популяции районов Африки южнее Сахары, оно включает значительное количество аборигенов Юго-Восточной Азии — выходцев из Индонезии, откуда они мигрировали на Мадагаскар примерно в первом веке нашей эры (Cole, 1965). Население также включает африканцев и гибриды двух рас. Пропорции этих трёх групп в населении точно неизвестны, но предполагается, что преобладают потомки африканцев. Средний IQ для Мадагаскара (82 балла) выше любого из показателей IQ африканцев из районов Африки южнее Сахары, представленных в Таблице 4.1, за исключением сомнительных 89 баллов для Кении в строке 19. Коэффициент интеллекта населения в рассматриваемой популяции является промежуточным между африканцами (65-70 баллов) и аборигенами Юго-Восточной Азии (приблизительно 87, см. Главу 7), хотя он несколько ближе к IQ африканцев, как можно ожидать для смешанной популяции выходцев из Юго-Восточной Азии и африканцев, в которой преобладают
52
Ричард Линн
африканские гены. Исходя из теории средовой обусловленности IQ, невозможно дять очевидного объяснения того, почему IQ населения Мадагаскара выше, чем всюду в материковой Африке южнее Сахары.
В строке 22 дан IQ в 64 балла для Мозамбика. Выборка состояла из испытуемых, проучившихся 3 с половиной года в школе и включала несколько человек из бантустана Транскей и республики Малави. В строке 23 приведён IQ в 70 баллов для Нигерии, полученный у детей (возраст не указан), посещавших школы в городе Зария. Элитная выборка мальчиков из гимназии в этом же городе (п=179) показала IQ 81 балл. Применялся тест Леона, описанный автором как «разработанный африканцем для африканских детей» (Farron, 1966, стр. 53). Результат противоречит часто высказываемому утверждению, что африканцы испытывают затруднения при выполнении тестов, разработанных европейцами. В строках 24 и 25 даны значения IQ в 64 и 69 баллов по двум последующим исследованиям в Нигерии.
В строках 26 и 27 даны IQ в 64 балла для двух выборок взрослых в Сьерра-Леоне. В строке 28 приведён IQ в 65 баллов, полученный в первом исследовании интеллекта африканцев в Южной Африке, проведённом в 1920-х гг. В строках 29-40 даны оценки IQ в диапазоне от 58 до 77 баллов, полученные в двенадцати более поздних исследованиях. В строке 34 дан IQ 71 для тюремных надзирателей, имевших 9-12 летнее образование. В строке 35 приведён IQ в 77 баллов, который является самым высоким для ощепопуляционных выборок в Африке южнее Сахары, но он был получен с помощью теста «нарисуй человека», который является довольно плохим тестом общего интеллекта. В строке 38 представлен IQ 58 для большой выборки 16-летних школьников-африканцев, имевших примерно десятилетнее образование. Результаты сравнивались с южноафриканскими европеоидами. В строке 39 приведён IQ 69 для группы взрослых испытуемых, описанных как «компетентные мужчины, работающие длительное время в сложной окружающей обстановке...» (Nell, 2000, стр. 27). В строке 40 дан IQ 68 для подростков с несколькими годами школьного обучения из Северного Трансвааля. В строках 41 и 42 даны IQ 67 и 64 для двух выборок подростков из школ г. Соуэто. В строке 43 приведён IQ в 67 баллов для школьников третьего класса — зулусов из Наталя.
В строке 44 представлен IQ 69 для Судана, измерявшийся у детей и подростков народности Шиллук, описанной автором как «одно из примитивных негритянских нилотских племен» (стр. 164) Южного Судана. Приведённый IQ является средним значением измерений по четырём тестам: Лабиринты Годдарда и Портеуса, Александр пассалонг, и «нарисуй человека». В строках 45,46 и 47 даны IQ в 64,74 и 73 балла для трёх последующих исследований в Судане.
В строке 48 дан IQ в 78 баллов для группы учащихся средней школы в Танзании, это исключительно высокое значение для африканцев. Автор
Расовые различия в интеллекте
53
исследования объясняет, что причина этого в том, что выборка была высоко селективна, так как «число мест в средней школе чрезвычайно ограничено и поступление происходит на конкурсной основе» (Klingelhofer, 1967, стр. 207). Высокий IQ этой выборки не может считаться репрезентативным. Однако результат информативен, поскольку показывает, что IQ в элитной выборке из средней школы равен 78 баллам, а это свидетельствует о том, что типичные для Африки южнее Сахары значения IQ в диапазоне 65-72 баллов соответствуют действительности. В строках 49 и 50 приведены значения IQ в 65 и 72 балла для ещё двух репрезентативных выборок из Танзании, что согласуется с обычными значениями для Африки южнее Сахары. В строке 51 приведён IQ 80 для селективной выборки школьников в Уганде, описанных Верноном (1969, стр. 182) как «значительно превосходящих средний уровень восточно-африканского населения». Это объясняет, почему IQ в этой группе выше, чем в репрезентативных выборках из районов Африки южнее Сахары. В строке 52 дан IQ в 73 балла для большой репрезентативной выборки угандийских детей. В строках 53 и 54 приведены IQ 77 и 64 для Замбии. В строках 55 и 56 даны величины IQ в 61 и 70 баллов для Зимбабве, полученные Зинди, африканским психологом из Университета Зимбабве.
Самая поразительная особенность значений IQ африканцев из Африки южнее Сахары состоит в том, что они устойчиво значительно ниже, чем у европейцев (см. Таблицу 3.1 Главы 3). Медиана величин IQ составляет 67 баллов, что является наиболее достоверной оценкой IQ африканцев. За исключением значения IQ в 82 балла для Мадагаскара, которое является самым высоким в таблице из-за присутствия в населении выходцев из Юго-Восточной Азии, а также IQ в 78 (строка 48) и 80 баллов (строка 51) для селективных выборок из элитных средних школ в Танзании и Уганде, и сомнительных 89 баллов для Кении (строка 19), все значения IQ лежат в диапазоне 59-77 баллов, в то время как все оценки IQ европейцев попадают в диапазон 87-105 баллов. Диапазоны значений IQ в этих двух популяциях не перекрываются. Географическая изменчивость IQ африканцев не просматривается, вариации IQ могут быть следствием ошибок выборок и измерения. Величина IQ африканцев не претерпела никакого изменения со времени опубликования Пиком данных первого исследования (Pick, 1929), в котором IQ африканцев в Южной Африке был оценен в 65 баллов. В четырёх более поздних исследованиях африканцев в Южной Африке, выполненных в 1990-е гг., были определены фактически такие же IQ: 69 (Nell, 2000), 68 (Sonke, 2000), 67 и 64 балла (Skuy и др., 2001).
4
Ричард Лит
Студенты университетов в Африке
Опубликованы данные тринадцати исследований интеллекта африкан (ев — студентов университетов в Южной Африке. В некоторых работа? акже приведены оценки IQ европейских студентов, тестировавшихся в тс <е время. Результаты исследований приведены в Таблице 4.2.
'аблица 4.2. Показатели интеллекта африканских и европейских универси чешских студентов в Африке

В строке 1 представлен IQ в 75 баллов для африканцев — студентов ниверситета Легон в Гане, полученный с помощью методики Кубики !ооса из теста Векслера. Во всех остальных строках приведены результа- ы для Южной Африки. В строке 2 даны IQ в 84 балла для африканских и 103 балла для европейских университетских студентов, вычисленные от- осительно американских норм для взрослых, опубликованных в работе авена, Курта и Равен (Raven, Court, Raven, 1994). В строках 3 и 4 приве
Расовые различия в интеллекте
55
дены результаты студентов по тесту Блокса (Blox), показатели IQ африканцев представлены в сравнении с показателями для европеоидных южноафриканских студентов, принятыми за 100 баллов. Строка 5 даёт результаты тестирования с помощью теста WAIS-R студентов (средний возраст 25 лет) в африканских университетах Форт Харе, Зулуленд, Северный и Медицинского университета Южной Африки. Вербальный IQ составил 78, IQ достижений — 73 балла, что ещё раз показывает, что африканцы имеют низкий IQ по всем основным познавательным способностям и противоречит иногда высказываемому утверждению о том, что африканцы испытывают затруднения в выполнении языковых задач. В строке 6 приведён IQ в 100 баллов для студентов научных специальностей из Северного университета. В строке 7 дан IQ 77 для студентов менее престижного африканского университета. В строке 8 представлен IQ в 83 балла для студентов университета Витватерсранда и университета Африкаанс Ранда в Йоханнесбурге. В строке 9 представлен IQ 82 для африканских студентов из университета Венда в Северном Трансваале. Европейская группа, результаты тестирования которой используются для сравнения, была из университета Тилберга в Нидерландах. В строке 10 дан IQ 81 для студентов-психологов из университета Витватерсранда. В строке 11 представлен IQ 93 для студентов-первокурсников инженерных специальностей из университета Витватерсранда. В строке 12 дан IQ 99 для немного уменьшенной группы тех же студентов, оцененный 16 месяцев спустя с помощью прогрессивных матриц повышенной сложности. И африканцы, и европейцы показали результаты приблизительно на 6 баллов выше при втором тестировании, вероятно вследствие тренировки. В строке 13 приведён IQ в 101 балл для другой выборки студентов-африканцев инженерных специальностей из университета Витватерсранда; видно, что африканские студенты имели IQ на 15 баллов ниже, чем белые европейцы.
Средние величины IQ общих выборок студентов, представленные в строках 1-5 и 7-9, попадают в узкий диапазон значений 72-84 балла с медианой 81. Приведённые в строках 6, 11, 12 и 13 величины IQ в 100, 93, 99 и 101 балл выше остальных из-за того, что они были получены у студентов научных и инженерных специальностей, поступавших в университеты на основе вступительных экзаменов по математике и физике и, как правило, имевших более высокую способность к рассуждению, чем студенты большинства других специальностей. К примеру, в Иране IQ 18-летних учащихся, изучавших математику, был на 10 баллов выше, чем у изучавших литературу (Mehryar, Shapurian, Bassiri, 1972). Британские студенты, получавшие степень в области науки, имели IQ на 9 баллов выше, чем получавшие степень в области искусств (Heim, 1968). Величины IQ студентов- европеоидов в Южной Африке находятся в диапазоне от 100 до 105, что почти совпадает со значениями студентов-европеоидов в других странах
56
Ричард Линн
(см. Глава 3, Таблица 3.3). Интерес этих данных состоит в том, что они показывают, что типичные африканские студенты, имевшие примерно 12летнее школьное образование и поступившие в университет, имеют IQ в диапазоне 72-84 балла. Так как они — африканская интеллектуальная элита, эти данные позволяют сказать, что IQ для генеральной популяции около 70 баллов достоверен и в общем правилен. Результаты также свидетельствуют о том, что IQ африканских студентов в Южной Африке в среднем на 20 баллов ниже, чем у европеоидных и что значительный разрыв значений между величинами IQ африканцев и европейцев сохраняется и тогда, когда они имеют одинаковую продолжительность обучения. Африканские студенты университетов, имевшие за плечами десять-двенадцать лет систематического образования, помимо изучавших математику и физику, получали оценки IQ в диапазоне 72-84 баллов. Коэффициент интеллекта студентов был на 10-12 баллов выше средне-африканского вследствие того, что они являются селективной группой.
Африканцы островов Карибского бассейна и Латинской Америки
Результаты определений IQ у африканцев с островов Карибского бассейна и из Латинской Америки приведены в Таблице 4.3.
В строке 1 дан IQ 80 для детей на Барбадосе. Это значение, как сообщается в исследовании, было вычислено из значения IQ в 83 балла для хорошо питавшихся и в 68 баллов для голодавших детей с учётом показателя преобладания умеренного и серьёзного недоедания дошкольников на Барбадосе (16,5 процентов — по данным обследования 1968 г. (см. Galler, Ramsay, Solimano и др., 1983)). В строке 2 дан IQ 70 для африканцев в Бразилии, посещавших школу в фавеле (трущобный район) в г. Бразилиа. В строке 3 дан IQ 64 для матерей этих детей. В строке 4 дан IQ 71 для африканцев в г. Сан-Паулу в Бразилии. В строке 5 представлен IQ 67 для 3-летних африканских детей из Доминиканской республики. Низкий IQ этих младенцев даёт основание предполагать, что плохое образование не является фактором, ответственным за низкие величины IQ африканцев островов Карибского бассейна. В строках 6-12 приведены результаты семи исследований IQ на Ямайке, давших оценку в диапазоне 60-75 баллов с медианой 67. В строке 13 дан IQ 60 для 4-летних детей с о. Сент-Люсия, а в строке 14 — IQ 70 для детей с о. Сент-Винсент. Медиана оценок четырнадцати исследований интеллекта африканцев с островов Карибского бассейна и из Латинской Америки составила 71 балл IQ. Это немного выше, чем средний IQ африканцев в Африке южнее Сахары, составляющий 67 бал
Расовые различия в интеллекте
57
лов. Возможное объяснение этого в том, что у африканцев островов Карибского бассейна и Латинской Америки есть некоторая примесь европейских генов. Как было оценено, доля европейских генов в африканском населении Ямайки составляет 6,8 процентов (Parra, Marcini, Akey, 1998).
Таблица 4.3. Показатели интеллекта африканцев с островов Карибского бассейна и из Латинской Америки

Все сокращения, в том числе в других таблицах — см. Таблицу 4.1.
Афроамериканцы в Соединенных Штатах Америки
В Соединенных Штатах проводились сотни исследований интеллекта афроамериканцев. Результаты наиболее важных из них приведены в Таблице 4.4.
В строке 1 даны результаты первого из больших исследований — среди призывников в период Первой Мировой войны, проводившегося с помощью объединённых армейских Альфа и Бета тестов, которые измеряли невербальный и вербальный IQ и впоследствии легли в основу тестов Векслера. Число европеоидов составляло 93.973 чел. В строке 2 приведены данные для призывников Второй Мировой войны, а в строке 3 — результаты
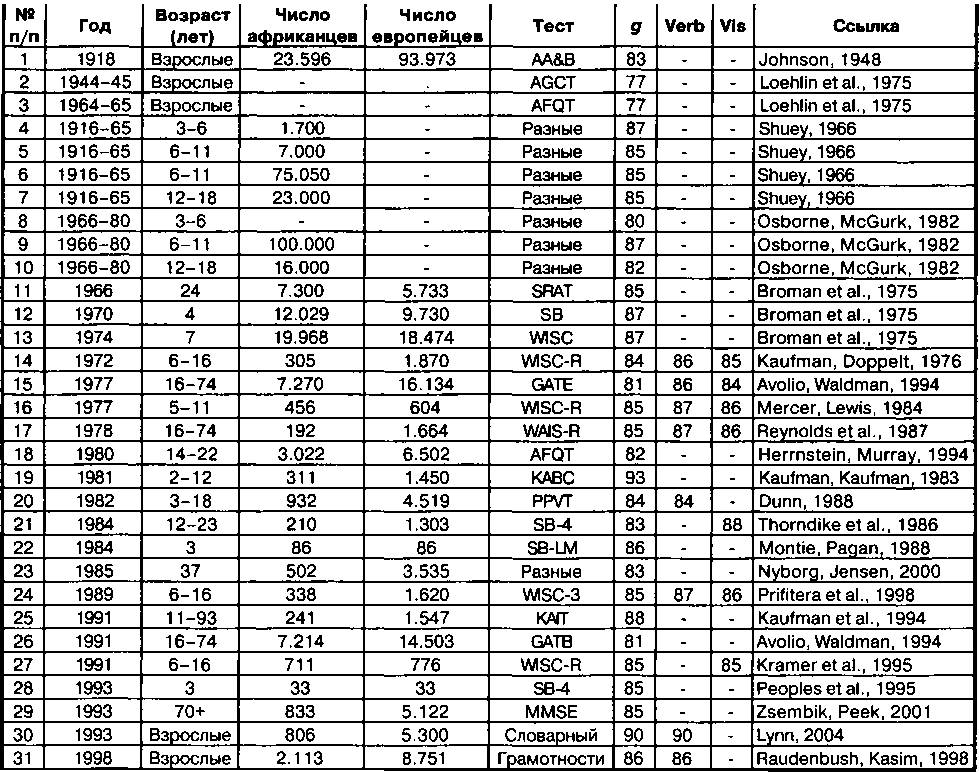
Расовые различия в интеллекте
59
тестирования призывников Вьетнамской войны. Примечательно, что средний IQ африканцев в периоды Второй Мировой и Вьетнамской войн был ниже (77 баллов), чем в период Первой Мировой войны, и ниже обычно приводимого среднего значения IQ афроамериканцев в Соединенных Штатах (85 баллов). В строках 4-7 приведены результаты компиляции всех американских исследований, выполненной Шу. В строке 4 дан IQ 87, полученный по данным 17 исследований среди дошкольников. В строке 5 представлен IQ 85, определённый по результатам 26 исследований среди учащихся начальной школы с использованием индивидуальных тестов, таких как тест Стенфорд-Бине. В строке 6 приведён IQ 85 для учащихся начальной школы, полученный по 103 исследованиям с применением тестов вербальной способности и 41 исследованию с помощью тестов невербальной способности. В строке 7 дан IQ 85 для студентов высшей школы. В строках 8,9 и 10 представлены результаты Осборна и Мак-Гурка (Osbome, McGurk, 1982), дополнивших суммарные данные американских исследований результатами работ, опубликованных в период 1976-1980 гг. В строке 8 дан IQ 80, полученный из результатов 66 исследований среди дошкольников 3-5 лет. В строке 9 приведён IQ 87, полученный по данным 126 исследований среди учащихся начальной школы, а в строке 10 — IQ 82, полученный по данным 17 исследований среди учащихся средней школы.
В строках 11,12 и 13 (Broman и др., 1975) даны результаты тестирования трёх больших выборок испытуемых, не представленных в обзоре Осборна и Мак-Гурка. В строке 11 приведён IQ в 85 баллов для африканских матерей, тестировавшихся в ходе выполнения Национального Коллабо- ративного Перинатального проекта, а в строках 12 и 13 — IQ их детей в возрасте 4 и 7 лет, составивший 87 баллов. В строке 14 представлен IQ общего интеллекта африканцев в 86 баллов по данным выборки теста стандартизации теста WISC-R. В строке 15 дан IQ в 81 балл для общего интеллекта, в 86 баллов для вербального интеллекта и в 84 балла для способности к визуализации среди работающих по найму (данные собраны Службой занятости Соединенных Штатов). В строке 16 дан IQ 85 для общего интеллекта афроамериканцев, 87 для вербального и 86 для способности к визуализации по выборке из Калифорнии. В строке 17 приведён IQ 85 для общего интеллекта, 87 для вербальной способности и 86 для способности к визуализации (по данным выборки стандартизации теста WAIS-R). В строке 18 представлен IQ 82, полученный с помощью теста AFQT. В строке 19 дан IQ 93 по данным выборки стандартизации теста К-АВС. В строке 20 даётся словарный IQ в 85 баллов по данным выборки стандартизации словарного теста Пибоди в картинках. В строке 21 представлен IQ в 83 балла, полученный по данным выборки стандартизации теста Стенфорд-Бине-4. Афроамериканцы из этой выборки получили IQ для кратковременной памяти в 89 баллов, что согласуется с результатами многих других исследователей, согласно
60
Ричард Линн
которым афроамериканцы сравнительно успешно выполняют тесты на кратковременную память. В строке 22 дан IQ 86 для 3-летних детей по данным выборки стандартизации теста Стенфорд-Бине-LM. В строке 23 приведён IQ 83, вычисленный из значений первого главного фактора как меры общего интеллекта, полученных у военнослужащих. В строке 24 представлен IQ 85 по данным выборки стандартизации теста WISC-3. В строке 25 дан IQ 88 по данным выборки стандартизации теста интеллекта Кауфмана для подростков и взрослых. В строке 26 дан IQ 81 для выборки работающих по найму (данные собраны Службой занятости Соединенных Штатов). В строке 27 приведён IQ 85 для способности к визуализации из оценок субтеста упорядочивания блоков теста WISC-R (по данным национальной выборки NHANES III).
Приведённый в строке 28IQ в 85 баллов получен у детей в возрасте от 3,0 до 3,4 лет поданным выборки стандартизации теста Стенфорд-Бине-4. В строке 29 дан IQ 85 для репрезентативной выборки испытуемых возраста 70 лет и старше из континентальных Соединенных Штатов (т. е., исключая Аляску и Гавайи). В строке 30 дан словарный IQ взрослых афроамериканцев в 90 баллов, полученный в исследованиях NORC за 1990-96 гг. на репрезентативной выборке из континентальных Соединенных Штатах. Это необычно высокое значение можно приписать краткости теста, состоящего в определении значений десяти слов. В строке 31 приведён IQ в 86 баллов по данным Национального исследования грамотности взрослых 1992 года, тест состоял в оценке понимания вербального материала и выполнении арифметических заданий испытуемыми репрезентативной выборки из континентальных Соединенных Штатов.
Из результатов исследований интеллекта афроамериканцев можно сделать пять выводов. Во-первых, медиана IQ составляет 85 баллов, это значение многими считается наилучшей оценкой IQ афроамериканцев. Эта оценка близка к значению в 83,5 балла, полученному в итоге мета-анализа данных 105 исследований, проведённых в общей сложности среди 6.246.729 испытуемых (Roth, Bevier, Bobko, Switzer, Tyler, 2001). Вариации средних значений, полученных в разных исследованиях, вызваны, вероятно, различиями выборок, ошибками измерения и различиями способностей испытуемых, измеряемых разными тестами. Во многих исследованиях было показано, что африканцы относительно хорошо справляются с тестами памяти, таким образом, величина африканско-европейского различия отражает в некоторой мере степень, в которой тесты памяти представлены в показателях интеллекта. Например, одно из наиболее высоких значений IQ в таблице (88 баллов) было получено по тесту KAIT Кауфмана. Этот тест содержит семь субтестов, из которых один оценивает память на лица, в нём требуется опознать лица известных людей. По этому субтесту африканцы получили средний IQ в 92,5 балла.
Расовые различия в интеллекте
61
Во-вторых, как можно видеть в строках 22 и 28, коэффициент интеллекта около 85 баллов наблюдается у 3-летних афроамериканцев. Эти результаты противоречат утверждению, часто высказываемому сторонниками средовой теории, что плохое образование и расизм полностью или в определённой степени ответственны за низкий IQ африканцев. Даже у 2-летних африканцев IQ составляет 93 балла (строка 19). Этот показатель не так низок, как в других исследованиях, потому что африканские младенцы созревают быстрее европейских до возраста двух лет (Lynn, 1998d; Rushton, 2000). Только на третьем году их IQ падает ниже европейского, и лишь на четвертом достигает величины порядка 85 баллов, как показано в строках 22 и 28.
В-третьих, характерный для афроамериканцев IQ около 85 баллов существенно выше среднего значения IQ африканцев в Африке южнее Сахары, составляющего примерно 67 баллов. Это различие можно объяснить двумя факторами. Прежде всего, афроамериканцы по многим параметрам живут в гораздо лучших условиях, чем африканцы в Африке, включая намного более высокий жизненный уровень, лучшее питание и здравоохранение. Кроме того, афроамериканцы имеют в среднем около 25 процентов европейских генов и это делает их IQ выше, чем у африканцев в Африке (Reed, 1969; Chakraborty, Kamboh, Nwanko and Ferrell, 1992).
В-четвертых, в пяти работах, где приводятся оценки вербального и визуального IQ, визуальный IQ афроамериканцев на один или два балла выше вербального. Результаты оценки вербального IQ более подвержены влиянию культурного окружения, таким образом, это противоречит утверждению, часто высказываемому сторонниками средовой теории, что африканцы плохо выполняют тесты потому, что тесты ориентированы не на них. Это подтверждает выводы Мак-Гурка (McGurk, 1953а, 1953b) о том, что афроамериканцы более не испытывают отрицательного влияния того, что считается «культурно-смещёнными общими информационными проблемами», и Дженсена (Jensen, 1980) о том, что тесты не являются культурно-смещёнными относительно афроамериканцев.
В-пятых, представляется, что в течение двадцатого столетия не происходило увеличения IQ афроамериканцев. Так, по данным четырнадцати исследований, выполнявшихся с 1980 г. по 1998 г., средний IQ составил 85 баллов, такое же значение определялось и в более ранних исследованиях. В пользу такого вывода говорит отсутствие какой-либо тенденции к уменьшению разрыва между афроамериканцами и европейцами в младших возрастных группах. Данные о различиях величин IQ у африканцев и европейцев разных возрастных групп были приведены в работе Рейнольдса и соавт. (Reynolds, Chastain, Kaufman, McLean, 1987) на материале выборки стандартизации теста WAIS-R 1978 г. Величины IQ африканцев составили 86 баллов у 16-19 летних, 85 баллов у 20-34 и у 35-54 летних, и 86
62
Ричард Линн
баллов у 55-74 летних испытуемых. Проводившиеся два раза в год тестирования показали также, что различие в интеллекте между афроамериканцами и европейцами не изменялось за период 1974—1996 гг. (Lynn, 1998е). Наконец, в выборке стандартизации теста KAIT (Тест интеллекта Кауфмана для подростков и взрослых; Kaufman и др., 1994) не было значимого различия между самой младшей и самой старшей возрастными группами. В самой младшей возрастной группе (родившиеся с 1980 по 1991 гг.) IQ был даже немного ниже (83 балла), чем в самой старшей (в среднем 1921 года рождения) — 88 баллов.
Африканцы в Великобритании
Африканцы начали мигрировать в Великобританию в значительном количестве вскоре после окончания Второй Мировой войны. Первые иммигранты приезжали в основном с островов Карибского бассейна, а в последней четверти двадцатого столетия значительное их число прибыло из Африки. Исследования IQ у африканцев в Великобритании публикуются с 1960-х гг. Результаты этих исследований приведены в Таблице 4.5.
В строке 1 приведён IQ в 88 баллов. По-видимому, это первая опубликованная работа по оценке IQ у детей негроидных иммигрантов из Вест- Индии. Исследование проводилось в Лондоне, где обосновалось большинство этих иммигрантов. В строке 2 дан IQ в 82 балла, вычисленный Верноном (Vernon, 1969, стр. 169) по другой выборке, тестировавшейся в Лондоне в 1960-х гг. В строке 3 приведён показатель способности к рассуждению в 88 баллов и словарный IQ в 82 балла для детей выходцев из Вест-Индии в сравнении с детьми англичан — европеоидов, посещавших одну и ту же среднюю школу в Лондонском районе Харингей. Это бедный район, где у европеоидных детей следует ожидать оценок IQ ниже среднего национального уровня, что будет повышать показатели выходцев из Вест-Индии. Чтобы скорректировать это смещение, IQ европейцев в этом исследовании был принят за 95. В строке 4 дан IQ 89 для группы детей из Лондона. В строке 5 представлен IQ 86 для выборок детей из Бирмингема и в Лондонского района Дептфорд.
В строке 6 приведён IQ в 104 балла для 9 африканских детей, принятых в учреждения опеки в младенческом возрасте из-за того, что их матери были неспособны заботиться о них. В этом же исследовании определялся IQ расово-смешанных и белых детей, также принятых в учреждения опеки; у расово-смешанных детей IQ составил 110 (п=15), а у белых 104 балла (п=36). Эти результаты не соответствуют всем остальным приведённым в таблице результатам, свидетельствующим о том, что у африканс-
Расовые различия в интеллекте
Таблица 4.5. Показатели интеллекта африканцев в Великобритании

ких детей в Великобритании показатели интеллекта значительно ниже, у белых. Кроме того, можно было бы ожидать, что IQ тестировавпп детей будет ниже среднего уровня, так как их матери, поместившие ] учреждения опеки, по преимуществу не владели необходимыми навык и, вероятно, обладали интеллектом ниже среднего. Данные об интелл* этих детей примечательны и нуждаются в дополнительной проверке. Е бы проверка их подтвердила, из них бы вытекало, что чернокожие мап не обеспечивают такие же хорошие условия детям, как белые приём
64
Ричард Линн
родители, воспитывавшие этих детей, но для такого вывода оснований немного. Число детей было очень небольшим (п=9), и возможно это просто случайный результат.
В строке 7 дан IQ в 86 баллов для общенациональной выборки афро- карибских детей в Великобритании. В строках 8 и 9 приведены значения IQ в 73 балла для детей из Великобритании, родившихся на Карибах, и в 82 балла для родившихся в Великобритании. Значение IQ в 73 балла у уроженцев островов Карибского бассейна близко к 71 — значению для аборигенных карибских детей. В строке 10 представлен вербальный IQ 86 для детей из Вест-Индии, тестировавшихся с помощью английского словарного теста в картинках. В строке 11 дан IQ 85 для детей выходцев из Вест- Индии в общеобразовательной средней школе города Ильфорд графства Эссекс; этот IQ ниже, чем у детей индусов, обучавшихся в той же школе, IQ которых составил 91 балл. Это первое в ряду исследований в Великобритании, продемонстрировавших, что у выходцев с островов Карибского бассейна показатели интеллекта ниже, чем у иммигрантов из Индии. В строке 12 дан словарный IQ в 78 баллов для всех детей выходцев из Вест- Индии, обучавшихся в общественных школах в Мидлендсе7, находящихся в ведении органов народного образования.
В строке 13 приведён IQ в 86 баллов, полученный с помощью теста чтения на очень большой национальной выборке (п= 12.530) 15-летних подростков. В строке 14 дан IQ в 85 баллов, полученный с помощью словарного теста у детей выходцев из Вест-Индии на Севере Англии в сравнении с 851 европейцами, посещавшими те же школы. В строке 15 представлен IQ в 86 баллов у детей выходцев из Вест-Индии из городской школы в Мидлендсе; дети выходцев из Индии, посещавшие те же школы, получили оценку IQ в 96 баллов. Это еще раз показывает, что выходцы из Южной Азии, находящиеся в одинаковых с африканцами условиях, получают более высокие оценки IQ. В строке 16 приведён IQ в 90 баллов для выборки детей выходцев из Вест-Индии в Лондоне. В строке 17 представлен IQ 87 для группы 4-летних детей выходцев из Вест-Индии.
В строке 18 приведён IQ в 89 баллов для национальной выборки карибских детей со всей Великобритании, родившихся в 1958 г. и проживших в Великобритании больше 4 лет; испытуемые другой группы (п=39), прожившие в Великобритании менее 4 лет, получили оценку IQ в 83 балла. Это даёт основание предполагать, что IQ карибских детей повышается вследствие проживания в Великобритании приблизительно на 6 баллов. Иногда высказывалось предположение, что вследствие того, что многие дети недавних иммигрантов с островов Карибского бассейна говорят на креольском Вест-Индийском диалекте английского языка, им трудно по
7 Так называют центральные графства Англии (прим. перев.)
Расовые различия в интеллекте
65
нимать учителей. Но то, что иммигранты из Вест-Индии выполняют тесты на невербальное рассуждение примерно на том же уровне, что и тесты на вербальное понимание, делает это предположение маловероятным. В строке 19 дан IQ в 92 балла для национальной британской выборки испытуемых, родившихся в 1970 г. Высокий IQ этой выборки мог бы быть индикатором того, что IQ детей выходцев с островов Карибского бассейна немного увеличился, но данные последующих работ, представленные в таблице, не показывают улучшения IQ африканских детей в период с 1960х по 1980-е гг., так что это может быть случайным результатом.
В строке 20 приведён IQ 88 для группы африканских школьников в лондонских школах, большинство из которых родилось в Великобритании. В строке 21 дан IQ в 92 балла для группы африканских школьников в Кембриджшире. Величина IQ дана относительно детей англичан, посещавших те же самые школы, у которых она, вероятно, была ниже национальных норм, так как британцы более высокого социально-экономического статуса предпочитают не посылать детей в школы, где учится заметное число иммигрантов. Это должно было повысить оценки интеллекта африканцев. Для применённого теста не существует национальных норм, поэтому IQ африканцев не мог быть точно расчитан относительно британцев. Вероятно, IQ британцев в этом исследовании составлял приблизительно 95 баллов, и, следовательно, IQ африканской группы составит около 87 баллов относительно британских национальных норм, то есть будет примерно таким же, что и в других выборках африканцев в Великобритании. В строке 22 приведён IQ в 89 баллов для группы 65-75-летних африканцев из Лондона, тестировавшихся в 1996-98 гг. в сравнении с национальной выборкой автохтонных британцев (п=5.379).
Результаты исследований интеллекта африканцев в Великобритании интересны в трёх отношениях. Во-первых, медианное значение IQ составляет 86 баллов, что почти точно совпадает со средним значением африканцев в Соединенных Штатах, равным 85 баллам. Эти оценки существенно выше медианных значений IQ в 67 баллов у африканцев в районах Африки южнее Сахары и в 71 балл у выходцев с островов Карибского бассейна, откуда большинство проживающих в Великобритании африканцев прибыло в период после окончания Второй Мировой войны. Во-вторых, более высокий IQ африканцев в Великобритании может быть объяснён влиянием лучших жизненных условий. Это влияние показано в исследовании, результаты которого представлены в строках 7 и 8 (Yule, Berger, Rutter, Yule, 1975). Видно, что IQ африканцев, родившихся в Вест-Индии, составляет 73 балла против 82 баллов у родившихся в Великобритании, что позволяет предположить, что IQ детей выходцев с островов Карибского бассейна увеличивается вследствие проживания в Великобритании на 9 баллов. Этот результат был подтвержден в работе Макинтоша и Масси-Тейлора
66
Ричард Линн
(Macintosh, Mascie-Taylor, 1985), результаты которой представлены в строке 18. В их исследовании IQ детей выходцев с островов Карибского бассейна, проживших в Великобритании более 4 лет, составил 89 баллов, в то время как в группе проживших Великобритании менее 4 лет (n=39) IQ был равен 83 баллам, из чего можно предположить, что интеллект карибских детей возрастает вследствие проживания в Великобритании примерно на 6 баллов. Эти два результата весьма сходны и говорят о том, что проживание в Великобритании увеличивает интеллект африканцев на 7-8 баллов. Это увеличение, видимо, является главным образом результатом лучшего питания и здравоохранения и, возможно, образования. Хотя нет никаких свидетельств тому, что образование в Вест-Индии хуже, чем в Великобритании; иногда утверждается что оно лучше. Влияние улучшеного питания на иммигрантов из Вест-Индии показано Юлом с соавторами (Yule и др.,
, определившими, что африканцы из Вест-Индии, родившиеся в Великобритании, выше ростом родившихся на островах Карибского бассейна и приехавших в Великобританию детьми; различие составило 0.67d(единицы стандартного отклонения). В-третьих, величина IQ для выборки 4-летних детей выходцев из Вест-Индии, приведённая в строке 17, была почти такой же (87 баллов), что и у более старших детей выходцев из Вест- Индии, обучавшихся в школе. Это говорит о том, что за низкие показатели интеллекта детей выходцев из Вест-Индии не могут нести ответственности школы, предубеждение учителей, трудности понимая детьми разговорного английского языка учителей и так далее. Этот результат подтверждают полученные в Соединенных Штатах данные о том, что IQ африканцев низок в дошкольном возрасте.
Африканцы в Нидерландах
В течение второй половины двадцатого столетия значительное число африканцев мигрировало в Нидерланды из бывшей голландской колонии Суринам на Северо-Востоке Южной Америки и с Нидерландских Антильских островов, бывшей голландской колонии в Карибском море. Результаты исследований их интеллекта представлены в Таблице 4.6.
В строке 1 представлен IQ в 86 баллов для детей иммигрантов из Суринама. Данные об использованном тесте и возрасте испытуемых не приведены. Население Суринама состоит на 35 процентов из креолов с африканско-европейской родословной, на 10 процентов из африканцев, на 33 процента из выходцев из Индии, на 16 процентов из индонезийцев и на 3 процента из американских индейцев. Величина IQ в 86 баллов — это примерно то, что можно было бы предсказать для этого расово-смешанного населения, так как у наи-
Расовые различия в интеллекте
67
Таблица 4.6. Показатели интеллекта африканцев в Нидерландах

большей группы, креолов, IQ будет примерно средним между IQ африканцев в Африке (67) и европейцев в Северо-Западной Европе (100), а у второй по численности группы, индусов, IQ должен был бы составить примерно 82 (см. Главу 6). В строке 2 дан IQ в 84 балла для выборки детей иммигрантов первого поколения из Суринама и с Нидерландских Антильских островов. В строке 3 приведён IQ 88 для выборки детей иммигрантов второго поколения из Суринама и с Нидерландских Антильских островов. У этих детей наблюдается увеличение IQ на четыре балла в сравнении со значением для детей иммигрантов первого поколения, представленным в строке 2. Это подтверждает исследования в Великобритании, показавшие, что иммигранты второго поколения получают более высокие оценки IQ, чем иммигранты первого поколения. Строка 4 дает IQ 85 для ещё одной выборки детей иммигрантов из Суринама. Данные об использованном тесте и возрасте испытуемых не приведены. В строке 5 дан IQ 83 для другой выборки иммигрантов из Суринама и с Нидерландских Антильских островов. В строке 6 приведён IQ 85 для взрослых иммигрантов из Суринама. В строке 7 представлен IQ 85 для иммигрантов с Нидерландских Антильских островов, население которых состоит на 85 процентов из африканцев и лиц со смешанной африканско-европейский родословной. Величины IQ, полученные в приведённых исследованиях, весьма сходны. Медианное значение составляет 85 баллов — такое же, как у африканцев в Соединенных Штатах.
Африканцы в Израиле
Некоторое количество эфиопских иудеев-африканцев мигрировало в Израиль в течение последних десятилетий двадцатого века. Каниель и Фишерман (Kaniel, Fisherman, 1991) оценили интеллект 15-летних испытуемых (п=250) с использованием стандартных прогрессивных матриц. Авторы пишут:
68
Ричард Линн
«Эфиопские иудеи были протестированы спустя год после прибытия в Израиль. Все испытуемые были протестированы в группах в своих школах с использованием стандартной процедуры. Каждой группе демонстрировали тренировочный первый пункт теста и выполнили его вместе с ними. Особое внимание уделяли тому, чтобы удостовериться в том, что эфиопские иудеи поняли, как организован тест, и в их способности заполнить бланк ответов. Никаких временных ограничений не было» (стр. 28).
Авторы сделали ошибки при вычислении коэффициента интеллекта эфиопов. В моей работе представлены правильные значения (Lynn, 1994b). Средняя оценка в тесте составила 27, что эквивалентно первому процен- тилю по нормам британской стандартизации 1979 г. и значению IQ в 65 балов. Израильские данные были получены в 1989 г., a IQ британцев, как предполагается, за период с 1979 по 1989 г. возрос на 2 балла. Чтобы учесть это, необходимо ввести поправку на это увеличение, соответственно IQ эфиопских иудеев должен быть уменьшен до 63.
Второе исследование показателя интеллекта эфиопских иудеев было опубликовано Козулином (Kozulin, 1998). Оно было проведено среди 14— 16-летних испытуемых, проживших в Израиле четыре или более лет и посещавших израильские школы-интернаты, для тестирования использовались прогрессивные матрицы. Средний IQ этих испытуемых составил 65 баллов. Эти результаты говорят о том, что образование в школах западного типа не способствует повышению IQ африканцев.
Кратковременная память и скорость восприятия у африканцев
До настоящего времени мы рассматривали интеллект африканцев в терминах фактора g (общий интеллект), способности к рассуждению, вербальных и визуализационных способностей. Рассмотрим теперь исследования кратковременной памяти и скорости восприятия африканцев. Кратковременная память обычно измеряется тестом повторения цифровых рядов (Digit Span), заключающемся в оценке способности запомнить и воспроизвести набор цифр либо в том порядке, в котором они были представлены (прямой Digit Span) или в обратном порядке (обратный Digit Span). Измерение скорости восприятия обычно проводится субтестами кодирования (Coding) и замещения символов цифрами (Digit Symbol) тестов Векслера, которые требуют точного и быстрого просмотра визуальной информации. Эти исследования показали, что у африканцев достаточно хорошие кратковременная память и скорость восприятия. Результаты работ приведены Таблице 4.7.
В строке 1 представлены величины IQ в 75 и 76 баллов для африканских 10-12-летних детей (п=1.123) в сравнении с европейскими (п=1.489) для
Расовые различия в интеллекте
69
Таблица 4.7. Первичные способности африканцев

Сокращения: AR — абстрактное рассуждение; VR — вербальное рассуждение; Verb — вербальное понимание; Vis — визуализация; STM — кратковременная память; PS — скорость восприятия.
невербального рассуждения и вербальной способности, измеренных с помощью теста Лорж-Торндайка; существенно более высокие оценки были получены для кратковременной памяти (IQ=90, тест Digit Span) и, особенно, для скорости восприятия (IQ=102). Авторы пишут по этому поводу: «При предъявлении теста, оценивающего только скорость, но не содержащего ощутимой когнитивной задачи, негритянские дети выполняют его так же хорошо или даже лучше, чем европейские» (Jensen, Rohwer, 1970, стр. 60). В строке 2 представлен типичный IQ в 85 балла для вербальной шкалы и шкалы действия теста WISC-R, предъявлявшегося 622 африканским детям 5-11-летнего возраста в сравнении с 669 европейскими, а также IQ кратковременной памяти в 94 балла как среднее значение оценок тестов прямой Digit Span (IQ=96) и обратный Digit Span (IQ=92). В строке 3 приведены оценки IQ африканских детей 12-18 лет, полученные из корпуса данных проекта «Талант»; согласно им IQ для кратковременной памяти (94 балла) является относительно высоким сравнительно с IQ для абстрактного рассуждения (89 баллов). В строке 4 дан IQ 94 для кратковременной памяти 5-9-летних детей африканских иммигрантов из Суринама в Нидерландах (п=183) в сравнении с европейскими детьми; тест состоял в предъявления десяти рисунков, каждому из которых было дано произвольное название, задача состояла в запоминании как можно большего числа названий. В строке 5 приведены оценки IQ африканских детей возраста 6-16 лет (п=711) относительно европейских детей (п=776) в 82 балла для вербальной способности, в 78 баллов для способности к визуализации и более высокое значение в 90 баллов для кратковременной памяти, измеренное тестом Digit Span. В строке 6 дан IQ в 94 балла для кратковременной памяти африканцев, полученный как результат мета-анализа
70
Ричард Линн
данных 31 исследования детей и взрослых. В строке 7 приведены оценки первичных способностей испытуемых из Южной Африки. Авторы работы протестировали 1.093 африканца и 1.056 европеоидов 16-летнего возраста с помощью юношеского теста способностей, разработанного в Южной Африке и дающего оценки абстрактного рассуждения (AR), вербального рассуждения (VR), вербального понимания (Verb), визуализации (Vis), кратковременной памяти (STM) и скорости восприятия (PS); у испытуемых были оценены также механические способности, оценка составила 68 баллов. Как можно видеть, IQ кратковременной памяти африканцев (79 баллов) и скорости восприятия (69 баллов) выше способности к абстрактному рассуждению (58 балов) и вербальному рассуждению (63 балла), что подтверждает данные американских исследований. В этой выборке визуализация и механические способности также были более выражены, чем способность к абстрактному и вербальному рассуждению. В строке 8 дан IQ кратковременной памяти в 79 баллов для выборки из 196 10-летних детей на Ямайке в сравнении с оценкой в 67 баллов — медианой результатов семи исследований, приведённых в Таблице 4.3.
Дженсен (Jensen, 1998) интерпретировал эти и другие результаты как указывающие на то, что африканско-европейские различия в интеллекте являются в значительной степени различиями в факторе gСпирмена. Согласно этой теории, кратковременная память и скорость восприятия являются слабыми оценками фактора g, поэтому африканцы сравнительно успешны в таких тестах. Теория получила немало подтверждений, их обзор представлен в работе Раштона (Rushton, 2003), но также вызвала и некоторую критику со стороны Долана и Хамакера (Dolan, Hamaker, 2001).
Музыкальные способности
Часто высказывалось предположение о том, что у африканцев хорошие музыкальные способности и особенно выражено чувство ритма. Кажется, это было впервые сказано в четырнадцатом веке арабским автором Ибн Бутланом, который писал, что если бы африканцу «привелось упасть с неба на землю, он отбивал бы такт, падая вниз» (Lewis, 1990, стр. 94). Музыкальные способности связаны с интеллектом, так что интересно посмотреть, имеют ли африканцы хорошие музыкальные способности, часто им приписываемые, или же у них слабые музыкальные способности в соответствии с низкими оценками IQ, получаемыми ими в тестах интеллекта.
Музыкальные способности измеряются простыми тестами, такими как распознавание изменения высоты тона (нужно определить, выше или ниже один тон другого; в первых пробах различие между тонами велико, но по
Расовые различия в интеллекте
71
степенно тоны становятся ближе друг к другу, пока не станет крайне сложно определить, который выше); музыкальная память (мелодия играется дважды и при втором исполнении одна нота изменяется; задача состоит в том, чтобы определить, какая нота изменилась); анализ аккорда (определение количества нот в аккорде) и чувство ритма (играются два музыкальных произведения и нужно определить, в одном или в разных ритмах). Связь между интеллектом и музыкальными способностями была показана в двух исследованиях, выполненных Линном, Вильсоном и Голтом (Lynn, Wilson, Gault, 1986). В первом исследовании группе 10-летних детей (п=217) представляли ряд тестов на рассуждение, словарные способности, визуализацию и скорость восприятия вместе с четырьмя тестами музыкальных способностей (высота тона, музыкальная память, анализ аккорда и чувство ритма). Все тесты были положительно интеркоррелированы и имели нагрузки по первой главной компоненте как мере общего интеллекта (фактор g).Нагрузки четырех музыкальных тестов лежали в пределах от 0,45 (аккорды) до 0,59 (ритм). Это показывает, что музыкальные тесты являются мерой фактора g. Во втором исследовании 93 испытуемым 9-11-летнего возраста предлагали три теста музыкальных способностей (изменение тона, анализ аккорда и музыкальная память) вместе со стандартными прогрессивными матрицами, как мерой фактора g.Три музыкальных теста имели значимые корреляции с прогрессивными матрицами: 0,27; 0,40 и 0,37, соответственно. Это подтверждает, что музыкальные способности связаны с интеллектом. Новые свидетельства этой связи были представлены Кэрролом (Carroll, 1993).
Таблица 4.8. Музыкальный (MQ) и ритмовый (RQ) коэффициенты афроамериканцев

Исследования музыкальных способностей афроамериканцев проводились, но они малоизвестны, так как не были обобщены в базовых учебниках по интеллекту, например у Броди (Brody, 1992) или Макинтоша (Mackintosh, 1998) или же в специальных учебниках по расовым различиям в интеллекте, таких как Лоелина, Линдзи и Спулера (Loehlin, Lindzey, Spuhler, 1975) или Дженсена (Jensen, 1980, 1998). Общий вывод из этих исследований: афроамериканцы хуже европейцев выполняют тесты музыкальных способностей на различение высоты, различение тона и музыкальную
72
Ричард Линн
память, но получают примерно одинаковые с европейцами оценки в тестах чувства ритма. Чтобы выявить эту структуру музыкальных способностей, результаты исследований были сгруппированы так, чтобы получить музыкальный коэффициент (MQ), выведенный из тестов музыкальных способностей за исключением чувства ритма и ритмовый коэффициент (RQ). Результаты этих исследований приведены в Таблице 4.8.
В строке 1 приведены результаты тестирования большой группы афроамериканцев в Северной Каролине, Южной Каролине и Виржинии с помощью теста Сишора. По данным тестирования был вычислен MQ (музыкальный коэффициент) в 90 баллов и более высокий RQ (ритмовый коэффициент) в 106 баллов. В строке 2 представлены результаты сравнения 272 европейцев и 288 афроамериканцев, обучавшихся в колледжах штата Теннеси, они вновь показывают, что европейцы получают более высокие оценки в тестах общих музыкальных способностей (высота тон, интенсивность, время звучания, гармония (iconsonance) и общая музыкальная память), но афроамериканцы получили более высокую оценку RQ (ритмовый коэффициент) —102 балла. В строке 3 представлены результаты обследования выборки из бедного пригорода Вашингтона, округ Колумбия, выявившего MQ 83 и RQ 96 баллов. В строке 4 приведены результаты для группы афроамериканцев возраста 5-8 лет из Рочестера в штате Нью-Йорк: MQ в 89 и RQ в 104 балла. В строке 5 даны результаты тестирования выборки 18-летних афроамериканцев (старшеклассников средней школы в основном из Техаса, но некоторые были из Иллинойса и Рочестера штата Нью-Йорк), получен MQ в 86 (по тестам тональной памяти и различения высоты тона) и RQ в 100 баллов. Группой сравнения были европеоиды (п=541), посещавшие те же школы. Все исследования показывают, что у афроамериканцев ритмовый IQ существенно выше общего музыкального IQ — примерно на 15 баллов. По-видимому, музыкальные способности африканцев не изменились примерно за полувековой период с 1920-х до конца 1970-х гг., в который проводились исследования. Сравнительно хорошее чувство ритма африканцев выражено в их музыке, в которой часто присутствует сильный ритмический элемент. Это особенно заметно в гимнах, исполняемых конгрегациями в африканских и афроамериканских церквях. Также это проявляется в джазе, который первоначально был создан афроамериканцами в Новом Орлеане в первые годы двадцатого столетия, и в развившемся из него впоследствии направлении «свинг», с его сильно синкопированными ритмами.
Несколько близнецовых исследований показали, что музыкальные способности имеют генетическую основу. Например, Ванденберг (Vandenberg. 1962), изучавший наследуемость «ритмовой способности», оцененной по корреляциям 33 пар однояйцовых и 43 пар разнояйцовых близнецов одного пола, вычислил уровень наследуемости в 0,52 (без поправки на ошибку измерения). При такой величине наследуемости представляется вероятным,
Расовые различия в интеллекте
73
что слабые музыкальные способности и высокая ритмовая способность африканцев имеют некоторое генетическое основание.
Слабые музыкальные способности африканцев, за исключением их выраженного чувства ритма, согласуются с их в целом скромными достижениями в классической музыке. Нет африканцев — композиторов, дирижеров или музыкантов первого ранга, нечасто можно увидеть африканских исполнителей и в ведущих симфонических оркестрах.
Время реакции
Время реакции представляет собой скорость реакции на простой стимул, такой, как вспышка света. Задача состоит в том, чтобы нажать кнопку в тот момент, когда это происходит. Время реакции — время, потраченное на ответ, оно составляет обычно около трети секунды. Результаты большого количества исследований, представленных в монографиях Дженсена (Jensen, 1998) и Дири (Deary, 2000), показали, что время реакции положительно связано с интеллектом, значение корреляции составляет от 0,2 до 0,3. Как убедительно аргументировал Дженсен (Jensen, 1998), время реакции является мерой неврологической эффективности обработки информации мозгом. Это побуждает нас задаться вопросом, не сопровождаются ли различия интеллекта между европейцами и африканцами также и различиями во времени реакции. Если да, то это означает, что расовые различия в эффективности деятельности мозга существуют. Если нет, это означает, что расовые различия в эффективности работы мозга отсутствуют и различия в интеллекте обусловлены другими факторами, такими как разница в возможностях научения решению тестовых задач, образовательный опыт или смещённость тестов.
Наиболее исчерпывающие исследования африканско-европейских различий во времени реакции были выполнены Дженсеном (Jensen, 1993) в Соединенных Штатах, а также Линном и Холмшоу (Lynn, Holmshaw, 1990) и Сонке (Sonke, 2000) в Южной Африке. Дженсен сравнивал 585 европейцев и 235 африканцев 10-летнего возраста, коэффициенты интеллекта которых по оценке с использованием прогрессивных матриц различались на 11 баллов IQ. В исследовании Линна и Холмшоу сравнивались 350 африканцев и 239 британцев 9-летнего возраста, интеллект которых различался на 37 баллов IQ. В обоих исследованиях был использован один и тот же управляемый компьютером аппарат, так что «человеческий фактор» не мог повлиять на результат измерения. В обоих исследованиях измерялись двенадцать компонентов времени реакции. Было измерено время трёх видов реакции: время простой реакции (SRT), представляющее
74
Ричард Линн
собой реакцию на единственную вспышку света, время реакции различения (CRT), когда надо реагировать на одну из восьми возможных вспышек, и время реакции на «третьего лишнего» (OMRT), когда надо реагировать на наиболее отстоящую от двух других вспышку. В каждой из этих трёх реакций фиксировались четыре показателя: собственно время реакции (время принятия решения), время движения (время, потраченное на нажатие кнопки) и стандартные отклонения (SD) времени реакции и движения. Полученные результаты приведены в Таблице 4.9.
Таблица 4.9. Корреляции между временем реакции и IQ, и различия во времени реакции между африканцами и европейцами

В колонке 1 указаны различные меры времени реакции. В колонках 2 и 3 приведены данные Дженсена по корреляции времени реакции с величиной IQ, оцененной по прогрессивным матрицам и различия между африканцами и европейцами (в d— единицах стандартного отклонения); отрицательные знаки означают разницу «в пользу» европейцев. В колонках 4 и 5 даны такие же показатели из работы Линна и Холмшоу. Корреляции между временем реакции и IQ устойчиво положительны во всех случаях, 16 из 24 корреляций статистически значимы (отмечены звездочками), но коэффициенты корреляции очень низки. Время реакции, приведённое в строках 1, 5 и 9, было меньше у европейцев, чем у африканцев, за исключением CRT в работе Дженсена. Время простого движения не различа
Расовые различия в интеллекте
75
лось, но по данным Линна и Холмшоу африканцы были значительно быстрее европейцев как по СМТ, так и по ОММТ. Более быстрые движения африканцев могут быть фактором, определяющим их высокую спринтерскую скорость, засвидетельствованную олимпийскими рекордами. Стандартные отклонения были устойчиво больше у африканцев по данным Линна и Холмшоу, и в четырех из шести случаев по данным Дженсена. В целом, африканско-европейские различия намного больше и более устойчивы по данным Линна и Холмшоу, чем по данным Дженсена. Этого можно было ожидать ввиду того, что в исследовании Линна и Холмшоу разница в интеллекте между сопоставляемыми группами была приблизительно в четыре раза больше. Однако, согласно данным Линна и Холмшоу среднее арифметическое различий шести показателей времени реакции и стандартных отклонений между африканцами и европейцами составляет только 0,67dв сравнении с различием в IQ 2,5d.Наилучшее объяснение этих результатов состоит в том, что приблизительно четверть африканско-европейского различия в интеллекте может быть объяснена скоростью неврологической обработки, в то время как остальная часть должна быть приписана другим процессам.
Хотя наследуемость времени реакции составляет около 50 процентов (Deary, 2000), на неё также влияет питание. В одном итальянском исследовании было показано, что у детей возраста 6-10 лет из йодо-дефицитных местностей было замедлено время реакции и снижены показатели IQ (Vitti и др., 1992). О замедленном времени реакции у детей из йодо-дефицитных местностей также сообщалось Бличродтом и соавторами (Bleichrodt и др., 1987).
Сонке (Sonke, 2000) опубликовал результаты обследования трёх групп: 26 неграмотных африканцев из Южной Африки 16-летнего возраста со «всего лишь несколькими классами образования», 29 африканских студентов из университета Венда в Северном Трансваале и 30 европейских студентов — голландцев из университета Тилберга. В этих трёх группах оценивали интеллект (по прогрессивным матрицам Равена) и измеряли время простой и сложной реакции, фиксировали также длительность латентного периода вызванного потенциала (РЗ) на ЭЭГ в ответ на представленный стимул, как меру скорости регистрации стимула мозгом. Количество мужчин и женщин было одинаковым во всех трёх группах. Полученные результаты приведены в Таблице 4.10.
Таблица 4.10. Времена реакции и ЭЭГ африканцев и европейцев

76
Ричард Линн
В строке 1 приведены коэффициенты интеллекта трех групп. В строке 2 дано среднее время простой реакции (RT-S), самыми «медленными» были неграмотные африканцы, самыми «быстрыми» — европейские студенты. В строке 3 представлено время сложной реакции (RT-C), различия между группами аналогичны. В строке 4 приведён латентный период вызванного потенциала для задачи В1 (EEG), самый длительный латентный период был у неграмотных африканцев, самый короткий — у европейских студентов. Все различия между группами были статистически значимыми.
В этом исследовании имеются шесть интересных моментов. Во-первых, IQ неграмотных африканцев по результатам тестирования с помощью прогрессивных матриц составил 68 баллов, что очень близко к значениям для большого количества выборок африканцев из Южной Африки и других стран в районах южнее Сахары. Во-вторых, у африканцев — студентов университетов IQ был несколько выше (82 балла), что также сходно с IQ других выборок африканских студентов университетов Южной Африки. В-третьих, выявлены значимые различия между группами во времени реакции, что подтверждает результаты других исследований, представленные в этой главе. В-четвертых, выявлены существенные африканско-европейские различия вызванных потенциалов ЭЭГ, указывающие на то, что у европейских студентов мозг реагирует на стимул быстрее, чем у африканских. В-пятых, имеется статистически значимая корреляция (0,213) между временем сложной реакции и результатом по прогрессивным матрицам, подтверждающая результаты многих других исследований этой связи. И, в-шестых, корреляция междурезультатом по прогрессивным матрицам и вызванными потенциалами ЭЭГ не является статистически значимой. Различия между неграмотными африканцами и африканскими студентами во времени реакции и вызванных потенциалах следует, вероятно, отнести на счёт более высокого IQ студентов.
Размер мозга африканцев и европейцев
Исследования, показывающие, что у африканцев средний размер мозга меньше, чем у европейцев, представлены в Таблице 4.11.
Размеры мозга в таблице приведены в кубических сантиметрах (в эти единицы были пересчитаны кубические дюймы, в которых представляли данные Мортон и Гулд, и граммы, в которых проводил измерения Хо (Но и др., 1980)). Нужно отметить, что оценки объёма черепа до некоторой степени зависят от метода измерения. Объём черепа измеряют, наполняя его свинцовой дробью или семенем горчицы и измеряя объём дроби или семени. Свинцовая дробь даёт немного больший объем, чем семя горчи
Расовые различия в интеллекте
77
цы, так как не может быть сжата так же плотно. У живых людей объём мозга вычисляют по длине, ширине и высоте головы или по её окружности. Использование разных методов измерения объясняет некоторые из различий, полученных в исследованиях. Несмотря на использование разных методов измерения, результаты различных исследований в значительной степени согласуются. В Таблице 4.11 приведены результаты восьми исследований размера мозга в выборках европейцев и африканцев и различия между средними значениями.
Таблица 4.11. Размер мозга (см3) у европейцев и африканцев

Все исследования показывают, что у европейцев средний размер мозга больше, чем у африканцев. В строке 1 приведены результаты первого исследования, выявившего различия в размере мозга у африканцев и европейцев, опубликованного в девятнадцатом столетии американским врачом Сэмюэлем Мортоном (Morton, 1849), собравшим коллекцию черепов, распределившим их по расам и вычислившем их средние объёмы. Его результаты критиковались Гулдом (Gould, 1996), обвинившем его в преувеличении значений с целью показать, что у европейцев мозг больше, чем у африканцев. Гулд повторно вычислил размеры черепов коллекции Мортона и получил очень близкие результаты. В таблице приведены значения, полученные Гулдом. Он отклонил различие в 41см3 как незначительное. Характерно, что Гулд не сослался ни на одно из других исследований, которые все подтвердили выводы Мортона и в которых были выявлены ещё большие различия. Отметим, что количество черепов в коллекции Мортона (52 черепа европейцев и 29 черепов африканцев) было весьма небольшим в сравнении с другими исследованиями.
78
Ричард Линн
В строке 2 даны результаты исследования намного большей коллекции черепов, хранящихся в Университете Западного Резерва в Огайо, различие в размере мозга между африканцами и европейцами составило 50см3. В строке 3 приведены результаты, представленные ревностным сторонником эгалитаризма Тобиасом, утверждающим, что расовых различий в размере мозга нет, хотя в действительности его результаты демонстрируют ещё большее африканско-европейское различие, чем у Мортона. В строке 4 представлены данные, полученные при вскрытии трупов в Соединенных Штатах, показавшие большее, чем в других исследованиях, африканско- европейское различие — 103см3. В строке 5 приведены результаты измерений наибольшей коллекции черепов, примерно 20 тысяч со всех континентов, выполненных американским антропологом Кеннетом Билсом. В строке 6 представлены результаты расчётов, выполненных Гровсом (Groves, 1991) на основе объединения оценок объёма мозга 36 выборок мужчин из данных Куна, Молнара и Мартина, а также Сэллера. Эти результаты вновь показывают, что у европейцев средний объём мозга больше, чем у африканцев. В строке 7 приведены результаты для военнослужащих в Соединенных Штатах, скорректированнные на рост и вес. Размер мозга европейцев был практически идентичен определённому Хо и соавт. (строка 4), но размер мозга африканцев был существенно больше: 1.359 см3 по сравнению с 1.267 см3. Объяснение этому состоит в том, что американские военные власти оценивают интеллект поступающих на военную службу и отклоняют тех, у кого IQ ниже 81 (Nyborg и Jensen, 2000). Флинн определил (Flynn, 1980), что доля не принятых на военную службу из-за низкого IQ составляет 3,4 процента для европейцев и 30 процентов для африканцев, в результате этого средний IQ африканцев в вооруженных силах США составляет 91,5. В результате отказа в приёме на военную службу африканцам с низким IQ происходит отсев тех из них, кто обладает низким интеллектом и малым объёмом мозга, что делает африканско-европейское различие размера мозга в этой выборке намного меньше, чем в других. В строке 8 приводится средний размер мозга для шести выборок европейцев мужского пола из Северной Америки и Европы и двух выборок из районов Африки южнее Сахары; данные компилированы Юргенсом, Ауне и Пипером (Jurgens, Aune, Pieper, 1990) и проанализированы Раштоном (Rushton, 2000, стр. 124), выявлено преимущество европейцев в 109см3. Все восемь наборов данных свидетельствуют о том, что у европейцев размер мозга больше, чем у африканцев. Результаты является достаточно согласующимися, принимая во внимание то, что они были собраны с использованием разных методов и на различных видах выборок, включая паталогоанатомические исследования (Но и др., 1980), определение объема черепа (Reals и др., 1984) или внешние обмеры головы живых людей.
Расовые различия в интеллекте
79
Эти результаты были подкреплены другим крупномасштабным исследованием, выполненным Броманом и соавторами (Broman и др., 1987) среди детей. Они наблюдали и обследовали приблизительно 17 тыс. европейских и 19 тыс. африканских детей в Соединенных Штатах в период от зачатия до возраста 7 лет. К 7 годам типичный разрыв в интеллекте двух групп составлял приблизительно 15 баллов IQ. В этом возрасте окружность головы, рассчитанная на основе опубликованных данных, составляла 50,9 см (станд. откл. 1,6) у африканцев и 51,7 см (станд. откл. 1,6) у европейцев. Это различие статистически высоко значимо и даёт приближённую меру различий в размере мозга, так как корреляция окружности головы и размера мозга составляет около 0,8 (Brandt, 1978). Объём мозга был оценен Раштоном (Rushton, 1997) в 1.134 см3 у африканцев и 1.150 см3 у европейцев. Это различие намного меньше, чем в других выборках вероятно потому, что европейцы созревают позже, чем африканцы. Наблюдавшиеся в этом исследовании африканские дети были немного более рослыми, чем европейские, что даёт основание предположить, что возможные различия в питании вряд ли ответственны за различия в размере головы.
Показатели интеллекта африканско-европейских гибридов
Теперь мы рассмотрим исследования интеллекта африканско-европейских гибридов. Генетическая теория расовых различий предсказывает, что величина IQ межрасовых гибридов должна попасть примерно на середину интервала между европейцами и африканцами. Чтобы проверить это предсказание, мы суммировали результаты исследований африканско-европейских расовых гибридов в Таблице 4.12.
В строке 1 приведены полученные в Бразилии результаты, показывающие, что гибриды, известные как «коричневые», имеют промежуточные оценки между европейцами и африканцами. В строке 2 представлены результаты исследования в Германии (Eyferth, 1961), показавшего, что IQ африканско-европейских гибридных детей составлял 94 балла относительно 100 у европейских детей. Средний IQ африканско-европейских гибридов составил 96,5, но он был уменьшен в таблице до 94, чтобы учесть долговременное увеличение IQ со времени стандартизации. В строке 3 приведены данные из Южной Африки для европейцев, африканцев и цветных, являющихся в значительной степени африкано-европейскими гибридами. Величина IQ цветных (83 балла) оказалась точно посредине интервала между европейцами (100 балла) и африканцами (65 баллов). В строке 4 приведены результаты более свежего исследования в Южной Африке,
80
Ричард Линн
Таблица 4.12. Показатели интеллекта европейцев, африканско-европейских гибридов и африканцев

проведённого примерно шестьдесят лет спустя и выявившего в выборке цветных IQ в 86 баллов в сравнении со 100 баллами европейцев. Африканцы не были включены в это исследование, но IQ цветных в 86 баллов был намного выше, чем у чистокровных африканцев в Южной Африке. В строке 5 приведены результаты последующего исследования в Южной Африке, давшего оценку IQ цветных в 80 баллов.
В строках 6-9 даны четыре результата для гибридов в Соединенных Штатах Америки. В строке 6 приведены результаты Миннесотского исследования межрасового усыновления; полученная оценка IQ гибридов находится посредине между оценками афроамериканцев и европейцев. Численности выборок очень низки, но результаты информативны, потому что все три группы воспитывались европейскими приёмными родителями, что исключает любую разумную средовую интерпретацию различий. В строке 7 представлены другие данные из Соединенных Штатов, еще раз указывающие на то, что гибриды имеют промежуточные оценки IQ между европейцами и афроамериканцами (результаты этого исследования были представлены Лоэлином, Линдзи и Спулером — Loehlin, Lindzey, Spuhler, 1975). В строке 8 даны результаты ещё одной работы из Соединенных Штатов. Её авторы разделяли афроамериканцев на темнокожих и менее темнокожих и показали, что обладающие относительно светлой кожей афроамериканцы, что принимали за показатель гибридизации с европейцами, имели IQ 92 — среднее значение между европейцами и африканцами. Представленный в строке 9 последний результат показывает, что гибриды афроамериканцев с европейцами имеют IQ 97 и вновь оказываются в промежутке между афроамериканцами и европейцами.
Расовые различия в интеллекте
81
Наследуемость интеллекта у афроамериканцев
Было проведено три исследования наследуемости интеллекта афроамериканцев в Соединенных Штатах. Все они были выполнены посредством сравнения монозиготных (Mz) и дизиготных близнецов (Dz). Результаты представлены в Таблице 4.13.
В таблице приведены возраст испытуемых, количество монозиготных (Mz) и дизиготных (Dz) близнецов, корреляции между парами близнецов, наследуемость, полученная путём удвоения разницы между корреляциями Mz и Dz, и исправленная наследуемость, скорректированная на уменьшение показателя наследуемости в силу не стопроцентной надёжности теста, принимая надёжность теста в 0,9.
Таблица 4.13. Наследуемость интеллекта у афроамериканцев

В строке 1 представлены данные Лоэлина, Линдзи и Спулера (Loehlin, Lindzey, Spuhler, 1975), полученные из докторской диссертации П. Л. Николса для 4-летних испытуемых, обследованных с помощью теста Стенфорд-Бине. Они дают исправленную наследуемость в 0,56—немного выше, чем у европейцев того же возраста. В строке 2 приведены данные из работы Осборна (Osborne, 1980, стр. 72) для общего интеллекта, вычисленного как среднее значение результатов двенадцати тестов; согласно им величина исправленной наследуемости оказалась равной 1,00. В строке 3 приведены результаты тестирования с помощью прогрессивных матриц (Scarr, 1981, стр. 282), согласно которым уровень исправленной наследуемости равен 0,60. Объединение результатов трёх работ даёт наследуемость интеллекта у афроамериканцев 0,72, она более высока у подростков (2 исследования), чем у 4-летних детей. Наследуемость интеллекта у афроамериканцев практически такая же, что и у европейцев (см. Главу 3).
82
Ричард Линн
Генетические и средовые объяснения низкого интеллекта африканцев
Вопрос о вкладах наследственности и среды в низкий IQ африканцев обсуждается с первых десятилетий двадцатого века, особенно в связи с проблемой низких показателей интеллекта афроамериканцев в Соединенных Штатах. Этой проблеме были посвящены сотни статей и множество книг, невозможно рассмотреть их все в достаточной мере. По рассматриваемому вопросу имеются три точки зрения:
Различие IQ между чернокожими и белыми полностью детерминировано средой или же, по крайней мере, нет убедительных свидетельств какого-либо вклада генетического фактора в низкий IQ чернокожих. Этой позиции придерживаются Флинн (Flynn, 1980), Макинтош (Mackintosh, 1998), Нисбетт (Nisbett, 1998), Фиш (Fish, 2002), Броди (Brody, 2003) и многие другие.
Различие IQ определено комбинацией генетических и средовых факторов. Эту позицию занимают Лоэлин, Линдзи и Спулер (Loehlin, Lindzey, Spuhler, 1975), Вернон (Vernon, 1979), а также Вальдман, Вай- нберг и Скарр (Waldman, Weinberg, Scarr, 1994, стр. 31), выполнившие одно из наиболее важных исследований по этой проблеме, в котором изучался IQ чернокожих детей, усыновлённых белыми супругами.
Различие IQ детерминировано преимущественно генетически. Эту точку зрения приняли Гаррет (Garrett, 1945,1961); Мак Гурк (McGurk, 1953а, 1953b), показавший, что при тестировании чернокожих и белых одинакового социально-экономического статуса чернокожие получают оценку IQ на 7,5 баллов ниже, чем белые; Каттнер (Kuttner, 1962), утверждавший, что различия в интеллекте между чернокжими и белыми отражаются в особенностях постройки зданий ранних цивилизаций; Шу (Shuey, 1966), который впервые свёл воедино результаты работ по изучению различий IQ чернокожих и белых (с 1916 до 1965 гг.); Осборн и Мак Гурк (Osborne, McGurk, 1982), дополнившие компиляцию Шу данными, полученными за период 1966-1980 гг., и Дженсен (Jensen, 1969, 1974, 1980, 1998), внесший огромный вклад в разработку этой проблемы и пришедший к заключению, что приблизительно две трети различия IQ между чернокожими и белыми в США обусловлено генетическими факторами. Другими авторами, принявшими преимущественно генетическую теорию, являются Шокли (Shockley, 1969), Айзенк (Eysenck, 1971), Бейкер (Baker, 1974),
Расовые различия в интеллекте
83
Леви (Levin, 1997), Раштон (Rushton, 2003) и автор настоящей книги (Lynn, 1994с, 2001).
Имеется шесть основных доводов в пользу присутствия некоторой генетической детерминации различия интеллекта африканцев и европейцев.
Во-первых, две расы эволюционировали независимо в различной среде обитания в течение приблизительно 100.000 лет (Mellars, Stringer, 1989; Cavalli-Sforza, 2000). Когда две популяции эволюционируют в условиях почти совершенной изоляции друг от друга в течение такого периода времени, между ними неизбежно развиваются генетические различия по всем признакам, подверженным генетической изменчивости. Эти различия развиваются в результате дрейфа генов, мутаций, эффекта основателя и, что наиболее важно, адаптации к различной среде обитания. Крайняя точка зрения сторонников средовой теории, состоящая в том, что нет никакого генетического различия между двумя расами в отношении интеллекта, бросает вызов этому основному принципу эволюционной биологии и должна быть отведена.
Во-вторых, постоянство, с которым африканцы получают низкие оценки IQ в самых различных местах проживания, может быть объяснено только действием сильного генетического фактора. Если бы за различия IQ разных популяций были ответственны только средовые факторы, то можно было бы ожидать найти некоторые страны, где бы африканцы получали более высокие оценки IQ, чем европейцы. То, что ни одной такой страны найти не удалось, указывает на присутствие сильного генетического фактора.
В-третьих, высокая наследуемость интеллекта, обнаруженная в близнецовых исследованиях чернокожих и белых в Соединенных Штатах, в Европе, Японии и в Индии, указывает на то, что интеллект в значительной степени определён генетическими факторами и делает маловероятным предположение, что различия между африканцами и европейцами или между любыми другими двумя расами могут быть детерминированы исключительно средовыми факторами.
В-четвертых, различие размера мозга между чернокожими и белыми указывает на генетическую детерминацию, учитывая высокую наследуемость размера мозга (примерно 0,9) и корреляцию между размером мозга и интеллектом (приблизительно 0,4).
В-пятых, некоторые эгалитаристы утверждают, что белый расизм может быть ответственным за снижение IQ чернокожих. Так, Вайнберг, Скарр и Вальдман пишут, что полученные ими данные о низком IQ чернокожих детей, усыновлённых белыми, «могут быть индикатором средовых влияний, таких как всепроникающее влияние расизма в американской жизни» (Weinberg, Scarr, Waldman, 1992, стр. 41) и «результаты оценки IQ совместимы с расово-ориентированными средовыми влияниями в том, что
84
Ричард Линн
касается порядка групповых средних» (там же, стр. 40). Макинтош (Macintosh, 1998, стр. 152) также возвращается к белому расизму в своей последней попытке обоснования того, что низкий IQ чернокожих приёмных детей может объясняться влиянием среды и предполагает, что вероятно «именно опыт жизни, будучи чёрным, в обществе, пропитанном белым расизмом, ответственен за снижение оценок IQ у чёрных детей», Эти эгалитаристы не объясняют, как гипотетический белый расизм мог снизить интеллект чернокожих детей, воспитанных белыми приёмными родителями из среднего класса. Нет никакого известного или вероятного механизма, посредством которого предполагаемый белый расизм мог бы понизить IQ чернокожих. Не пытаются они объяснить и то, как получается, что повсеместно в Африке южнее Сахары у африканцев, которые не подвергаются действию белого расизма, кроме как в Южной Африке, IQ составляет приблизительно 67 баллов.
Более того, если расизм снижает интеллект, то поразительно, что у евреев в Соединенных Штатах и Великобритании коэффициент интеллекта составляет около 108, как показано Линном (Lynn, 2004), хотя евреи в некоторой степени испытывали действие расизма в течение многих столетий. О высоком IQ американских евреев хорошо известно с 1930-х гг., данные широко публиковались Сторфером (Storfer, 1990), Мак Дональдом (MacDonald, 1994) и Хернсгайном и Мюрреем (Hermstein, Murray, 1994), но они удивительным образом остаются незамеченными сторонниками средовой теории, такими как Флинн (Flynn, 1980), Броди (Brody, 1992,2003), Найссер (Neisser,
, Макинтош (Macintosh, 1998), Дженкс и Филлипс (Jencks, Phillips, 1998), Нисбетт (Nisbett, 1998), Монтагю (Montagu, 1999) и Фиш (Fish, 2002).
В-шестых, Миннесотское исследование межрасового усыновления, выполненное Вальдманом, Скарр и Вайнбергом (Waldman, Scarr, Weinberg,
, было призвано показать, что в случае усыновления чернокожих младенцев белыми приёмными родителями они будут иметь такой же IQ, как белые. Авторы этого исследования изучили группы чернокожих, белых и расово-смешанных младенцев, все они были усыновлены белыми супругами среднего класса. В итоге оказалось, что к возрасту 17 лет IQ составлял 89 у чернокожих, 98 у межрасовых гибридов и 106 у белых. Таким образом, разрыв в 17 баллов IQ между чернокожими и белыми остаётся и тогда, когда они воспитываются в одинаковых условиях. Воспитание белыми приёмными родителями не оказывало положительного влияния на интеллект чернокожих детей, так как их IQ составлял 89 баллов, то есть был таким же, как у чернокожих в северных и центральных штатах США, откуда были родом младенцы. Межрасовая группа с её IQ в 98 баллов оказалась посредине между чернокожими и белыми, что предсказуемо, исходя из генетической природы различия. Полный анализ и обсуждение этого исследования были даны в работах Леви (Levin, 1994) и
Расовые различия в интеллекте
85
Линна (Lynn, 1994с), вместе с неубедительным ответом Вальдмана, Скарр и Вайнберга (Waldman, Weinberg, Scarr, 1994, стр. 43), в котором они утверждают, что «мы чувствуем, что баланс свидетельств, хотя и не окончательный, благоприятствует теории преобладания средовой этиологии расовых различий в интеллекте и что бремя доказательств лежит на тех исследователях, которые высказываются в пользу преобладания генетической природы расовых различий». Заметьте, что использование ими термина «преобладание средовой этиология» говорит о признании ими того, что генетические факторы также присутствуют.
Хотя результаты этого исследования показывают, что различия семейных условий не могут объяснить низкий IQ чернокожих, остаётся возможным предположить, что для чернокожих характерны худшие пренатальные условия как результат более плохого питания беременных чернокожих женщин или, вероятно, выкуривания большего количенства сигарет, что могло бы нарушить рост мозга эмбриона. Эти возможности предоставляются неправдоподобными, если учесть результаты исследований, показавших, что питание чернокожих американцев в течение двадцатого столетия было не хуже, чем у белых (см. Главу 13, Параграф 7). Другая возможность состоит в том, что у чернокожих младенцев мозг мог в большей степени повреждаться вследствие того, что чернокожие беременные женщины, может быть, выкуривают больше сигарет, так как есть свидетельства о том, что курение замедляет рост плода. Это предположение также представляется неправдоподобным, так как многочисленными исследованиями показано, что чернокожие курят меньше, чем белые.
Интересно отметить, что, несмотря на приверженность Вальдмана, Скарр и Вайнберга эгалитарным взглядам, они заключают, что их данные указывают на то, что и генетические и средовые факторы вносят вклад в различие IQ чернокожих и белых: «Мы думаем, что чрезвычайно неправдоподобно то, что эти различия определяются исключительно генетически или исключительно средой» (стр. 31). Таким образом, хотя в их данных нет ничего, что могло бы подтвердить данное заключение, так как они не представляют подтверждений какого либо вклада среды в низкий IQ чернокожих, их окончательная позиция существенно не отличается от взглядов Дженсена (Jensen, 1969), полагающего, что и генетические, и средовые факторы ответственны за низкий IQ чернокожих. Но в то время, как Дженсен предположил, что относительный вклад генетических факторов равняется примерно двум третям, а средовых—одной трети, Вальдман, Скарр и Вайнберг делают вывод лишь о вовлечении обоих факторов, хотя представляют количественную оценку вкладов соответствующих факторов.
В действительности, результаты Миннесотского Исследования Меж- расового Усыновления показывают, что оба вывода неверны. Заключение, которое должно быть сделано из этого исследования состоит в том,
86
Ричард Линн
что воспитание чернокожих детей в условиях жизни белой семьи среднего класса никак не влияет на их IQ к 17 годам.
Оценка генотипного IQ африканцев
Величины IQ в популяциях африканцев в Африке южнее Сахары на уровне около 67 баллов, приведённые в Таблице 4.1, являются функцией как генетических, так и средовых факторов. Теперь мы предпримем попытку оценить генотипный IQ африканцев. Генотипный IQ — это такой IQ, какой имели бы африканцы, если бы они были помещены в одинаковые с европейцами условия. Отправной точкой этого анализа является Миннесотское Исследование Межрасового Усыновления, результаты которого приведены в разделе 14 и которое показало, что различие IQ афроамериканцев и европейцев в 17 баллов сохраняется и тогда, когда они воспитываются в одинаковых семейных условиях. Из этого следует вывод о том, что различие IQ афроамериканцев и европейцев в Соединенных Штатах полностью генетически детерминировано. Хотя в рассматриваемом исследовании выявлено африканско-европейское различие IQ в 17 баллов, резонно предположить, что истинное афро-американско-европейское различие составляет 15 баллов, как это показано результатами многочисленных исследований, приведёнными в Таблице 4.4, и что различие в 17 баллов, полученное в данном исследовании, являются ошибкой выборки. Поэтому мы делаем вывод о том, что генотипный IQ афроамериканцев на 15 баллов ниже, чем у американских европеоидов. Следующий аргумент в пользу того, что IQ афроамериканцев полностью определён генетически состоит в том, что он остаётся постоянным в течение приблизительно 80 лет, несмотря на значительное улучшение их жизненных условий относительно европейцев.
Заключение о том, что афроамериканцы имеют генотипный IQ в 85 баллов, не означает, что у африканцев в Африке южнее Сахары генотипный IQ также составляет 85 баллов. Афроамериканцы не чистые африканцы, а являются гибридным населением с существенной долей европейских предков. Эта доля была оценена в 25 процентов Ридом (Reed, 1971) и Чак- раборты и соавт. (Chakraborty и др., 1992). Мы можем оценить, что у чистых африканцев в Африке и в Соединенных Штатах генотипный IQ равен 80, и что этот IQ увеличивается на 0,2 балла IQ на каждый процент генов европеоидов. Таким образом, у среднего афроамериканца IQ будет составлять 85 (80 + 25 х 0,2 = 85), такое значение подтверждено многочисленными исследованиями, приведёнными в Таблице 4.4. В Юго-Восточных штатах процент европейских генов у афроамериканцев весьма низок. Например, в Южной Каролине он был оценен в 6 процентов (Workman,
Расовые различия в интеллекте
87
1968), а в Джорджии в 11 процентов (Reed, 1969). Такая примесь европейских генов должна поднять их IQ на 1,2 и 2,2 балла соответственно, приводя к значениям IQ в 81,2 и 82,2. Это предсказание было подтверждено исследованием 1.800 афроамериканцев в пяти Юго-Восточных штатах (Kennedy, Van der Reit, White, 1963), в котором их IQ оказался равным 80,7 баллам по тесту Стенфорд-Бине 1960 г.
У афроамериканцев с 50-процентной примесью европейских генов IQ составит 90 (80 + {50 х 0,2 = 10} = 90). Это примерно средний IQ афроамериканцев в Северных штатах, где доля европейских предков приближается к 50 процентам. У афроамериканцев с 75 процентами европейских генов IQ будет на 15 баллов выше и составит 95 (80 + {75 х 0,2 = 15} = 95), что очень близко к показателю интеллекта 94 у расово-смешанных детей в Миннесотском Исследовании Межрасового Усыновления. У европейцев с 100 процентами европейских генов IQ будет 100 баллов.
Оценка генотипного IQ африканцев в 80 баллов означает, что средний IQ, который получили бы африканцы, если бы условия их существования были такими же, как у европейцев, составил бы 80 баллов. Везде в Африке южнее Сахары средний IQ африканцев равен приблизительно 67 баллам. Таким образом, можно заключить, что неблагоприятные условия существования в Африке южнее Сахары уменьшают IQ африканцев примерно на 13 баллов.
г ж с тти н шшн
Интеллект бушменов
Размер мозга бушменов
Пигмеи
Бушмены, иногда называемые также койсанами, санидами или капо- идами, и пигмеи являются двумя малыми расами районов Африки южнее Сахары в таксономиях классической антропологии, например Куна, Гарна и Бёрдселла (Coon, Gam, Birdsell, 1950). Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994) в их генетическом анализе человеческих популяций подтвердили, что эти два народа имеют отличные, но очень сходные генетические характеристики и образуют два связанных «кластера». Бушмены наряду с пигмеями и африканцами эволюционировали от прародителей Homo sapiensв экваториальной Восточной Африке. Предки бушменов мигрировали к югу и примерно 100.000 лет назад населяли почти всю Южную Африку. Огромное количество человеческих костей и артефактов было найдено в Пограничной пещере (Border Cave,в настоящее время в Свазиленде), они были датированы возрастом приблизительно 100.000 лет. Морфология костных останков указывает на то, что эти люди являлись гибридами африканцев и бушменов (Beaumont, deVilliers, Vogel, 1978).
Примерно полторы тысячи лет назад бушмены населяли большую часть Южной Африки, а пигмеи дождевые леса Западной и Центральной Африки. Примерно с 500 г. нашей эры африканцы (негроиды) с Севера начали вторгаться в их земли, множество их истребили и вытеснили большинство уцелевших бушменов в пустыню Калахари, а пигмеев в густые дождевые леса Центральной Африки. Родственными бушменам являются готтентоты, небольшие группы которых выжили в нескольких районах Южной Африки. Хотя эти две группы генетически очень близки, есть несколько генетических различий, таких как низкая частота группы крови В у бушменов и высокая её частота у готтентотов.
Многие готтентоты являются межрасовыми гибридами с бушменской и европейской родословной, что дало им более светлую кожу и более высокий рост, чем у бушменов (Cole, 1965). Бушмены выжили преимущественно в пустыне Калахари, где их численность составляет около 50 тысяч.
Расовые различия в интеллекте
89
Примерно столько же готтентотов. Наибольшая из выживших групп — племя Нама в Юго-Западной Африке, где их около 24 тыс. (Cole, 1965), есть ещё несколько более мелких групп к северу от Оранжевой реки.
Бушмены имеют ряд физических особенностей, отличающих их от аф- риканцев-негроидов. Волосы бушменов похожи на горошины перца, они растут спиралями с открытыми участками между пучками, тогда как у большинства африканцев густые курчавые спиралевидные волосы, спутывающиеся в плотную массу. Предполагается, что волосы бушменов в виде горошин перца развились как адаптивный признак в жарких и влажных лесах, где они жили многие тысячелетия, так как они защищают их от сильного солнечного света, но в то же время открытые участки между пучками позволяют поту испаряться. У пигмеев, оставшихся в тропических дождевых лесах, такие же волосы в виде горошин перца. Спутанные густые курчавые волосы африканских негроидов лучше подходят для жизни в сухой жаркой среде, так как обеспечивают более надёжную защиту от сильного солнечного света и уменьшают потоотделение. Кожа бушменов желтовато-коричневого цвета, в то время как у африканских негроидов она чёрная или тёмная. У некоторых бушменов встречается эпикантус на верхнем веке, подобный имеющемуся у аборигенов Восточной Азии и коренного населения Арктики, но менее выраженный. Преимущество эпикантуса для бушменов состоит вероятно в том, что он уменьшает ослепляющий эффект яркого солнечного света, отраженного от поверхности пустыни, подобно тому, как он уменьшает влияние ослепляющего блеска снега в случае аборигенов Восточной Азии и коренных жителей Арктики. Эта особенность наверняка возникла независимо в ходе конвергентной эволюции.
Отличительной особенностью бушменов является стеатопигия — очень большие ягодицы у женщин. Её адаптивное значение, вероятно, в сохранении питательных веществ и воды во время голода и недостатка воды. Гениталии бушменов уникальны среди всех человеческих рас. Пенис у бушменов торчит горизонтально, в то время как у бушменских женщин огромные малые срамные губы, свисающие приблизительно на 3 дюйма ниже влагалища. Адаптивные преимущества этих особенностей неизвестны.
Интеллект бушменов
Имеется всего три исследования интеллекта бушменов. В 1930-х гг. Портеус оценил интеллект 25 бушменов с помощью своего теста лабиринтов (Porteus, 1937), в котором нужно провести карандашом правильный путь через ряд лабиринтов возрастающей сложности. По этому тесту имеются нормы для европейских детей каждого возраста, относительно которых
90
Ричард Линн
умственный возраст бушменов составил семь с половиной лет, что соответствует IQ около 48 баллов. Во втором исследовании Портеус протестировал 197 взрослых бушменов с помощью Международной шкалы действия Лейтер и пришёл к заключению, что их умственный возраст составляет приблизительно 10 лет, что соответствует 62 баллам IQ. В третьем исследовании южноафриканский психолог Реунинг (Reuning, 1972) протестировал 108 бушменов и 159 африканцев с помощью теста завершения паттерна, заключающемся в выборе детали, завершающей рисунок. Основываясь на своём опыте применения теста Реунинг заключил, что он «может использоваться как надежный инструмент для оценки интеллекта на низших уровнях когнитивного развития и среди дописьменных народов» (Reuning, 1968, стр. 469). По результатам этого теста бушмены уступали африканцам приблизительно 15 баллов IQ, а ввиду того, что средний IQ африканцев составляет примерно 67 (см. Главу 4), это даёт оценку IQ бушменов приблизительно в 52 балла.
Реунинг также исследовал бушменов и африканцев с помощью рисуночного теста, в котором требовалось нарисовать человека. Этот тест идентичен тесту «нарисуй человека» Гудинаф (DAM), который является приемлемой мерой интеллекта. Рисунки, выполненные в тесте Гудинаф, оцениваются по количеству деталей и сложности, которые увеличиваются по мере взросления детей. Маленькие дети обычно рисуют людей в виде палочек с небольшим количеством деталей, в то время как старшие дети рисуют людей с объёмным телом и множеством деталей, таких как брови, большие пальцы и так далее. Реунинг писал (Reuning, 1968, стр. 476), что уровень рисунков бушменов был значительно ниже, чем у протестированных им шахтёров-негроидов, 76 процентов из которых были неграмотны. Он описал рисунки бушменов как характеризующиеся «чрезвычайной простотой..., большинство представляло собой фигурки из палочек... никаких деталей — пальцев на руках и ногах, волос, глаз и т. д...... Простота
бушменских рисунков «контрастирует с тенденцией негроидов изображать на своих рисунках много мелких деталей (пуговицы, волосы, пальцы рук и ног, курительная трубка и т. д.)». Различие в сложности рисунков бушменов и африканцев даёт дополнительное подтверждение более низкого интеллекта бушменов.
Реунинг отмечал, что среди бушменов наблюдалась значительная вариабельность в интеллекте, так же как и среди других народов, и что сами бушмены признают, что некоторые из них умны, в то время как другие тупоумные. В их языках есть слово «умный» для описания этого качества. Он пишет: «Когда тестирующий в конце теста хвалил испытуемого за хорошее выполнение, они сообщали нам через переводчика: «Мы могли бы вам сказать это, он (она) умный»» (1968, стр. 479). Более того, на основании интеркорреляции ряда тестов у бушменов пустыни Калахари был вы
Расовые различия в интеллекте
91
явлен фактор общего интелелкта (g) и также была выявлена корреляция результата выполнения тестов с общим мнением о том, кто умён.
В дополнение к тестированию интеллекта Реунинг оценил бушменов в сравнении с группами африканцев и белых на константность размера — способность оценить размер объекта на расстоянии. Он обнаружил, что бушмены по этой способности превосходили африканцев и европейцев и приписал это тому, что эта способность требуется для стрельбы из лука по животным с большого расстояния. Он нашел, что у бушменок также хорошая константность размера, хотя они и не охотятся. Он предположил, что развитие этой способности следует отнести, вероятно, на счёт преимущества, которое она даёт на охоте. Если это верно, то следует предположить, что эта способность могла ухудшиться у европейцев и аборигенов Восточной Азии, которые отошли от охоты приблизительно 8.000 лет назад и занялись сельским хозяйством.
Величины IQ, полученные в трёх исследованиях бушменов Портеуса и Реунинга, равняются 48,62 и 52 баллам, что при усреднении даёт 54 балла. Можно задаться вопросом, могли ли люди со средним IQ в 54 балла выжить как охотники-собиратели в пустыне Калахари, и, следовательно, верна ли эта оценка их интеллекта? Величина IQ в 54 балла находится в нижней части диапазона умеренной умственной отсталости для населения экономически развитых стран. Это не такая большая проблема, как можно подумать. Значительное большинство умеренно умственно отсталых в экономически развитых странах не находится в больницах или закрытых учреждениях, но живёт нормальной жизнью в обществе. Многие из них имеют детей и выполняют обязанности по дому или же занимаются работой, не требующей умственного напряжения. Коэффициент интеллекта в 54 балла соответствует умственному возрасту среднего европейского 8-летнего ребенка, а средний 8-летний европеец умеет читать, писать, знает арифметику и без труда освоил бы навыки сбора пищи и охоты, чем занимаются бушмены. Среднего 8-летнего европейца легко научить собирать ягоды, складывать их в корзины и относить домой, собирать страусиные яйца и использовать раковины для хранения воды или попадать в цель с некоторого расстояния стрелами из лука. До повсеместного введения всеобщего образования детей в Северной Америке и Европе во второй половине девятнадцатого столетия значительное большинство 8-летних детей продуктивно работало на фермах и иногда трубочистами или на фабриках и шахтах. И сегодня многие дети этого возраста в Африке, Индии, Пакистане, Бангладеш и в значительном числе повсеместно в Латинской Америке и в других развивающихся странах работают на фермах, а некоторые из них выполняют работу средней квалификации, такую как выработка ковров и обслуживание швейных машин. Коэффициент интеллекта большинства бушменов лежит в диапазоне от 35 до 75 баллов. Показатель в 35 баллов
92
Ричард Линн
приблизительно соответствует умственному возрасту средних европейских детей пяти с половиной лет, a IQ в 75 баллов это примерно умственный возраст средних европейских детей одиннадцати с половиной лет. Средний европейский ребенок пяти с половиной лет свободно разговаривает и способен к выполнению низко квалифицированной работы, и даже наименее интеллектуально развитые бушмены должны быть способны к этому.
Более того, обезьяны с умственными способностями примерно 4-летних детей весьма хорошо выживают как собиратели и по случаю охотники, также как в прошлом ранние гоминиды с IQ около 40 баллов и размером мозга намного меньшим, чем у современных бушменов. Поэтому нет ничего удивительного в том, что современные бушмены со средним IQ примерно в 54 балла и диапазоном IQ в основном в пределах от 35 до 75 баллов способны выжить как охотники-собиратели и выполняя работу низкой и средней квалификации на фермах, чем некоторые из них занимались в последние десятилетия двадцатого века.
Размер мозга
Размер мозга бушменов был оценен в 1.250 см3 Дреннаном (Drennan, 1937) и немного выше — в 1.270 см3 Смитом и Билсом (Smith, Beals, 1990). Массив данных Смита и Билса включал также африканцев-негроидов, размер мозга которых составил 1.282 см3, то есть немного больше, чем у бушменов. Это согласуется с более высоким средним IQ африканцев в 67 баллов в сравнении с 54 баллами бушменов, хотя такое различие размеров мозга может объяснить лишь небольшую долю различия интеллекта. Меньший размер мозга и более низкий интеллект бушменов в сравнении с африканцами говорит о том, что мозг африканцев увеличился за последние 100.000 лет или около того, так как современные африканцы и бушмены произошли от одних и тех же предков. Первоначально размер их мозга должен был быть одинаковыми, но затем мозг африканцев увеличился либо в результате более жёсткого давления отбора, либо в результате благоприятных мутаций.
Пигмеи
Пигмеи обитают в экваториальных дождевых лесах Заира, теперь называемого Конго, и в Центральноафриканской Республике. В конце двадцатого столетия предполагалось, что их численность лежит в пределах от
Расовые различия в интеллекте
93
100 до 200 тыс. Самые чистые пигмеи — племяМбути, обитающие в лесу Итури Северо-Восточного Конго, предполагается что их от 30 до 60 тыс. или около того. Другие популяции пигмеев более гибридизированы с африканцами. Средний рост Мбути составляет приблизительно 1,4 м. Дети пигмеев до достижения пубертатного возраста имеют нормальный рост, но когда они становятся подростками, у них не происходит скачка роста, как у других народов, из-за низкой выработки у них инсулино-подобного фактора роста 1. Большинство пигмеев остались охотниками-собирателя- ми. Обычно они живут маленькими группами приблизительно в 30 человек и переходят с места на место. Они не преуспели в одомашнивании животных и окультуривании растений. В начале двадцать первого столетия пигмеи в Конго были описаны Ченгом (Cheung, 2003) как живущие «в глухих лесах, перебиваясь охотой и собирательством».
Было проведено всего одно исследование интеллекта пигмеев. Это работа Вудворта (Woodworth, 1910) использовавшего тест «Доски Сеге- на». В этом тесте кубики различной формы нужно вставлять в соответствующие отверстия. Он обнаружил, что пигмеи выполняют тест намного хуже, чем другие народы, включая эскимосов, американских индейцев и филиппинцев, но он не измерил их способности. Судя по образу жизни, их интеллект, по-видимому, ниже, чем у африканцев-негроидов. Большинство пигмеев всё ещё сохраняет примитивный образ жизни охотников-со- бирателей, в то время как многие африканцы-негроиды стали фермерами в течение последних нескольких сотен лет. В двадцатом столетии некоторое число пигмеев работало на фермах у африканцев-негроидов, где они «всегда являются низшей кастой, будучи «наследственными слугами» фермеров» согласно Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994, стр. 178). Термин «наследственные слуги» является, по-видимому, эвфемизмом для слова «рабы». Порабощение пигмеев аф- риканцами-негроидами согласуется с общим принципом, состоящим в том, что более интеллектуальные расы обычно побеждают и порабощают менее интеллектуальные. Точно так же европейцы и аборигены Южной Азии часто порабощали африканцев, но не наоборот.
Г Ж с
ЛШИГШ1 40Ш410Й ЛЗКН Н (ШР410В ЯФРШ
Интеллект аборигенов Южной Азии и Северной Африки
Выходцы из Южной Азии и Северной Африки в Великобритании и Австралии
Выходцы из Южной Азии и Северной Африки в континентальной Европе
Индусы в Африке, на Фиджи, в Малайзии и на Маврикии
Учащиеся полной средней школы и студенты университетов
Размер мозга
Наследуемость интеллекта у аборигенов Южной Азии и Северной Африки
Генетические и средовые детерминанты интеллекта аборигенов Южной Азии и Северной Африки
Интеллект в Израиле
Аборигены Южной Азии и Северной Африки — автохтонное население Южной Азии от Бангладеш на Востоке и далее на Запад — Индии, Пакистана, Ирака, Ирана, стран Персидского залива и Ближнего Востока, Турции, а также Африки севернее пустыни Сахара. Они близко родственны европейцам и в некоторых таксономиях классической антропологии, к примеру к Куна, Гарна и Бёрдселла (Coon, Gam, Birdsel, 1950), эти народы рассматривались как одна раса, называемая кавказоидами. Однако Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994) путём генетического анализа различий человечества показали, что аборигены Южной Азии и Северной Африки образуют отдельный генетический «кластер», отличный от европейцев. Поэтому они рассматриваются нами как отдельная раса.
Расовые различия в интеллекте
95
Интеллект аборигенов Южной Азии и Северной Африки
Данные исследований интеллекта аборигенов Южной Азии и Северной Африки представлены в таблице 6.1. Приводятся значения только для общего интеллекта (g), так как практически нет данных об их вербальных или визуализационных способностях.
В строках с первой по тринадцатую приведены результаты двенадцати исследований, выполненных в разных районах Индии и давших результаты от 78 до 88 баллов с медианой 82. В строках с четырнадцатой по семнадцатую приведены результаты в 84, 83, 89 и 80 баллов IQ для выборок школьников в Иране, все выборки из города Шираз. В строках 18 и 19 приведены значения IQ в 87 баллов для детей и взрослых в Ираке. В строке 20 представлен IQ для арабов в Израиле, полученный на выборке стандартизации теста WISC-R в сравнении с евреями (IQ евреев в Израиле лежит в пределах 90-97 и рассматривается в параграфе 9). В строке 21 дан IQ 84 для Иордании, рассчитанный по данным выборки стандартизации теста КАВС. В строке 22 приведён IQ в 86 баллов для Кувейта, полученный из данных выборки стандартизации прогрессивных матриц у школьников. В строке 23 дан IQ 82 для школьников из разных районов Ливана. В строке 24 дан IQ в 78 баллов для Непала, полученный у детей из 34 школ в городах, деревнях и в районах джунглей. В строке 25 представлен IQ 84 для подростков из Исламабадского региона Пакистана. Группа сравнения состояла из 707 европейских детей из Канады, данные о ней приведены в отчёте об этом исследовании. В строке 26 дан IQ 84 для детей в Пакистане из 9 школ, расположенных в бедных и богатых районах неподалёку от Карачи. В строке 27 приведён IQ в 78 баллов для школьников из Катара, в строке 28 — IQ в 79 баллов для учащихся младших классов сельской школы на Шри-Ланке. В строке 29 представлен IQ 83 учащихся первого класса школ Сирии в сравнении с двумястами школьниками того же возраста из Германии. В строках с тридцатой по тридцать вторую даны значения IQ в 84,90 и 96 баллов для школьников из Турции. В строке 33 приведён IQ 85 для Йемена, полученный из стандартизации теста СРМ на выборке 6-11-летних детей.
В строках с 34 по 37 даны значения IQ для выборок из Северной Африки. В строке 34 приведён IQ 84, полученный на смешанной выборке аборигенов Северной Африки из Алжира, Марокко и Туниса. В строках 35, 36 и 37 представлены значения IQ в 77, 81 и 83 балла для школьников из Египта. Тридцать семь значений IQ аборигенов Южной Азии и Северной
96
Ричард Линн
Таблица 6.1. IQ аборигенов Южной Азии и Северной Африки

Расовые различия в интеллекте
97
Африки достаточно однородны. За исключением Турции, все величины IQ находятся в диапазоне от 77 до 89 баллов. Медианное значение IQ по всем выборкам, составившее 84 балла, представляется наилучшей оценкой IQ аборигенов Южной Азии и Северной Африки. Медианное значение IQ результатов трёх исследований в Турции составило 90 баллов, что выше, чем в остальных районах Южной Азии и Северной Африки. Скорее всего это можно объяснить тем, что Турция в течение многих столетий распространялась на Европу и Азию. Географическая близость Турции и Юго-Восточной Европы обусловила смешение турецких и европейских генов, что произвело европейско-южноазиатскую клину признаков или генетически гибридное население, так что в результате современные турки и греки генетически весьма сходны, как это показано Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994) и о чём уже говорилось в Главе 3.
Выходцы из Южной Азии и Северной Африки в Великобритании и Австралии
Величины IQ выходцев из Южной Азии в Европе и в Австралии приведены Таблице 6. 2.
Таблица 6.2. IQ выходцев из Южной Азии и Северной Африки в Великобритании и Австралии

В строке 1 дан IQ 87 для детей индусов в Лондоне, вычисленный Верноном (Vernon, 1969, стр. 169) по данным, собранным в середине 1960-х гг.
98
Ричард Линн
Управлением Образования Старого Лондона (ILEA). В строке 2 представлен IQ 93 для выборки пакистанских детей из Лондона. В строке 3 дан IQ в 91 балл школьников-индусов из общеобразовательной школы в Эссексе, полученный в исследовании, в котором афро-карибские дети из той же школы получили оценку IQ в 85 баллов. В строках 4 и 5 приведены значения IQ в 94 и 89 баллов для индийских и пакистанских детей из городов Британского Мидлендса. Афро-карибские дети из той же школы получили оценку IQ в 86 баллов, что подтверждает результат, представленный в строке 3, свидетельствующий о более высоком IQ у аборигенов Южной Азии, чем у африканцев. В строках 6 и 7 приведены значения IQ 83 и 97 для индусов, а в строках 8 и 9IQ 93 и 96 баллов для пакистанских детей по всей Великобритании.
В строках с десятой по двенадцатую представлены данные исследования, в котором дети из Пакистана, Индии и Бангладеш получили оценки IQ в 93,92 и 92 балла, что свидетельствует об отсутствии различий IQ между этими тремя группами населения субконтинента Индии. Эти величины IQ, рассчитанные относительно 100 баллов для посещавших те же школы белых детей, вероятно несколько завышены, так как около 7 процентов белых детей, в основном из среднего класса с более высоким IQ, посещают частные школы, а белые родители среднего класса, которые посылают детей в государственные школы, стремятся не отправлять их в школы с большим количеством иммигрантов. Результатом этого будет завышение IQ аборигенов Южной Азии относительно британских национальных норм. Для использованных тестов нет национальных норм, ввиду чего величина завышения IQ не может быть определена количественно, но вероятно она составляет около 5 баллов. В строке 13 дан IQ 89 для аборигенов Южной Азии, иммигрировавших в Австралию.
Разброс значений IQ выходцев из Южной Азии в Великобритании весьма велик, от 83 до 97 баллов. Одна из причин этого значительного разброса в том, что IQ растёт со временем проживания в Великобритании, это было показано в двух исследованиях. Во-первых, в строках 6 и 7 дан IQ для невербального рассуждения в 83 балла для индийских детей, проживших в Великобритании меньше 4 лет, и в 97 баллов для проживших четыре или более лет: выявлено увеличение IQ на 14 баллов вследствие проживания в Великобритании. Любопытно отметить, что IQ индийских детей, проживших в Великобритании менее 4 лет (83 балла) почти равен IQ индусов в Индии (82 балла), приведённому в Таблице 6.1. Во- вторых, в строках 8 и 9 даны значения IQ для невербального рассуждения в 93 балла, полученные у пакистанских детей, проживших в Великобритании менее 4 лет и в 96 баллов для проживших четыре года и более, показан прирост IQ на 3 балла при более длительном проживании в Великобритании.
Расовые различия в интеллекте
99
Медианное значение IQ у выходцев из Южной Азии, проживающих в Великобритании, составило 89 баллов, такое же значение, приведённое в последней строке таблицы, выявлено у выходцев из Южной Азии в Австралии. Это немного выше, чем IQ местного населения в Южной Азии (84 балла), и согласуется с данными, говорящими об увеличении IQ по мере возрастания срока проживания в Великобритании и Австралии. Прирост IQ может быть обусловлен разными факторами. Недавние иммигранты испытывают трудности в общении на английском языке и могут хуже выполнять даже невербальные тесты вследствие трудности понимания инструкций, предъявляемых на английском. Вдобавок к этому, родившиеся в Великобритании могут иметь преимущества за счёт лучшего питания и образования, чем у сравниваемых с ними детей в их собственных странах.
Выходцы из Южной Азии и Северной Африки в континентальной Европе
Значения IQ выходцев из Южной Азии и Северной Африки в континентальной Европе представлены Таблице 6.3.
В строке 1 приведён IQ в 86 баллов для турок, иммигрировавших в Германию. В строках со второй по семнадцатую представлены результаты 16 исследований по определению IQ у иммигрантов из Южной Азии и Северной Африки в Нидерландах. Полезный обзор ряда таких работ был опубликован Те Нийенуисом и ван дер Флиером (Те Nijenhuis, van der Flier, 2001). В строке 2 дан IQ в 78 баллов для выборки детей иммигрантов первого поколения, а в строке 3 — в 79 баллов для выборки детей иммигрантов второго поколения из Турции. Оба значения IQ низки и указывают на отсутствие повышения интеллекта у иммигрантов второго поколения. В строке 4 приведён IQ в 75 баллов для выборки детей иммигрантов первого поколения из Марокко, а в строке 5 — в 79 баллов для выборки детей марокканцев, иммигрантов второго поколения. Вновь оба значения IQ низки, хотя, по-видимому, имеется некоторое повышение интеллекта у иммигрантов второго поколения, как это было найдено в исследованиях среди иммигрантов в Великобритании.
В строке 6 приведён IQ в 83 балла для детей иммигрантов из Марокко и Турции. В строках 7 и 8 даны значения IQ в 85 и 84 балла для двух других выборок детей марокканских и турецких иммигрантов. В строке 9 дан IQ в 88 баллов для выборки индусов, что на 6 баллов выше медианного значения IQ индусов в Индии (82 балла). В строке 10 представлен IQ 93 для марокканских и турецких детей, что является средней величиной значений IQ 92 и 94, полученных с помощью тестов Отиса и CITO, оба теста в
100
Ричард Линн
Таблица 6.3. IQ выходцев из Южной Азии и Северной Африки в континентальной Европе

основном вербальные. В строке 11 даны значения IQ в 84 балла, полученные у турецких (п=24) и марокканских (п=9) детей из выборки стандартизации невербального теста Снайдерса-Оомана; величины IQ этих родившихся в Нидерландах детей были такими же, как и у проживших в стране от 1 до 6 лет. В строках 12 и 13 приведены IQ в 85 и 84 балла для выборок марокканских и турецких детей. В строках 14 и 15 даны IQ 84 и 88 для взрослых марокканцев и турок, полученные с помощью батареи тестов общих способностей (GATB) — голландского теста, включающего восемь субтестов на оценку словарной и арифметической способностей, скорости восприятия и т.д. Иммигранты из Турции и Марокко плохо выполняли словарный тест, так как плохо владели голландским языком, ввиду чего результаты этого субтеста были исключены при вычислении IQ. Значения фактора g представляют собой среднее по остальным семи субтестам. В строке 16 представлен IQ в 92 балла для детей проживающих в Нидерлан
Расовые различия в интеллекте
101
дах мусульман из Турции и Марокко в сравнении примерно с 69 тысячами детей голландцев — европейцев, это результат арифметического теста, он введён как показатель вербальнго IQ. Средний словарный IQ в этой выборке составил 85 баллов, но его не учитывали, так как для большинства этих детей голландский язык не был родным. В строке 17 дан IQ в 94 балла для детей иммигрантов второго поколения, из которых 72 процента были из Турции и Марокко и 10 процентов из Суринама и с Нидерландских Антильских островов. Их вербальный IQ составил 80 баллов, но его исключили из рассмотрения на том основании, что для большинства детей голландский язык не был родным. Результаты исследований в Нидерландах близки к полученным в Великобритании. Медианное значение величин IQ для первых восьми исследований среди иммигрантов первого поколения из Южной Азии и Северной Африки составило 84 балла—столько же, сколько в местах их исконного проживания. В строке 18 дан IQ 83 для цыган из Словакии. Этот результат приведён здесь потому, что цыгане, или как их стали называть «рома», являются южноазиатским племенем, мигрировавшим из Северо-Западной Индии в период между девятым и четырнадцатым веками нашей эры. Это показал лингвистический анализ их языка «Романи», который, как был обнаружено, имеет индийское происхождение, а также генетическими исследованиями (Pearson, 1985; Fraser,
. Они остались в значительной степени изолированными от европейцев и их IQ типичен для аборигенов Южной Азии.
Индусы в Африке, на Фиджи, в Малайзии и на Маврикии
В нескольких странах Африки есть популяции индусов. В Южной Африке их численность составляет приблизительно один миллион, примерно 84 процента проживает в Натале и 14 процентов в Трансваале. Индусы проживают также в Кении и Танзании. Предки этих индусов были ввезены британцами и немцами во время колониального правления для выполнения разных работ, включая прокладку железных дорог. Данные исследований по определению IQ индусов в Африке приведены в Таблице 6.4.
В строке 1 представлен IQ в 77 баллов по результатам первого исследования IQ у индусов в Южной Африке в сравнении с 65 баллами африканцев. В строке 2 дан IQ в 88 баллов для студентов — индусов, изучавших компьютерное программирование, в сравнении со 100 баллами в сопоставимой выборке из 243 белых. В строке 3 представлен IQ 86, вычисленный по данным выборок стандартизации теста Юношеских южноафриканских индивидуальных шкал. Этот тест напоминает тест Векслера. Нормы для
102
Ричард Линн
Таблица 6.4. Показатели интеллекта индусов в Африке, на Фиджи, в Малайзии и на Маврики

индусов были вычислены относительно выборки стандартизации южноафриканского белого населения. Тест содержит шкалу числовой способности, по которой индусы получили IQ 86 и которая вносит вклад в их общий IQ. В строке 4 приведён IQ 91 относительно британских норм 1979 г. У белого населения Южной Африки IQ составил 98; следовательно, индусы уступают 7 баллов IQ южноафриканским белым. В строке 5 дан IQ 83 относительно южноафриканских белых по данным Южноафриканского юношеского теста способностей. Этот тест включает два субтеста на память, по которым индусы получили IQ в 89 баллов. В том же исследовании африканцы получили оценку IQ в 63 балла, что вновь показывает, что оценки IQ индусов в Южной Африке значительно выше, чем у африканцев.
Медианное значение результатов пяти исследований IQ индусов в Южной Африке составило 86 баллов. Это немного выше медианного значения IQ индусов в Индии (82 балла) и немного ниже, чем у индусов, родившихся в Великобритании (89 баллов). Вероятно, причина этих различий в том, что уровень жизни ниже всего в Индии, выше в Южной Африке и самый высокий в Великобритании, что в некоторой мере повлияло на уровень интеллекта. Возможно также, что имелись различия в интеллекте предков мигрантов, от которых произошли индусы, проживающие ныне в Южной Африке и Великобритании. Предков южноафриканских индусов нанимали в основном для работы на плантациях сахарного тростника и табака, и возможно их IQ не был столь высок, как у мигрировавших в Великобританию во второй половине двадцатого столетия.
В строке 6 дан IQ в 91 балл для индусов в Танзании. Выборка состояла из учащихся средних школ, сдавших вступительный экзамен, следовательно это значение IQ несколько завышено. Вероятно, величина IQ в этой
Расовые различия в интеллекте
103
выборке примерно на 8 баллов выше, чем во всей популяции индусов в Танзании, IQ которых, следовательно, может быть оценен приблизительно в 83 балла, что близко к IQ индусов в Индии (82 балла), приведённому в Таблице 6.1. В этом же исследовании IQ африканцев из тех же элитных школах составил 78 баллов. Это различие подтверждают данные многих исследований в Южной Африке и в Великобритании, говорящие о том, что индусы получают значительно более высокие оценки IQ, чем живущие в тех же условиях африканцы.
В строке 7 представлен IQ в 82 балла для индусов на о-вах Фиджи, где проживают примерно равные количества индусов и автохтонных фиджийцев. Фиджийцы в этом исследовании получили среднюю оценку IQ в 84 балла. В строке 8 дан IQ 88 для индусов в Малайзии, полученный при стандартизации стандартных прогрессивных матриц Равена. В строке 9 приведён IQ 89 для группы 11-летних детей на о-ве Маврикий, описанной как «выборка местного сообщества», в которой 69 процентов были индусами, а остальные креолами — смесью европейцев и выходцев из районов Африки южнее Сахары, IQ последних был на 2,5 балла ниже чем, у индусов (Raine и др., 2002). Значения IQ, приведённые в Таблице 6.4, лежат в диапазоне от 77 до 91 баллов с медианой 88 — немного выше, чем в Индии и почти на том же уровне, как у индусов в Европе. Вероятно, это объясняется тем, что индусы вне Индии живут в лучших условиях. Другая возможная причина в том, что те, кто эмигрировал из Индии, имели интеллект выше среднего.
Учащиеся полной средней школы и студенты
Исследования интеллекта учащихся полной средней школы, студентов колледжей и университетов в Южной Азии и Северной Африке, приведены в Таблице 6.5. Можно ожидать, что их показатели будут несколько выше, чем в общепопуляционных выборках, и мы увидим, что это действительно так.
В строке 1 дан IQ в 85 баллов для студентов Александрийского университета в Египте. Это лишь незначительно выше приведённой в Таблице 6.1 оценки IQ в 83 балла, полученной с помощью того же теста по всей популяции. В строке 2 приведён результат раннего исследования 1926 года, в котором студенты второго курса университета Калькутты получили оценку IQ в 95 баллов. Применялся тест Стенфорд, по которому американские студенты из Стенфордского университета получили среднюю оценку в 113 баллов. В строке 3 представлен IQ в 93 балла для 14-летних учащихся школы Св. Хавьера в Дели, про которых сообщалось, что они происходят из
104
Ричард Линн
Таблица 6.5. IQ учащихся полной средней школы и университетов в Южной Азии и Северной Африке

семей, принадлежащих к высшему слою общества. В строках 4 и 5 даны IQ в 90 и 88 баллов для студентов из Пенджабского университета. В строке 6 приведён IQ 88 для студенток разных колледжей индийского города Амритсар. В строке 7 дан IQ 90 для университетских студентов инженерных, экономических и гуманитарных специальностей из Тегерана. В строке 8 дан IQ в 92 балла для учащихся полной средней школы в Багдаде, описанных автором исследования как «высоко селективная группа, так как обучение в полной средней школе не обязательно, и достигшие этого уровня учащиеся должны выдержать строгие экзамены» (Alzohaie, 1966, стр. 476).
В строках 9 и 10 даны IQ в 98 и 102 балла для студентов технических специальностей из Университета Витватерсранда в Южной Африке. В этом исследовании европейские студенты с тех же факультетов получили оценки IQ в 106 и 113 баллов, а чернокожие африканские студенты в 93 и 99 баллов. В строке 11 приведён IQ в 106 баллов ещё для одной выборки студентов технических специальностей из Университета Витватерсранда в Южной Африке. В этом исследовании европейские студенты с тех же факультетов получили IQ в 116 баллов, а чернокожие африканские студенты в 101 балл. Таким образом, в этих трёх исследованиях студентов величины IQ индусов попадают в диапазон между оценками белых и чернокожих,
Расовые различия в интеллекте
105
также как и в общепопуляционных выборках. Кроме того, в этих исследованиях показатели интеллекта индусов оказались несколько выше, чем в Южной Азии и Северной Африке. Вероятно, это можно отнести за счёт того, что IQ индусов в Южной Африке выше, а также того, что на инженерный факультет Университета Витватерсранда принимают сравнительно талантливых студентов.
В строках 12 и 13 даны IQ в 92 и 101 балл для студентов университетов в Турции. В строке 12 приведён IQ 92 для 15 студенток и 24 студентов- стоматологов из университета Ататюрка в городе Эрзерум. В строке 13 дан IQ в 101 балл для студентов-медиков того же университета.
Медиана значений по всем исследованиям составила 92 балла IQ — на восемь баллов выше, чем по общепопуляционным выборкам аборигенов Южной Азии и Северной Африки. Эти исследования интересны тем, что они показывают, что университетские студенты — выходцы из Южной Азии и Северной Африки с основательным образованием, принадлежащие к семьям высшего и среднего классов, получают оценки IQ ниже среднеевропейских. Это указывает на то, что недостаточное образование вряд ли является главным фактором, ответственным за низкие оценки IQ в общепопуляционных выборках. Величины IQ студентов из Южной Азии и Северной Африки также оказались ниже медианного значения для европейских студентов колледжей, равного 105 баллам (Таблица 3.3). Таким образом, разница IQ в 15 баллов между европейцами и выходцами из Южной Азии и Северной Африки в общепопуляционных выборках близка к разнице в 14 баллов между студентами колледжей.
Размер мозга аборигенов Южной Азии
В Таблице 6.6. представлено четыре набора данных измерений размера мозга аборигенов Южной Азии в сравнении с европейцами.
Таблица 6.6. Размер мозга (см3) европейцев и аборигенов Южной Азии

В строке 1 представлены данные, полученные Смитом и Билсом (Smith, Beals, 1990) на материале коллекции приблизительно в 20.000 черепов, собранных со всего мира. Эти данные демонстрируют преимущество европейцев
106
Ричард Линн
примерно на 84см3. В строке 2 приведены данные, собранные из различных источников Гровсом, утверждающим, что расовых различий в размере мозга нет, но тем не менее его данные показывают преимущество европейцев в 63см3. В строке 3 приведён средний размер мозга для шести выборок европейцев из Северной Америки и Европы и для двух выборок из Индии по данным, собранным Юргенсом и соавторами (Jurgens и др., 1990) и проанализированным Раштоном (Rushton, 2000, стр. 124); показано преимущество европейцев в 126см3. В строке 4 представлены средние значения, полученные американским Национальным управлением по аэронавтике и исследованиям космического пространства (НАСА) на 19 выборках военнослужащих — европейцев мужского пола, а также на выборке иранских военнослужащих-мужчин; данные показывают преимущество европейцев в 114см3. Значения этих четырех наборов данных говорят о большем размере мозга европейцев, их можно считать согласованными, учитывая, что они были собраны с использованием различных методов. Смит и Билс получили свои результаты путём измерения объёма черепов, данные Гровса получены из различных источников, а наборы данных в строках 3 и 4 были вычислены по внешним измерениям головы живых людей. В среднем по четырём наборам данных европейские кавказоиды имеют преимущество в 97см3.
Наследуемость интеллекта у аборигенов Южной Азии и Северной Африки
Имеется два исследования наследуемости интеллекта у индусов, оба были выполнены с использованием метода сравнения величин IQ монозиготных (MZ) и дизиготных (DZ) близнецов. Пал, Шиам и Сингх (Pal, Shyam, Singh, 1997) опубликовали результаты исследования взрослых близнецов (30 пар MZ и 30 пар однополых DZ), вычисленная наследуемость составила 0,81. При введении поправки на не 100-процентную надёжность теста, полагая надёжность теста в 0,9, наследуемость станет равной 0,90. Авторы другого исследования (Nathawat, Puri, 1995) получили наследуемость 0,9, которая после введения поправки, полагая надёжность теста в 0,9, увеличивается до 1,0. Таким образом, наследуемость интеллекта в Индии немного выше, чем у европейцев (0,83).
Расовые различия в интеллекте
107
Генетические и средовые детерминанты интеллекта аборигенов Южной Азии и Северной Африки
Мы видели, что средняя величина представленных в таблице 6.1 оценок IQ у аборигенов Южной Азии и Северной Африки равна 84 баллам. Эта величина занижена вследствие низкого уровня жизни этих народов. В отчёте ЮНИСЕФ о недоедании в Южной Азии и Северной Африке в начале 1990-х гг. (UNICEF, 1996) говорится о задержке роста у 24 процентов детей на Ближнем Востоке и в Северной Африке и у 60 процентов детей в Южной Азии, что указывает на недостаточность питания. Несомненно, это неблагоприятно сказывается на интеллекте. Но представляется вероятным, что генетические факторы также вовлечены. Во-первых, очень высокая наследуемость интеллекта у аборигенов Южной Азии и у европейцев говорит о том, что генетические факторы в значительной степени ответственны за различия в интеллекте этих двух популяций, ввиду чего вполне вероятно, что они вносят свой вклад в различия между ними. Во- вторых, было показано, что у выходцев из Южной Азии и Северной Африки, живущих вполне обеспеченно в Великобритании, Австралии и Нидерландах, медианные значения IQ составляют 89, 89 и 94 балла, соответственно. Все эти значения выше средней оценки IQ в 84 балла у их соотечественников, живущих в худших условиях в странах своего исконного проживания. Эти цифры показывают, что у выходцев из Южной Азии и Северной Африки, выросших в благоприятной среде европейских стран, IQ увеличивается, но не достигает уровня европейцев. Это предлагает наличие генетических факторов. В-третьих, IQ индусов в Южной Африке равен 86 баллам. Это выше 82 баллов их соотечественников в Индии, что может быть отнесено на счёт лучших условий жизни, но существенно ниже IQ европейцев. Индусы были привезены в Наталь в 1850-х гг. для работы на плантациях сахарного тростника (Johnston, 1930). Они приспосабливались к условиям Южной Африки на протяжении жизни четырёх — шести поколений, живут в этой стране приблизительно в тех же условиях, что и европейцы, однако значительное различие в IQ остаётся, что говорит о генетическом различии между этими двумя популяциями. В-четвертых, средний размер мозга у аборигенов Южной Азии приблизительно на 8 процентов меньше, чем у европейцев, что отчасти может быть обусловлено неоптимальным питанием, но вероятно, для этого есть и некоторое генетическое основание, что вносит свой вклад в различия интеллекта.
108
Ричард Линн
Интеллект в Израиле
Интеллект в Израиле выше, чем в других странах Южной Азии и Северной Африки. Результаты восьми исследований интеллекта в Израиле приведены в Таблице 6.7.
Таблица 6.7. Интеллект в Израиле
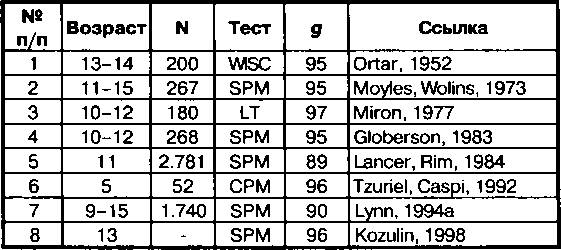
Представленные показатели интеллекта лежат в диапазоне от 89 до 97 баллов с медианой в 95. Это существенно выше медианного значения остального населения Южной Азии (84 балла), то есть у евреев более высокий IQ, чем у остального населения региона. В Израиле около 20 процентов населения составляют арабы, IQ которых равен 86 баллам (см. Таблицу 6.1, строку 20), практически такой же, как и у других аборигенов Южной Азии на Ближнем Востоке. Сорок процентов населения составляют европейские евреи (главным образом ашкенази из России и Восточной Европы) и 40 процентов — восточные евреи (мицраим8) из Азии и Северной Африки. В трёх выполненных в Израиле исследованиях обнаружено, что у ашкенази средний IQ приблизительно на 12 баллов выше, чем у восточных евреев (Zeidner, 1987а; Burg, Belmont, 1990; Lieblich, Ninio, Kugelmass, 1972). Величина IQ для Израиля в 95 баллов является средним взвешенным значений евреев ашкенази (103 балла), восточных евреев (91 балл — на 12 баллов ниже) и арабов (86 баллов). Более низкий IQ арабов в Израиле по сравнению с евреями подтвержден Цайднером (Zeidner, 1987а), представившим сообщение, что арабские абитуриенты при поступлении в университет получают оценку IQ на 15 баллов ниже, чем еврейские.
Имеется два вопроса относительно еврейского IQ, требующих объяснения. Во-первых, почему у евреев-ашкенази в Израиле IQ составляет 103
8 Восточные евреи, более известные как «сефарды» (прим. пер.)
Расовые различия в интеллекте
109
балла. Это не очень удивительно, так как имеются многочисленные свидетельства о том, что у евреев-ашкенази в Соединенных Штатах и Великобритании показатели IQ существенно выше, чем у неевреев. В опубликованном в 1920-х гг. исследовании сообщалось, что у 10-летних еврейских детей в Соединенных Штатах IQ по тесту Стенфорд-Бине был на 13 баллов выше, чем у европейцев неевреев (п=110 и 689, соответственно) (Веге, 1924). В 1940-х гг. Нарди (Nardi, 1948) сообщал об IQ 12-летних еврейских подростков (п=1.210) в 110 баллов по тесту Стенфорд-Бине, а в 1950-х гг. Левинсон (Levinson, 1957) также с помощью теста Стенфорд-Бине оценил IQ 12-летних еврейских подростков (п=2.083) в 109 баллов. Хернстайн и Мюррей (Herrnstein, Murray, 1994) сообщают о том, что в их Национальном лонгитюдном исследовании молодежи IQ еврейских подростков составил 112,6 баллов, а по данным самого последнего исследования IQ в национальной репрезентативной выборке взрослых евреев (п=150) составил 107,5 баллов (Lynn, 2004). Также сообщалось о высоком IQ еврейских детей в Великобритании. В 1920-х гг. Девис и Хьюдж (Davies, Hughes, 1927) нашли, что у 8-14-летних еврейских детей в Лондоне (n=1.081) IQ составляет 110 баллов в сравнении со 100 баллами детей британцев неевреев. В 1960-х гг. у 10-летних еврейских детей в Глазго (п=907) величина IQ была оценена в 117,8 баллов в сравнении с шотландскими детьми из того же города (Vincent, 1966). Однако, это значение нельзя соотносить с британскими детьми в целом, так как IQ детей в Глазго равен 93,7 баллам относительно среднего национального значения 100 (Lynn, 1979). Чтобы сравнить среднее значение IQ еврейских детей в Глазго с IQ белых британцев неевреев, мы должны вычесть 6,3 балла IQ из их оценки, что даёт средний IQ в 111,5 баллов. Таким образом, средние оценки IQ евреев в Соединенных Штатах и Великобритании лежат между 107 и 115 баллами, и таким образом выше 103 баллов евреев-ашкенази в Израиле. Возможные объяснения этого состоят в следующем. Лишь незначительное число американских и британских евреев эмигрировало в Израиль. Большинство евреев-ашкенази в Соединенных Штатах и Великобритании бежало от преследований из России и Восточной Европы в период между 1880 и 1914 гг. и из Германии с 1933 до 1939 г. Кажется вероятным, что те, кто предвидел опасность дальнейшего пребывания в стране и были в состоянии организовать эмиграцию, были более интеллектуальными. Те же, кто остался в России и Восточной Европе, вероятно, были несколько менее интеллектуальными. Это они эмигрировали в Израиль после Второй Мировой Войны, чтобы избежать преследований и бедности, и это их IQ немного ниже, чем у евреев-ашкенази в Соединенных Штатах и Великобритании. Другим фактором является то, что многие из предполагаемых европейских евреев вовсе ими не являются, но объявили себя евреями, чтобы получить разрешение на выезд из Советского Союза (Abbink, 2002).
110
Ричард Линн
Вторая проблема, касающаяся интеллекта евреев, состоит в следующем. Все евреи имеют общее происхождение. Так почему же интеллект евреев-ашкенази приблизительно на 12 баллов IQ выше, чем у восточных евреев? Вероятно, может быть два ответа на этот вопрос. Во-первых, несмотря на строгие еврейские запреты на экзогамию, всегда имелось некоторое число смешанных браков и половых связей между евреями и неевре- ями, живущими в одной местности. Даже небольшой доли экзогамии во многих поколениях было достаточно, чтобы внести существенные пропорции нееврейских генов в еврейский генофонд. Эффекты этого заметны у европейских евреев, некоторое количество которых имеют светлые волосы и голубые глаза. В результате этого евреи-ашкенази в Европе должны были получить значительную долю присущих европейцам генов, обуславливающих более высокий интеллект, а восточные евреи на Ближнем Востоке и в Северной Африке воспринять от автохтонного населения этих регионов значительную долю генов, обуславливающих более низкий интеллект. Другой фактор, который вероятно работал в сторону увеличения интеллекта евреев-ашкенази в Европе и Соединенных Штатах по сравнению с восточными евреями это то, что евреи ашкенази больше подвергались преследованиям. Евреи Юго-Западной Азии и Северной Африки в течение многих столетий меньше подвергались притеснениям. Они испытывали некоторые притеснения, но не в такой степени, как евреи-ашкенази в Европе. Этого оказалось достаточно, чтобы поднять их IQ до 91 балла в сравнении с 84 баллами у других аборигенов Южной Азии и Северной Африки.
Различие в 12 баллов IQ между евреями-ашкенази и восточными евреями в Израиле почти наверняка в некоторой степени обусловлено генетически. Генетический анализ, выполненный Хаммером, Реддом и Вудом (Hummer, Redd, Wood, 2000) показал, что все евреи в определённой мере генетически близки (за исключением эфиопских евреев) вследствие их общего происхождения с Ближнего Востока, но европейские и восточные евреи образуют две генетические семьи: европейские евреи с небольшим генетическим родством с европейскими неевреями и восточные евреи с некоторой долей генетической близости с аборигенами Юго-Западной Азии и Северной Африки.
Г(1Ш7
ЛШИГШ140Г0-И0СТ0Ч410Н ШП
Интеллект аборигенов Юго-Восточной Азии
Аборигены Юго-Восточной Азии в Соединенных Штатах и Нидерландах
Размер мозга аборигенов Юго-Восточной Азии
Генетические и средовые детерминанты IQ аборигенов Юго-Восточной Азии
К аборигенами Юго-Восточной Азии принадлежит автохтонное население Бирмы, Таиланда, Камбоджи, Вьетнама, Малайзии, Индонезии, Филиппин и о-ва Борнео. В классической антропологии они именовались малайцами (Morton, 1849; Coon, Gam, Birdsell, 1950) или индонезийскими малайцами (Cole, 1965). Их расовая особенность была подтверждена генетическим анализом, выполненным Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994), согласно результатам которого эти народы составляют отдельный генетический «кластер». Они имеют генетическое родство с аборигенами Восточной Азии, с которыми до некоторой степени смешаны, но нос у них не такой плоский и эпикантус менее заметен.
Интеллект аборигенов Юго-Восточной Азии
Показатели IQ выборок аборигенов Юго-Восточной Азии из пяти стран представлены в Таблице 7.1.
В строках 1-4 даны величины IQ для Индонезии. В строке 1 приведён IQ в 86 баллов для детей из города Бандунг на о-ве Ява. В строке 2 дан IQ в 87 балла для детей и подростков из двух деревень в центральной Яве, в строке 3 — в 87 баллов для детей из семей работников чайных плантаций. В строке 4 приведён IQ 87 для детей из северной Джакарты. В строке 5 приведён IQ 90 для детей народности Лао, проживающих в деревне но «не из семей, живущих в крайней бедности». В строке 6 дан IQ 88 для матерей детей, чьи данные представлены в строке 5. В строке 7 представлен IQ в 89 баллов для малайцев из Малайзии, полученный в ходе стандартизации
112
Ричард Линн
Таблица 7.1. IQ аборигенов Юго-Восточной Азии

стандартных прогрессивных матриц. В строке 8 приведён IQ 85 для малайцев — студентов Международного Исламского университета в г. Куала-Лумпур в сравнении со студентами университетов в Германии, России и в Соединенных Штатах. В строке 9 представлен IQ филиппинцев в 86 баллов, определявшийся у школьников в Маниле. В строке 10 представлен IQ 93 для 13-летних малайцев из школы в Сингапуре. В строке 11 дан IQ 91 для школьников изтаиландской провинции Чон Бури — сельскохозяйственной области на Восточном побережье. Показатели IQ лежат в диапазоне от 86 до 93 баллов с медианой 87.
Аборигены Юго-Восточной Азии в Соединенных Штатах и Нидерландах
Величины IQ выходцев из Юго-Восточной Азии в Соединенных Штатах и Нидерландах представлены в Таблице 7.2.
В строке 1 приведён IQ в 96 баллов по данным раннего исследования выборки филиппинских детей на Гавайях, протестированных с помощью лабиринтов Портеуса. В строке 2 дан IQ 89 для выборки филиппинцев из Гонолулу (данные были собраны Смитом в 1924 и 1938 гг. (Smith, 1942)). В строке 3 представлен IQ 91 для филиппинских детей на Гавайском острове Кауаи. В строке 4 дан IQ 93 для выборки филиппинцев на Гавайях, полученный по математическому субтесту теста STAS. В строке 5 приведён IQ
Расовые различия в интеллекте
113
Таблица 7.2. IQ аборигенов Юго-Восточной Азии в Соединенных Штатах и Нидерландах

в 87 баллов для выборки филиппинцев в Соединенных Штатах, вычисленный Флинном (Flynn, 1991). В строке 6 дан IQ 92 для выборки индонезийцев — иммигрантов второго поколения в Нидерландах. В строке 7 приведён вычисленный Флинном (Flynn, 1991) IQ в 94 балла для выборки учащихся американской городской полной средней школы, большинство испытуемых были вьетнамцами. Вербальный IQ испытуемых этой выборки, оцененный с помощью словарных шкал Милл Хилл, составил 87 баллов. Вероятно, это значение немного занижено относительно их IQ для невербального рассуждения, так как многие из испытуемых не говорили свободно по-английски. Медианное значение по семи исследованиям составило 93 балла, что немного выше IQ аборигенов Юго-Восточной Азии в странах их исконного проживания (87 баллов). Возможно, это различие объясняется отчасти тем, что при миграции в Соединенные Штаты и в Нидерланды происходил некоторый отбор. Другим возможным фактором является то, что у выходцев из Юго-Восточной Азии в Соединенных Штатах и Нидерландах более высокий уровень жизни и лучшее питание, чем у местного населения стран Юго-Восточной Азии.
Размер мозга аборигенов Юго-Восточной Азии
Результаты исследований различий размера мозга между европейцами и аборигенами Юго-Восточной Азии приведены в Таблице 7.3.
В строке 1 даны результаты, вычисленные Гулдом (Gould, 1981) на материале коллекции черепов, собранных в девятнадцатом столетии американским врачом Сэмюэлем Мортоном (Morton, 1849). Число черепов было весьма невелико, а именно 18 черепов аборигенов Юго-Восточной
114
Ричард Линн
Таблица 7.3. Различия размера мозга (см3) европейцев и аборигенов Юго-Восточной Азии

Азии и 52 европейцев, ввиду чего эти данные не имеют большого веса и представлены здесь главным образом из исторического интереса. В строке 2 приведены данные для шести популяций аборигенов Юго-Восточной Азии в сравнении с девятью популяциями европейцев, различие составляет 37 см3. Стандартные отклонения представлены Билсом и соавт. (Beals et al. 1984). Количества наблюдений не приведены, но эти выборки являются частью коллекции, численностью приблизительно в 20 тысяч черепов и, как можно предполагать, в сумме составляли несколько тысяч. Несмотря на небольшой размер выборки Мортона и вопреки обвинению его Гулдом, в том, что он подогнал свои результаты «в пользу» европейцев, результаты Мортона весьма близки к полученным в более поздней работе Смита и Билса. В строке 3 представлено намного большее различие мозга по измерениям 190 выборок европейцев и 20 выборок аборигенов ЮгоВосточной Азии. Таким образом, все три набора данных указывают на меньший размер мозга у аборигенов Юго-Восточной Азии в сравнении с европейцами, что согласуется с показателями их IQ.
Генетические и средовые детерминанты IQ аборигенов Юго-Восточной Азии
Величина IQ выходцев из Юго-Восточной Азии в Соединенных Штатах выше (93 балла), чем у местного населения Юго-Восточной Азии (87 баллов). Это различие можно приписать более высокому уровню жизни в Соединенных Штатах: лучшему питанию, образованию и социальному обеспечению. В результате разрыв величины IQ между выходцами из ЮгоВосточной Азии и европейцами уменьшается примерно вдвое. Но разница в 7 баллов IQ остаётся и тогда, когда выходцы из Юго-Восточной Азии и европейцы воспитываются и живут приблизительно в одинаковых условиях. Это говорит о том, что генетические факторы вносят вклад в различие интеллекта этих двух рас. Меньший средний размер мозга аборигенов Юго-Восточной Азии по сравнению с европейцами также говорит о генетическом различии.
Г(1Ш о
ШМИИЙШ ЛШИГШ1
Интеллект австралийских аборигенов
Гибриды австралийских аборигенов с европейцами
Интеллект по Пиаже
Пространственная память
Размер мозга
Генотипный интеллект
Интеллект населения Новой Гвинеи
Заключение
Австралийские аборигены являются автохтонным населением Австралии. Они известны также как австралиды и давно признаны отдельной расой в классической антропологии. К примеру, в таксономии Куна, Гарна и Бёрдселла (Coon, Gam, Birdsell, 1950) они являются одной из семи главных рас. Они имеют характерный профиль групп крови: приблизительно у 73 процентов присутствует группа 0 (у европейцев её частота немного меньше 50%), у остальных 27 процентов группа А, а группа В практически не встречается. Расовая особенность австралийских аборигенов была подтверждена Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994) с помощью генетического анализа, по результатам которого австралийские аборигены вместе с туземным населением Новой Гвинеи образуют отдельный генетический «кластер». Причина близкого генетического родства австралийских аборигенов и автохтонных новогвинейцев в том, что приблизительно 60 тысяч лет назад предки австралийских аборигенов мигрировали из Новой Гвинеи в Австралию (Bradshaw,
. Мигрировавшие отделились от оставшихся на Новой Гвинее, обитающих ныне во внутренних горных районах острова. Близко родственными австралийским аборигенам были также вымершие ныне тасманийцы. Последний чистокровный тасманиец умер в 1876 г., но сохраняется небольшое число расово-смешанных тасманийцев.
Было оценено, что до появления европейцев в Австралии там проживало около 300 тысяч аборигенов. Их количество значительно сократилось после колонизации Австралии европейцами, отчасти вследствие передавшихся им от европейцев болезней, к которым у них не было иммунитета, а частично в результате истребления их европейцами. Во второй половине
116
Ричард Линн
двадцатого столетия количество аборигенов по переписям населения 1961, 1971 и 1981 гг. составляло приблизительно 106.000, 139.000 и 171.000 чел. Быстрое увеличение численности явилось результатом высокой рождаемости и сокращения младенческой и детской смертности.
Во второй половине двадцатого столетия имелось три группы австралийских аборигенов. Первая обитала в государственных резервациях преимущественно на Севере и в центре Австралии. Вторая группа жила в предместьях провинциальных городов и животноводческих хозяйств. Третья проживала в более крупных городах. И вторая, и третья группы обычно посещали те же школы, что и европейцы. Многие аборигены второй и третьей групп имееют некоторую долю европейских предков, в то время как в резервациях проживают в основном чистокровные аборигены.
Интеллект австралийских аборигенов
Первая попытка оценить интеллект австралийских аборигенов была предпринята Гальтоном (Galton, 1869). На основе сообщений путешественников о достижениях аборигенов он оценил их интеллект примерно на три «деления» ниже, чем у англичан. Одно деление шкалы Гальтона эквивалентно 10,4 баллам IQ. Следовательно, в терминах шкалы IQ его оценка интеллекта австралийских аборигенов составила 68,8 баллов. Дальнейшие исследования интеллекта австралийских аборигенов с помощью тестов интеллекта показали, что это оценка была весьма точной. Результаты этих исследований приведены в Таблице 8.1.
В строке 1 представлены результаты первого исследования, в котором Портеусом была получена оценка IQ в 66 баллов с помощью его теста лабиринтов. Тест представляет собой серию нарисованных на бумаге лабиринтов возрастающей сложности, путь через которые нужно отметить карандашом. Умственный возраст испытуемых измеряется путём соотнесения с уровнем достижений среднего ребёнка соответствующего хронологического возраста. Тест лабиринтов, позже включённый в тесты Векслера, даёт оценку фактора g и способности к визуализации. Средний умственный возраст взрослых аборигенов в этом исследовании составил 10,5 лет, что соответствует приблизительно 66 баллам IQ. В строке 2 даны результаты другого исследования, где были использованы лабиринты Пор- теуса для тестирования выборки аборигенов из г. Ла Г ранде Бей в СевероЗападной Австралии. Умственный возраст мужчин оказался равным 10,5, а женщин — 8,6 лет. Средний умственный возраст для обоих полов составил 9,55 лет, что эквивалентно IQ в 59 баллов. В строке 3 приведён сходный результат, полученный Портеусом у взрослых в миссии Бигл Бей в
Расовые различия в интеллекте
117
Таблица 8.1. Значения IQ австралийских аборигенов

области Кимберли; умственный возраст аборигенов оказался равным 9,35 лет, что эквивалентно 58 баллам IQ. В строке 4 представлен IQ 69, полученный по результатам двух тестов визуализации (.Alexander passalong и Fergusson Form Boards). В строке 5 приведён IQ 70 по данным исследования аборигенов племени Вайлбири в Центральной Австралии, проведённого Портеусом и Грегором в 1960-х гг. В строке 6 дан IQ в 58 баллов для выборки учеников начальной школы в г. Манингрида на Северных Территориях. В строке 7 представлен IQ 74 для выборки взрослых, умственный возраст которых был оценен в 11,8 лет. В строках 8 и 9 приведены IQ в 62 и 64 балла для двух выборок детей аборигенов, посещавших городские школы вместе с белыми детьми в Новом Южном Уэльсе.
В строке 10 дан IQ в 53 балла для выборки взрослых аборигенов, тестировавшихся с помощью цветных прогрессивных матриц. В строке 11 приведён вербальный IQ в 64 балла для 3- и 4-летних детей аборигенов, посещавших вместе с белыми детьми группы дошкольной подготовки в г. Бурке. В строке 12 дан вербальный IQ 52 для детей, посещавших школу миссии Хермансберг в Центральной Австралии. В строке 13 приведён IQ 58 для аборигенов, вычисленный относительно норм европейских детей в Новой Зеландии. В строке 14 приведено значение IQ в 62 балла, полученное
118
Ричард Линн
с помощью теста на рассуждение Spiral Omnibusв выборке 13-летнех детей аборигенов, посещавших школу в резервации в Квинсленде. В строке 15 дан вербальный IQ в 59 баллов для выборки детей аборигенов возраста 610 лет из г. Элис Спрингс в Центральной Австралии. В строке 16 приведён IQ в 60 баллов для способности к рассуждению для выборки взрослых (средний возраст 25 лет). В строке 17 приведён словарный IQ в 61 балл для выборки 4-летних детей.
Оценки IQ располагаются в пределах от 52 до 74 баллов. Медианное значение IQ по семнадцати исследованиям составило 62 балла, что представляется наилучшей оценкой среднего интеллекта австралийских аборигенов. Их вербальная способность была немного ниже способности к визуализации, медианные значения составляют 62 и 68 баллов, соответственно. Низкий интеллект австралийских аборигенов был подтвержден в исследовании, показавшем, что для них характерно длительное время реакции (Davidson, 1974).
Гибриды австралийских аборигенов с европейцами
Имеется ряд исследований интеллекта гибридов аборигенов с европейцами. Результаты приведены в Таблице 8.2.
Таблица 8.2. Показатели интеллекта гибридов австралийских аборигенов с европейцами

В строке 1 дан IQ 95 для первого из таких исследований, выполненного Портеусом в животноводческом хозяйстве миссии в Порт МакЛей в Южной Австралии. В строках 2 и 3 приведены результаты исследования, в котором сравнивались 19 аборигено-европейских гибридов с европейскими 5летними детьми, посещавшими те же школы в Новом Южном Уэльсе. Относительно IQ 100 европейских детей аборигено-европейские гибриды получили оценки IQ в 79 баллов в тесте PPVT (Словарный тест Пибоди в картинках) и 77 баллов в тесте ITPA (Иллинойский тест психолингвиста-
Расовые различия в интеллекте
119
ческих способностей). В строке 4 дан вербальный IQ в 69 баллов для 13 нечистокровных аборигенов возраста 6-12 лет, что немного выше 64 баллов у 40 чистокровных аборигенов, тестировавшихся в том же исследовании. Величина IQ визуализации в 95 баллов, приведённая в строке 1, намного выше значений вербального IQ в 79,77 и 69 баллов, представленных строках 2,3 и 4, соответствено. Все величины IQ гибридов аборигенов с европейцами, приведённые в Таблице 8.2, были выше медианы значений IQ чистокровных аборигенов, представленных в Таблице 8.1. Это могло произойти из-за примеси европейских генов, повышающих интеллект аборигенов. Альтернативным объяснением может являться то, что аборигено-европейские гибриды обычно воспитываются в лучших условиях, что касается уровня жизни и питания. Ни в одном из этих исследований не даётся оценки доли европейских предков у нечистокровных аборигенов.
Интеллект по Пиаже
Интеллект австралийских аборигенов был оценен тестами Пиаже вдобавок к обычным тестам интеллекта. Эта работа была выполнена в рамках теории развития интеллекта детей, разработанной швейцарским психологом Жаном Пиаже. Согласно этой теории, дети проходят четыре стадии когнитивного развития. Первая из них — сенсо-моторная стадия, на которой ребенок узнает о свойствах объектов, пространстве, времени и причинности. Приблизительно в два года дети переходят к дооперацио- нальной стадии, на которой они овладевают языком и отвлечёнными понятиями, но ещё не способны понять логические принципы. Эта стадия длится приблизительно до шестилетнего возраста. В Западных обществах примерно к семи годам дети переходят к стадии конкретных операций, когда они могут осознавать логические принципы, но только на конкретном материале. Примерно к двенадцатилетнему возрасту европейские дети переходят к четвертой и заключительной стадии формальных операций, когда они становятся способными мыслить логически в терминах общих принципов, оторванных от конкретных примеров. В ряде исследований было показано, что сформированность понятий, измеряемая с помощью задач Пиаже, высоко коррелирует с величинами IQ, оцениваемыми стандартными тестами интеллекта (Jensen, 1980).
Методика, использованная в исследованиях интеллекта детей австралийских аборигенов «по Пиаже», состояла в установлении того, достигают ли они стадий когнитивного развития в том же самом возрасте, что и европейские дети. Эти исследования в большинстве случаев оценивали возраст, в котором дети аборигенов достигают стадий конкретных операций
120
Ричард Линн
и формальных операций. Стадия конкретных операций наиболее часто оценивалась тестами, определяющими, сформировалось ли у ребенка понятие «сохранения». Это — понимание принципа, что объём и вес вещества остаются теми же (то есть, «сохраняются»), когда изменяется его форма. Стандартный тест для проверки способности к пониманию принципа сохранения количества состоит в том, что тестирующий переливает воду или какое-либо другое вещество (например, бусинки) из стеклянного стакана в высокий тонкий сосуд. Ребенка спрашивают, осталось ли количество воды или другого вещества тем же самым. Маленькие дети обычно полагают, что воды или другого вещества больше в высоком тонком сосуде, сосредотачиваясь, очевидно, на его большей высоте и игнорируя меньшую ширину. Когда дети улавливают, что объём остаётся тем же вне зависимости от формы сосуда — они достигли понимания принципа сохранения.
Первые исследования понимания взрослыми австралийскими аборигенами принципа сохранения были выполнены де Лемос (de Lemos, 1969, 1979). Она показывала 12 женщинам-аборигенкам два стакана сахара. Один, высокий и тонкий, был наполнен чашкой сахара, в то время как другой, широкий и низкий, был наполнен половиной чашки сахара. Женщинам дали на выбор два этих стакана и восемь из них выбрали широкий и низкий стакан, в котором сахара было меньше. Автор резюмировала: «Согласно теории Пиаже, это понятие является базовым для всего логического мышления и, следовательно, это отставание указывает на более низкий уровень функционирования интеллекта в сравнении с обычно достигаемым в европейской культуре» (1969, стр. 15). Отсутствие понимания принципа сохранения двумя третями этих взрослых женщин даёт основание полагать, что они находились на уровне умственного развития примерно 8-летних белых детей, что говорит о том, что их IQ должен составлять около 50 баллов. Де Лемос также нашла, что межрасовые гибриды аборигенов и белых лучше выполняли тест на сохранение, чем чистокровные аборигены, хотя и не так хорошо, как белые (De Lemos, 1969).
Во втором исследовании Де Лемос (De Lemos, 1969) давала задачи Пиаже на сохранение 38 чистокровным детям аборигенов и 34 имевшим приблизительно одну восьмую долю европейских предков. Она описала условия их жизни следующим образом: «Не было никаких явных различий в условиях существования детей нечистокровных и чистокровных аборигенов... которые формировали единое интегрированное сообщество, дети воспитывались в условиях одной миссии и посещали одну и ту же школу» (стр. 257). Нечистокровные дети аборигенов получали значительно более высокие оценки в задачах, чем чистокровные аборигены, но не было возможности оценить результаты количественно в баллах IQ. Де Лемос заключила, что поскольку эти две группы жили в одинаковых условиях, только генетическая гипотеза может объяснить различие.
Расовые различия в интеллекте
121
Исследование Дасена (Dasen, 1973) дало сходные результаты. Он давал задачи Пиаже на сохранение двум выборкам аборигенов в Центральной Австралии (55 детей и 90 взрослых) и 80 белым детям в Канберре. Все дети аборигенов посещали школы. Белые дети достигли этой стадии в среднем в 8-летнем возрасте, в то время как дети аборигенов приблизительно в 15 лет. Двадцать три процента взрослых аборигенов достигали стадии, достигавшейся европейскими детьми в среднем в возрасте 7-8 лет. Дасен заключил (Dasen, 1973, стр. 92), что «у значительной доли аборигенов конкретные операциональные понятия не развиваются вообще, даже в взрослом состоянии». Результаты указывают на то, что IQ аборигенов составлял около 55 баллов. Другая часть исследования проведена Дасеном среди приблизительно 30 чистокровных детей аборигенов и 30 имевших смешанную родословную. Он обнаружил, что нечистокровные аборигены выполняли задачи лучше чистокровных аборигенов, но не намного.
Ещё одно исследование формирования понятия сохранения у детей австралийских аборигенов было проведено Сигримом и Лендоном (Seagrim, Lendon, 1980). Они нашли, что у 10 процентов 7-8-летних, 35 процентов 9-10-летних и у 70 процентов 12-летних детей сформировалось это понятие. Таким образом, 12-летние аборигены находятся приблизительно на том же ментальном уровне, что и 7-8-летние белые дети. Это позволяет дать им оценку IQ приблизительно в 60 баллов.
На основе своей работы со швейцарскими детьми Пиаже заключил, что все, кроме умственно отсталых, достигают всех стадий когнитивного развития к тому времени, когда становятся взрослыми. Исследования австралийских аборигенов показали, что это неверно, и что многие из них никогда не достигают последней стадии логического мышления. Эти исследования, демонстрирующие отставание в развитии интеллекта по Пиаже, дают дальнейшее подтверждение низкого интеллекта австралийских аборигенов.
Пространственная память
В замечательном исследовании Кеаринс (Kearins, 1981) было обнаружено, что у детей аборигенов пространственная память намного лучше, чем у европейцев. В этом исследовании 132 детям аборигенов возраста 716 лет и такому же числу детей белых австралийцев предъявляли различные тесты пространственной памяти. Общая схема тестов состояла в следующем: раскладывали 20 объектов и просили ребенка посмотреть на них в течение 30 секунд и попытаться запомнить их расположение. Затем объекты убирали и просили ребенка вновь разложить их на те же места. Во всех
122
Ричард Линн
задачах дети аборигенов показывали лучшие результаты, чем белые. Их общее преимущество выражается оценкой IQ пространственной памяти в 119 баллов. Кеаринс утверждала, что наиболее вероятное объяснение наличия у аборигенов хорошей пространственной памяти состоит в том, что она развилась у них вследствие малого количества ориентиров в пустынях Центральной Австралии. Кочевое аборигенное население должно было замечать и запоминать местоположение таких ориентиров, чтобы построить в уме пространственные планы окрестностей с целью найти путь домой с охоты. Для подтверждения этого она протестировала выборку городских аборигенов, чьи семьи на протяжении жизни нескольких поколений жили в городах. Испытуемые этой выборки так же хорошо выполняли тесты на пространственную память, как и жители пустыни. По утверждению Кеаринс это указывает на то, что среда обитания не «несёт ответственности» за хорошую пространственную память аборигенов и согласуется с представлением о генетической обусловленности этой способности.
Результаты Кеаринс, однако, оспаривались. Дринкуотер (Drinkwater,
сравнил способности 12-летних детей (22 аборигена и 22 белых) с помощью сходных тестов и нашёл, что обе группы выполняли эти тесты на одинаковом уровне. Но участвовавшие в этом исследовании аборигены происходили из прибрежной области, где хорошая пространственная память, требующаяся согласно теории Кеаринс, не будет необходима и не должна бы развиться. Тем не менее примечательно, учитывая низкий общий интеллект аборигенов, что они выполняли тесты на пространственную память так же хорошо, как белые. Харрис обнаружил (Harris, 1977 — неопубликованная докторская диссертация), что аборигены из пустыни справляются с этой задачей хуже белых. Кнапп и Сигрим (Knapp, Seagrim, 1981) также нашли, что аборигены из пустыни справляются с тестами хуже белых. К сожалению, авторы не представили данные в таком виде, который бы позволил вычислить величину преимущества белых. Несмотря на эти отрицательные результаты, данные Кеаринс о пространственной памяти аборигенов остаются впечатляющими и заслуживают дальнейшего исследования австралийскими психологами. Хорошая пространственная память аборигенов, если это будет подтверждено, имеет параллель в хорошей визуальной памяти эскимосов, о которой сообщает Кляйнфельд (Kleinfeld, 1971), объясняющая её адаптацией к существованию в промёрзшей тундре, имеющей мало ориентиров и в этом отношении подобной пустыням Австралии (см. Главу 11).
Расовые различия в интеллекте
123
Размер мозга
Данные семи исследований размера мозга австралийских аборигенов в сравнении европейцами приведены в Таблице 8.3.
Таблица 8.3. Размер мозга (см3) австралийских аборигенов и европейцев (численность выборок в круглых скобках)

В строке 1 приведены данные Мортона, уточнённые Гулдом (Gould,
. Во всех исследованиях показано, что размер мозга австралийских аборигенов меньше, чем у европейцев. Эти результаты подтверждены в исследовании 6-11-летних детей аборигенов из начальной школы (п=281) Эдвардса и Крэд дока (Edwards, Craddock, 1973), обнаружившими, что средняя главная окружность головы детей соответствовала 10-му процентилю белых в Соединенных Штатах и Австралии. Главная окружность головы является приближённой оценкой размера мозга. Поскольку размер мозга — важная детерминанта интеллекта (Vernon и др., 2000), меньший средний размер мозга аборигенов можно считать отчасти ответственным за их более низкий IQ. Клекамп и соавторы сообщали (Klekamp et al., 1987), что у австралийских аборигенов визуальная кора правого полушария занимает большую площадь, чем у европейцев. Правое полушарие ответственно за пространственные, а левое — за вербальные способности, так что наличие у аборигенов относительно более развитого правого полушария согласуется с их хорошей пространственной памятью, что было найдено Кеаринс (Kearins, 1981 — см. параграф 4), предложившей теорию, что у аборигенов развилось относительно большее правое полушарие мозга и визуальная кора для того, чтобы решать визуальные и пространственные задачи, с которыми сталкиваются кочевые народы в лишённом примет ландшафте пустыни.
124
Ричард Линн
Генотипный интеллект австралийских аборигенов
На наличие некой генетической составляющей низкого интеллекта австралийских аборигенов указывают восемь групп фактов.
Во-первых, наиболее удовлетворительным методом оценки степени, в которой генетические факторы обуславливают низкий интеллект аборигенов, было бы кросс-расовое исследование, в котором аборигенные младенцы усыновлялись бы белыми семьями. Средовая теория предсказывает, что у них будет такой же IQ, как у белых, в то время как согласно генетической теории их IQ останется таким же, как у аборигенов. Если их средний IQ окажется промежуточным между значениями аборигенов и белых, можно будет заключить, что вовлечены и генетические, и средовые факторы. В единственном исследовании такого типа (Dasen, de Lacey, Seagrim, 1973) исследовались дети аборигенов (п=35), усыновлённые белыми супругами в г. Аделаида и его окрестностях. Семнадцать из этих детей были аборигенами наполовину, из остальных 7 были чистокровными аборигенами, 2 — на три четверти, 4 — на одну четверть, один — на одну восьмую и у 4 родословная была неизвестна. В среднем они были аборигенами примерно наполовину. Средний возраст усыновлённых составлял 18 месяцев. Между 5 и 13 годами детей тестировали с помощью шести тестов: четыре теста Пиаже, тест «переклассификации» Никсона и словарный тест Пибоди в картинках (PPVT). Приводятся результаты для усыновлённых аборигенов и групп сравнения, состоявших из европейцев и чистокровных аборигенов из Центральной Австралии. Ни один из результатов тестов не может быть точно оценен количественно, так как они приведены в виде графиков. Насколько можно из них понять, в двух тестах Пиаже (сохранение количества и веса) результаты аборигенов находились примерно в середине диапазона между европейцами и чистокровными аборигенами. Поскольку усыновлённые аборигены были полукровками, полученные данные являются ожидаемыми и говорят о том, что опыт жизни в приёмной семье не имел благоприятного эффекта. В третьем тесте (сохранение горизонтальности) усыновлённые аборигены показали несколько худшие результаты в сравнении с европейской группой, но существенно лучшие, чем чистокровные аборигены. Четвертый, пятый и шестой тесты, описанные как меры «сериации длины», «переклассификации» (ни один из этих терминов не объяснён) и PPVT, аборигены выполняли примерно так же, как европейская группа сравнения. Таким образом, несмотря на то, что успешность выполнения разных тестов усыновлёнными детьми, являющимися отчасти аборигенами, была различна, в целом они
Расовые различия в интеллекте
125
уступали европейским детям. Это согласуется с замечанием авторов, что «о большинстве детей их родители сообщали, что их успеваемость в школе была ниже средней; чаще всего говорилось об особых трудностях в математике» (стр. 98). В то время как приёмные аборигены-полукровки выполняли задания хуже европейцев, они, по-видимому, выполняли их лучше, чем аборигены-полукровки, воспитываемые их биологическими родителями. Следовательно, результаты говорят о том, что как генетические, так и средовые факторы ответственны за низкий интеллект аборигенов. Следует отметить, что средний возраст детей во время тестирования составлял приблизительно 8 лет и что в американском исследовании чернокожих детей, усыновлённых белыми родителями (Weinberg, Scarr, Waldman, 1992), было обнаружено, что в возрасте 7 лет они имели средний IQ в 95 баллов, но к 17 годам он понизился до 89. Это говорит о том, что IQ малолетних чернокожих детей благодаря усыновлению возрастает, но затем прирост постепенно исчезает в старшем подростковом возрасте (см. Главу 4).
Во-вторых, медианное значение IQ аборигенов по результатам 16 исследований, приведённым в Таблице 8.1, составляет 62 балла, в то время как медианный IQ аборигено-европейских гибридов по данным четырёх исследований, приведённым в Таблице 8.2, равен 78 баллам. Данные о более высоком IQ гибридов согласуются с генетической гипотезой, предсказывающей, что величина IQ гибридов должна быть промежуточной между значениями IQ родительских рас. Однако может статься, что гибриды жили в лучших условиях и их более высокий IQ может быть объяснён средовыми влияниями.
В-третьих, все дети аборигенов в исследованиях, результаты которых приведены в Таблицах 8.1 и 8.2, посещали школы, а в трёх исследованиях (строки 6, 7 и 9 Таблицы 8.1) дети аборигенов посещали школы вместе с белыми детьми, так что их низкие IQ не могут быть приписаны недостатку возможности приобрести ментальные навыки (оцениваемые тестами интеллекта) или коренными различиями среды обитания.
В-четвертых, низкие показатели IQ аборигенов фиксируются и у детей 4-летнего возраста (Таблица 8.1, строки 11 и 17), что также говорит о невозможности приписать их неадекватному обучению.
В-пятых, низкие значения IQ аборигенов наблюдаются в широком диапазоне способностей, включая рассуждение, вербальное понимание, словарную способность, пространственную способность (измеренную лабиринтами Портеуса) и в задачах сохранения Пиаже. Это показывает, что их низкий IQ не может быть объяснён смещённостью какого-либо отдельного теста.
В-шестых, не наблюдается тенденции роста показателей IQ аборигенов за примерно полувековой период от первых двух исследований, выполненных приблизительно в 1930 г. (давших оценки IQ в 66 и 59 баллов)
126
Ричард Линн
и двух исследований, проведённых в 1980-х и 1990-х гг. (давших оценки IQ в 60 и 61 балл, см. Таблицу 8.1), несмотря на улучшение условий жизни аборигенов вследствие повышения благосостояния и улучшения медицинского обслуживания.
В-седьмых, если интеллект некоторых аборигенов снижен вследствие неблагоприятных условий существования, то наиболее вероятной причиной этого будет плохое питание. Распространенность недоедания среди аборигенов была исследована в двух работах. В первой из них Эдвардс (Edwards, 1970) обследовал 82 аборигена-дошкольника в Новом Южном Уэльсе и обнаружил, что 31 процент из них недоедал. В другой работе среди детей аборигенов (п=281) было выявлено, что доля недоедающих составляет 21 процент (Edwards, Craddock, 1973). Неполноценное питание в младенчестве отрицательно влияет на интеллект, но по данным этих двух исследований, вместе взятых, доля недоедающих аборигенов составляла только около 25 процентов. Эдвардс и Крэддок (Edwards, Craddock, 1973) провели тестирование интеллекта 29 недоедавших и 29 нормально питавшихся детей аборигенов возраста 6-10 лет и нашли, что IQ недоедавших детей был в среднем на 8 баллов ниже, чем у получавших нормальное питание. Поскольку недоедают приблизительно 25 процентов аборигенов, недоедание должно снижать IQ всей популяции примерно на 2 балла. Это позволяет предположить, что недостаточное питание лишь в незначительной степени ответственно за низкий IQ аборигенов.
В-восьмых, малый размер мозга аборигенов — главная неврологическая и генетическая детерминанта их низкого интеллекта. Размер мозга влияет на интеллект и в значительной мере наследуется. Размер мозга может быть снижен вследствие недостаточного питания, но поскольку только около 25 процентов аборигенов недоедают, малый размер их мозга определён в значительной степени генетически и является важной детерминантой их низкого интеллекта.
Интеллект новогвинейцев
Аборигены Новой Гвинеи населяют её внутренние горные районы, куда они были вытеснены меланезийцами и аборигенами Юго-Восточной Азии индонезийского происхождения в течение последних 3.000 лет или около того. Ныне население Новой Гвинеи состоит из местных аборигенов, выходцев с островов Тихого океана, выходцев из Юго-Восточной Азии и гибридов. Обычно исследователи не указывают, к какой из этих групп принадлежат их выборки, и это должно быть выведено из их местоположения. Было проведено два исследования интеллекта аборигенов Новой
Расовые различия в интеллекте
127
Гвинеи при помощи тестов интеллекта. В первом исследовании (McElwain, Kearney, 1970) величина IQ у 26 мужчин возраста 20-29 лет была оценена с помощью невербального Квинслендского теста в 65 баллов в сравнении с белыми австралийцами. В другом исследовании (Berry, 1971) выборка взрослых (п=70) была протестирована с помощью цветных прогрессивных матриц. Показатели новогвинейцев находились значительно ниже первого процентиля взрослых британцев, их IQ может быть оценен в приблизительно в 62 балла — такой же, как у австралийских аборигенов.
Было выполнено три исследования интеллекта новогвинейских аборигенов по Пиаже. Первое из них выполнил Принс (Prince, 1968) на большой выборке (п=2.700) школьников и студентов педагогического колледжа. Он пришёл к заключению, что новогвинейцы «демонстрируют ожидаемый паттерн стадий Пиаже, хотя стадия сохранения не достигается до значительно более позднего возраста, чем в западноевропейской культуре» (стр. 65). Даже студенты колледжа продемонстрировали «значительно худшее развитие по всем заданиям теста, требующим понятия сохранения» (стр. 64). В то время как принцип сохранения понимается приблизительно 85 процентами европейских 8-летних детей и практически всеми 12-летними, сохранение вещества понимали 22 процента новогвинейских 8-летних детей и 85 процентов 18-летних, а сохранение площади не понималось ни одним 8-летним и 50 процентами 18-летних. Эти результаты дают основание полагать, что 18-летние новогвинейские аборигены обладают умственным возрастом приблизительно 8-летних европейцев, что эквивалентно примерно 50 баллам IQ.
Второе исследование 432 детей и подростков в возрасте 6-19 лет (в средним 11 лет) было выполнено Келли (Kelly, 1977). Результаты были таковы: 31 процент испытуемых достиг понятия сохранения количества и ни один не достиг стадии формальных операций. Поскольку приблизительно 70 процентов европейских детей достигают понятия сохранения к возрасту 7 лет и все, кроме умственно отсталых, достигают стадии формальных операций к двенадцати годам, то, что к 7 годам лишь 31 процент испытуемых новогвинейской выборки достиг стадии конкретных операций и никто из них не достиг стадии формальных операций, свидетельствует о том, что их средний IQ составляет около 55 баллов. В этом исследовании новогвинейцев делили на посещавших и не посещавших школу. Среди мальчиков не было выявлено различия между двумя этими группами, что говорит о том, что позднее формирование понятия сохранения не объясняется отсутствием образования. Среди девочек результаты были лучше у посещавших школу, но это не обязательно является эффектом обучения. Для обучения в школе отбирали более интеллектуально развитых и две группы были из разных племён, проживающих в разных местах. Автор пишет: «Результаты мальчиков согласуются с часто повторяющимся
128
Ричард Линн
положением Пиаже, что те структуры, которые он описывает, не подвержены сколь-нибудь значительному влиянию со стороны школьного обучения» (Kelly, 1977, стр. 183).
Келли суммирует другое исследование Джонса, обнаружившего, что среди учащихся старших классов и студентов университета возраста 16-19 лет, отобранных в старшие классы или в колледж на основании конкурсного экзамена и имевших за плечами как минимум девять классов школы, 67 процентов были способны выполнять задачи на сохранение. Следовательно, остальные 33 процента имели умственный возраст средних 7-летних европейских детей. Эти результаты дают основание полагать, что в целом по группе умственный возраст испытуемых примерно соответствовал 10-летним европейцам и, таким образом, их IQ составлял приблизительно 63 балла. Результаты пяти исследований в достаточной мере согласуются и дают медианное значение IQ в 63 балла, что почти совпадает с 62 баллами австралийских аборигенов.
Заключение
Результаты тестов интеллекта, выявивших его низкий уровень у австралийских аборигенов, подтверждают наблюдения антропологов, описавших эти народы в конце девятнадцатого и первой половине двадцатого столетий и считавших, что австралийские аборигены имеют низкие умственные способности и являются уцелевшими примитивными людьми каменного века. Так, Вейк (Wake, 1872, стр. 80) писал, что «австралийские аборигены все ещё дети по своему общему умственному развитию». В первом десятилетии двадцатого века Клаатч опубликовал первую из ряда своих работ (Klaatsch, 1908, стр. 164), в которых было показано, что мозг аборигенов меньше европейского и пришёл к заключению, что «австралийские аборигены являются реликтом самого древнего типа человечества». Несколько лет спустя антрополог сэр Артур Кейт (Keith, 1922, стр. xi) писал, что австралийские аборигены «представляют собой изначальную расу, от которой произошли три большие современных расы: негроиды, европеоиды и монголоиды». Но во второй половине двадцатого столетия антропологи стали утверждать, что аборигены столь же интеллектуальны, как и европейцы. Так, А.Р.Е. (1960, стр. 714), в статье в Энциклопедии Britannica, описал примитивную культуру каменного века аборигенов, но продолжил это описание утверждением, что «эта материальная бедность не является результатом низкого интеллекта, но условий существования. Размер мозга аборигенов находится в пределах европейского диапазона, и нет никаких данных, чтобы говорить, что это не верно для их интеллекта». Ис
Расовые различия в интеллекте
129
пользование фразы «находится в пределах европейского диапазона» для размера мозга аборигенов говорит о том, что автору было хорошо известно, что средние размеры их мозга приходятся на нижний конец европейского диапазона, но ему очевидно хотелось это замолчать, в то время как утверждение о том, что аборигены столь же интеллектуальны как европейцы, можно, вероятно, приписать полному невежеству.
Джаред Даймонд идёт ещё дальше в своей книге «Ружья, микробы и сталь» («Guns, Germs, and Steel»).Он начинает с описания того, как во время его работы на Новой Гвинее туземец по имени Йали спросил его: «Почему это вы, белые люди, создали так много груза и привезли его на Новую Гвинею, а у нас, чёрных людей, мало собственного груза?» (стр. 14). «Груз» («cargo»)на новогвинейском жаргоне означает товары. Даймонд говорит, что написал эту книгу, чтобы ответить на этот вопрос. Он утверждает, что ответ кроется не в различиях интеллекта разных народов и что «новогвинейцы показались мне в среднем более интеллектуальными, чемсредний европеец или американец» (Diamond, 1998, стр. 20). Он не упоминает об исследованиях, выявивших низкий IQ этого народа с помощью тестов интеллекта и задач Пиаже.
гмш ттт остгаш тихого ошим
Интеллект новозеландских маори
Другие аборигены островов Тихого океана
Гибридное население Гавайских островов
Размер мозга
Средовые и генетические детерминанты интеллекта аборигенов островов Тихого океана
Аборигены островов Тихого океана — туземные народы многочисленных островов Тихого океана, главными из которых являются Новая Зеландия, группы островов Микронезии, Меланезии, Полинезии и Гавайев, и изолированный остров Пасхи. Эти острова оставались необитаемыми приблизительно до 6.000-1.000 лет до нашей эры, когда Микронезия, Меланезия и Западная Полинезия начали заселяться выходцами из ЮгоВосточной Азии. Только примерно к 650 г. нашей эры все основные острова Полинезии были заселены. Последней из тихоокеанских островов была колонизирована Новая Зеландия, в которой приблизительно к 800 г. нашей эры обосновались выходцы из Полинезии — предки современных маори. В классической антропологии аборигены островов Тихого океана были признаны одной из семи главных рас Куном, Гарном и Бёрдселлом (Coon, Garn, Birdsell, 1950). Это было подтверждено Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994) в их генетической классификации, в которой микронезийцы, меланезийцы и полинезийцы выступают как «кластер». Аборигены островов Тихого океана — одна из малых рас численностью примерно 1,5 миллиона. В конце двадцатого столетия в Новой Зеландии проживало приблизительно 350.000 маори. Население Соломоновых островов составляет приблизительно 380.000 чел., примерно столько же проживает на островах Фиджи. Население Западного Самоа составляет приблизительно 170.000 чел., а о-вов Тонга — примерно 100.000 чел.
Расовые различия в интеллекте
131
Интеллект новозеландских маори
Аборигены островов Тихого океана, чей интеллект изучен лучше всего — это маори Новой Зеландии. Результаты оценок их IQ приведены в Таблице 9.1.
Таблица 9.1. Величины IQ новозеландских маори

В строке 1 дан общий IQ в 91 балл, а также вербальный и визуализа- ционный IQ в 92 и 94 балла. В строке 2 приведены величины IQ для 13летних испытуемых: 90 баллов для фактора g и рассуждения, 94 для вербальной способности и 87 для визуализации. В строке 3 представлен IQ 82 для выборки 15-летних испытуемых, полученный по результатам теста Отиса — преимущественно вербального теста общего интеллекта. В строках с четвёртой по тринадцатую даны значения IQ в диапазоне от 81 до 95 баллов. В строке 14 приведён IQ 92 по данным национального когортного исследования. В строке 15 дан IQ в 92 балла для вербального рассуждения, полученный у претендентов на посты в правительственной организации в сравнении с 55 претендентами — европейцами. Ни одна из групп не может быть признана репрезентативной для соответствующих популяций, но
132
Ричард Линн
различие интеллекта остаётся приблизительно в середине диапазона данных других исследований.
Все исследования дают сходные оценки IQ в диапазоне от 81 до 96 баллов с медианой в 90. Показатели интеллекта маори единообразно находятся на уровне около 90 баллов по тестам рассуждения, вербальных и невербальных способностей.
Другие аборигены островов Тихого океана
Результаты исследований интеллекта аборигенов островов Тихого океана кроме новозеландских маори приведены в Таблице 9.2.
Таблица 9.2. Показатели интеллекта аборигенов островов Тихого океана

В строке 1 представлен IQ в 85 баллов, полученный в середине 1920-х гг. у туземных детей гавайской начальной школы. В строках 2 и 3 даны значения IQ в 90 и 82 балла для туземных гавайских школьников, полученные в 1924 и 1938 гг.; эти значения даны относительно 100 баллов европейских детей, тестировавшихся в то же время. В строке 4 дан IQ 81 для детей, посещавших школу на Марианских островах. В строке 5 приведён IQ в 84 балла для подростков с Маршалловых островов. В строке 6 представлен IQ 90 для выборки
Расовые различия в интеллекте
133
школьников с островов Самоа, вычисленный по данным словарного теста и теста вербального понимания. В строке 7 дан IQ 82 для выборки аборигенов с островов Папуа-Новой Гвинеи. Испытуемые этой выборки имели в среднем девятилетнее образование и претендовали на поступление в австралийский военно-морской флот. Их IQ вычислен относительно 100 для белых австралийских претендентов также с девятилетним образованием. В строке 8 приведён IQ 89 для дошкольников возраста 4-6 лет с островов Кука. В строке 9 дан IQ 84 для 12-летних школьников с островов Фиджи. В строке 10 приведён IQ детей начальной школы с островов Тонга, равный 86 баллам. В строке 11 представлен IQ 83 для детей начальной школы на Папуа- Новой Гвинее, в значительной степени являвшихся аборигенами островов Тихого океана, а не новогвинейскими аборигенами, и посещавшими школы вместе с белыми австралийскими детьми. В строке 12 приведён IQ 89 для большой выборки филиппинцев на Гавайях, полученный из математического субтеста теста STAS. В строке 13 дан IQ 85 для выборки детей с французской заморской территории Новая Каледония, полученный с помощью кубиков Кооса. В строке 14 приведён IQ 88 для выборки школьников — выходцев с островов Тихого океана, посещавших школы в Новой Зеландии. Медианный IQ аборигенов островов Тихого океана, кроме новозеландских маори, составил 85 баллов, немного ниже медианного значения маори (90 баллов). Объяснение более высокого интеллекта маори состоит в том, что многие из них гибридизированы с европейцами и уровень жизни и здравоохранения у них выше, чем у других аборигенов островов Тихого океана. Поэтому мы считаем, что IQ остальных аборигенов островов Тихого океана в 85 баллов — это наилучшая оценка их интеллекта.
Гибридное население Гавайских островов
В Таблице 9.3. представлены величины IQ детей, у которых один из родителей был представителем туземного населения Гавайских островов, а другой европейцем или китайцем, полученные Смитом (Smith, 1942) в его исследованиях, проведённых в Гонолулу в 1924 и 1938 гг.
Таблица 9.3. Величины IQ гибридов европейцев, китайцев и аборигенов островов Тихого океана

134
Ричард Линн
Величины IQ двух гибридных групп были несколько выше средних значений IQ двух родительских рас. Средний IQ европейцев и гавайцев составляет 90,5 баллов, в то время как IQ их детей — 93 балла. Аналогично, средний IQ китайцев и гавайцев составляет 90, a IQ их детей — 91 балл. То, что величины IQ детей от родителей разных рас несколько выше ожидаемых значений, может объясняться гибридной силой или эффектом гетерозиса, который часто проявляется у гибридов двух пород. Тот же феномен был обнаружен на Гавайях в исследовании азиатско-европейских детей, величины IQ которых были 4 на балла выше, чем у азиатских и европейских детей (Nagoshi, Johnson, 1986). В этом исследовании во всех трех группах родители имели одинаковое образование и социально-экономический статус, что предполагает действие генетического фактора.
Размер мозга аборигенов островов Тихого океана
Оказалось возможным найти только одно исследование размера мозга аборигенов островов Тихого океана. Смит и Билс (Smith, Beals, 1990) дают размеры мозга для шести популяций, среднее значение которых составило 1.317 см3. Они также приводят размер мозга европейцев в 1.369 см3. Различие в 52 см3 представляется достаточно существенным, что позволяет сделать шаг в сторону объяснения различия интеллекта между этими двумя народами.
Средовые и генетические детерминанты интеллекта аборигенов островов Тихого океана
Исследования наследуемости интеллекта аборигенов островов Тихого Океана отсутствуют, но вероятно, и средовые, и генетические факторы ответственны за их более низкий интеллект в сравнении с европейцами. У маори в Новой Зеландии и гавайцев на Гавайях приблизительно та же среда обитания, что и у европейцев, поскольку они пользуются преимуществами проживания в богатых европейских обществах с высокими стандартами питания, образования и социального обеспечения. Величина IQ маори в 90 баллов выше 85 баллов других аборигенов островов Тихого Океана, что указывает на благоприятное воздействие проживания в богатом европейском окружении, но оно остается значительно ниже IQ европейцев. По результатам четырёх исследований средний IQ аборигенов Гавайских островов составил 84 балла — практически столько же, сколько и у других аборигенов островов Тихого Океана (85 баллов). Это гово
Расовые различия в интеллекте
135
рит о том, что обеспеченные условия жизни на Гавайских островах не повышают их интеллект. Размер мозга аборигенов островов Тихого Океана приблизительно на 4 процента меньше, чем у европейцев, что, вероятно, имеет некоторое генетическое основание и вносит вклад в различие интеллекта.
ты ю
ЛШИГШ1Ш0Ч410Й лэнн
Интеллект аборигенов Восточной Азии
Аборигены Восточной Азии в Соединенных Штатах
Другие исследования аборигенов Восточной Азии вне Восточной Азии
Аборигены восточной Азии, усыновлённые европейцами
Гибриды аборигенов Восточной Азии с европейцами
Время реакции
Зрительная память
Размер мозга
Наследуемость интеллекта у аборигенов Восточной Азии
Средовые и генетические объяснения IQ аборигенов Восточной Азии
Аборигены Восточной Азии — коренное население современных Китая, Японии, Кореи, Монголии и Тибета. Их часто называли монголоидами, они были признаны одной из больших рас в классической антропологии, начиная с таксономий Линнея (Linnaeus, 1758) и Блюменбаха (Blumenbach, 1776) и являются одной из семи главных рас в классификации, предложенной Куном, Гарном и Бёрдселлом (Coon, Gam, Birdsell, 1950). Их идентичность как генетического «кластера» была подтверждена Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994) в их классификации, основанной на ряде генетических маркеров, выявлявшихся в выборках ненцев, монголов, тибетцев, корейцев и японцев. Основные отличительные особенности аборигенов Восточной Азии: прямые чёрные волосы, плоский нос, желтоватый цвет кожи и эпикантус — складка над верхним веком, которая обуславливает узкий разрез их глаз.
Интеллект аборигенов Восточной Азии
Исследования интеллекта аборигенов восточной Азии проводились в Китае, Японии, Гонконге, Южной Корее, Тайване и в Сингапуре, где эт
Расовые различия в интеллекте
137
нические китайцы составляют 76 процентов населения. Результаты этих исследований приведены в Таблице 10.1.
Таблица 10.1. Значения IQ аборигенов Восточной Азии
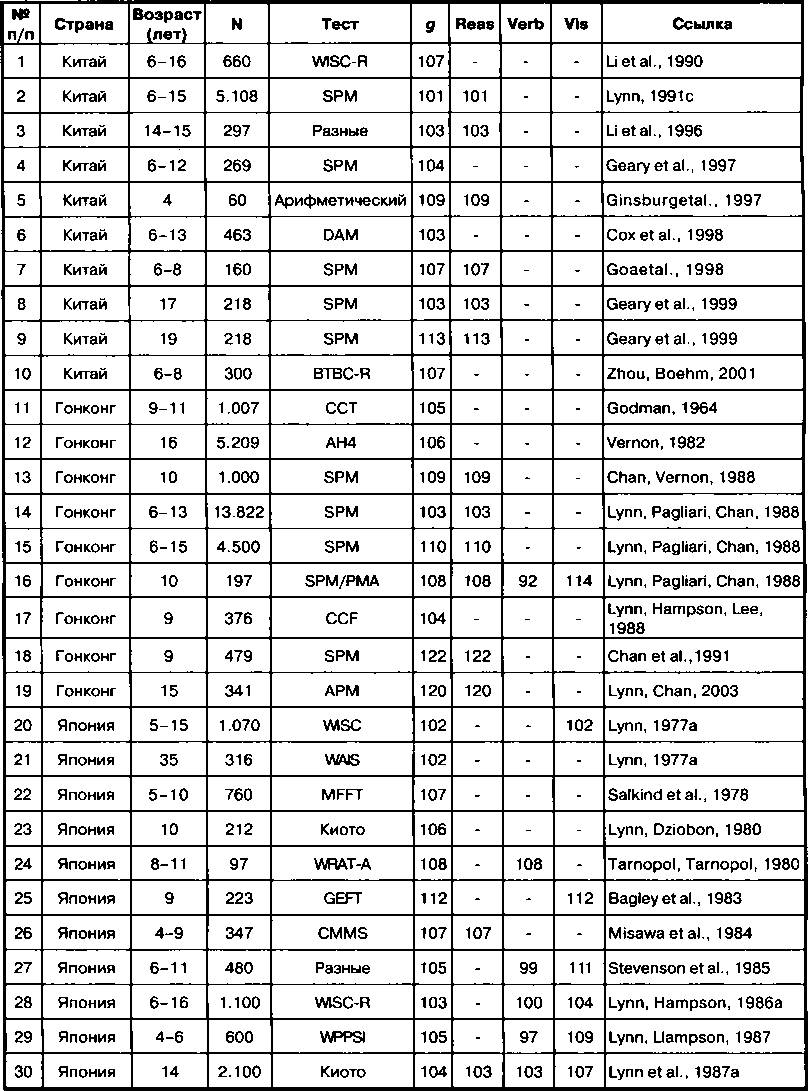
138
Ричард Линн

В строках 1-10 приведены результаты для Китайской Народной Республики. В строке 1 дан IQ в 107 баллов по данным стандартизации теста WISC-R в Шанхае. Это величина, вероятно, несколько выше, чем в Китае
Расовые различия в интеллекте
139
в целом, потому что IQ в Шанхае немного превышает общекитайское значение. В строке 2 представлен IQ в 101 балл, вычисленный из данных стандартизации стандартных прогрессивных матриц в Китае у испытуемых возраста от 6 до 15 лет. В строке 3 дан IQ 103 для 14- и 15-летних подростков, тестировавшихся с помощью нескольких тестов на рассуждение в середине 1990-х гг. В строке 4 приведён IQ 104 для 12-18-летних испытуемых в Шанхае по сравнению с американцами из штатов Миссури и Джорджия. По 10 арифметическим тестам на вычисления и арифметическое рассуждение китайцы имели более высокие оценки, в среднем на 1,37d,что эквивалентно 20 баллам IQ. В этом исследовании также приводятся результаты сравнения успешности выполнения тестов пожилыми китайцами (п=56, возраст 66 лет) и американцами (п=47, возраст 70 лет); китайцы получили среднюю оценку IQ на 8 баллов ниже, чем американцы. Не представлено никакой информации о том, насколько репрезентативны выборки, поэтому мы считаем этот результат недостаточно надёжным для включения в таблицу. В строке 5 дан IQ 109 по тесту арифметического рассуждения для выборки 4-летних дошкольников в Пекине в сравнении с выборкой американских детей (п=156). В строке 6 приведён IQ 103 по тесту рисунка человека и лошади, напоминающему тест «нарисуй человека»; китайские дети учились в школе в Пекине и сравнивались с выборкой британских детей (п=489). В строке 7 представлен IQ 107 для объединённой выборки городских и сельских детей. В строке 8 приведён IQ 103 для выборки 17-летних учащихся полной средней школы в Шанхае в сравнении с выборкой из 55 американских учащихся полной средней школы в г. Колумбия, шт. Миссури. В строке 9 дан IQ 113 для выборки студентов колледжа в Восточном Китайском Нормальном Университете в Шанхае по сравнению с выборкой из 239 американских студентов колледжа университета шт. Миссури. В строке 10 дан IQ в 107 баллов для выборки 7-8-летних школьников в Пекине.
В строках 11-19 представлены результаты девяти исследований в Гонконге. В строке 11 приведён IQ 105, полученный с помощью культурно беспристрастного теста в репрезентативной выборке 9-11-летних китайских школьников из пяти начальных школ. В строке 12 дан IQ 106, полученный в большой выборке 16-летних испытуемых с помощью теста АН4. Так как для этого теста нет удовлетворительных британских возрастных норм, группой сравнения служила выборка канадцев 16-летнего возраста (MacLean, McGhie, 1980). В строках 13-16 приведены величины IQ в 109, 103,110 и 108 баллов, полученные с помощью стандартных прогрессивных матриц (SPM). В строке 16 даны результаты 10-летних испытуемых, у которых оценивали способность к рассуждению с помощью SPM, пространственную способность с помощью теста пространственных отношений, входящего в тест первичных умственных способностей, а вербальную
140
Ричард Линн
способность оценивали по беглости речи. В этом исследовании выявлен ярко выраженный восточно-азиатский паттерн высокого IQ рассуждения (108), ещё более высокого пространственного IQ (114) и более низкого вербального IQ (92). В строке 17 дан IQ в 104 балла, полученный с помощью культурно беспристрастного теста. В строке 18 приведён необычно высокий IQ для выборки 9-летних детей — 122 балла. В строке 19 приведено близкое значение IQ (120 баллов), полученное с помощью теста прогрессивных матриц повышенной сложности в Гонконгской выборке стандартизации, которая, по-видимому, была исключительно тщательно сформирована.
В строках 20-42 приведены значения IQ по исследованиям в Японии. В строке 20 дан IQ в 102 балла для всей Японии, вычисленный по данным японской выборки стандартизации теста WISC и основанный на результатах пяти тестов действия и теста повторения цифровых рядов Digit Span (вербальные тесты в японской версии теста были изменены и поэтому нами не учитываются); IQ визуализации (102 балла) вычислен по субтестам лабиринтов и складывания кубиков. В строке 21 дан IQ для Японии также в 102 балла, вычисленный по данным выборки стандартизации теста WAIS и основанный на данных субтестов замещения символов цифрами (Digit Symbol), складывания кубиков и Digit Span — лишь эти тесты не были изменены в японской версии. В строке 22 дан IQ в 107 баллов для японских детей возраста 5-10 лет по тесту MFFT, вычисленный по показателю количества ошибок по сравнению с американской выборкой (п=2.676). В строке 23 представлен IQ 106 для 10-летних японских детей, полученный с помощью японского теста Киото в сравнении с британскими детьми. В строке 24 приведён IQ 108 для выборки детей из Хиросимы по результатам арифметического субтеста WRAT. В строке 25 дан IQ 112 для японских детей из г. Нагоя и Хамамацу. В строке 26 приведён IQ 107, полученный на японской выборке стандартизации Колумбийской шкалы умственной зрелости.
В строке 27 приведены результаты исследования Стивенсона и соавторов, которые сравнивали шести- и одиннадцатилетних испытуемых из выборок, сформированных в г. Миннеаполис в Соединенных Штатах, Сендай в Японии и Тайбэй на Тайване. В то время как Сендай и Тайбэй приемлемы для формирования репрезентативной выборки городских детей в Японии и на Тайване, о Миннеаполисе же нельзя сказать, что он является репрезентативным представителем американских городов. Миннеаполис — главный город шт. Миннесота, при этом имеется немало свидетельств о том, что уровень интеллекта в Миннесоте выше, чем в Соединенных Штатах в целом. При отборе в армию в период Первой Мировой войны белые из Миннесоты получили наивысшие оценки среди всех американских штатов по Американскому армейскому тесту бета (Montagu,
Расовые различия в интеллекте
141
1945Ь). Во время корейской войны процент признанных непригодными для военной службы по причине низкого интеллекта в Миннесоте был предпоследним среди американских штатов (Jensen, 1973, стр. 107), что указывает на высокий средний уровень интеллекта. Результаты выполнения математического теста, предъявлявшегося учащимся 8-го класса из шт. Миннесота в 2003 г. в рамках Национальной программы мониторинга образования (NAEP), были наилучшими среди всех американских штатов (National Center for Education Statistics, 2003). Флинн вычислил (Flynn, 1980, стр. 107), что средний IQ белых в Миннесоте равен 105 баллам. Примем это как наилучшую оценку. Следовательно, для сравнения с IQ белых американцев, равным 100 баллам, необходимо к данным выборок из Японии и Тайваня прибавить 5 баллов, что даст им оценку IQ в 105 баллов — сопоставимую с результатами многих других исследований.
В строке 28 дан общий (для всей шкалы) IQ в 103 балла, полученный на японской выборке стандартизации теста WISC-R, вербальный IQ в 100 баллов (по данным пяти вербальных субтестов) и IQ визуализации в 104 балла (по субтесту складывания кубиков). В строке 29 дан общий (для всей шкалы) IQ в 103 балла, полученный из данных японской выборки стандартизации теста WPPSI, вербальный IQ в 97 (по пяти вербальным субтестам) и IQ визуализации в 109 баллов (по четырём субтестам действия).
В строке 30 приведён IQ в 103 балла для рассуждения, в 103 для вербальной способности и 107 баллов для способности к визуализации, полученные по результатам тестировании репрезентативной выборки британских детей с помощью теста Киото; эти три показателя IQ были усреднены, что дало 104 балла для общего IQ. В строке 31 приведены значения IQ в 104 балла для рассуждения и в 114 баллов для способности к визуализации, полученные с помощью теста DAT на выборке 13-15-летних японских подростков. В строке 32 дан общий IQ в 103 балла, в 102 балла для «последовательной обработки» (приблизительный эквивалент вербальной способности) и в 105 баллов для «одновременной обработки» (приблизительный эквивалент способности к визуализации), вычисленный по данным японской выборки стандартизации теста Маккарти. В строке 33 приведён IQ 103, полученный из анализа Кауфмана и соавторов (Kaufman et al., 1989) данных японской выборки стандартизации теста WISC-R для последовательного и одновременного факторов Кауфмана. Корреляция «последовательной обработки» (приблизительная мера вербальной способности) с вербальным IQ по тесту Векслера составила 0,44, а «одновременной обработки» (приблизительная мера способности к визуализации) с IQ действия по Векслеру оказалась равной 0,73. Два эти показателя интеллекта были усреднены, чтобы получить меру g.Кроме того, этот тест содержит матричный тест аналогий, сходный с прогрессивными матрицами, результаты которого были включены в таблицу как IQ рассуждения.
142
Ричард Линн
В строке 34 приведён IQ в 113 баллов для выборки подростков из школы в Кейо в сравнении с американской выборкой школьников (п=121) из школы во Флориде; вербальный IQ в 116 баллов был вычислен по результатам арифметического теста, a IQ в 110 баллов для визуализации — по результатам тестов вращения в уме и лабиринтов. В строке 35 дан IQ в 110 баллов для выборки 9-летних детей из Токио. В строке 36 приведены результаты сравнения показателей японских детей из г. Нагоя с канадскими нормами для Канадского теста когнитивных способностей (ССАТ). Средний японский IQ для рассуждения, полученный в японской выборке (106 баллов), типичен для ряда других исследований, но вербальный IQ в 121 балл необычайно высок для японских детей. В этом исследовании у японских детей также был определён количественный IQ в 112 баллов. Возрастной диапазон детей составлял 5-7 лет, и преимущество японских 5-летних детей было так же велико, как и 6-7-летних. Пятилетние посещали детский сад. Высокие оценки IQ 5-летних японцев делают практически невероятным предположение о том, что преимущество японцев может быть результатом более эффективного обучения, выдвинутое Стивенсоном и соавторами (Stevenson et al., 1985). В строке 37 представлены результаты сравнения японских детей из города средних размеров Мацуяма с американскими нормами Матричного теста аналогий; средний IQ японских детей составил 105 баллов.
В строке 38 приведены величины IQ в 103 балла для рассуждения и в 109 баллов для визуализации, полученные в середине 1990-х гг. у 14-15летних испытуемых как средние значения данных нескольких тестов. Этот результат — часть того же исследования, в котором IQ детей того же возраста в Китае был оценен в 103 балла, что даёт основание предположить, что в середине 1990-х гг. величины IQ китайских и японских детей были одинаковыми. В строке 39 дан вербальный IQ в 100 баллов, выведенный из теста общих знаний, предъявленного шести- и семнадцатилетним испытуемым в японском городе Сендай в сравнении с данными для американского города Миннеаполис. Поскольку средний IQ белых в Миннеаполисе оценивается в 105 баллов (см. комментарий к строке 27), среднее значение японцев было увеличено на 5 баллов. В строке 40 приведён IQ визуализации в 102 балла, полученный путём сравнения выборки японских старшеклассников и студентов университетов с выборкой из 52 европейцев, студентов университетского колледжа в Дублине. В строке 41 дан IQ 102, полученный у 7-летних и 11-летних японских детей в сравнении с сопоставимой выборкой из 60 британских детей. В строке 42 дан вербальный IQ 105 для 17-летних японцев, выведенный из теста общих знаний в сравнении с американцами из Миннеаполиса; среднее значение японцев было увеличено на 5 баллов по причине, указанной в комментарии к строке 39.
Расовые различия в интеллекте
143
В строках 43 и 44 приведены результаты для Сингапура. В строке 43 дан IQ в 107 баллов для выборки 13-летних подростков. В строке 44 дан IQ в 114 баллов для 15-летних испытуемых, полученный с помощью прогрессивных матриц повышенной сложности. Это значение существенно выше, хотя и не столь высоко, как в двух последних исследованиях в Гонконге.
В строках 45-48 даны четыре результата для Южной Кореи. В строке 45 приведён IQ 113, полученный по данным выборки стандартизации теста К-ABC Кауфмана — исключительно хорошо построенного и стандартизированного американского теста. В этом исследовании показан типичный паттерн аборигенов Восточной Азии: высокий IQ рассуждения (110, по результатам матричного теста аналогий), высокий пространственный IQ (120) и более низкий вербальный IQ (106). В строке 46 представлен IQ109 и сходный паттерн более низкой вербальной способности в сравнении со способностью к визуализации. В строке 47 приведён IQ в 103 балла для социально репрезентативной выборки 4-летних испытуемых из дошкольного учреждения в районе г. Пусан в сравнении со 156 американскими детьми. В строке 48 дан IQ 100, базирующийся на выборке стандартизации теста WISC-III.
В строках 49-59 представлены одиннадцать результатов для Тайваня. В строке 49 дан IQ 102, полученный из результатов раннего исследования 1950-х гг. В строках 50, 51 и 53 даны оценки IQ в 110, 105 и 104 балла, полученные учащимися начальной школы. В строке 52 приведён IQ 107 для выборки детей в Тайбэе по данным арифметического субтеста WRAT. В строке 54 дан IQ 105, полученный путём сравнения тайваньских детей с американской выборкой из Миннеаполиса, где средний IQ белых был оценен в 105 баллов (как объяснено в комментарии к строке 27), соответственно средний IQ тайваньцев был увеличен на 5 баллов. В строке 55 дан IQ 100 для общей информированности или теста знаний, предъявленного выборкам испытуемых из Соединенных Штатов (п= 1.052) и из Тайбэя на Тайване. В строке 56 приведён IQ 6-8-летних детей из начальных школ Тайбэя и провинциальных городов и деревень, равный 105 баллам. Общие знания — компонент вербального интеллекта, как показано посредством многократно выполнявшегося факторного анализа результатов тестов Векслера (Carroll, 1993), результаты этого теста введены как вербальный IQ. В строке 57 дан IQ в 105 баллов для невербального рассуждения. В строке 58 приведён IQ 107 для теста общих знаний, предъявленного выборкам испытуемых в Соединенных Штатах (п=1.052) и в Тайбэе. Тайваньская выборка получила оценку на 2 балла выше, чем американская, но американская выборка была из г. Миннеаполис, где средний IQ белых оценивается в 105 баллов (как объяснено в комментарии к строке 27), поэтому среднее значение тайваньцев было увеличено на 5 баллов до 107. Медианное значение IQ результатов десяти исследований на Тайване составляет 105 баллов.
144
Ричард Линн
По результатам работ, приведённым в Таблице 10.1, можно сделать два вывода. Во-первых, все величины IQ аборигенов Восточной Азии немного выше, чем у европейцев, за исключением результатов оценки общей информированности Чена и соавторов (Chen et al., 1996) в Японии и на Тайване, и Георгаса и соавторов (Georgas et al, 2003) в Южной Корее; обе работы дают аборигенам Восточной Азии оценку IQ в 100 баллов. Диапазон оценок IQ лежит в пределах от 100 до 122 баллов. Медианная величина IQ по всем исследованиям составляет 105 баллов и её следует принять как наилучшую оценку IQ аборигенов Восточной Азии. Во-вторых, в одиннадцати работах приводятся меры вербальной и визуализаци- онной способностей и в десяти из них IQ визуализации был выше вербального IQ (исследование, приведённое в строке 36 является исключением). Среднее арифметическое и медиана разностей этих двух способностей равны 12 баллам IQ. Это различие проявляется в разных тестах. Более высокая способность к визуализации и более низкая вербальная способность аборигенов Восточной Азии в сравнении с европейцами выявляется так последовательно и это является настолько выраженным, что представляется реальным феноменом.
Аборигены Восточной Азии в Соединенных Штатах
Аборигены восточной Азии обосновались во многих странах, включая Соединенные Штаты, Канаду, Европу, Бразилию и Малайзию. Наибольшее число исследований интеллекта аборигенов Восточной Азии вне Восточной Азии было выполнено в Соединенных Штатах. Они были собраны воедино и обсуждены Верноном (Vernon, 1982) и Флинном (Flynn,
. Вернон пришёл к заключению, что у выходцев из Восточной Азии в США вербальный IQ составляет 97 баллов, a IQ по невербальным и пространственным тестам — 110 баллов (стр. 28). Его анализ имеет два слабых места. Во-первых, нет общепринятого значения понятия «невербальный» интеллект. Это нечёткое понятие является неудовлетворительным, так как может включать любую способность, не являющуюся вербальной, включая абстрактное рассуждение, визуализацию, пространственные и перцептивные способности. Во-вторых, Вернон не принял во внимание долговременное возрастание тестовых норм, в результате которого группы, протестированные с помощью теста, нормировавшегося в более ранний период, получают завышенные значения IQ. Анализ Флинна лучше, поскольку он делает поправку IQ на долговременное возрастание тестовых норм, но он также анализирует интеллект в терминах вербального и «невербального» IQ и усредняет их, чтобы получить «общий IQ». Флинн
Расовые различия в интеллекте
145
делает вывод (стр. 65), что у американских этнических китайцев и японцев вербальный IQ составляет 95,3, а «невербальный IQ» — 99,6 баллов, и он усредняет их, получая «общий IQ» в 97,6 баллов. Это неудовлетворительный анализ, потому что «невербальный IQ» не признаётся сегодня осмысленным понятием. Более того, то, как он использует понятия вербального и невербального интеллекта, даёт вербальной способности при вычислении общего интеллекта такой же вес, как и всем другим способностям, а поскольку аборигены Восточной Азии имеют относительно слабую вербальную способность, это вызывает ложное занижение их IQ. Общий интеллект или gлучше всего может быть измерен либо тестами невербального рассуждения, либо как среднее значение вербальной способности и способностей к рассуждению и визуализации. Несмотря на эту концептуальную слабость, Флинн выполнил полезный литературный обзор и анализ, и в целом я принимаю его оценки.
Проблема с исследованиями выходцев из Восточной Азии в Соединенных Штатах и в других местах состоит в том, что многие из них продолжают говорить на японском, китайском или корейском как на первом языке и вследствие этого плохо выполняют вербальные тесты на английском. Во многих случаях невозможно говорить о степени этого затруднения.
Результаты исследований выходцев из Восточной Азии в Соединенных Штатах приведены в Таблице 10.2.
В строке 1 представлен IQ 97 для китайских детей в Сан-Франциско. Они обследовались с помощью теста Бине, являющегося в значительной степени вербальным тестом, и это вероятно создавало трудности детям, многие из которых говорили на английском как на втором языке. В строке 2 дан IQ для невербального рассуждения в 99 баллов и IQ по тесту на знание слов в 95 баллов для китайских детей на Гавайях. В строке 3 дан IQ в 101 балл для китайцев и японцев, полученный на выборке стандартизации теста «Нарисуй человека». В строках 4 и 5 приведены величины IQ в 100 и 103 балла для китайцев и японцев на Гавайях, полученные при помощи теста «Лабиринты Портеуса». В строках 6, 7 и 8 даны IQ для невербального рассуждения в 99,101 и 102 балла для этнических китайцев, японцев и корейцев в Гонолулу по сравнению с проживающими там же белыми. Китайцы, японцы и корейцы существенно уступали белым в вербальных тестах (IQ 89, 86 и 88 баллов, соответственно). Невозможно определить, в какой степени низкие значения вербального IQ китайцев, японцев и корейцев были обусловлены их общением дома на родном языке и возникающими отсюда трудностями выполнения вербальных тестов на английском, а насколько они определялись типичным для аборигенов Восточной Азии паттерном более низкой вербальной способности по сравнению со способностью к рассуждению. Вероятно, «работали» оба фактора. В строке 9 приведено вычисленное Флинном (Flynn, 1991) значение вербального
146
Ричард Линн
Таблица 10.2. IQ аборигенов Восточной Азии в Соединенных Штатах

Расовые различия в интеллекте
147
IQ в 96 баллов для 18-летних японцев, интернированных во время Второй Мировой войны из которых 91 процент были иммигрантами второго поколения и 8 процентов — иммигрантами третьего поколения. Как и в исследовании в Гонолулу, неясно, у какого процента из них основным языком общения дома был японский и, следовательно, были трудности при выполнении вербальных тестов на английском. В строке 10 приведён вербальный IQ в 97 и IQ визуализации в 106 баллов для выборки 6-летних китайцев в Нью-Йорке. Неизвестно, говорили ли некоторые из этих детей на китайском языке дома и, следовательно, имели сложности при выполнении вербальных тестов. Предполагается, что половина из них говорила, ввиду чего вербальному IQ была присвоена только половина веса IQ визуализации при вычислении величины общего интеллекта, которая составила таким образом 103 балла.
В строке 11 приведены величины IQ этнических китайцев, японцев и корейцев в 100 баллов для рассуждения и в 97 баллов для вербальной способности, рассчитанные Флинном по данным общенационального исследования Колмена. Величина IQ рассуждения была принята как мера фактора g.В строке 12 дан вербальный IQ 96 для этнических китайцев и японцев на Гавайях, вычисленный Флинном. В строке 13 приведены результаты японских детей на острове Кауаи Гавайского архипелага. Средние оценки IQ составляли: 107 баллов для вербальной способности, 105 для пространственной, 112 для рассуждения, 105 для восприятия и 106 баллов для числовой способности. Флинн подсчитал (Flynn, 1991), что нормы теста устарели на 33 года, и поэтому из оценок IQ следует вычесть 10 баллов. В результате он пришёл к оценке вербального IQ в 97 баллов, невербального (вычисленный как среднее остальных четырёх тестов) в 99 баллов, и, отсюда, общего IQ в 98 баллов. Результаты необычны в плане относительно низкого IQ (95 баллов) для пространственной способности (визуализации). В строке 14 дан общий IQ 99 и вербальный IQ 95 для 17-летних испытуемых из Лос-Анджелеса, вычисленные Флинном по данным 1969-1970 гг. В строке 15 приведён IQ в 103 балла для китайцев из Сан-Франциско, имевших сходный с белыми социально-экономический статус, обследованных с помощью теста понимания карт, охарактеризованного как мера пространственного рассуждения. В том же исследовании чернокожие, также имевшие сходный с белыми социально-экономический статус, получили оценку IQ 90 баллов.
В строке 16 приведены IQ в 101 балл по словарному субтесту и в 102 балла по субтесту складывания кубиков теста WISC, полученные в общенациональном исследовании и давшие усредненный IQ в 101 балл для фактора g(Флинн пропустил это исследование в своём обзоре). В строке 17 даны IQ невербального рассуждения 98 и вербальный IQ 99 (по результатам тестов информированности и знания английского языка, проводившихся в
148
Ричард Линн
рамках общенационального исследования, рассчитанные Флинном). Более высокий вербальный IQ в сравнении с IQ рассуждения необычен, это находится в таком противоречии с обычным паттерном аборигенов Восточной Азии (IQ способности к рассуждению выше IQ вербальной способности), что данный результат не может считаться надёжным. В строке 18 даны невербальный IQ действия в 101 и вербальный IQ в 91 балл, полученные по результатам обследования с помощью теста WISC небольшой выборки 9-летних детей из китайского квартала Сан-Франциско, вычисленные Флинном (Flynn, 1991). Дети посещали китайские частные школы, и их низкий вербальный IQ следует, вероятно, объяснить тем, что многие дети говорили по-китайски и в школе, и дома. Ввиду этого, IQ действия можно принять как наилучшую меру их общего интеллекта. В строке 19 дан IQ в 101 балл для китайской выборки из Калифорнии.
В строке 20 дан IQ 99, определённый Флинном по данным разных исследований, собранных Совеллом. Использованные в этих исследованиях тесты не идентифицированы и их результаты взяты как мера фактора g.В строке 21 приведён IQ в 107 баллов, полученный с помощью теста математических навыков на большой выборке японских подростков на Гавайях в сравнении с выборкой европейцев (п=3.722), также сформированной на Гавайях.
В строке 22 дан IQ 101 для китайских детей в Калифорнии, данные собраны Дженсеном в 1975 г. и проанализированы Флинном. Половина детей родилась за границей (главным образом в Гонконге), что, вероятно, объясняет их низкий вербальный IQ — 89 баллов.
В строке 23 приведены результаты, собранные Дженсеном в богатом районе г. Беркли, Калифорния, и обработанные Флинном. Как дети китайцев и японцев, так и дети белых, получили высокие оценки. Цифры, помещённые в таблицу, являются оценками Флинна для детей выходцев из Восточной Азии, вычисленными относительно национальных норм. В сравнении с белыми детьми из того же района (п= 1.506) дети выходцев из Восточной Азии получили IQ в 98 баллов для рассуждения и в 95 баллов для вербального интеллекта. Эта выборка не является репрезентативной. В строке 24 приведены результаты для выборки китайцев из Калифорнии. В строке 25 дан IQ в 103 балла для небольшой, но национально репрезентативной выборки подростков. В строке 26 дан IQ 104 для американских выходцев из Азии по данным выборки стандартизации дифференциальной шкалы способностей (Differential Ability Scale). Американская перепись населения 1980 г. показала, что приблизительно половина американских выходцев из Азии — выходцы из Восточной Азии: этнические китайцы, японцы, корейцы и некоторое количество вьетнамцев9. Остальные — главным
9 Часть выходцев из северных районов Вьетнама являются этническими китайцам (персональное сообщение автора).
Расовые различия в интеллекте
149
образом филиппинцы, вьетнамцы, тайцы, камбоджийцы и другие выходцы из Юго-Восточной Азии. У выходцев из Юго-Восточной Азии IQ ниже европейского и, таким образом, приведённые в таблице результаты преуменьшают интеллект американских выходцев из Восточной Азии. В строке 27 дан IQ в 109 баллов для маленькой выборки выходцев из Восточной Азии, оцененный по данным Национального Коллаборативного Перинатального Проекта, полученным приблизительно в 1966 г.
Медианное значение IQ по результатам девяти исследований первой половины двадцатого века составило 101 балл, что немного ниже медианного значения IQ по данным девяти исследований, выполненных начиная с 1980 г. — 104 балла (почти совпадающего со 105 баллами аборигенов Восточной Азии в странах их исконного проживания). Имеется три возможных объяснения повышения интеллекта выходцев Восточной Азии в Соединенных Штатах в течение двадцатого столетия. Во-первых, многие из обследованных в первой половине двадцатого века говорили на китайском или японском языке как на первом языке, что затрудняло им выполнение тестов на английском языке. Во-вторых, возможно интеллект аборигенов Восточной Азии, эмигрировавших в Соединенные Штаты, был в целом немного ниже среднего интеллекта тех, кто оставался в Восточной Азии. Китайцы и японцы, эмигрировавшие в Соединенные Штаты во второй половине девятнадцатого столетия, были в основномкрестьянами, приехавшими для выполнения низкоквалифицированных работ на строительстве железных дорог и других строительных работ. Этот вариант, вероятно, не казался привлекательным более интеллектуальным людям, которые обычно достаточно хорошо устроены в их собственных странах. У детей ранних мигрантов, обжившихся в Соединенных Штатах, IQ несколько «прогрессировал» к среднему значению аборигенов Восточной Азии, равному 105 баллам.
В-третьих, в пяти исследованиях содержатся оценки вербальной и ви- зуализационной способностей, и в четырех из них IQ способности к визуализации был выше вербального IQ (исследование в строке 13 является исключением). Среднее различие величин этих двух способностей составило 4,4 балла IQ и отмечалось в исследованиях с использованием различных тестов. Это согласутеся с паттерном результатов аборигенов Восточной Азии, проживающих с своих странах (Таблица 10.1).
150
Ричард Линн
Другие исследования аборигенов Восточной Азии вне Северо-Восточной Азии
Исследования интеллекта аборигенов Восточной Азии, проживающих вне Северо-Восточной Азии и Соединенных Штатов, представлены в Таблице 10.3.
Таблица 10.3. Другие исследования аборигенов Восточной Азии вне Северо-восточной Азии

В строке 1 приведён IQ в 99 баллов для этнических японцев в Бразилии. В строке 2 дан IQ в 107 баллов для японских детей в Лондоне. Действительный IQ этой выборки составлял 115 баллов, но родители детей были в основном бизнесменами, дипломатами и представителями свободных профессий и, следовательно, IQ в этой выборке должен быть завышен. Обычно различие в IQ между верхними и нижними социально-экономическими классами составляет приблизительно 15 баллов (см. напр., Nettle, 2003). Таким образом, величина IQ средних японских детей должна составлять около 107 баллов. В строках 3 и 4 даны IQ в 104 и 95 баллов по ранним исследованиям японцев и китайцев в Ванкувере. Согласно Вернону (Vernon, 1982), китайские иммигранты происходили из бедных крестьян, в то время как японцы были квалифицированными рабочими и представителями среднего класса, что объясняет, почему японцы выполняли тесты лучше. В строке 5 дан IQ в 101 балл по данным более позднего исследования китайцев в Ванкувере. В строках 6 и 7 приведены результаты двух последующих исследований китайцев в Канаде, давших оценки IQ в 101 и 103 балла. В строке 8 представлен IQ 99 для этнических китайцев в Малайзии — на 10 баллов выше, чем у малайцев (см. Таблицу 7.1). Причи
Расовые различия в интеллекте
151
ной того, что IQ этнических китайцев в Малайзии оказался немного ниже, чем у других представителей коренного населения Восточной Азии, может служить то, что они — относительно недавние иммигранты, принятые на низкоквалифицированную работу, и, возможно, IQ этих иммигрантов был несколько ниже среднего китайского значения. В строке 9 дан IQ для числового рассуждения в 102 балла для детей китайских иммигрантов в Нидерландах. Их IQ вербального понимании составил 85 баллов, но так как они говорили на китайском как на первом языке, этот результат нельзя считать валидным. Медианное значение IQ по девяти исследованиям составило 101 балл — точно такое же, как у выходцев из Восточной Азии в Соединенных Штатах.
Из результатов исследований, представленных в Таблице 10.3, можно сделать три вывода. Во-первых, среднее значение IQ выходцев из Восточной Азии немного выше, чем у европейцев, живущих в сходных условиях. В Бразилии IQ этнических японцев составляет 99 — на 4 балла выше, чем у европейцев (см. Таблицу 3.2). Предки японцев в этой стране были приняты на работу в качестве сельскохозяйственных чернорабочих в конце девятнадцатого столетия и, возможно, величина их IQ была несколько ниже среднего японского уровня. Кроме того, в Бразилии значения IQ как у японцев, так и у европейцев немного снижены (до 95 баллов, см. Таблицу 3.2), вероятно отчасти из-за низкого жизненного уровня в Бразилии (6.625 $ валового дохода на душу населения в год в 1998 г. против 20.336 $ в Великобритании). Медианное значение IQ по реультатам четырёх канадских исследований составило 101 балл. Медианное значение IQ по всем исследованиям в Соединенных Штатах также было равно 101 баллу. Это немного выше оценки Флинна (97,6 балла), так как Флинн не рассматривает исследования Симондса (Symonds, 1924), Гудинаф (Goodenough, 1926b), Фельдмана (Feldman, 1971), Департамента Здравоохранения, Образования и Социального Обеспечения Соединенных Штатов (1971) и четыре последние исследования, которые были опубликованы позже его анализа. Таким образом, IQ выходцев из Восточной Азии в Соединенных Штатах немного ниже 105 баллов аборигенов Восточной Азии в их исконных странах. Для этого есть четыре возможных объяснения.
Во-первых, IQ мигрировавших в Соединенные Штаты мог быть несколько ниже среднего уровня оставшихся в Азии. Это возможно потому, что аборигены Восточной Азии в Соединенных Штатах и в других местах вне Восточной Азии являются потомками иммигрантов, которые мигрировали, чтобы устроиться на низко квалифицированную работу и могли иметь интеллект немного ниже среднего по популяциям, из которых они произошли.
Во-вторых, многие китайцы и японцы говорили на китайском и японском языке как на первом языке, что затрудняло им выполнение некоторых тестов.
152
Ричард Линн
В-третьих, может потребоваться два поколения для того, чтобы иммигранты из бедных стран смогли преодолеть последствия недостаточного питания и смогли полностью раскрыть свой потенциал. Средний IQ выходцев из Восточной Азии по последним шести представленным в Таблице 10.2 исследованиям, опубликованным начиная с 1990 г., составил 105 баллов — такой же, как у аборигенов Восточной Азии в их собственных странах.
В-четвертых, в трёх работах представлены меры вербальной способности и способности к визуализации, во всех случаях IQ визуализации был выше вербального IQ. Среднее различие между этими двумя способностями составило 7 баллов. Это подтверждает паттерн «низкая вербальная способность — высокая способность к визуализации», выявленный в Восточной Азии и в Соединенных Штатах. О дальнейшем исследовании, выявившим этот паттерн способностей, сообщено Раштоном (Rushton, 1992а), изучавшим интеллект студентов университетов в Канаде. Средний вербальный IQ выборки студентов из Восточной Азии составил 112,8 баллов, средний IQ действия (главным образом визуализации) — 120,6 баллов, в то время как у студентов — европейцев средний вербальный IQ составлял 117,7, а средний IQ действия (главным образом визуализации) — 118,8 баллов.
Аборигены Восточной Азии, усыновлённые европейцами
Было проведено шесть исследований интеллекта младенцев из Восточной Азии, усыновлённых европейскими семьями в Европе и Соединенных Штатах. Результаты представлены в Таблице 10.4.
Таблица 10.4. IQ детей из Восточной Азии, усыновлённых европейцами

В строках 1-3 приведены величины IQ корейских детей, воспитываемых белыми американскими приёмными родителями. Выборка была раз
Расовые различия в интеллекте
153
делена на три группы: «тяжёлая степень недоедания в младенчестве» (строка 1), «умеренная степень недоедания в младенчестве» (строка 2) и «хорошо питавшиеся в младенчестве» (строка 3). Величины IQ в этих трёх группах оказались связаны с уровнем питания. Оценки в серьёзно недоедавшей группе существенно не отличались от оценок американских белых, но испытуемые двух других групп получили более высокие оценки. Не приводится подробного описания способа определения величин IQ, которые выводились из школьных отметок, и, вероятно, были завышены вследствие использования устаревших норм. В строке 4 дан вербальный IQ в 115 баллов для выборки из 25 выходцев преимущественно из Восточной Азии (12 человек было из Вьетнама (главным образом этнические китайцы), 10 из Кореи, 3 из Камбоджи и 2 — из Таиланда).
В строках 5 и 6 даны результаты аналогичных исследований в Европе. В строке 5 приведён IQ в 110 баллов для корейских детей, усыновлённых младенцами в Бельгии; их способность к визуализации превышала вербальную способность, что типично для аборигенов Восточной Азии. В строке 6 приведён IQ в 108 баллов для 36 корейских детей, усыновлённых голландскими семьями в Нидерландах.
Среднее значение IQ по шести исследованиям составило 109 баллов, а если исключить первые два исследования плохо питавшихся младенцев, то 111 баллов. Вероятно, одна из причин такого высокого значения IQ состоит в том, что это были маленькие дети, усыновлённые в основном семьями среднего класса. Из работы Вайнберга, Скарр и Вальдмана (Weinberg, Scarr, Waldman, 1992) известно, что воспитание приёмными родителями среднего класса повышает IQ приёмных детей в раннем и среднем детском возрасте, но этот эффект постепенно исчезает в старшем подростковом периоде и при достижении совершенно л ения. В этом исследовании у чернокожих младенцев, усыновлённых белыми родителями среднего класса, средний IQ в семь лет был равен 95 баллам, но он упал до 89 баллов в 17 лет; это указывает на то, что воспитание в белых семьях среднего класса повышает IQ воспитанников в детстве на 6 баллов (Levin, 1994; Lynn, 1994с).
Применив этот результат к 112 баллам IQ у нормально питавшихся приёмных детей из Восточной Азии, мы можем предположить, что ко взрослому состоянию их IQ уменьшился бы на 6 баллов — до 106, то есть практически до величины, характерной для аборигенов Восточной Азии в своих странах и, согласно данным последних исследований, в Соединенных Штатах.
Представленные в строке 5 результаты приёмных детей в Бельгии еще раз демонстрируют паттерн низкой вербальной способности и высокой способности к визуализации у аборигенов Восточной Азии, проживающих в Восточной Азии, в Соединенных Штатах и повсюду. Этот расовый
154
Ричард Линн
паттерн обнаруживается последовательно и в разных местах, что свидетельствует о его генетической природе.
Гибриды аборигенов Восточной Азии с европейцами
В Главе 4 приводилось немало свидетельств того, что величина интеллекта гибридов африканцев с европейцами находится в промежутке между величинами африканцев и европейцев. Можно ожидать, что уровень интеллекта гибридов аборигенов Восточной Азии с европейцами также будет промежуточным между уровнями двух родительских рас. Имеется всего одна работа по этой проблеме — исследование Раштона в рамках анализа данных Американского Национального Коллаборативного Перинатального Проекта (Rushton, 1997). В исследовании участвовало 53.043 ребёнка, по которым получали информацию разного рода при рождении, в младенчестве и в возрасте семи лет, когда у них измеряли IQ (с помощью теста WISC) и главную окружность головы. Величины IQ выходцев из Восточной Азии, гибридов аборигенов Восточной Азии с европейцами (37 случаев, в 5 случаях с чернокожими, но эти данные не были отброшены), европейцев и афроамериканцев в возрасте 7 лет приведены в Таблице 10.5.
Таблица 10.5. Величины IQ и размер мозга (см?) гибридов аборигенов Восточной Азии с европейцами

В строке 1 приведены количества детей в группах. В строке 2 даны представленные автором средние величины IQ. В строке 3 приведены значения IQ с поправкой на долговременное увеличение норм теста с 1949 г. (год стандартизации теста WISC) по 1966 г («медианный» год сбора данных): из представленных автором значений IQ вычитали по 5 баллов. В строке 4 дан размер мозга в кубических сантиметрах, оцененный по главной окружности головы. Обратите внимание, что по всем трём измерениям гибриды попадают в промежуток между аборигенами Восточной Азии и европейцами. То, что они ближе к европейцам можно объяснить тем, что в 5 случаях это были гибриды аборигенов Восточной Азии с африканцами.
Расовые различия в интеллекте
155
Время реакции
Время реакции представляет собой скорость реакции на простой стимул, такой как вспышка света. Во многих исследованиях было показано, что время реакции положительно связано с интеллектом, значение корреляция лежит в пределах от 0,2 до 0,3 (см. Jensen, 1998; Deary, 2000). Согласно аргументации Дженсена (Jensen, 1998), время реакции является мерой неврологической эффективности мозга при обработке информации. В Главе 4 мы видели, что по сравнению с европейцами африканцы характеризуются более медленным временем реакции, что согласуется с их более низким IQ. Рассмотрим теперь, обладают ли аборигены Восточной Азии более быстрым по сравнению с европейцами временем реакции согласно их более высокому IQ.
Результаты трёх исследований по этой проблеме приведены в Таблице 10.6. Во всех трёх исследованиях время реакции (RT) являлось средним значением для трёх задач на время реакции, а именно, времени простой реакции (скорость реакции на единственный световой стимул), времени реакции различения (когда надо реагировать на один из восьми стимулов) и времени реакции «третий лишний» (когда надо реагировать на наиболее отстоящий от двух других стимул). В строке 1 сравниваются японские и британские 9-летние дети. Величина IQ японских детей составила 110 баллов, или на 0,66dвыше, а время реакции было на 0,50*/ выше, чем у британских детей. В строке 2 приведены результаты аналогично построенного исследования, в котором сравнивались китайские дети из Гонконга с белыми британскими детьми. Величина IQ китайских детей составила 122 балла, или на 1,33dвыше, а время реакции было на 0,96dвыше, чем у британских детей. В строке 3 даны результаты другого исследования детей этнических китайцев в Калифорнии в сравнении с 77 детьми европейцев. Величина IQ китайских детей была на 6 баллов, или на 0,40dвыше, а среднее время реакции по трём тестам — на 0,25dвыше, чем у белых. Результаты всех трёх исследований показывают, что преимущество детей аборигенов Восточной Азии во времени реакции, отражающим неврологическую эффективность мозга при простой обработке информации, составляет приблизительно две трети от их преимущества в интеллекте. В этих исследованиях также сообщается о различиях в вариабельности времени реакции и во времени движения, они имели приблизительно такую же величину, что и время реакции. Дженсен (Jensen, 1998) даёт более детальное описание и обсуждение этих исследований. Джири и соавт. (Geary et. al., 1997) также сообщали о более быстрой реакции и более высоком IQ китайских детей в
156
Ричард Линн
сравнении с американскими, но они не приводят стандартных отклонений, ввиду чего различие не может быть выражено в величинах d.
Таблица 10.6. Различия между аборигенами восточной Азии и европейцами во времени реакции

Сокращения: N — число испытуемых; IQ — коэффициент интеллекта; IQd— коэффициент интеллекта в единицах стандартного отклонения; RTd— время реакции в единицах стандартного отклонения.
Зрительная память
В тестах интеллекта зрительная память обычно не оценивается. Было выполнено четыре исследования зрительной памяти японцев, результаты которых представлены в Таблице 10.7.
Таблица 10.7. Различия между аборигенами Восточной Азии и европейцами в зрительной памяти

В строке 1 приведён IQ 5-10-летних японских детей (107 баллов по данным теста MFFT), значение вычислено по числу неправильных ответов в сравнении с американской выборкой (п=2.676). В тесте MFFT испытуемым показывают рисунки ряда объектов (например, лодка, курица и т. д.), к которым следует подобрать идентичный рисунок из нескольких, очень похожих друг на друга. Задача состоит в запоминании деталей рисунка, чтобы найти точно совпадающий. Результаты выполнения этой задачи кор
Расовые различия в интеллекте
157
релируют на уровне 0,38 со шкалой действия WISC (Plomin, Buss, 1973), так что это слабый тест как способности к визуализации и общего интеллекта, так и зрительной памяти. В строке 2 дан IQ зрительной памяти в 105 баллов для этнических японцев в Америке в сравнении с американскими европеоидами. Этот результат получен в помощью двух тестов зрительной памяти, заключавшихся в показе испытуемым в течение 25 секунд 20 объектов, которые затем убирали. Задача состояла в запоминании и восстановлении их расположения. В строке 3 дан IQ зрительной памяти в 110 баллов, полученный при сравнении выборки японских учащихся старших классов средней школы и студентов университета с выборкой европейских студентов (п=52) из университетского колледжа г. Дублина. В строке 4 дан IQ зрительной памяти в 113 баллов по субтестам зрительного воспроизведения пересмотренных шкал памяти Векслера, полученный в японской выборке стандартизации этого теста в сравнении с американской выборкой стандартизации. Для выполнения теста необходимо нарисовать по памяти геометрические узоры, которые показывают испытуемым в течение 10 секунд. По мнению авторов, объяснение превосходства японцев может состоять в том, что японские дети изучают японское идеографическое письмо кандзи, что развивает зрительную память. Однако, эта гипотеза очевидно опровергается исследованием Флайерти и Коннолли (Flaherty, Connolly, 1996), результаты которых приведены в строке 2. Некоторые участники исследования, американские этнические японцы, владели кандзи, в то время как другие не владели, но между владевшими и не владевшими кандзи не было выявлено различия в зрительной памяти. Это опровергает теорию, что преимущество аборигенов Восточной Азии в задачах визуализации является результатом их практики в визуализации идеографического письма.
Размер мозга
Исследования различий в размере мозга у аборигенов Восточной Азии и европейцев приведены в Таблице 10.8. Средние значения и стандартные отклонения объёма мозга выражены в кубических сантиметрах.
В строке 1 даны результаты, вычисленные Гулдом (Gould, 1981) на материале коллекции черепов, собранных в начале девятнадцатого столетия американским врачом Сэмюэлем Мортоном (Morton, 1849), распределившим их по расам и вычислившим их средние объёмы. Гулд обвинил Мортона в ошибках и повторно измерил черепа, показав, к своему удовлетворению, что объём мозга аборигенов Восточной Азии и европейцев одинаков. Но число черепов было очень невелико, а именно 10 черепов
158
Ричард Линн
Таблица 10.8. Различия размер мозга (см3) у европейцев и аборигенов Восточной Азии

аборигенов Восточной Азии и 52 — европейцев. Вес этих данных слишком мал для того, чтобы придавать им большое значение, они приведены здесь преимущественно из исторического интереса. В строке 2 даны результаты для наибольшей из когда-либо собранных коллекции черепов (около 20 тысяч), показывающие, что объём мозга аборигенов Восточной Азии на 1,2d(единицы стандартного отклонения) больше, чем у европейцев. В строке 3 приведено различие в 20 см3 по результатам исследования американских 7-летних детей, выполненного Броманом и соавт. (Broman et al., 1987). Размеры мозга были вычислены по их данным Раштоном (Rushton, 1997). В строке 4 даны результаты, полученные по собранным Юргенсом, Ауне и Пипером (Jurgens, Aune, Pieper, 1990) данным для многих тысяч 25-45-летних. Значения в таблице были скорректированы на размер тела Раштоном (Rushton, 2000). В строке 5 даны результаты для массива данных, собранных Гровсом (Groves, 1991), объединившим оценки объёмов мозга 36 выборок мужчин (из представленых Куном, Молна- ром, Мартином и Селл ером значений). Эти размеры мозга больше, чем представленные в строке 2 ввиду того, что это — данные для мужчин, но различие между европейцами и аборигенами Северо-Востока Азии было примерно таким же, хотя и немного больше. В строке 6 даны результаты Раштона по размеру мозга с поправкой на размер тела для 6.325 военнослужащих в Соединенных Штатах. В строке 7 приведены рассчитанные Раштоном суммарные данные о размере мозга по большому числу массивов данных с поправкой на размер тела. Таким образом, во всех исследованиях (кроме приведённого в строке 1 исследования Мортона, пересмотренного Гулдом) обнаружено, что у аборигенов Восточной Азии средний объём мозга больше, чем у европейцев.
Расовые различия в интеллекте
159
Наследуемость интеллекта у аборигенов Восточной Азии
Опубликовано всего лишь одно исследование наследуемости интеллекта у аборигенов Восточной Азии (Lynn, Hattori, 1990). В этой работе изучались корреляции между объединёнными показателями 23 тестов для 543 пар идентичных и 134 пар неидентичных близнецов в возрасте 12 лет. Корреляция составила 0,782 для идентичных и 0,491 для неидентичных близнецов. Наследуемость, рассчитанная путём удвоения разницы между этими двумя корреляциями, составляет 0,582. После внесения поправки на надёжность тестов, принимая коэффициент надёжности за 0,9 (Bouchard, 1993), наследуемость принимает значение 0,65. Это почти совпадает со значением наследуемости у европейцев такого же возраста — см. Параграф 4 Главы 3. Резонно предположить, что степень наследуемости интеллекта у европейцев и аборигенов Восточной Азии приблизительно одинакова.
Средовые и генетические объяснения IQ аборигенов Восточной Азии
Устойчиво высокие величины IQ аборигенов Восточной Азии в местах их исконного проживания в Восточной Азии, а также в Европе и на обоих материках Америки, представляют проблему для сторонников сре- довой теории. Им достаточно легко объяснить низкие показатели IQ африканцев, которые они приписывают бедности, плохому образованию, смещённости тестов и расизму. Но ни одна из этих причин не может объяснить более низкий IQ европейцев по сравнению с аборигенами Восточной Азии. Сторонники средовой теории избрали три стратегии, чтобы справиться с этой проблемой. Первая — проигнорировать её. Такое решение проблемы избрано в большинстве общих учебников и в специальных монографиях на тему расы и интеллекта Фиша (Fish, 2002), Гулда (Gould, 1996) и Монтагю (Montagu, 1999). Вторая стратегия состоит в том, чтобы поставить под сомнение или преуменьшить проблему. Так, вскоре после опубликования данных первого исследования, выявившего высокий IQ японцев по тесту WISC-R, Стивенсон и Азума (Stevenson, Azuma, 1983) утверждали, что в японской выборке стандартизации были недостаточно представлены группы с низким IQ. Позже, когда были опубликованы другие работы, подтверждающие высокий IQ японцев, стало невозможно оспаривать это, и защитники средовой теории начали утверждать, что различие было лишь
160
Ричард Линн
незначительным. Например, Броди (Brody, 2000, стр. 219) пишет об исследованиях, в которых было найдено, что интеллект в Японии выше, чем в Соединенных Штатах: «Свидетельств в пользу того, что имеются большие различия в IQ этих групп, мало, или их вовсе нет». Он не уточняет, что он подразумевает под словом «большие». В сходном духе Макинтош (Mackintosh, 1998, стр. 168) пишет: «Нет никаких убедительных причин считать, что китайцы или японцы серьёзно превосходят белых в тестах интеллекта». Он не детализирует, что он подразумевает под «серьёзно». Третья стратегия, принятая защитниками средовой теории, состоит в утверждении, что даже если допустить, что аборигены Восточной Азии имеют более высокий в сравнении с европейцами IQ, «нет данных для решения вопроса, имеют ли такие различия средовое или генетическое происхождение» (Mackintosh, 1998, стр. 168).
Вопреки этому утверждению, приведённые в этой главе исследования указывают на сильную генетическую детерминацию более высокого IQ аборигенов Восточной Азии по сравнению с европейцами.
Во-первых, IQ аборигенов Восточной Азии постоянно оказывается выше европейского во многих странах, включая Китай, Японию, Гонконг, Сингапур и Южную Корею (см. Таблицу 10.1).
Во-вторых, высокие оценки IQ, получаемые аборигенами Восточной Азии в их собственных странах, в целом подтверждаются исследованиями вне Восточной Азии, результаты которых приведены в Таблице 10.2. Медиана значений IQ выходцев из Восточной Азии в Соединенных Штатах составила по всем исследованиям 101 балл — немного ниже, чем 105 баллов аборигенов Восточной Азии, проживающих в своих собственных странах. Для этого есть две возможные причины. Первая: величина IQ тех, кто мигрировал в Соединенные Штаты, могла быть немного ниже среднего значения тех, кто остался в Азии. Другой причиной является то, что иммигранты первого и второго поколений обычно продолжают общаться на своём собственном языке, а английский является для них вторым языком. Это может осложнять им выполнение тестов, требующих знания языка. Средний IQ по последним шести приведённым в Таблице 10.2 исследованиям, выполненным с 1990 г, составил 105 баллов, такой же, как у аборигенов Восточной Азии в странах Восточной Азии.
Из Таблицы 10.3 видно, что аборигены Восточной Азии постоянно получают немного более высокие средние оценки IQ, чем европейцы, проживающие в сходных условиях. В Бразилии IQ этнических японцев составляет 99 — на 4 балла выше, чем у европейцев (см. Таблицу 3.2), хотя японцы были ввезены в страну в качестве сельскохозяйственных чернорабочих после отмены рабства в 1888 г. и вряд ли могли иметь более высокие коэффициенты интеллекта, чем население Японии в целом. В Великобритании выходцы из Восточной Азии получили оценку IQ 107 баллов, а в Нидер
Расовые различия в интеллекте
161
ландах — 102 балла. В Малайзии их IQ составил 99 баллов — на 10 баллов выше, чем у автохтонных малайцев.
В-третьих, приверженцы средовой теории не предлагают какого-либо объяснения устойчиво высокого IQ аборигенов Восточной Азии, и сомнительно, что какое-нибудь правдоподобное средовое объяснение может быть найдено. Интеллект подвержен влиянию уровня жизни, но уровень жизни многих аборигенов Восточной Азии ниже, чем у европейцев. Уровень жизни аборигенов Восточной Азии в Японии, Гонконге и Сингапуре сопоставим с уровнем жизни европейцев в Северной и Западной Европе, Соединенных Штатах, Канаде, Австралии и Новой Зеландии, но жизненный уровень в Китае, Южной Корее и на Тайване существенно ниже, и однако же IQ в этих странах приблизительно на 5 баллов выше, чем у европейцев. Различие наблюдается повсюду и нет никакого правдоподобного средо- вого объяснения превосходства аборигенов Восточной Азии.
В-четвертых, все шесть приведённых в Таблице 10.4 исследований интеллекта корейских младенцев, усыновлённых европейскими семьями в Европе и в Соединенных Штатах, показывают, что IQ этих детей выше, чем у европейцев, в среде которых они выросли. Кажется невероятным, что эти младенцы, отданные на усыновление, были селективной выборкой с IQ выше среднего. Однако следует отметить, что возраст этих детей был весьма мал и они, вероятно, были усыновлены преимущественно семьями среднего класса, которые обеспечили им некоторые «средовые преимущества». Трудно оценить, насколько велик мог быть этот эффект, но маловероятно, что он мог достигать 11 баллов IQ для четырёх выборок приёмных детей из Восточной Азии, получавших полноценное питание. В исследовании Вайнберга, Скарр и Вальдмана (Weinberg, Scarr, Waldman,
, рассматривавшемся в Параграфе 13 Главы 4, было показано, что чернокожие младенцы, усыновлённые белыми семьями среднего класса, получили оценку IQ 95 баллов в 7 лет и 89 баллов в 17 лет. Это свидетельствует о том, что эффект среды в 7-летнем возрасте составлял 6 баллов IQ. Если применить это же эмпирическое правило к приёмным детям аборигенов Восточной Азии, то средний IQ в выборках детей, получавших полноценное питание (111 баллов), должен быть уменьшен на 6 баллов, что даёт истинный IQ в 105 баллов — точно такой же, как у аборигенов Восточной Азии в странах их исконного проживания.
В-пятых, более быстрая реакция детей аборигенов Восточной Азии (см. Таблицу 10.6) указывает на то, что их нервная система более эффективно обрабатывает информацию, что вносит существенный вклад в их более высокий IQ. И вновь это превосходство во времени реакции не может быть правдоподобно объяснено средовой теорией.
В-шестых, в нескольких из приведённых в Таблице 10.8 исследований показано, что средний размер мозга у аборигенов Восточной Азии выше,
6 Ричард Линн
162
Ричард Линн
чем у европейцев. Размер мозга является важной неврологической детерминантой интеллекта и имеет высокую наследуемость. Сомнительно, возможно ли вообще какое-либо средовое объяснение большего размера мозга у аборигенов Восточной Азии. Как можно оценить, больший размер мозга должен дать им преимущество в 10 баллов IQ. Этого преимущества в IQ за счёт большего мозга следует ожидать в тех случаях, когда они с детства живут в одинаковых с европейцами условиях. Так как IQ европейцев оценивается в 99 баллов, а аборигенов Восточной Азии в 105 баллов, фактическая разница IQ составляет 6 баллов. Может быть предложено два возможных объяснения того, почему реальное преимущество аборигенов Восточной Азии составляет лишь 6 баллов IQ, в то время как предсказанное — 10 баллов. Во-первых, часть большего мозга аборигенов Восточной Азии может обеспечивать их лучшую зрительную память, не измеряемую тестами интеллекта. Во-вторых, испытуемые многих выборок были зачаты и выросли в худших по сравнению с европейцами условиях, что может несколько снижать их IQ. Если это так, то можно предсказать, что когда условия жизни в Восточной Азии сравняются с европейскими, разница в IQ достигнет 10 баллов и станет полностью объяснима различием в размере мозга.
ш т11
МПНШМ 4MP044J
Интеллект арктических народов
Зрительная память
Размер мозга
Генотипный интеллект арктических народов
Арктические народы представлены автохтонными инуитами (ранее называемыми эскимосами) Аляски, северного побережья Канады, о-ва Гренландия, алеутами Алеутских островов, а также северными тюркскими народами и чукчами крайнего Северо-Востока Азии. Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994) выделели их в отдельный генетический «кластер» в своей классификации народов, основанной на ряде генетических маркеров. Арктические народы отличаются генетически от америндов (американских индейцев) высокой частотой группы крови В, не встречающейся у америндов. Они отличаются от америндов и от аборигенов Восточной Азии тем, что существенно лучше приспособлены к холоду. У них более короткие ноги и руки и широкое туловище, что способствует сохранению тепла, более развит эпикантус, нос более уплощённый, что уменьшает риск обморожения. Причина, по которой арктические народы развились в отдельную расу, состоит в том, что их предки были изолированы от аборигенов Восточной Азии горным хребтом Черского в Северо-Восточной Азии. Инуиты отделились от чукчей Северо-Востока России, мигрировав через Берингов пролив в Северную Америку приблизительно за 10-11 тыс. лет до нашей эры. Несколько их доисторических стоянок были обнаружены в долине реки Ненана в центральной Аляске, где находки датируются возрастом от 11.300 до 10.000 лет (Dixon, 1999). В середине двадцатого столетия насчитывалось приблизительно 50.000 инуитов и приблизительно 5.600 жителей Алеутских островов
Интеллект арктических народов
Данные исследований интеллекта арктических народов приведены в Таблице 11.1.
164
Ричард Линн
Таблица 11.1. Оценки IQ арктических народов

В строке 1 представлен IQ в 89 баллов по результатам первого исследования большой выборки инуитских детей, тестировавшихся с помощью теста «нарисуй человека» Гудинаф (DAM). В строке 2 дан IQ 92 для выборки алеутских детей, также протестированных с помощью теста DAM тем же автором. Это единственное исследование интеллекта алеутских детей. Все остальные исследования проведены среди инуитов. В строках 3 и 4 представлены результаты проведённой в 1962 г. оценки IQ детей инуитов в репрезентативных выборках учащихся начальной и средней школ на Юконе и на Северо-Западных территориях Канады. Ученики начальной школы получили оценку IQ в 94 балла, а учащиеся средней школы — 84 балла. В строке 5 приведён самый низкий из всех IQ в 78 балла, полученный на выборке молодых людей. Он настолько отличается от других результатов, что его следует считать сомнительным. В строке 6 дан IQ способности к визуализации взрослых инуитов в 93 балла. В строке 7 приведён IQ 91 для 10-летних испытуемых. В строке 8 приведён IQ в 90 баллов, полученный Верноном с помощью матричного теста, словарного теста и методики «кубики Кооса». Величина IQ по словарному тесту (80 баллов) могла оказаться заниженной по причине того, что дети, возможно, говорили дома на своем родном языке. Величина IQ в 90 баллов для рассуждения была взята как наиболее разумная оценка общего интеллекта. В этом исследовании Вернон также использовал тест DAM, по которому инуитс- кие дети получили IQ в 95 баллов. Это более или менее согласуется с ре
Расовые различия в интеллекте
165
зультатами, представленными в строках 1 и 2, где дети получили по тесту DAM оценки IQ 89 и 92 балла.
В строке 9 представлен IQ в 91 балл для значительной выборки 6-12летних детей, полученный с помощью шкалы действия WISC; эти дети получили намного более низкие оценки вербального IQ, но они не говорили на английском как на перовом языке и их низкий вербальный IQ не может считаться валидным. В строке 10 дан IQ рассуждения в 96 баллов для выборки 9-12 летних детей. В строке 11 приведены оценки вербального IQ и IQ действия в 78 и 93 балла для небольшой выборки 7-летних детей, обследованных с помощью теста WPPSI. Английский являлся для детей вторым языком, ввиду чего их вербальный IQ оказался заниженным, поэтому в качестве наилучшей меры общего интеллекта был взят IQ действия, равный 93 баллам. В строке 12 приведён IQ рассуждения в 95 баллов для выборки 7-10-летних детей. В строке 13 дан IQ 91 для значительной выборки 7-14-летних испытуемых, полученный с помощью шкалы действия WISC-R. Вербальную шкалу не использовали, так как английский не был первым языком испытуемых. В строке 14 приведён IQ 92 для 5летних инуитов из арктической зоны Квебека. Авторы заявляют, что результаты у инуитских детей были выше, чем у американских, но это вызвано тем, что американские нормы снижены из-за включения в них этнических меньшинств и тем, что авторы не делали поправки на долговременное увеличение тестовых оценок (детей также повторно тестировали в возрасте 6 и 7 лет, в этих тестированиях их результаты были выше, что можно приписать эффекту тренировки). В строке 15 дан IQ невербального рассуждения в 86 баллов и словарный IQ в 77 баллов для 15-летних инуитов на Аляске.
Медианное значение IQ по всем исследованиям составило 91 балл, что мы предлагаем рассматривать как наилучшую оценку интеллекта арктических народов. Оценки IQ визуализации оказались несколько выше оценок вербального IQ, как показано в работе Вернона (строка 8), где значения IQ визуализации и вербального IQ составили 88 и 80 баллов, и в исследовании Тейлора и Сканеса (строка 11), где IQ визуализации составил 93, а вербальный IQ — 78 баллов. Усреднение этих двух результатов даёт оценку IQ визуализации, превышающую оценку вербального IQ на 11 баллов. Этот паттерн высокой способности к визуализации и низкой вербальной способности наличествует также у родственных арктическим народам аборигенов Восточной Азии и у американских индейцев. Представляется, что не было тенденции увеличения интеллекта инуитов за приблизительно 60-летний период со времени первого исследования начала 1930-х гг. (Eells, 1933), в котором их IQ был оценен в 89 баллов, до 90-х гг., когда Вилгош и соавт. (Wilgosh et al., 1986) получили оценку IQ в 91 балл.
166
Ричард Линн
Зрительная память
У инуитов чрезвычайно хорошая зрительная память, которая не измеряется стандартными тестами интеллекта. Это было показано Кляйн- фельд (Kleinfeld, 1971) в исследовании зрительной памяти у 125 инуитских детей возраста 9-16 лет из сельской местности Аляски в сравнении с 501 белым ребёнком из Анкориджа и Фэрбенкса, двух основных городов Аляски. Тест заключался в показе детям рисунков в течение короткого промежутка времени, после чего они получали задание нарисовать их по памяти. Инуитские дети получили средний IQ в 106 баллов относительно среднего значения белых в 100 баллов. Кляйнфельд отмечает, что этот результат согласуется с наблюдениями путешественников, сопровождавших инуитов в длительных охотничьих экспедициях. Она пишет, что: «Кавка- зоиды, путешествовавшие с инуитами, часто отмечают их экстраординарную способность находить дорогу в, казалось бы, лишённой примет местности, внимательно рассматривая мельчайшие ориентиры и запоминая их пространственное расположение».
Хорошая зрительная память инуитов может объяснить, почему они относительно успешны в правописании. В исследовании Вернона показано (Vernon, 1969), что 10-летние инуиты имели IQ правописания в 95 баллов, что значительно выше их вербального IQ (80 баллов), компонентом которого обычно считают правописание (Carroll, 1993). Хорошая зрительная память способствует правописанию, так как позволяет запомнить начертание слов. Вероятно, именно поэтому женщины существенно успешнее в правописании, чем мужчины (Lynn, 1992): у них лучше зрительная память (Halpem, 2000; Kimura, 2002).
Вероятно, что хорошая зрительная память инуитов имеет генетическую основу. Осборн и Грегор обнаружили (Osborne, Gregor, 1966), что зрительная память имеет высокую наследуемость. Даже у 9-летних инуитских детей зрительная память была значительно лучше, чем у европейцев, и кажется маловероятным, что дети столь юного возраста приобрели бы эту выраженную способность посредством обучения, даже если это было бы возможно. Наиболее вероятное объяснение хорошей зрительной памяти у инуитских детей состоит в том, что эта способность развивалась генетически посредством естественного отбора в силу необходимости для живущих в Арктике людей запоминать мельчайшие детали ландшафта, чтобы найти обратный путь, возвращаясь с продолжительной охоты. Пейзаж промёрзшей тундры имеет мало ориентиров, ввиду чего охотники должны были замечать и запоминать имеющееся небольшое количество при
Расовые различия в интеллекте
167
знаков. Хорошая зрительная память характерна также для аборигенов Восточной Азии (IQ=107; Глава 10, параграф 7) и американских индейцев, у которых Ломбарди(Lombardi, 1970) оценил IQ зрительной памяти в 104 балла, что очень близко к IQ инуитов (106 баллов по данным Кляйн- фельд). Возможно, в предковой популяции населения Северо-Восточной Азии хорошая зрительная память развилась до того, как оно разделилась на аборигенов Восточной Азии, американских индейцев и арктические народы. Хорошая зрительная память инуитов имеет параллель у австралийских аборигенов, о которых сообщает Кеаринс (Kearins, 1981); у последних это объясняется адаптацией к проживанию в пустынях с малым количеством ориентиров, подобных в этом отношении промёрзшей тундре Арктики (см. Главу 8).
Размер мозга
Удалось найти всего одно исследование размера мозга арктических народов. Смит и Билс (Smith, Beals, 1990) приводят размеры мозга для десяти популяций, среднее значение по которым составило 1.444 см3. Размер мозга европейцев, представленный ими, составил 1.368 см3. Различие в 76 см3 является существенным. Имеется связь размера мозга с интеллектом индивидов, такую же связь следует ожидать и для групп. Больший размер мозга арктических народов позволяет ожидать, что они будут иметь более высокие в сравнении с европейцами оценки IQ, но это не так.
Есть два вероятных объяснения этой аномалии. Во-первых, какая-то часть большого мозга арктических народов, вероятно, обслуживает их хорошую зрительную память, обнаруженную Кляйнфельд (Kleinfeld, 1971) и рассмотренную в Параграфе 2. Во-вторых, размер мозга - не единственная детерминанта интеллекта. Некоторые нейрофизиологические процессы, ответственные за более высокий интеллект, могли развиться у европейцев в результате генетических мутаций, не появившихся у арктических народов. Причина этого, вероятно, в том, что европейцы были намного более многочисленны и, соответственно, имели больше шансов на благоприятные мутации, обуславливающие более высокий интеллект.
Генотипный интеллект
Кажется вероятным, что и генетические, и средовые факторы вносят вклад в низкий IQ арктических народов. Имеются две группы фактов, говорящих о некоторой генетической детерминации. Во-первых, как отмечено
168
Ричард Линн
в Параграфе 1, IQ арктических народов не претерпел увеличения относительно европейцев с начала 1930-х гг., хотя условия их жизни улучшились, поскольку во второй половине двадцатого столетия они получали увеличенные социальные пособия и лучшее образование. Если бы интеллект арктических народов был снижен неблагоприятными условиями жизни в 1930-х гг., то он должен бы был увеличиться к началу 1980-х гг. Во-вторых, во всех исследованиях, приведённых в Таблице 11.1, дети арктических народов учились в школе и, значит, были знакомы с процедурами тестирования, следовательно, нет причин предполагать, что в этом отношении они были «ущербны».
Г(1Ш 12 ШИШМ 1ЩЩ41
Интеллект американских индейцев в Северной Америке
Коэффициент интеллекта по тесту «нарисуй человека»
Латинская Америка
Зрительная память
Гибриды американских индейцев с европейцами
Музыкальные способности
Размер мозга
Испаноговорящие в Соединённых Штатах
Генотипный интеллект американских индейцев
Автохтонные американцы, называемые также индейцами, являются коренным населением обоих континентов Америки, их предки мигрировали с крайнего Северо-Востока Азии через Берингов пролив на современную Аляску. Они — одна из больших рас в таксономиях классических антропологов: Линнея (Linnaeus, 1758), Блюменбаха (Blumenbach, 1776), а также Куна, Гарна и Бёрдселла (Coon, Gam, Birdsell, 1950). Кавалли-Сфорца, Меноцци и Пиацца (Cavalli-Sforza, Menozzi, Piazza, 1994) подтвердили, что американские индейцы образуют особый генетический «кластер», что отделяет их от других народов. Наиболее характерными особенностями американских индейцев, отличающими их от аборигенов Восточной Азии, является более тёмная, иногда красноватая кожа, крючковатый или прямой нос и отсутствие присущего аборигенам Восточной Азии полного эпикантуса, хотя складка на внутренней части века иногда имеется. В Соединенных Штатах по переписи населения 1990 г. проживало приблизительно два миллиона индейцев. Примерно половина из них жила в резервациях или в их окрестностях. В Центральной и Южной Америке в конце двадцатого столетия насчитывалось приблизительно 70 миллионов индейцев и около 140 миллионов метисов, имеющих индейских и европейских предков.
170
Ричард Линн
Интеллект американских индейцев в Северной Америке
Интеллект американских индейцев в Соединенных Штатах начал изучаться в 1920-х гг., а с 1960-х гг. начали публиковаться подобные исследования индейцев в Канаде. Результаты этих исследований приведены в Таблице 12.1.
Таблица 12.1. Оценки IQ американских индейцев в Северной Америке

В строке 1 приведён IQ в 86 баллов из первого исследования интеллекта американских индейцев в Соединенных Штатах, опубликованного ещё в 1922 г. В строке 9 дан IQ невербального рассуждения в 91 балл, полученный у 8-17-летних испытуемых в работе Колемана — наиболее массовом из когда-либо опубликованных исследований, посвящённых изучению способностей индейцев — учащихся школ. Их вербальный IQ был немного ниже (87 баллов).
Медиана результатов 21 исследования составляет 86 баллов IQ. Американские индейцы получали более высокие оценки способности к визуа
Расовые различия в интеллекте
171
лизации в сравнении с вербальной способностью во всех шести исследованиях, в которых тестировались обе эти способности. Медианное значение IQ визуализации в этих исследованиях составляет 89,5, а медианное значение вербального IQ — 81 балл. Такой же профиль высокой способности к визуализации и низкой вербальной способности обнаруживается и у аборигенов Восточной Азии (см. Главу 10, параграф 1), с которыми американские индейцы генетически близко родственны.
Коэффициент интеллекта по тесту «Нарисуй человека»
Было проведено несколько исследований интеллекта американских индейцев с помощью теста Гудинаф «Нарисуй человека» (DAM). Поскольку это невербальный тест, то, стало быть, обходится проблема, состоящая в том, что испытуемые ряда выборок индейцев не говорили по-английски как на первом языке. Поэтому целесообразно рассмотреть эти исследования отдельно. Тест DAM был разработан Флоренс Гудинаф (Goodenough, 1926а, 1926b) и представляет собой задание нарисовать мужчину и женщину. Рисунки оценивают по присутствию деталей, таких как уши, брови и т. д. Оценки по тесту DAM коррелируют с показателями других общепринятых тестов интеллекта со значением порядка 0,40-0,60. Например, Абель, Вуд и Либман (Abell, Wood, Liebman, 2001) на основании исследования у 100 детей сообщают, что корреляция данных теста DAM с тестами WISC- R и WISC-III составила 0,46 и 0,35 для вербального IQ, 0,57 и 0,48 для IQ действия, и 0,54 и 0,45 для общего IQ. В исследовании, в котором тест DAM предъявляли выборке из 217 10-летних детей наряду с рядом других тестов (тесты словарной, пространственной и перцепцтивной способностей, а также способности к рассуждению), тест DAM имел нагрузку 0,48 по первой главной компоненте, в то время как нагрузки других когнитивных тестов имели значения в диапазоне от 0,58 до 0,70 (Lynn, Wilson, Gault, 1989). Таким образом, тест DAM — адекватная, хотя не сильная мера фактора g, и, по-видимому, является скорее мерой способности к визуализации, чем вербальной способности. Причина этого вероятно в том, что прежде чем нарисовать человеческое тело, ребенок должен визуализировать его. Результаты исследований IQ индейцев Северной Америки с помощью теста DAM представлены в Таблице 12.2. Медианное значение IQ составляет 90 баллов. Это почти совпадает с медианой IQ визуализации (89,5 баллов) по данным исследований, приведённых в Таблице 12.1, и согласуется с интерпретацией DAM как теста, оценивающего преимущественно способность к визуализации.
172
Ричард Линн
Таблица 12.2. Значения IQ американских индейцев по тесту «Нарисуй человека»

Латинская Америка
Результаты исследований интеллекта американских индейцев в Латинской Америке приведены в Таблице 12.3.
Таблица 12.3. Значения IQ индейцев в Латинской Америке

В строке 1 дан IQ в 84 балла, полученный по тесту количественного рассуждения у 4-летних детей в Колумбии, про которых сказано, что они были «приблизительно одинаково распределены по группам социальноэкономического положения» (стр. 172). Они сравнивались с группой из
Расовые различия в интеллекте
173
156 американских детей, которую характеризовали как репрезентативную для Соединенных Штатов. Население Колумбии состоит на 75-процентов из индейцев и метисов, на 20 процентов из европейцев и на 5 процентов из африканцев. Резонно предположить, что более высокий IQ европейцев и более низкий IQ африканцев приблизительно уравновесят друг друга, и что IQ в 84 балла репрезентативен для интеллекта индейцев. В строках со второй по пятую представлены четыре значения IQ для Эквадора (89, 88, 80, и 91 балл). Значение IQ в 91 балл, приведённое в строке 5, получено у 8летних детей народности кечуа в двух деревнях, некоторые из испытуемых были чистокровными индейцами, а другие имели смешанную расовую родословную. В строке 6 дан IQ в 79 баллов для Гватемалы. В строках 7,8, и 9 приведены три значения IQ для Мексики (87,92 и 83 балла), последнее значение получено у индейцев в Баха-Калифорнии. В строках 10 и 11 даны IQ в 87 и 85 баллов для индейцев в Перу.
Величины IQ лежат в диапазоне 79-92 баллов и достаточно сходны, учитывая диапазон стран, в которых были сформированы выборки. Медиана IQ по всем исследованиям составила 86 баллов — такая же, как у индейцев Северной Америки (по данным исследований, приведённых в Таблице 12.1). Поэтому наилучшей оценкой IQ американских индейцев в Северной и в Южной Америке будет 86 баллов.
Зрительная память
Зрительная память — способность, обычно не оцениваемая тестами интеллекта. По некоторым свидетельствам, у американских индейцев эта способность хорошо развита. Ломбарди в своём исследовании (Lombardi, 1970) сравненивал 80 индейских детей с 80 белыми детьми возраста 6-8 лет с помощью Иллинойского теста психолингвистических способностей и обнаружил, что у индейцев вербальный IQ составлял 73, a IQ визуализации — 93 балла. Величина IQ визуализации определялась по сумме оценок шести субтестов способностей к визуализации, по одной из которых, зрительной памяти, IQ индейцев составил 104 балла. Этот субтест был единственным, по которому индейцы получили более высокую оценку, чем белые.
Хорошей зрительной памятью американских индейцев может объясняться то, почему они относительно успешны в правописании. В исследовании академической успеваемости приблизительно 13.000 индейских детей в 11 штатах США было найдено, что они, как оказалось, продемонстрировали наихудшие результаты по словарному запасу при чтении, вероятно потому, что многие из них говорили на английском как на втором языке, а наилучшие — в правописании (Coombs, 1958). Хорошая зрительная память
174
Ричард Линн
помогает правописанию, так как даёт возможность запомнить начертание слов. Это согласуется с тем фактом, что женщины имеют относительно лучшую зрительную память в сравнении с мужчинами (Halpem, 2000) и более успешны в правописании (Lynn, 1992).
Гибриды американских индейцев с европейцами
Было проведено всего несколько исследований, в которых интеллект чистокровных американских индейцев сравнивался с интеллектом гибридов индейцев с европейцами. Эти исследования показали, что гибриды получают более высокие средние оценки IQ, чем чистокровные индейцы. Результаты исследований представлены в Таблице 12.4. В строке 1 приведена намного более низкая оценка IQ чистокровных индейцев (67 баллов), чем у гибридов (93 балла), но IQ чистокровных индейцев следует считать заниженным, так как тест Отиса — вербальный тест, а индейцы говорили на английском как на втором языке. В работе нет информации о первом языке гибридов. В этом исследовании гибриды были поделены на имевших четверть, половину, три четверти индейской крови и чистокровных индейцев. Была выявлено, что корреляция между долей белой родословной и оценкой IQ составляет 0,41. В строке 2 даны IQ в 89 баллов для гибридов и в 86 баллов для небольшой выборки испытуемых, у которых доля индейской крови составляла от 80 до 100 процентов. В строке 3 даны IQ 94 для метисов и 83 для чистокровных индейцев в Мексике.
Таблицы 12.4. Величины IQ гибридов американских индейцев с европейцами

Музыкальные способности
Простые музыкальные способности, такие как определение изменения высоты тона и запоминание мелодий коррелируют с интеллектом и могут считаться компонентом интеллекта (Carroll, 1993). Ввиду этого интересно выяснить, являются ли музыкальные способности американских индейцев плохими в соответствии с их низким IQ. Единственное исследо
Расовые различия в интеллекте
175
вание этой проблемы — работа Гарта (Garth, 1931), в которой приводятся результаты оценки определения высоты тона и запоминания мелодий в выборке 757 школьников. Их MQ (Музыкальный коэффициент), основывающийся на этих двух тестах, составил приблизительно 92 балла — несколько выше, чем их IQ, оцененный в Параграфе 1 (86 баллов). Однако, в тесте на ритм они были успешнее белых учащихся, получив оценку MQ приблизительно в 104 балла. В этом отношении американские индейцы сходны с африканцами, которые также получают в тестах на ритм более высокие по сравнению с белыми оценки (см. Главу 4). Не известно, связана ли способность оценивать ритм с интеллектом, и нет очевидного объяснения этой способности американских индейцев и африканцев.
Размер мозга
Результаты исследований размера мозга американских индейцев в сравнении с европейцами приведены в Таблице 12.5. В строке 1 приведены результаты, вычисленные Гулдом (Gould, 1981) на материале коллекции черепов, собранных в начале девятнадцатого столетия американским врачом Сэмюэлем Мортоном (Morton, 1849). Гулд обвинял Мортона в подгонке данных в сторону увеличения объёма мозга европейцев, но, как можно видеть, представленное Мортоном различие практически такое же, как в работе Билса и Смита (строка 2), полученное на материале коллекции, содержащей приблизительно 20.000 человеческих черепов и намного меньше, чем среднее различие, данное Юргенсом, Ауне и Пипером (Jurgens, Aune, Pieper, 1990) для нескольких тысяч 25-45-летних (строка 3). Очевидно, что хотя во всех трёх исследованиях размер мозга у европейцев больше, чем у индейцев, величина различия варьирует весьма значительно. В первых двух исследованиях выявленное различие очень мало, но различие в 79 см3, выявленное в третьем исследовании, значительно.
Таблица 12.5. Размер мозга (см3) американских индейцев и европейцев

176
Ричард Линн
Испаноговорящие в Соединенных Штатах
В Соединенных Штатах термин «испаноговорящие» (Hispanics) обозначает выходцев из Латинской Америки и стран Карибского бассейна, говорящих по-испански. Выходцы из Латинской Америки могут быть чисто белыми, чернокожими, гибридами белых и чернокожих, индейцами или метисами, имеющими белых и индейских предков. В Соединенных Штатах насчитывается пять основных групп испаноговорящих. Это выходцы из Мексики, из остальной части Латинской Америки, с Кубы, с Пуэрто-Рико и с других карибских островов. Бюро переписи населения Соединенных Штатов сообщало (United States Bureau of the Census, 1989), что 63 процента испаноговорящих были из Мексики, 13 процентов из Пуэрто-Рико, 10 процентов из Центральной и Южной Америки кроме Мексики, 6 процентов с Кубы и 8 процентов из других мест, главным образом с карибских островов, особенно из Доминиканской республики. Так образом, несомненно, что наибольшая группа — выходцы из Мексики, где 9 процентов населения составляют белые, 60 процентов метисы и 30 процентов — индейцы (Philip’s, 1996). Многие выходцы из остальных частей Латинской Америки также являются метисами. Следовательно, большинство испаноговорящих в Соединенных Штатах — метисы.
Результаты исследований IQ испаноговорящих приведены в Таблице 12.6.
В строках 1 и 2 даны IQ в 89 и 87 баллов для двух первых исследований, проведённых в 1920-х гг. В строке 3 приведён IQ невербального рассуждения в 92 балла, полученный с помощью прогрессивных матриц, теста Лорж-Торндайка и теста копирования фигур Гезелла, а также вербальный IQ в 91 балл для выборки мексиканских детей возраста 6-13 лет в сравнении с 638 белыми. В строке 4 дан IQ невербального рассуждения в 90 баллов для выборки мексиканских детей возраста 6-12 лет в сравнении с 638 белыми, тестировавшимися с помощью Цветных прогрессивных матриц; оценка их IQ по Словарному тесту Пибоди в картинках (PPVT) была несколько ниже (84 балла), возможно отчасти или полностью за счёт того, что некоторые из мексиканских детей использовали английский как второй язык. В строке 5 приведён IQ в 95 баллов для мексиканских детей в Калифорнии в сравнении с 744 белыми детьми. В строке 6 дан IQ в 83 балла, полученный с помощью Цветных прогрессивных матриц в 1972 г у испаноговорящих детей в Калифорнии, выборка охарактеризована как репрезентативная. В строке 7 дан IQ в 94 балла для 6-11-летних, а строке 8 — в 84 балла для 9-12-летних испаноговорящих детей в Техасе.
Таблица 12.6. Показатели интеллекта испаноговорящих в Соединенных Штатах

Расовые различия в интеллекте
178
Ричард Линн
В строке 9 представлены данные из стандартизации теста Стенофорд- Бине 4, согласно которым IQ невербального рассуждения испаноговорящих равен 99 баллам; IQ вербального рассуждения в этой выборке составил 93 балла. В строке 10 дан IQ 87, полученный из данных выборки стандартизации пересмотренного теста PPVT. В строках 11 и 12 даны величины IQ в 84 и 83 балла для Пуэрто-Рико, население которого на 80 процентов состоит из белых, на 8 процентов из чернокожих и на 11 процентов является расово-смешанным (Philip’s, 1996).
В строке 13 дан общий IQ в 93 и вербальный IQ в 85 баллов, полученные в выборке стандартизации теста K-BIT. В строке 14 приведён IQ латиноамериканцев в 86 баллов (в основном по вербальному тесту g — данные Национального лонгитюдного исследования молодежи). В строке 15 приведён IQ 92, полученный в выборке стандартизации теста KAIT. В строке 16 даны значения IQ в 88 баллов для общей способности (g),в 91 балл для вербальной и в 94 балла для пространственной способности у работающих по найму, собранные Службой занятости Соединенных Штатов. В строке 17 дан IQ 91, полученный в выборке стандартизации теста WISC- III. В строке 18 приведён IQ 81 для выборки американцев мексиканского происхождения из шт. Аризона. В строке 19 дан IQ 92, полученный в выборке стандартизации теста WAIS-III. В строке 20 приведён IQ 89, полученный путём мета-анализа данных 39 исследований взрослых испаноговорящих, работающих по найму. Медианное значение IQ результатов исследований с первого по девятнадцатое составляет 89 баллов, что совпадает с результатом мета-анализа, представленным в строке 20.
Генотипный интеллект американских индейцев
Низкий интеллект значительного числа индейцев Южной и Центральной Америки можно отчасти приписать плохому питанию. По оценкам, 21 процент индейских детей «малорослые», то есть имеют задержку роста в результате недостаточного питания, а 30 процентов беременных женщин страдают анемией в результате дефицита железа (De Маеуег и Adiels- Tegman, 1985; UNICEF, 1996). Широко распространённый дефицит йода вызывает высокую встречаемость зоба и кретинизма, что вызывает задержку роста и снижает интеллект. В сельских горных районах Эквадора уровень распространенности кретинизма оценивается на уровне около 7 процентов (Fierro-Benitez, Cazar, Sandoval, 1989). По оценкам, на 1 процент страдающего кретинизмом населения, приходится 3 процента, имеющих какое-либо повреждение головного мозга, вызывающее снижение интеллекта, а у 30 процентов наблюдается общая слабость вследствие гипоти
Расовые различия в интеллекте
179
реоза (Hetzel, 1994). Таким образом, в горной местности Эквадора приблизительно у 21 процента населения интеллект может быть снижен в результате субклинического кретинизма, присутствует также некоторое снижение работоспособности. Может показаться удивительным, что при таких пищевых дефицитах оценки IQ индейцев Южной и Центральной Америки достигают величины 86 баллов. Индейцы в Северной Америке находятся в более благоприятных условиях, так как в Соединенных Штатах и Канаде им обеспечен более высокий уровень жизни, питания и здравоохранения, поэтому может показаться неожиданным, что их IQ является таким же (86 баллов), как у индейцев Южной и Центральной Америки.
Низкий интеллект американских индейцев наиболее обоснованно отнести на счёт действия как генетических, так и средовых факторов. Имеются пять групп свидетельств, указывающих на некоторую генетическую детерминацию. Во-первых, только 20-30 процентов индейцев Южной и Центральной Америки получают недостаточное питание, могущее объяснить их низкий интеллект. Во-вторых, интеллект индейцев в Соединенных Штатах и Канаде не проявил тенденции к повышению соотносительно с европейцами с 1920-х гг., несмотря на значительное улучшение их жизненного уровня и среды обитания. В-третьих, интеллект гибридов индейцев с европейцами коррелирует с долей европейской родословной, как было показано в Параграфе 5. В-четвертых, испаноговорящее население США, в значительной мере являющееся гибридами индейцев и европейцев, обладает уровнем интеллекта, промежуточным между двумя родительскими расами. В-пятых, исследование Кундика, Готтфредсона и Вильсона (Cundick, Gottfredson, Willson, 1974) показало, что у 84 индейских детей, отданных в белые патронатные семьи среднего класса сроком на шесть лет, не было прироста интеллекта. Это демонстрирует, что различные средовые преимущества, связанные с воспитанием в белой семье среднего класса, не оказывают благоприятного воздействия на интеллект американских индейцев и говорит о том, что их IQ до некоторой степени детерминирован генетически.
\(\т п
ЩОД«0СТ4 Н ШНДШ1, М(ОШХ МЭЛНЧНН й И41ТШШЕ
Итоговый обзор расовых различий в интеллекте
Надёжность оценок IQ рас
Валидность оценок Ю рас: Понятия числа
Валидность оценок IQ рас: Успехи в образовании
Валидность оценок IQ рас: Доход на душу населения
и экономический рост
Данные об интеллекте разных рас были подробно изложены в предыдущих главах. В этой главе мы даем суммарный обзор этих различий. Затем мы рассмотрим надежность и валидность представленных значений IQ.
Итоговый обзор расовых различий в интеллекте
В Таблице 13.1 приведены суммарные данные о расовых различиях в интеллекте, которые были изложены в деталях отдельно по каждой расе в Главах 3-12. В таблице дан перечень рас, ранжированных в порядке возрастания уровня их интеллекта, приведено их географическое местоположение, число исследований, число стран, в которых выполнялись исследования, медианные значения и диапазон величин IQ.
В строке 1 приведено медианное значение IQ в 54 балла для бушменов пустыни Калахари, полученное по трём исследованиям, в которых величины IQ колеблются в пределах от 48 до 62 баллов. В строках 2 и 3 даны IQ в 62 и 63 балла для аборигенов Австралии и Новой Гвинеи. Объединение этих двух результатов дает взвешенное значение IQ в 62 балла. В строке 4 дан IQ 67 для африканцев в районах Африке южнее Сахары, вычисленный по результатам 57 исследований. В строке 5 представлен IQ 71 для африканцев с островов Карибского бассейна и из Бразилии, определённый по данным 14 исследований. В строке 6 приведён IQ в 85 баллов для африканцев в Соединенных Штатах, определённый по результатам 29 исследований. В строке 7 дан такой же IQ в 85 баллов для африканцев в Нидерлан-
Расовые различия в интеллекте
181
Таблица 13.1. Расовые различия в интеллекте

182
Ричард Линн
дах по данным 7 исследований. В строке 8 представлен почти идентичный IQ в 86 баллов для африканцев в Великобритании (по данным 18 исследований). Таким образом, африканцы вне Африки стабильно получают более высокие оценки IQ, чем местное население Африки. Этому есть два объяснения. Во-первых, многие африканцы в регионе Карибского бассейна, в Соединенных Штатах, Нидерландах и Великобритании являются межрасовыми гибридами с заметной долей белых предков. Во-вторых, африканцы вне Африки живут в сообществах, руководимых преимущественно европейцами, которые обеспечивают более высокий уровень жизни, что оказывает положительное средовое влияние на IQ африканцев, главным образом за счёт лучшего питания и здравоохранения.
В строках 9-13 приведены величины IQ аборигенов Южной Азии и Северной Африки из разных регионов. В строке 9 дан IQ местного населения Южной Азии и Северной Африки в 84 балла (по результатам 37 исследований в 17 странах). В строке 10 дан IQ 92 для выходцев из Южной Азии в Великобритании, вычисленный по данным 16 исследований. В строке 11 дан IQ 85 для выходцев из Южной Азии в континентальной Европе, полученный по результатам 18 исследований в трех странах. В строке 12 приведён IQ 86 для выходцев из Южной Азии, проживающих в Африке (по 6 исследованиям в двух странах). В строке 13 дан IQ 85 для выходцев из Южной Азии, вычисленный по данным трёх исследований на о-вах Фиджи, в Малайзии и на о. Маврикий. Величины IQ аборигенов Южной Азии и Северной Африки довольно сходны и варьируют в пределах от 84 до 91 балла.
В строке 14 дан IQ в 85 баллов для аборигенов островов Тихого океана, определённый по 14 выборкам, а в строке 15 — в 90 баллов, оцененный по результатам 12 исследований среди народа маори в Новой Зеландии. Более высокий IQ маори в Новой Зеландии обусловлен, вероятно, тем, что уровень их жизни выше в силу того, что они живут в стране, управляемой европейцами. По этой причине IQ в 85 баллов принят нами как наилучшая оценка IQ аборигенов островов Тихого океана.
В строке 16 представлен IQ в 87 баллов для местного населения ЮгоВосточной Азии, определённый по 11 выборкам из 6 стран. В строке 17 дан IQ 93 для выходцев из Юго-Восточной Азии в Соединенных Штатах и Нидерландах. Причина того, что это значение выше IQ, чем в Юго-Восточной Азии кроется, вероятно, в том, что их миграция происходила отчасти селективно, а также в более высоком уровне жизни.
В строке 18 дан IQ в 86 баллов для индейцев Северной Америки по результатам 19 исследований. В строке 19 приведено такое же значение IQ в 86 баллов для индейцев Латинской Америки, полученное по результатам 9 исследованиям в 5 странах. Значение в 86 баллов принято нами как наилучшая оценка IQ американских индейцев. В строке 20 дан IQ 91 для
Расовые различия в интеллекте
183
арктических народов в Северной Америке по 15 исследованиям на Аляске и в Канаде.
В строке 21 приведён IQ европейцев в 99 баллов по данным 66 исследований в 25 странах (при определении медианы исключались балканские страны, население которых представляет собой европейско-южноазиатскую переходную форму). В строке 22 дан IQ 99 для европейцев вне Европы по данным 23 исследований в 12 странах. Медианное значение по данным всех исследований составляет 99 баллов и принято нами как наилучшая оценка IQ европейцев.
В строке 23 представлен IQ в 105 баллов для местного населения Восточной Азии по 60 исследованиям в 7 странах. В строке 24 дан IQ 101 для выходцев из Восточной Азии в Соединенных Штатах, определённый по данным 26 исследований. В строке 25 приведён IQ 102 для аборигенов Восточной Азии, проживающих в других странах, полученный по результатам 9 исследований в 5 государствах (Бельгия, Бразилия, Канада, Малайзия и Нидерланды). Причина того, что IQ аборигенов Восточной Азии в их собственных странах выше, чем в Соединенных Штатах и в других странах, заключается, видимо, в том, что мигрировали преимущественно лица с интеллектом ниже среднего. В случае Бельгии, Канады и Нидерландов это не может быть объяснено жизненным уровнем, так как он столь же высок, как и в Восточной Азии. Возможно, более низкий IQ выходцев из Восточной Азии в Бразилии и Малайзии (99 баллов) может частично определяться более низким уровнем жизни в этих странах. Так как миграция аборигенов Восточной Азии в страны вне Северо-Востока Азии могла происходить с некоторой долей селективности, величина IQ местного населения Восточной Азии в 105 баллов принята нами в качестве наиболее точной оценки.
Надёжность оценок IQ рас
Значения IQ многих выборок будут, очевидно, в некоторой степени неточны вследствие ошибок выборки и измерения. Точность результатов, называемая их надёжностью, оценивается по тому, насколько будут сходны результаты для двух выборок, сформированных в одной и той же стране. Корреляция между двумя полученными в тех же странах величинами IQ (там, где были доступны три или более оценки IQ — по двум крайним оценкам) составляет 0,94. Это показывает, что величины IQ высоко надежны.
Валидность оценок IQ — вопрос о степени, в которой они обеспечивают подлинные или валидные меры когнитивных способностей в выборках. Часто утверждается, что народы, которые получают низкие оценки
184
Ричард Линн
IQ, в действительности столь же интеллектуальны, как и европейцы, но тесты направлены против них. Проблема смещения оценок тестов подробно обсуждается Дженсеном (Jensen, 1980) в книге «Смещённость в тестах умственных способностей»(«Bias in Mental Tests») и, как было показано, это опасение не является обоснованным. Индивидумы и расы, которые успешно справляются с тестами интеллекта, также в большинстве случаев получают хорошее образование, имеют высокие заработки, успешно справляются с работой и имеют высокий социально-экономический статус (Jensen, 1980, 1998; Hermstein, Murray, 1994). Австралийский психолог Мюррей Дюк (Dyck, 1996, стр. 67) вынес следующее заключение о смещённости тестов:
«По имеющимся данным, когнитивные тесты одинаково надежны для разных рас, их сложность по заданиям для разных рас эквивалентна, корреляции субтестов сходны...и факторный анализ даёт близкие результаты. На вопрос, имеют ли стандартные тесты интеллекта культурное смещение, ответ будет: нет».
Ещё один вердикт о смещённости тестов был недавно вынесен Брауном, Рейнольдсом и Витакером (Brown, Reynolds, Whitaker, 1999, стр. 215): «В выполненных до настоящего времени исследованиях последовательно обнаруживается, что стандартизированные когнитивные тесты не смещены в терминах прогностической или конструктной валидности».
Валидность оценок IQ рас: Понятия числа
Под валидностью оценок IQ рас подразумевается степень, в которой они являются мерой реальных различий умственных способностей помимо умения решать задачи, представляемые в тестах интеллекта. Имеется несколько способов установить валидность расовых различий в интеллекте. Рассмотрим сначала расовые различия в развитии числовых понятий. Как показал Буттерворс (Butterworth, 1999), сложные числовые системы, имеющие обозначения чисел от единицы до десяти, десяток, тысяч, десяток тысяч и сотен тысяч, были разработаны аборигенами Южной Азии и Северной Африки и аборигенами Восточной Азии четыре-пять тысяч лет назад и несколько позже европейцами и американскими индейцами. Бушмены, африканцы, австралийские аборигены и аборигены Новой Гвинеи изобрели лишь числа обозначающие один, два, мало и много. На некоторых языках бушменов и австралийских аборигенов можно выразить числа до шести или семи, используя воспроизведение одного и двух. Так, семь выражается как два, два, два, один. Большие значения не могут быть выражены, так как становится слишком трудно запомнить число двоек и единиц. Построение сложных числовых систем требует умеренно высокого
Расовые различия в интеллекте
185
интеллекта и то, что в этом существуют расовые различия, позволяет предполагать, что расовые различия в интеллекте существовали уже несколько тысяч лет назад.
Валидность оценок IQ рас: Успехи в образовании
С первых лет двадцатого столетия валидность тестов интеллекта проверялась по степени корреляции их результатов с успехами в образовании. В многочисленных исследованиях было установлено, что корреляция величины IQ с достижениями в образовании лежит в пределах порядка от 0,6 до 0,7 (Jencks, 1972; Lynn, Hampson, Magee, 1984). Это свидетельствует о том, что тесты интеллекта служат валидной мерой общих когнитивных способностей, а не просто умения решать предъявляемые в тестах задачи.
Такую же методику мы использовали для оценки валидности расовых различий величин IQ. Задача состоит в определении того, связаны ли расовые различия в IQ с различиями успехов в образовании. Единицами анализа служат нации. Вычисляются IQ наций, затем нации распределяются по расам и оценивается, насколько IQ наций связан с национальными оценками по математике и естественным наукам. Величины IQ наций были получены из данных, представленных в предыдущих главах. Если для нации имелось две или более оценки IQ, брались медианные значения. Значение IQ для Латвии было взято из работы Линна и Ванханена (Lynn, Vanhanen, 2002). Для мультирасовых обществ величины IQ вычислялись с использованием взвешивая IQ этих рас согласно их пропорциями в популяции. Величина IQ для Чили была вычислена по данным нормативного исследования 5-16-летних испытуемых (п=4.213), тестировавшихся с помощью прогрессивных матриц Маринковичем и соавт. (Marinkovich et al., 2000).
Национальные оценки по математике и естественным наукам были взяты из Международных исследований достижений в математике и естественных науках. Серии этих исследований выполнялись в период между серединой 1960-х гг. и 1994 г., в них репрезентативным выборкам учащихся начальной и средней школ из ряда стран давали тесты по математике и естественным наукам. Некоторые результаты имеются для всех 52 стран, но не все страны участвовали во всех исследованиях, ввиду чего довольно многих данных недостаёт. В Таблице 13.2 представлены использованные нами пять наборов данных национальных оценок по математике и естественным наукам.
186
Ричард Линн
Таблица 13.2. Национальные IQ и достижения в математике и естественных науках

Расовые различия в интеллекте
187

В столбце 1 представлены нации, распределённые по расам. В столбце 2 даны значения IQ наций. В столбце 3 представлены данные первых двух Международных исследований достижений в математике и естественных науках, выполненных в период с середины 1960-х гг. по 1986 г. Данные были объединены Ханушеком и Кимко (Hanushek, Kimko, 2000) для того, чтобы получить единые оценки для каждой нации по шкале со средним значением 50 и стандартным отклонением 10. В столбцах 4, 5, 6 и 7 представлены оценки по математике и естественным наукам для 10- и 14летних по результатам Третьего международного исследования по математике и естественным наукам, проведённого в 1994 г. Данные были представлены Битоном и соавт. (Beaton, et al., 1996; 1996а).
188
Ричард Линн
Нации распределены по расам согласно расовому составу их населения. Для каждой расы в первой строке даны медианное значение IQ и медианы оценок достижений в образовании. Первыми идут шесть стран Восточной Азии (Китай, Гонконг, Япония, Сингапур, Южная Корея и Тайвань). Они получили самую высокую оценку IQ (105 баллов) и наиболее высокие значения по всем всех пяти показателям достижений в математике и естественных науках. Далее представлены 30 европейских стран. К ним отнесены и государства вне Европы, населенные преимущественно европейцами: Австралия, Новая Зеландия и Соединенные Штаты. У них второй по величине IQ (98 баллов) и второе место по оценкам успехов в математике и естественных науках. Третьими приведены три страны Южной Америки с мультирасовым составом населения (Бразилия, Чили и Колумбия). Медианное значение их IQ составляет 86 баллов. Их оценки по математике и естественным наукам значительно ниже, чем у европейских наций. Четвертую группу составляют девять стран Южной и ЮгоВосточной Азии. У них такой же IQ (86 баллов), как в странах Южной Америки. Их оценки по математике и естественным наукам немного превышают оценки южноамериканских стран, но количество данных очень ограничено. Оценки стран Южной и Юго-Восточной Азии по математике и естественным наукам значительно ниже, чем у европейских наций. Пятую группу составляют четыре страны района Африки южнее Сахары (данные для Свазиленда взяты из работы Бейкера и Джонса (Baker, Jones, 1993)). В этих странах самое низкое значение IQ (69 баллов) и самые низкие оценки по математике и естественным наукам.
В нижней строке таблицы приведены корреляции между IQ наций и оценками достижений в образовании. Корреляции лежат в пределах от 0,81 до 0,89 и все являются статистически значимыми на однопроцентном уровне. Эти корреляции ниже своих истинных значений вследствие ошибки измерения. Действительно, интеркорреляции между пятью оценками успехов в образовании составляют в среднем 0,78, что ниже среднего значения корреляции между IQ и пятью оценками достижений в образовании (0,83). Коррекция на ненадежность этих оценок (коррекция на аттенюа- цию) дает истинную корреляцию в 1,0 между национальными значениями IQ и достижениями наций в образовании. Это свидетельствует о валидности национальных IQ и показывает, что они измеряют важные когнитивные способности, а не просто умение выполнять тесты интеллекта.
Оценить величины расовых различий достижений в математике и естественных науках в сравнении с различиями в интеллекте можно,выразив различия рас в IQ и в оценках по математике и естественным наукам, приведённых в колонке 3, в единицах стандартного отклонения (ds).Эти сравнения приведены в Таблице 13.3. Величины IQ и оценки по математике и естественным наукам европейцев используются как стандарт, с кото
Расовые различия в интеллекте
189
рым сравниваются dдругих рас. Как можно заметить, во всех случаях различия рас по успехам в образовании больше различий в интеллекте. Так, у аборигенов Восточной Азии преимущество перед европейцами в IQ составляет 0,33d(5 баллов IQ), а преимущество успехов в образовании выше и составляет 0,44d.Это может быть следствием лучшего обучения в их школах, о чём я говорил применительно к Японии (Lynn, 1988). К примеру, учебный год в Японии длится 240 дней, а в Великобритании и Соединенных Штатах 180 дней, во Франции 155 дней. В Восточной Азии в школах обычно больше порядка и меньше проблем с дисциплиной, чем в европейских странах, ввиду чего восточно-азиатские школьники имеют возможность усвоить больше. Возможно, что у восточно-азиатских школьников более сильная, чем у европейцев, трудовая этика и это способствует их лучшим успехам в образовании.
В трёх других расовых и этнических группах дело с достижениями в образовании обстоит хуже, чем с интеллектом. Так, оценка интеллекта латиноамериканцев ниже европейской на 0,66d(10 баллов IQ), но по успехам в образовании — на 2,27d.Аборигены Южной Азии уступают европейцам в интеллекте 0,93d(14 баллов IQ), а по успехам в образовании 1,30d. У африканцев интеллект ниже европейского на 2,00J (30 баллов IQ), а достижения в образовании ниже на 2,44d.Очевидно, имеются другие факторы, помимо более низкого интеллекта, которые снижают достижения в образовании в этих экономически развивающихся странах. Возможно, это меньшая эффективность школьного обучения или более слабая трудовая этика. Причины расовых различий достижений в образовании были очень плохо исследованы специалистами в области педагогики, которые даже не признают, что различия в интеллекте — главный фактор, хотя, конечно, задействованы и другие факторы.
Таблица 13.3. Сравнение национальных IQ и достижений в образовании в области математики и естественных наук (ds)

190
Ричард Линн
Валидность оценок IQ рас: Доход на душу населения и экономический рост
Другой метод установления валидности тестов интеллекта состоит в исследовании связи показателей тестов с доходом на душу населения и экономическим ростом. В нескольких исследованиях, где единицами анализа выступали индивиды, было показано, что оценки IQ, полученные в детстве, коррелируют с заработком во взрослом возрасте с коэффициентом порядка от 0,35 (Jencks, 1972) до 0,37 (Murray, 1998). Поскольку интеллект измерялся в детстве, обычно считается, что IQ — детерминанта дохода во взрослом возрасте. Эта точка зрения подтверждается результатами многочисленных исследований, показывающими, что интеллект положительно связан с успешностью профессиональной деятельности, корреляция лежит в пределах от 0,4 до 0,6 (Hunter, Hunter, 1984).
Валидность расовых различий величин IQ была оценена посредством распространения этой закономерности на расы. В нашей работе (Lynn, Vanhanen, 2002) мы оценили IQ всех 185 наций мира численностью более 50 тыс. чел. Оценки IQ наций определяются расовым составом популяций, как было показано в предыдущем параграфе. Нами было обнаружено, что национальные IQ коррелировали с валовым внутренним продуктом (ВВП) на душу населения в 1998 г. (г=0,62) и с валовым национальным продуктом (ВНП) на душу населения в 1998 г (г=0,63). Это показывает, что величина IQ нации объясняет 38 и, соответственно, 40 процентов различий ВВП и ВНП на душу населения. Следовательно, IQ нации вносит значительный вклад в её национальный доход (по этим двум показателям). Корреляции не слишком велики ввиду того, что национальный доход на душу населения в значительной степени определяется другими факторами, из которых наиболее важные — обладание природными ресурсами и наличие рыночной экономики. Так, например, реальный ВВП на душу населения в 1998 г. в Катаре (IQ=78) составлял 20.987$ — примерно столько же, сколько в Великобритании (20.336$) и был намного выше, чем в странах Южной Азии приблизительно с тем же IQ, но не имеющих природных ресурсов, таких как Индия (IQ=82, ВВП=2.077$) и Иордания (IQ=84, ВВП=3.347$). Источником высокого ВВП в Катаре является владение нефтью, которой не обладают Индия или Иордания. Вторым главным фактором, определяющим национальный доход, является наличие рыночной экономики. Так, страны Восточной Европы имеют примерно такие же показатели IQ, что и в Западной Европе, но доходы на душу населения в них намного ниже (например, 6.460$ в России и 7.619$ в Польше) в сравнении
Расовые различия в интеллекте
191
с 20.336$ в Великобритании и 20.846$ во Франции. Сходным образом, доход на душу населения в Китае (3.105$ в 1998 г.) намного ниже, чем в Гонконге (20.763$) и Сингапуре (24.210$), хотя все они населены преимущественно или исключительно китайцами и имеют близкие величины IQ (Китай — 103, Гонконг — 107, Сингапур — 103). Причина в том, что экономическое развитие в Китае было заторможено коммунистической системой экономики, в то время как в Гонконге и Сингапуре рыночная экономика ему способствовала. Во всех странах, вместе взятых, коэффициент корреляции степени развития у них рыночной экономики, которую оценивали по индексу экономической свободы, с реальным ВВП на душу населения в 1998 г. составлял 0,71, что немного выше, чем корреляции IQ с реальным ВВП на душу населения в 1998 г. (0,62). Это показывает, что как экономическая свобода, так и IQ вносят существенный вклад в национальное благосостояние. Коэффициент корреляции степени экономической свободы с реальным ВНП на душу населения в 1998 г. составил 0,64, а коэффициент корреляции национального IQ и ВНП на душу населения был равен 0,63.
Наблюдается также значительная связь IQ нации со скоростью её экономического роста в течение двадцатого столетия. Так, корреляция национального IQ и роста ВВП с 1950 г. по 1990 г. составила 0,51. Это говорит о том, что страны с более высокими показателями IQ становятся богаче и разрыв между богатыми странами с высоким IQ и бедными странами с низким IQ увеличивается. Это ожидаемо, так как экономический рост стимулируется новыми высокотехнологичными товарами, такими как компьютеры, мобильные телефоны, фармацевтические препараты, самолёты, оружие, автомобили, и т. д., которые могут быть созданы только людьми с высоким IQ. Нации с низким IQ зарабатывают на жизнь преимущественно экспортом сельскохозяйственных продуктов, сырья и полезных ископаемых, которые менее востребованы и цены на которые ниже, ввиду чего темпы их экономического роста и доходы на душу населения остаются низкими.
Существенные корреляции между IQ наций, их доходами на душу населения и темпами экономического роста обеспечивают дальнейшую валидацию показателей национального интеллекта, так как они могут быть предсказаны, а предсказания верифицированы. Однако, требуется пояснение. Положительная связь между индивидуальными IQ и доходами внутри государств — односторонне направленное причинное отношение. Величина IQ, измеренная в детстве и подростковом возрасте, является детерминантой заработка взрослых, в то время как доходы взрослых — не детерминанта их интеллекта. Положительная связь между оценкой IQ нации и национальным доходом на душу населения не является столь однонаправленной. Это — двустороннее причинное взаимодействие, включающее положительную обратную связь. Национальные популяции с
192
Ричард Линн
высоким IQ в состоянии произвести сложные товары и услуги (компьютеры, мобильные телефоны, автомобили, самолёты, фармацевтические препараты и т. д.) высоко ценящиеся на международных рынках. Это обеспечивает высокий среднедушевой национальный доход, который позволяет этим популяциям создавать себе благоприятные средовые условия с высококачественным питанием, здравоохранением и образованием, что развивает интеллект их детей. Когда эти дети становятся взрослыми, они могут использовать свой высокий интеллект для того, чтобы производить более сложные товары и услуги, высоко ценящиеся на международных рынках. И так далее по этому благодатному кругу.
ГЛШ 14
оедшк и гжпшсмдомишш
ШКОШХ РАЗЛИЧИЙ О Н1ПШВД
Питание
Голландское исследование голодания в период Второй Мировой войны
Нейрофизиологический эффект недоедания
Образование
Генетические детерминанты расовых различий в интеллекте
Генотипно-средовая ковариация
Теперь мы рассмотрим вопрос о средовых и генетических детерминантах расовых различий в интеллекте. Возможны три точки зрения на эту проблему. Первая: различия между всеми десятью расами могут быть полностью определены средой обитания. Вторая: различия могут быть полностью определены генетически. И третья: различия могут определяться как генетическими, так и средовыми факторами. Безусловно, наиболее вероятно третье из этих положений — что и генетические и средовые факторы ответственны за различия в интеллекте.
Вопрос о том, вносит ли наследственность вклад в различия интеллекта, обсуждался на протяжении более столетия. Значительная часть, но отнюдь не все эти дебаты, касались различий между афроамериканцами и европейцами в Соединенных Штатах. Среди тех, кто высказывался в пользу существенного влияния наследственности — Гобино (Gobineau, 1853), Галь- тон (Galton, 1869), Гарретт (Garrett, 1945,1961), МакГурк (McGurk, 1953а, 1953Ъ), Шу (Shuey, 1966), Шокли (Shockley, 1968), Дженсен (Jensen, 1969, 1980, 1998), Вернон (Vernon, 1969, 1979), Айзенк (Eysenck, 1971), Бейкер (Baker, 1974), Лоэлин, Линдзи и Спулер (Loehlin, Lindzey, Spuhler, 1976), Раштон (Rushton, 1988, 2000), Раштон и Дженсен (Rushton, Jensen, 2005), Линн (Lynn, 1991, 1991b, 1997), Вальдман, Вайнберг и Скарр (Waldman, Weinberg, Scarr, 1994, стр. 38), Скарр (Scarr, 1995), Левин (Levin, 1997) и Готтфредсон (Gottfredson, 2005). Среди утверждающих об отсутствии существенной генетической детерминации расовых различий — Флинн (Flynn, 1980), Броди (Brody, 1992, 2003), Найссер (Neisser, 1996), Нисбетт
194
Ричард Линн
(Nisbett, 1998), Макинтош (Mackintosh, 1998), Дженкс и Филипс (Jencks, Phillips, 1998), а также Фиш (Fish, 2002). Целые тома были посвящены этому вопросу. Дженсен (Jensen, 1998) в своей монографии «Фактор g»посвящает этому главу из 113 страниц, которая сама по себе является почти что книгой, но даже в ней рассматриваются почти исключительно различия между чернокожими и белыми в Соединенных Штатах. Целью настоящей книги не является рассмотрение всех данных и доводов по этой проблеме, а скорее расширение дискуссии от локальной проблемы вклада средовых и генетических факторов в различие между чернокожими и белыми в Соединенных Штатах к намного более широкой глобальной проблеме детерминации различий между десятью расами, показатели IQ которых были приведены в Таблице 13.1.
Питание
Нет сомнения в том, что низкие величины IQ населения бедных стран третьего мира до некоторой степени определены средовыми факторами, наиболее важный из которых — плохое питание. Даже в богатых эконо- мически-развитых странах значительная доля населения питается плохо, что отрицательно воздействует на интеллект. Имеется много данных, подтверждающих это неблагоприятное действие, они были детально рассмотрены в моих работах (Lynn, 1990а, 1993, 1998b). Например, иногда случается, что близнецы при рождении имеют разную массу тела и размер мозга вследствие того, что близнец с большей массой получал в матке больше питательных веществ, чем близнец с меньшей массой. Недостаточное питание, получаемое близнецами с меньшей массой, оказывает постоянное отрицательное воздействие на их интеллект, что выражается в более низком IQ (в среднем примерно на 5 баллов) в подростковом возрасте и во взрослом состоянии. В моей работе (Lynn, 1990а) дан обзор семи исследований, в которых это было показано. В нескольких исследованиях в эко- номически-развитых обнаружено, что младенцы, которых кормили грудью, впоследствии получают более высокие оценки IQ чем те, которые питались смесями на основе коровьего молока (Lucas et al., 1992; Lucas, Morley, Cole, 1998). Объяснение этого состоит в том, что грудное молоко содержит питательные вещества, не присутствующие в молочных смесях, и железо из грудного молока у младенцев всасывается лучше, чем из коровьего молока. Было также показано, что введение пищевых добавок в рацион подростков, имевших пищевые дефициты, повышает их интеллект. Например, исследование подростков в социально неблагополучном городе Великобритании показало, что 17 процентов из них испытывали дефи
Расовые различия в интеллекте
195
цит железа. Ежедневное введение в их рацион железосодержащих пищевых добавок на протяжении трех месяцев привело к увеличению IQ на 5,8 баллов (Lynn, Harland, 1998). Другие исследования, в которых было продемонстрировано положительное влияние пищевых добавок на интеллект детей в экономически-развитых странах, описаны Бентоном и Робертсом (Benton, Roberts, 1988), Бентоном и Куком (Benton, Cook, 1991), а также Айзенком и Шойнталером (Eysenck, Schoenthaler 1997). Долговременное увеличение интеллекта, наблюдаемое в экономически-развитых странах на протяжении большей части двадцатого столетия, в значительной степени обязано улучшению питания, вызвавшего увеличение роста на сходную величину - примерно на половину стандартного отклонения за более чем пятьдесят лет. Обзор данных, свидетельствующих об этом, содержатся в моей работе (Lynn, 1990а).
Во многих бедных экономически развивающихся странах недостаточное питание широко распространено и имеется масса свидетельств тому, что это отрицательно воздействует на интеллект населения. Основные изученные виды неполноценного питания это: белково-калорийная недостаточность, дефицит железа и дефицит йода. Белково-калорийная недостаточность задерживает рост, а в крайних случаях вызывает квашиоркор10 и маразм. Дефицит железа вызывает анемию, общую слабость и снижает интеллект. Дефицит йода вызывает зоб, а у беременных женщин нарушает развитие мозга плода, приводящее к снижению интеллекта и кретинизму. Бличродт и Борн (Bleichrodt, Bom, 1994) выполнили мета-анализ данных 18 исследований, в которых сравнивался интеллект населения из йодо-дефицитных и из благополучных по этому показателю регионов, изучалось также влияние введения йода населению йодо-дефицитных популяций. Был сделан вывод о том, что тяжёлый дефицит йода вызывает снижение интеллекта на 13,5 баллов.
Недоедание подавляет физический рост, включая рост мозга, что является причиной снижения интеллекта. Недоедание оценивается по «задержке роста», «истощению», и «снижению массы тела».
Задержка роста—это низкий рост. Умеренной и серьезной задержкой роста считается рост, сниженный больше, чем на два стандартных отклонения от медианного значения роста в популяции того же возраста, получавшей полноценное питание. Задержка роста вызвана хронической недостаточностью белка, необходимого для роста костей.
10 Квашиоркор (иначе — изолированная белковая недостаточность; на языке коренного населения Ганы означает «золотой мальчик» или «красный мальчик») — тяжелое расстройство питания у детей. Наблюдается у детей до 5 лет, чаще возникает после прекращения грудного вскармливания. При квашиоркоре наблюдаются задержка роста, отёки, гипоальбуминемия, жировая дистрофия печени, нарушение пигментации кожи, психические расстройства (прим. перев.)
196
Ричард Линн
Умеренным и серьёзным истощением считается масса тела, сниженная более, чем на два стандартных отклонения от медианы массы тела лиц того же роста в нормально питающейся популяции.
Умеренно и серьёзно сниженной считается масса тела, уменьшенная более, чем на два стандартных отклонения от медианы массы тела лиц того же возраста в полноценно питающейся популяции.
В Таблице 14.1 представлены данные о распространенности умеренного и серьёзного недоедания в экономически развивающихся странах разных регионов в начале 1990-х гг. (по оценке ЮНИСЕФ — UNICEF, 1996) и анемии среди беременных женщин в период 1960-1982 гг. (по данным Всемирной Организации Здравоохранения — De Maeyer, Adiels- Tegman, 1985). Отчёты по отдельным странам подтверждают эти данные. К примеру, исследование, проведённое в Индии в 1980-х гг., выявило анемию приблизительно у 60 процентов детей до 3 лет и у 44 процентов 3-5 летних (Seshadri, Gopaldas, 1989). Неадекватное питание во многих экономически развивающихся странах отягощено болезнями, особенно диареей и корью, которые нарушают усвоение питательных веществ.
Таблица 14.1. Распространенность недоедания в экономически развивающихся странах (проценты)

Отрицательное воздействие недоедания на интеллект многих народов в развивающихся странах продемонстрировано многими исследованиями, в которых сравнивали IQ хорошо и плохо питавшихся детей. Симеон и Грантам-МакГрегор (Simeon, Grantham-McGregor, 1990) проанализировали результаты пятнадцати таких исследований и пришли к выводу, что в десяти из них плохо питавшиеся дети имели более низкий IQ, чем те, которых кормили хорошо. Отрицательное воздействие неадекватного питания на интеллект было показано также в ряде исследований, в которых недоедавшим детям давали пищевые добавки, результатом чего было уве
Расовые различия в интеллекте
197
личение интеллекта. Обзор семи таких исследований, выполненных в экономически развивающихся странах, был сделан Симеоном и Грантам- МакГрегорм (Simeon, Grantham-McGregor, 1990).
Хотя неадекватное питание, несомненно, ослабляет интеллект значительной доли населения экономически развивающихся стран, оно не объясняет полностью расовые различия. Приведённые в Таблице 14.1 значения говорят о том, что в экономически развивающихся странах Африки южнее Сахары, Ближнего Востока и Северной Африки, Восточной Азии, Тихоокеанских островов, Латинской Америки и на островов Карибского бассейна от недоедания страдает менее половины детей. Только в Южной Азии голодает больше половины детей, у 64 процентов которых снижена масса тела, а у 62 процентов имеется задержка роста. Хотя в нескольких исследованиях было показано, что IQ недоедающих детей в экономически развивающихся странах снижен в сравнении с хорошо питающимися, всё же IQ хорошо питающихся детей существенно ниже, чем у европейцев и аборигенов Восточной Азии в экономически развитых странах. Например, Геллер и её коллеги сообщили (Galler, Ramsey, Ford, 1986), что у детей на Барбадосе, недоедавших на первом году жизни, IQ в возрасте 9-15 лет составлял 68 баллов, в то время как в группе хорошо питавшихся детей IQ был равен 83 баллам. Различие в 15 баллов указывает на то, что недоедание должно уменьшать IQ на 15 баллов. Однако, лишь 16,5 процентов детей на Барбадосе недоедают и IQ в 83 балла у хорошо питающихся африканских детей значительно ниже IQ европейцев (99 баллов) и аборигенов Восточной Азии (105 баллов). В грубых цифрах, недоедание африканцев в Африке южнее Сахары и в бассейне Карибского моря объясняет, вероятно, приблизительно половину их низких значений IQ, оставляя другую половину на долю генетических факторов.
Сторонники средовой теории иногда утверждают, что плохое питание привносит свой вклад в низкий IQ афроамериканцев в Соединенных Штатах. Так, Ричардсон и Спирс (Richardson, Spears, 1972, стр. 82) пишут: «Мы имеем неоспоримые свидетельства того, что меньшинства, такие как чернокожие, всегда имеют тенденцию питаться хуже, чем большинство». Они не представляют никаких свидетельств в пользу этого утверждения и сомнительно, что оно верно. Плохое питание снижает рост, таким образом, низкоросл ость — показатель плохого питания. Но уже в 1918 г. рост американских призывников составляли 170,96 см у белых и несколько больше у чернокожих — 171,99 см (Nelson, 1933). Последующее исследование, опубликованное позже в двадцатом столетии, подтвердило, что различий в росте между американскими чернокожими и белыми не наблюдается как в возрасте 4 и 7 лет, так и у взрослых (Broman et al., 1985). Изучение рациона американских чернокожих и белых также не выявило каких-либо различий. Например, исследование 1987-88 гг. выявило, что в репрезентативной
198
Ричард Линн
выборке 9-10-летних девочек (п=2.379) 20 процентов чернокожих и 25 процентов белых получали витамин С в дозе ниже рекомендованной суточной нормы, равной 45мг (Simon et al., 1993). В Первом Исследовании Здоровья и Питания девочек возраста до 15 лет не было выявлено различий между чернокожими и белыми в отношении низкого потребления витамина С (National Center for Health Statistics, 1979). Другие пищевые дефициты вероятно будут сопутствовать дефициту витамина С, так что эти результаты позволяют полагать, что американские чернокожие испытывают дефицит питания не в большей степени, чем белые.
Что касается аборигенов Восточной Азии, то в исследовании корейских младенцев, усыновлённых американскими родителями в возрасте до 2 лет и обследованных с помощью тестов интеллекта в возрасте от 6 до 14 лет (Winick, Meyer, Harris, 1975) было обнаружено, что IQ серьёзно голодавших в младенчестве составлял 102 балла, у голодавших в младенчестве в умеренной степени — 106 баллов, в то время как у хорошо питавшихся он был равен 112 баллам. Результаты позволяют предположить, что серьезное недоедание в младенчестве ослабляет интеллект на 10 баллов. Однако, даже у аборигенов Восточной Азии, серьёзно недоедавших в младенчестве, IQ составлял 102 балла, что немного выше, чем у хорошо питающихся европейцев. Это говорит о том, что за более высокий IQ аборигенов Восточной Азии ответственны генетические факторы.
Голландское исследование голодания в период Второй Мировой войны
Основным исследованием, дающим основание полагать, что недоедание в пренатальный и ранний постнатальный периоды не оказывает отрицательного воздействия на интеллект детей, является Голландское исследование голодания в период Второй Мировой войны (Stein et al, 1972). В этом исследовании изучалось влияние голода в одном из районов Нидерландов зимой и весной 1944-45 гг., где население, включая беременных женщин, серьезно недоедало в течение шести месяцев. Рацион был снижен приблизительно до 700 калорий в день, что составляет примерно четверть — одна пятая рациона, обычно потребляемого в экономически развитых странах. В этот период масса тела новорожденных младенцев была снижена приблизительно на 300 граммов, но к возрасту 19 лет они имели такие же значения IQ, что и у проживавших в других районах Нидерландов и не подвергшихся воздействию голода в пренатальный период. Однако авторы предупреждают, что: «Результаты не следует распространять ни на последствия хронического недоедания с другим набором пищевых дефи
Расовые различия в интеллекте
199
цитов, какое часто имеет место в развивающихся странах, ни на неблагоприятные последствия недоедания в постнатальный период жизни» (стр. 712). Этот вопрос был детально рассмотрен Мартореллом (Martorell, 1997), утверждающим, что 6-месячное недоедание не оказывает отрицательного воздействия, если мать до того питалась хорошо, и ребёнок получал нормальное питание в остальной срок беременности и в младенчестве. Он полагает, что матери, вероятно, имели запасы микронутриентов, которые использовались в период голода. В экономически развивающихся странах многие люди хронически недоедают, и компенсация такого рода невозможна. Имеются сомнения относительно правильности этого объяснения в свете факта, что у младенцев, родившихся в охваченном голодом районе, масса тела при рождении была существенно ниже (приблизительно на 300 граммов) чем в районах, где не было голода, а также весьма весомых данных о том, что низкая масса тела при рождении часто сопровождается пониженным интеллектом. Если бы матери были в состоянии мобилизовать резервы питательных веществ, масса тела их младенцев при рождении была бы нормальной. Её значительное снижение указывает на то, что голодание действительно оказывало отрицательное воздействие. Наиболее вероятное объяснение полученных результатов состоит в том, что в районах голода доля младенцев, родившихся в семьях, не занимавшихся ручным трудом, была выше, чем в семьях, занимавшихся ручным трудом. Возможно, что не занимавшиеся ручным трудом семьи имели больше возможностей получать пищу из областей, не охваченных голодом, и это улучшало их питание и увеличило рождаемость по сравнению с семьями, занимающихся ручным трудом. Имеется тесная связь между интеллектом и социально-экономическим статусом, и, таким образом, увеличение доли младенцев, родившихся в семьях, не занимавшихся ручным трудом, компенсировало отрицательное воздействие недоедания в этой когорте.
Нейрофизиологический эффект недоедания
Нейрофизиологическим эффектом недоедания является нарушение роста мозга, вследствие чего он функционирует менее эффективно. Недоедание в пренатальный и ранний постнатальный периоды оказывает наиболее серьёзное отрицательное влияние на интеллект, так как приблизительно 70 процентов роста мозга приходится на внутриутробный период, а остающиеся 30 процентов, включая рост дендритов и синаптических ветвлений, завершаются к возрасту 18-24 месяцев (Dobbing, Smart, 1974). Недоедание оказывает на плод и новорожденного младенца разнообразные отрицательные воздействия, впоследствии ослабляющие интеллект. Из них лучше всех установлены следующие:
200
Ричард Линн
Недоедание нарушает рост мозга и уменьшает количество мозговых клеток, а размер мозга связан с интеллектом с корреляцией 0,40 (Vernon et al., 2000);
Дефицит железа снижает количество рецепторов дофамина, это ослабляет дофаминэргическую нейропередачу, что в свою очередь ухудшает обучаемость и функционирование мозга во взрослом состоянии;
Жирные кислоты необходимы для роста и эффективного функционирования мозга; приблизительно половина этих кислот усваивается во внутриутробном периоде, а другая половина в первые 12 месяцев жизни из грудного молока; этих жирных кислот нет в коровьем молоке и в большинстве молочных смесей для младенцев, что является одной из причин того, что младенцы, которых кормили грудью, в дальнейшем имеют более высокие значения IQ (Grantham-McGregor, Walker, Powell, 1994).
Образование
Вторым средовым фактором, который иногда считают ответственным за низкие величины IQ народов экономически развивающихся стран, является недостаток образования. К примеру, Фиш (Fish, 2002, стр. 14) пишет: «Недостаток систематического образования африканцев относительно европейских групп сравнения даёт очевидное объяснение их более низких результатов выполнения тестов». Бишевель (Biesheuvel, 1949) высказывает ту же точку зрения и цитирует выполненную в Южной Африке работу, в которой было показано, что у никогда не посещавших школу африканцев IQ был приблизительно на 10 баллов ниже, чем у посещавших школу, и это, утверждает он, говорит о том, что недостаток школьного обучения ослабляет интеллект. Обосновать это положение трудно, так как вероятно, что у детей, которых посылают в школу, родители имеют более высокие значения IQ и более высокий социально-экономический статус, чем у не посещающих школу детей, и более высокий IQ эти дети могли скорее приобрести от родителей, а не вследствие обучения в школе. В исследованиях влияния обучения на интеллект в экономически развитых странах были получены неоднозначные результаты. Цеси (Ceci, 1991) и Макинтош (Mackintosh, 1998) провели обзор нескольких исследований, выявивших увеличение интеллекта вследствие школьного обучения, но это увеличение было кратковременным и исчезало через несколько лет. Не было продемонстрировано, что это увеличение устойчиво. Имеются некоторые данные, говорящие об обратном. В Великобритании дети начинают учиться в школе в возрасте 5 лет, а почти во всех других странах в шесть или
Расовые различия в интеллекте
201
семь, но британские дети, как кажется, не получают преимущества за счёт дополнительного года обучения. В Соединенных Штатах в Отчёте Колме- на, имевшего полномочия исследовать влияние обучения, делается заключение, что «школа оказывает незначительное влияние в отношении достижений ребёнка, которые независимы от его подготовленности и общего социального контекста» (Coleman et al., 1966, стр. 325). Школы в Соединенных Штатах были объединены с 1960-х гг., когда чернокожие, белые, выходцы из Азии и представители других рас стали посещать одни и те же школы, куда во многих областях их начали возить автобусами, но IQ афроамериканцев и американских индейцев возрос незначительно или вообще не увеличился. В Великобритании и континентальной Европе представители всех рас посещают одни и те же школы, но расовые различия в интеллекте до сих пор существуют.
Большинство исследований интеллекта рас, рассмотренных в Главах 3-12, было выполнено среди школьников, во многих случаях дети посещали те же школы, что и европейцы. В Южной Африке в выборке Оуэна (Owen, 1992) 16-летние испытуемые имели 8-10-летнее образование и, однако же, получили типичные для этого региона средние оценки IQ в 63 балла. В двенадцати исследованиях среди африканцев — студентов университетов в Южной Африке, обучавшихся в школе от десяти до двенадцати лет, было обнаружено, что их IQ приблизительно на 20 баллов ниже, чем у белых студентов (см. Главу 4, Таблицу 4.2). Сходным образом, в трёх исследованиях интеллекта студентов университетов в Индии было обнаружено, что их IQ составлял 88,90 и 95 баллов — ощутимо ниже среднего европейского значения.
Более того, в нескольких исследованиях было показано, что расовые различия в интеллекте в полной мере представлены у дошкольников. Так, IQ 3-летних африканцев в Доминиканской республике составляет 67 баллов (Wein, Stevenson, 1972), а у 4-летних на острове Санта Лючия он равен 62 баллам (Murray, 1983). В Соединенных Штатах IQ 3-летних африканцев оценивается в 86 (Montie, Pagan, 1988) и 85 баллов (Peoples et al., 1995), а у 4-летних он составляет 87 баллов (Broman et al., 1975) — как раз столько же, сколько у афроамериканских подростков и взрослых. Эти исследования дошкольников позволяют говорить о том, что недостаток образования не является существенным фактором, определяющим расовые различия в интеллекте.
202
Ричард Линн
Генетические детерминанты расовых различий в интеллекте
Наряду с тем, что средовые факторы, несомненно, вносят вклад в расовые различия в интеллекте, имеется ряд соображений в пользу того, что генетические факторы также играют роль. В настоящем параграфе мы рассмотрим десять таких соображений.
Во-первых, принцип эволюционной биологии гласит, что когда субпопуляции биологического вида оказываются географически изолированными и попадают в разную среду обитания, они генетически дифференцируются, и, в конечном итоге, дивергируют настолько, что становятся разными видами. Например, у белок в Северной Америке образовался серый мех, а в Европе — рыжий. Изначальный предковый вид кошек эволюционировал во львов, леопардов и гепардов в Африке, тигров в Азии и в ягуаров и пум в Америке. Общий принцип был сформулирован Доукин- сом (Dawkins, 1988, стр. 238-239), писавшем, что когда две популяции становятся изолированными от друг друга:
«они становятся столь непохожими друг на друга, что через некоторое время натуралисты отнесли бы их к разным расам; через более длительное время они дивергируют настолько, что мы должны будем классифицировать их как различные виды..., теория видообразования вследствие изначальной географической изоляции долго была краеугольным камнем господствующего направления, ортодоксального неодарвинизма».
Процессы, посредством которых происходит это генетическое расхождение, были описаны в Главе 2. Именно в соответствии с этим принципом расы стали генетически дифференцированными по всем признакам, по которым имеется генетическая вариабельность, в том числе по форме тела; цвету кожи, волос и глаз; распространенности генетических болезней; группам крови. Невозможно представить, что интеллект будет единственным исключением в ряду этих различий. Некоторые расовые различия в интеллекте также должны были развиться на основании общего биологического принципа.
Во-вторых, результаты исследований, представленные в Таблице 13.1, свидетельствуют о постоянстве величин IQ рас в широком диапазоне географических регионов, что может быть объяснено только некоторой генетической детерминацией. Например, в 57 исследованиях общепопуляционных выборок африканцев в 17 африканских странах все значения IQ лежат в диапазоне от 59 до 88 баллов (Таблица 4.1), а в 14 странах Карибского бассейна и Латинской Америки все значения IQ находятся в диапазоне от 60 до 80 баллов (Таблица 4.3). Сходным образом, в 58 исследова
Расовые различия в интеллекте
203
ниях аборигенов Восточной Азии в 6 странах все значения IQ лежат в диапазоне от 100 до 120 баллов (Таблица 10.1). Лишь генетический фактор может объяснить устойчивость этих расовых различий в столь различающейся окружающей среде. Странно, что сторонники средовой теории расовых различий в интеллекте, такие как Найссер (Neisser, 1996), Макинтош (Macintosh, 1998), Дженкс и Филлипс (Jencks, Phillips, 1998), Нисбетт (Nisbett, 1998), Флинн (Flynn, 1980), Фиш (Fish, 2002) и Броди (Brody, 2003) никак не упоминают об устойчивости расовых различий в самых разных условиях среды обитания и в разных странах.
В-третьих, при жизни в одинаковых условиях представители разных рас устойчиво различаются по величине IQ. Так, африканцы и в Соединенных Штатах, и в Великобритании, и в Нидерландах и в Бразилии имеют более низкие значения IQ, чем белые. Это же верно для выходцев из Южной Азии и Северной Африки в Великобритании, континентальной Европе, в Африке, на о-вах Фиджи, в Малайзии и на Маврикии; для американских индейцев, проживающих вместе с европейцами в Соединенных Штатах, Канаде и Мексике; для арктических народов, живущих вместе с европейцами в Канаде; для австралийских аборигенов, живущих вместе с европейцами в Австралии и для аборигенов островов Тихого океана, проживающих вместе с европейцами в Новой Зеландии и на Гавайях. Все эти различия согласуются друг с другом и ложатся на чашу весов генетической теории.
В-четвертых, младенцы других рас, усыновлённые европейцами в Европе и в Соединенных Штатах, сохраняют IQ, характерный для своей расы. Это было показано для африканцев в Соединенных Штатах, где чернокожие младенцы, усыновлённые белыми родителями среднего класса, имели такой же IQ, как и чернокожие, воспитываемые в их собственных сообществах (Lynn, 1994с); для австралийских аборигенов в Австралии и для аборигенов Восточной Азии в Соединенных Штатах и Европе (у корейских младенцев, усыновлённых европейцами, величина IQ находится в диапазоне от 102 до 110, см. Таблицу 10.4) — в Главах 4, 8 и 10, соответственно.
В-пятых, лица со смешанной расовой родословной имеют значение IQ, промежуточное между значениями родительских рас. Так, в исследовании детей, усыновлённых семьями белых среднего класса (Weinberg, Scarr, Waldman, 1992), было показано, что к возрасту 17 лет IQ чернокожих составлял 89 баллов; у тех, у кого один из родителей был чернокожим, а другой белымй — 98 баллов; у белых - 106 баллов (Lynn, 1994с). Когда долю европейских предков у американских чернокожих оценивали по цвету кожи, темнокожие афроамериканцы имели IQ в 85 баллов, а у светлокожих он составлял 92 балла (Lynn, 2002а); связь между оттенком кожи и интеллектом была статистически значимой. Сходным образом, у австралийских аборигенов со смешанной расовой родословной значения IQ находятся в промежутке между значениями чистокровных аборигенов и
204
Ричард Линн
европейцев (Глава 8, Параграф 2), а величина IQ расово-смешанных американских индейцев является промежуточной между значениями чистокровных индейцев и европейцев (Глава 12, Таблица 12.4).
В-шестых, величины IQ рас объясняют степень, в какой они совершили неолитический переход от охотников-собирателей к оседлому сельскому хозяйству. Этот переход был совершён полностью более интеллектуальными расами: европейцами, аборигенами Южной Азии и Северной Африки, аборигенами Восточной Азии, аборигенами Юго-Восточной Азии и американскими индейцами; до некоторой степени аборигенами островов Тихого океана, которых ставила в невыгодные условия жизнь маленькими и рассеянными популяциями на небольших островах; минимально африканцами; он совсем не имел места у бушменов и австралийских аборигенов (чьи показатели интеллекта равны 54 и 62 баллам), которые не достигли практически никакого прогресса в переходе от охотников-собирателей к осёдлым сельскохозяйственным обществам. Единственная аномалия — арктические народы с их IQ 91, оставшиеся в значительной степени охотниками-собира- телями, но это произошло по причине их жизни очень маленькими рассеянными популяциями и сурового климата Заполярья.
В-седьмых, значения IQ рас согласуются с их достижениями в создании ранних городских цивилизаций с письменностью, системой счисления и кодифицированными законами, это было показано Бейкером (Baker, 1974), документально подтвердившим, что только аборигены Восточной Азии, европейцы, аборигены Южной Азии и Северной Африки и аборигены Юго-Восточной Азии создали ранние цивилизации. Менее интеллектуальные американские индейцы создавали «полуцивилизации», а остальные расы не смогли создать чего-либо, что можно было бы назвать цивилизациями. Что касается аборигенов островов Тихого океана и арктических народов с их IQ 90 и 91, которые не создали что-либо напоминающее цивилизацию, то это можно объяснить в случае аборигенов островов Тихого океана их расселением очень маленькими и рассеянными популяциями на изолированных островах, а в случае арктических народов — суровостью климата, который лишил их возможности поддерживать городские цивилизации. Эти расовые различия в развитии ранних цивилизаций в период приблизительно с 4.000 по 500 гг. до нашей эры, проанализированные Бейкером (Baker, 1974), сохраняются с 1 века нашей эры до настоящего времени. Фактически все достижения в науке, математике, технике и искусствах за последние две тысячи лет были сделаны аборигенами Восточной Азии и европейцами с небольшим вкладом аборигенов Южной Азии и Северной Африки. Это было детально документировано Мюрреем (Murray, 2003), хотя он анализирует эти достижения по географическим регионам и воздерживается от указания на то, что эти достижения были сделаны почти исключительно народами Восточной Азии
Расовые различия в интеллекте
205
и Европы. Достижения рас в осуществлении неолитического перехода, в создании ранних цивилизаций и в развитии зрелых цивилизаций за последние две тысячи лет показывают, что различия в интеллекте имеют многотысячелетнюю историю и являются ещё одним выражением генетически обусловленных расовых различий в интеллекте.
В-восьмых, во всех близнецовых исследованиях, выполнявшихся в Европе, Индии и Японии, а такжесреди чернокожих и белых в Соединенных Штатах, была выявлена высокая наследуемость интеллекта внутри национальных популяций. Невероятно, чтобы эта высокая наследуемость внутри рас могла сосуществовать с отсутствием какой-либо наследуемости межрасовых различий.
В-девятых, имеются расовые различия в размере мозга, которые связаны с различиями в интеллекте, а наследуемость размера мозга составляет 90 процентов (Вааге et al., 2001; см. также Rushton, Osborne, 1995). Единственной разумной интерпретацией этой связи является то, что у рас с более высоким интеллектом развился более крупный мозг, обеспечивающий им более высокий IQ. Этот вопрос более детально рассматривается в параграфах 3-6 Главы 16.
В-десятых, постоянство всех расовых различий в самых разных странах, в успешности развития ранних и более поздних цивилизаций и высокая наследуемость интеллекта везде, где это было исследовано — всё это следует рассмотреть в терминах теории логики научного объяснения Карла Поппера (Popper, 1959). Она гласит, что научная теория генерирует предсказания, которые доступны эмпирической проверке. Сильная теория базируется на небольшом числе предположений и позволяет сделать большое количество предсказаний, которые верифицируются опытным путем. Если предсказания не подтверждаются, теория теряет силу и может вообще быть отброшена, хотя единственный противоречащий факт обычно может быть объяснен, или же теория может быть модифицирована с его учётом. Что касается проблемы расовых различий в интеллекте, то теория, согласно которой они до некоторой степени генетически детерминированы, объясняет все многочисленные феномены, перечисленные выше, серьёзные аномалии отсутствуют. Теория, согласно которой расовые различия в интеллекте в значительной степени детерминированы генетически, удовлетворяет критерию сильной теории Поппера. Те, кто утверждают, что очевидных свидетельств в пользу генетической основы расовых различий в интеллекте нет, обнаруживают своё недостаточное понимание логики научного объяснения.
206
Ричард Линн
Генотипно-средовая ковариация
Проблема оценки сравнительных вкладов средовых и генетических факторов в расовые различия в интеллекте осложнена действием принци- пома генотипно-средовой ковариации. Этот принцип гласит, что генам, определяющим высокий интеллект, обычно сопутствует благоприятная среда, обеспечивающая оптимальное развитие интеллекта (Plomin, 1994). Так, интеллектуальные женщины, будучи беременными, обычно воздерживаются от курения, от чрезмерного употребления алкоголя и приёма наркотиков, так как знают, что это может нарушить рост мозга и в будущем интеллект вынашиваемых ими детей. Интеллектуальные родители стараются обеспечить своим детям хорошее питание, потому что им понятны общие принципы здорового питания, а здоровое питание — детерминанта интеллекта. Интеллектуальные родители также с большей вероятностью будут стимулировать познавательные способности своих детей, что по широко распространённому убеждению (не обязательно верному) способствует развитию детского интеллекта. Тот же принцип работает и в случае рас. Расы с высоким интеллектом обычно обеспечивают своим детям двойное преимущество передавая им благоприятные гены и предоставляя благоприятную среду обитания с хорошим питанием, здравоохранением и, возможно, образованием, что улучшает развитие интеллекта их детей. Напротив, дети менее интеллектуальных рас обычно находятся во вдвойне неблагоприятном положении за счёт плохой наследственности и плохих условий среды. Так что проблематично, следует ли считать плохое питание и здравоохранение, снижающие интеллект многих народов третьего мира, чисто средовым или же до некоторой степени генетическим эффектом, являющимся результатом низкого интеллекта популяций, что делает их неспособными обеспечить хорошее питание и здравоохранение для своих детей. Принцип генотипно-средовой ковариации подразумевает, что расовые различия в интеллекте, непосредственной причиной которых являются средовые различия, можно также отнести на счёт генетических факторов, которые вносят свой вклад в средовые различия.
Трудно не придти к выводу, что расовые различия в интеллекте определяются как средовыми, так и генетическими факторами. Степень наследуемости расовых различий в интеллекте, надо ожидать, варьирует в зависимости от того, какие пары рас сравниваются. Величина наследуемости зависит от вариабельности средовых детерминант интеллекта в популяции, а в случае двух популяций — от различий в средовых детерминантах между двумя популяциями. В случае сравнения африканцев в Африке и европейцев, средовые различия между двумя популяциями в качестве пи
Расовые различия в интеллекте
207
тания, здравоохранения и образования весьма значительны. Следовательно, они окажут существенное влияние и, возможно, объяснят около 50 процентов различия в интеллекте между этими двумя популяциями (см. Главу 4). В случае сравнения африканцев в Соединенных Штатах и европейцев средовые различия между этими популяциями намного меньше, так как у них примерно одинаковы качество питания и образования, так что эффект среды намного меньше, а наследуемость, соответственно, больше. Аналогично, в случае сравнения аборигенов Восточной Азии и европейцев, условия их жизни весьма сходны, поскольку уровни благосостояния, питания, здравоохранения и образования у них примерно одинаковы; так что немного более высокий IQ аборигенов Восточной Азии, вероятно, в значительной мере детерминирован генетически.
HIM IE
жищни шпимшп
Общие принципы эволюции интеллекта
Млекопитающие
Птицы
Приматы
Гоминиды
Ю обезьян, человекообразных обезьян, и гоминид до Homo sapiens
Вернёмся теперь к вопросу о том, как развился интеллект. На всём протяжении эволюции биологических видов прослеживается общая тенденция развития более высокого интеллекта. В этой главе рассматриваются механизмы, ответственные за это. Эволюция расовых различий в интеллекте продолжает эту тенденцию и обсуждается в двух последующих главах.
Общие принципы эволюции интеллекта
Общие принципы эволюции интеллекта в сторону увеличения в течение приблизительно 225 миллионов лет были сформулированы Джерисо- ном (Jerison, 1973, 2000). Он показал, что имели место два процесса. Во- первых, время от времени биологические виды занимают новые среды обитания или ниши, что требует повышенной познавательной активности. Когда это происходит, виды отвечают развитием большего мозга, обеспечивающего более высокий интеллект. Это позволяет живым организмам справляться с познавательными требованиями новой ниши. Второй процесс состоял в том, что плотоядные и травоядные животные были втянуты в своего рода «гонку вооружений», в которой плотоядные животные должны были стать более интеллектуальными, чтобы ловить травоядных животных, в то время как травоядные должны были стать более интеллектуальными, чтобы спасаться от хищников. Полезное рассмотрение этого процесса было сделано Доукинсом и Кребсом (Dawkins, Krebs, 1979).
При межвидовых сравнениях по размеру мозга и интеллекту возникает проблема, вызванная сильной кросс-видовой связью размеров мозга и тела. Причина этого в том, что значительная часть мозга обслуживает
Расовые различия в интеллекте
209
Таблица 15.1. Эволюция коэффициентов энцефализации

функции тела, ввиду чего виды с крупным телом имеют большой мозг. Чтобы учесть размер тела при межвидовых сравнениях размера мозга, Джерисон ввёл понятие коэффициента энцефализации (EQ), представляющий собой показатель размера мозга относительно размера тела. Он установил величину EQ для среднего живущего ныне млекопитающего в 1,0 и выразил значения EQ других вымерших и живущих ныне видов относительно этого
210
Ричард Линн
стандарта. Джерисон определяет интеллект биологических видов как их EQ, который обуславливает способность мозга к обработке информации.
Важные моменты в развитии более высокого EQ при появлении новых видов представлены в Таблице 15.1. Эти данные были скомпилированы из работ Джерисона (Jerison, 1973,2000), Катлера (Cutler, 1976), и Харви и Глуттон-Брока (Harvey, Glutton-Brock, 1985).
В строках 1, 2 и 3 показано, что 225 миллионов лет рыбы и рептилии имели низкий EQ, равный 0,05, и что он не претерпел увеличения ни 60 миллионов лет назад, ни к настоящему времени.
Млекопитающие
Из строки 4 видно, что EQ первых млекопитающих, которые появились приблизительно 225 миллионов лет назад, составлял 0,25. Это было пятикратным увеличением по сравнению с рептилиями, от которых они эволюционировали, и первым квантовым скачком в увеличении EQ и интеллекта.
Объяснение такого развития состоит в том, что рептилии были в значительной степени дневными животными, получавшими информацию об окружающем мире главным образом через зрение. Подобно живущим ныне рептилиям, их поведение состояло в значительной степени в жёстких ответах на визуальные знаки-стимулы. Первые млекопитающие были маленькими животными размером примерно с крысу. Они заняли ночную нишу: в течение дня спали, а ночью добывали пропитание. Эта ниша представляла преимущество ввиду того, что давала защиту от хищных рептилий, но и имела недостаток, так как зрение ночных животных плохо подходило для сбора информации об окружающем мире, хотя положение улучшалось в сумерках, на рассвете и лунными ночами. Выходом из этого положения явилось развитие у ранних ночных млекопитающих слуха, обоняния, осязания и «интегрирующего процессора» для получения и анализа информации от этих трёх органов чувств наряду со зрением. Тогда они стали в состоянии объединять информацию, получаемую от этих четырёх органов чувств, чтобы идентифицировать хищников, пищу и половых партнёров. Развитие способности к обработке слуховой, обонятельной и осязательной информации потребовало увеличения слуховых, ольфакторных и тактильных центров мозга, а также развития интеграционной способности для объединения информации, полученной от этих четырёх органов чувств. Эти новые познавательные функции потребовали пятикратного увеличения коэффициента энцефализации по сравнению со средним значением у рыб и рептилий (от 0,05 до 0,25).
Расовые различия в интеллекте
211
Из строки 5 видно, что 60 миллионов лет назад EQ средних млекопитающих увеличился до 0,75, что представляет тройное увеличение в сравнении с 0,25 первых млекопитающих. В строке 6 показано, что за следующие 60 миллионов лет EQ среднего млекопитающего ещё увеличился до 1,0. Таким образом, за 225 миллионов лет, прошедших с их появления, EQ средних млекопитающих увеличился приблизительно в четыре раза. Как представляется, это увеличение имело место в значительной степени вследствие действия принципа «гонки вооружений» между плотоядными и травоядными животными, которые оказывали давление отбора друг на друга в сторону увеличения интеллекта и соответствующего ему EQ.
Птицы
Как следует из строки 7, первые птицы появились приблизительно 150 миллионов лет назад. У первой птицы, археоптерикса, EQ составлял 0,10 — вдвое выше значения рептилий, от которых они эволюционировали. Это был второй квантовый скачок в увеличении EQ и интеллекта. В строках 8 и 9 показано, что EQ птиц увеличился приблизительно до 0,75 60 миллионов лет назад и ещё возрос до 1,0 за следующие 60 миллионов лет к настоящему времени. Таким образом, у средних живущих ныне птиц примерно такой же EQ, как и у средних живущих ныне млекопитающих. Объяснение увеличения EQ птиц следует искать, вероятно, в том, что нишей их обитания стала преимущественно воздушная среда. Это давало преимущество: они находились вдалеке от хищников. Невыгодным было то, что птенцов, вылупившихся в гнездах на вершинах деревьев нужно было кормить в течение нескольких недель, пока те не вырастут в достаточной степени для того, чтобы летать и заботиться о себе самостоятельно. Чтобы вырастить своих птенцов, родители должны были построить гнездо, запомнить положение гнезда на местности, создать брачную пару и сотрудничать в кормлении своих птенцов и в защите гнезда от хищников. Эти задачи требовали, очевидно, более высоких интеллекта, способности к обучению и EQ, чем те, которые были необходимы рыбам и рептилиям, не заботящимся о своей молоди. Более высокий интеллект птиц и млекопитающих, таких как собаки и кролики, был выявлен с помощью различных экспериментальных задач, это рассматривается в обзоре Разрина (Razrin, 1971). Возрастание со временем EQ птиц происходило, вероятно, в значительной степени вследствие «гонки вооружений» между хищными и нехищными птицами, которые оказывали давление отбора друг на друга, приводящее к возрастанию интеллекта и соответствующего ему EQ.
212
Ричард Линн
Приматы
В строке 10 представлен EQ первых приматов (0,75), появившихся приблизительно 60 миллионов лет назад вслед за исчезновением динозавров. Величина EQ первых приматов была примерно такой же, как и у живших тогда средних млекопитающих и птиц. В строках с 11 по 15 представлены значения EQ живущих ныне представителей первых приматов: древесной землеройки (0,85), западноафриканского лемура потто (1,1), галаго (1,2), изящного лемура (0,7) и чёрного лемура (1,6). Средний EQ этих пяти живущих ныне видов составляет 1,1; увеличение в сравнении с первыми приматами, жившими 60 миллионов лет назад, равняется приблизительно 50 процентам. В строке 16 приведён EQ первых обезьян (1,0), появившихся около 30 миллионов лет назад. В строках с 17 по 22 даны EQ шести типичных видов живущих ныне обезьян. Величины их EQ располагаются в пределах от 1,3 у серебристого лангура до 3,5 у бурого черноголового капуцина, так что все они имеют более высокий EQ, чем первые, жившие 30 миллионов лет назад обезьяны, с их EQ 1,0. В строке 23 приведён EQ первых человекообразных обезьян (2,0), появившихся примерно 16 миллионов лет назад. Основные отличия человекообразных от остальных обезьян заключаются в том, что у человекообразных обезьян нет хвоста и у них более гибкие плечи, что позволяют им поднимать руки выше головы и повисать на ветвях деревьев, тогда как другие обезьяны бегают по веткам. В строках с 24 по 28 даны EQ пяти видов живущих ныне крупных человекообразных обезьян. Значения их EQ следующие: 2,0 у горилл Центральной Африки, 2,1 у гиббона сиаманга из Юго-Восточной Азии и Индонезии, 2,4 у орангутанга с о-вов Борнео и Суматры, 2,6 у шимпанзе из Центральной Африки и 2,8 у белорукого гиббона или лара из Юго-Восточной Азии и Индонезии. Если рассматривать крупных человекообразных обезьян как род, то можно сказать, что у них, по-видимому, не развился более высокий EQ, чем у других обезьян. Средний EQ пяти видов крупных человекообразных обезьян равен 2,4, а у шести видов других обезьян он составляет 2,3. Некоторые из этих EQ определены по малому числу измерений и могут быть недостаточно точны вследствие ошибок выборки.
Стремительное развитие EQ у обезьян и человекообразных обезьян от 1,0 до 2,4 за 30 миллионов лет их существования было намного более быстрым, чем у других млекопитающих и птиц за тот же самый период. Это было третьим большим квантовым скачком в увеличении размера мозга и интеллекта. Представляется, что для столь быстрого увеличения EQ были две причины. Во-первых, в то время как ранние приматы, подоб
Расовые различия в интеллекте
213
но млекопитающим, от которых они эволюционировали, были ночными животными, (Byrne, 2002), обезьяны и человекообразные обезьяны стали животными дневными, бодрствующими днём и спящими ночью. Дневные виды получают информацию о внешнем мире в значительной степени посредством зрения, в соответствии с этим визуальные центры мозга обезьян и человекообразных обезьян увеличились в размере, обеспечив им повышенную способность к обработке визуальной информации. Во-вторых, они жили социальными группами, тогда как ранние приматы вели одиночный образ жизни. Проживание группами даёт преимущества в обеспечении исключительного использования территории и её ресурсов, в сотрудничестве в поиске пищи, в заботе о молодняке и в защите от хищников. Платой за это является необходимость усвоения особями сложных социальных навыков для гармоничного сосуществования с другими участниками группы, с которыми они должны сотрудничать при защите территории, но с которыми они и конкурируют за пищу и половых партнёров. Социальная система этих животных обычно представляет собой группу примерно от тридцати до восьмидесяти особей, в которой существует иерархия подчинения, где два-три доминирующих самца получают больше пищи, исключительный доступ к самкам, когда у тех происходит течка, и лучшие спальные места на деревьях. Чтобы сохранить своё положение, доминирующие самцы обычно формируют союзы для противодействия претензиям со стороны не доминирующих самцов. Не доминирующие самцы принадлежат к группе, однако должны проявлять уважительное отношение к положению доминирующих самцов, которые изгонят их из стаи, если почувствуют, что им брошен вызов. Тем не менее, не доминирующие самцы, как представляется, понимают, что если они будут вести себя искусно, настанет время, когда доминирующие самцы состарятся и ослабнут, и в конце концов умрут, и они смогут унаследовать их место. Чтобы поддержать своё положение в группе в ожидании этой возможности, не доминирующие самцы должны проявлять сдержанность и осмотрительность до тех пор, пока им не представится удобный случай успешно оспорить права доминирующих самцов и сместить их. До тех пор они вступают в союзы с другими не доминирующими самцами, чтобы поддержать своё положение в группе и увеличить свои шансы стать доминирующими. Приобретение этих социальных навыков требует быстрого научения и способности сдерживать претензии к доминирующим самцам. Эти социальные навыки получили название «социального интеллекта» и они, по-видимому, требуют относительно высокого EQ для понимания и управления социальными отношениями, наблюдения, фиксации и запоминания особенностей других участников группы, а также для подавления импульсивных действий. Самцы с высоким социальным интеллектом в конечном итоге становятся доминирующими и приобретают возможность размножаться, что повышает
214
Ричард Линн
социальный интеллект вида. Теория, что становление высоко социальными животными было нишей, которая повысила EQ обезьян и человекообразных обезьян, была разработана Данбаром (Dunbar, 1992). Он показал, что среди приматов размер социальной группы вида коррелирует с EQ; это говорит о том, что приматам, живущим большими группами, требуется более высокий EQ для участия в более сложных социальных отношениях между членами группы. Таким образом, обезьяны и человекообразные обезьяны заняли новую нишу как кооперированные социальные виды, что требовало более высокого интеллекта, обеспечиваемого более высоким EQ.
Обезьяны и человекообразные обезьяны демонстрируют разнообразные проявления высокого интеллекта, что согласуется с высокими значениями их EQ. Наиболее изученным видом является шимпанзе. В 1920-х гг. Кёлером было показано (Kohler, 1925), что когда перед шимпанзе ставят сложную задачу, например, достать банан, подвешенный к потолку слишком высоко, чтобы до него можно был дотянуться, они могут сообразить, как из ящиков можно построить платформу, на которую они могут забраться и схватить этот банан. Позже Гудолл показала (Goodall, 1986), что шимпанзе в дикой природе научаются изготавливать и использовать орудия для различных целей. Они берут ветки, от которых отрывают боковые отростки, облизывают их, чтобы сделать липкими, вставляют их в отверстия в горках термитов или гнездах Муравьёв, вытаскивают липкие ветки и съедают термитов или муравьев, прилипших к ним. Они делают пестики, чтобы измельчить древесную мякоть в съедобную пасту, долота чтобы открыть пчелиные гнёзда, используют камни для раскалывания орехов и листья как чаши для питья и чтобы почистить себя, а также поднимают упавшие сучья деревьев для обороны от хищников и при защите своей территории от вторгшихся чужаков. У них также имеется словарь приблизительно из дюжины криков, служащих для передачи информации о присутствии хищников, о вторжении на их территорию соседних групп обезьян, о расположении источников пищи, о готовности или нежелании поделиться пищей и так далее. Недавно было показано, что орангутанги также изготавливают и используют орудия (Fox, Sitompul, Van Schaik 1999). В лабораторных исследованиях только обезьяны и человекообразные обезьяны могут справиться с задачами выбора непохожего предмета, в которых предлагаются три предмета, два из которых одинаковы, и нужно выбрать третий лишний, а также обучаться с первого раза, когда предъявляются два различных объекта и правильный выбор меняется день ото дня.
Расовые различия в интеллекте
215
Гоминиды
Четвертый квантовый прыжок в EQ и интеллекте имел место с эволюцией гоминид. Это ряд видов, который в итоге привел к появлению Homo sapiens— Человека разумного. Началось это приблизительно 4 миллиона лет назад в Центральной Восточной Африке (там, где теперь находятся Кения и Танзания) с появлением австралопитеков, затем последовательно появились три вида: Homo habilis, Homo erectusи, наконец, Homo sapiens. Время существования этих видов и их EQ приведены в строках 29-32 Таблицы 15.1. Первый из них, австралопитеки, включал несколько видов. Первым появивился Australopithicus afarensis,эволюционировавший от человекообразной обезьяны, очень похожей на шимпанзе. За следующие два миллиона лет развились другие виды австралопитеков, включая africanus, robustusи boisei.Особи последних видов были крупнее и имели больший мозг, но не в пропорции к размеру тела, так что их коэффициенты энцефализации оставались теми же. Причина эволюции австралопитеков состояла в том, что человекообразные обезьяны приспособлены к жизни в лесах, но в Центральной Восточной Африке климат стал более засушливым, в результате чего большая часть лесов исчезла и сменилась лугами с вкраплениями кустарников и отдельных рощ деревьев. Таким образом, человекообразные обезьяны в Центральной Восточной Африке должны были приспособиться, чтобы выжить в новой нише открытой саванны. Тремя главными адаптивными признаками стали: прямохождение (тогда как обезьяны передвигаются на четвереньках), противопоставление большого пальца остальным и увеличение EQ. Адаптивное значение вертикального положения тела состояло в том, что оно предоставляло лучший обзор, позволяющий видеть хищников на большем расстоянии, давало возможность совершать длительные переходы в поисках пищи и освобождало руки. Освобождение рук и противопоставление большого пальца остальным позволило использовать руки для переноса пищи на стоянку, изготавливать каменные орудия и более ловко захватывать камни и куски дерева и прогонять ими хищников.
Величина EQ гоминидов увеличилась приблизительно втрое в течение примерно 4 миллионов лет от приблизительно 2,6 у человекообразных обезьян, от которых они эволюционировали, до 7,5 у Homo sapiens. Это было очень большой скоростью увеличения по сравнению с таким же увеличением за примерно 56 миллионов от 0,75 у первых приматов около 60 миллионов лет назад до 2,6 у наиболее энцефализированных обезьян и человекообразных обезьян. Объяснением такого увеличения является то,
216
Ричард Линн
что гоминиды заняли новую нишу открытой саванны, выживание в которой предъявляло более высокие требования к познавательной сфере, чем у человекообразных обезьян, от которых они эволюционировали. Требования к познавательной сфере в этой новой нише состояли преимущественно в том, что надо было находить множество различных видов пищи и защищать себя от хищников. Австралопитеки и последующие гоминиды, как и человекообразные обезьяны, от которых они эволюционировали, продолжали в значительной степени питаться растительной пищей, но в открытой саванне она была более разнообразной и распределенной по большей территории, чем у человекообразных обезьян. Чтобы найти эту пищу, гоминидам требовалось запоминать планы обширных территорий, а это требовало большего мозга. Виды пищи, поедавшейся ими, можно определить по изнашиванию зубов, которое показывает, что их рацион состоял в значительной степени из листьев и фруктов, но они также поедали клубни, орехи, семена трав и насекомых (Isaac, 1978; Parker, Gibson, 1977; Grin, Kay, 1988; Stahl, 1984). Некоторые из гоминидов жили по берегам озер Баринго и Туркана в сегодняшней Кении. Здесь они могли собирать моллюсков и разбивать их ударяя камнями, которые были в состоянии удерживать между большим пальцем и остальными пальцами рук.
Гоминиды пополняли свой рацион из растений и насекомых некоторым количеством мяса, подбирая останки животных и, возможно, иногда убивая мелких животных. Бабуины и шимпанзе иногда убивают мелких животных на корм, хотя мясо всегда остаётся лишь небольшой частью их рациона (Strum, 1981). Возможно, австралопитеки и более поздние гоминиды, Homo habilis, делали то же самое. Кроме того они собирали останки животных, задранных львами, гепардами и леопардами. На стоянках Homo habilisнаходят кости крупных травоядных животных со следами зубов хищников, поверх которых имеются следы, нанесённые гоминида- ми камнем. Это говорит о том, что эти крупные травоядные животные были убиты львами, гепардами и леопардами, и что Homo habilisподбирали кости и разбивали их для извлечения костного и головного мозга, до которых не могли добраться хищники (Binford, 1985; Blumenschine, 1989). Homo habilisс их EQ, увеличившемся до 4,3, стали первыми гоминидами с мозгом, дающим возможность изготавливать каменные орудия широкого ассортимента. Они откалывали куски кремня для производства острых режущих орудий, которые использовали для изготовления оружия, такого как копья, и ножей для расчленения туш крупных млекопитающих, задранных львами, гепардами и леопардами.
Помимо поиска пищи, другой главной проблемой гоминидов, живущих в открытых саваннах, являлась защита ото львов, гепардов и леопардов. Человекообразные и остальные обезьяны убегают от крупных кошек, забираясь на деревья или перепрыгивая с одного дерева на другое. Для
Расовые различия в интеллекте
217
австралопитеков и более поздних гоминидов в открытой саванне такой возможности больше не было. Они должны были отражать нападения львов, леопардов и гепардов бросая в них камни и поражая их дубинами, сделанными из сучьев немногих оставшихся деревьев. Развившиеся у них недавно большие пальцы, обеспечивающие большую силу захвата, должны были давать им в этом отношении большое преимущество. Шимпанзе иногда используют палки для отражения хищников, но они не собирают арсенал палок и камней для этой цели. Австралопитеки должны были делать это, что должно было требовать большей способности к предвидению и большего интеллекта. По предположению Александера (Alexander, 1989), ещё одно давление отбора, увеличившее EQ гоминидов, состояло, вероятно, в том, что более интеллектуальные особи были более искусными производителями орудий и охотниками и имели более высокий социальный интеллект, обеспечивающий им более высокий ранг в иерархии подчинения, благодаря которому они производили больше потомства.
Величины IQ обезьян, человекообразных обезьян и гоминид до Homo sapiens
Делались попытки оценить интеллект обезьян, человекообразных обезьян и гоминид до Homo sapiens,основываясь на теории Пиаже о развитии интеллекта у детей. Согласно теории Пиаже, дети проходят четыре стадии познавательного развития. Первая из них — сенсомоторная стадия, на которой ребенок узнает о свойствах объектов, места, времени и причинности. Приблизительно в возрасте двух лет дети совершают переход к «до- операционной» стадии, на которой они овладевают языком и абстрактными понятиями, но еще не в состоянии понять логические принципы. Эта стадия длится до возраста примерно шести лет. В западных обществах дети примерно в семь лет переходят к стадии «конкретных операций», когда они могут усвоить логические принципы, но только на конкретном материале. Примерно в возрасте 12 лет дети переходят к четвертой и заключительной стадии «формальных операций», когда они становятся способными мыслить логически в терминах общих принципов, абстрагированных от конкретных примеров.
Обзор попыток применения этой теории к интеллекту обезьян, человекообразных обезьян и дочеловеческих гоминидов представлен в монографии Паркера и МакКинни (Parker, McKinney, 1999). Согласно их заключению, обезьяны большинства видов не продвигаются дальше первой стадии Пиаже и остаются, таким образом, на когнитивном уровне примерно двухлетних детей. По шкале человеческого интеллекта их IQ составил
218
Ричард Линн
бы приблизительно 12 баллов. Человекообразные обезьяны находятся на ранних этапах дооперационной стадии Пиаже и достигают когнитивного уровня среднего европейского 3-4-летнего ребёнка. Их IQ составил бы примерно 22 балла. Попытки оценить уровень интеллекта по Пиаже, достигаемый последовательно появлявшимися видами гоминидов, исходя из изготавливаемых ими орудий, предпринимались Винном (Wynn, 1989). Согласно его заключению, Homo habilis,обитавшие в Восточной Африке приблизительно 2,4 миллиона лет назад, изготавливали простые каменные орудия, для чего им надо было находиться на первых этапах дооперационной стадии, примерно на тех же самых, на которых находятся человекообразные обезьяны. Homo erectus,появившиеся приблизительно 1,7 миллионов лет назад и имевшие несколько больший мозг, изготавливали более сложные ашельские11 каменные орудия, включая двусторонние ручные рубила, что требует мышления в стадии конкретных операций на уровне, достигаемом современными европейскими 7-8-летними детьми. Из этого можно вывести, что их IQ должен был составлять приблизительно 50 балов.
11 Характерные для Ашельской культуры раннего палеолита, названной по предместью г. Амьен (Сент-Ашель, Saint-Acheul) во Франции. Основные орудия — каменные ручные рубила. Хозяйство: охота и собирательство {прим. перев.).
тыic
mmiмел. тт моэгд и шгомш
Эволюция рас
Требования к когнитивной сфере в северных широтах
Расовые различия в размере мозга
Расовые различия: зимние температуры, размер мозга и Ю
Размер мозга и интеллект человека
Вклад расовых различий размера мозга в различия в интеллекте
Половые различия в интеллекте и размере мозга
Генетические процессы в эволюции расовых различий в Ю
В этой главе рассматриваются общие принципы развития расовых различий в интеллекте. Ключевым фактором, ответственным за развитие расовых различий в интеллекте, признаётся умеренный и холодный климат Северного полушария, предъявляющий повышенные «когнитивные требования» для выживания и действующий как давление отбора в сторону увеличения интеллекта. Аборигены Южной Азии и Северной Африки, европейцы, аборигены Восточной Азии, арктические народы и американские индейцы адаптировались к этим когнитивным требованиям, развив больший мозг и интеллект. Ответственные за это генетические процессы состояли в увеличении частот аллелей высокого интеллекта и в мутациях, обеспечивающих повышение интеллекта.
Эволюция рас
Общепризнанным в теории эволюции рас является то, что человек эволюционировал от человекообразных обезьян в Африке южнее Сахары в течение последних четырёх миллионов лет или около того. За это время последовательность видов, известных под собирательным названием гоминидов, эволюционировала в сторону увеличения мозга. Это были австралопитеки, за ними последовали Homo habilis,затем Homo erectus,появившиеся
220
Ричард Линн
примерно 1,5 миллиона лет назад, и, наконец, Homo sapiens (современный человек), появившийся приблизительно 150.000 лет назад (Relethford, 1988). Примерно 100.000 лет назад группы Homo sapiens начали мигрировать из экваториальной Африки в другие области мира и приблизительно 30.000 лет назад колонизировали большую часть земного шара. В начале этого периода они распространились по большей части Африки южнее Сахары и 100.000 лет назад они уже населяли Юг Африки, где эволюционировали в бушменов. Примерно 88.000 лет назад они обосновались в Юго-западной Азии. В период от 60 до 40 тыс. лет назад они расселились по всей Азии и примерно 40 тыс. лет назад заселили Европу, Индонезийский архипелаг, Австралию и оба континента Америки. В течение последних 6.000 лет или около того они колонизировали острова Тихого океана (Foley, 1997; Mellars, Stringer, 1999; Cavalli-Sforza, 2000). На рисунке 1 представлена составленная по данным археологических находок карта, показывающая приблизительное время и направления миграций современного человека.

Рисунок 1. Миграции современного человека, начавшиеся в Африке приблизительно 100.000 лет назад.
Когда популяции пространственно изолированы друг от друга, в них неизбежно развиваются генетические различия и они эволюционируют в разные породы или, в случае человека, расы. Это — общий принцип эволюционной биологии. Эти различия развиваются вследствие эффекта основателя, дрейфа генов, мутаций и адаптации к различной среде обита
Расовые различия в интеллекте
221
ния. Эффект основателя имеет место, когда небольшая группа отделяется от популяции, мигрирует к новому местообитанию и образует новую популяцию. Мигрирующая группа, вероятно, будет характеризоваться случайными генетическими отличиями от оставшейся, что приводит к возникновению двух групп с различными генетическими особенностями. Считается маловероятным, что этот процесс играл какую-либо существенную роль в развитии генетических различий в интеллекте между расами. Второй процесс, посредством которого расы расходятся генетически, это дрейф генов. Он состоит в том, что частоты одних генов увеличиваются, а других уменьшаются случайным образом. Возможно, расовые различия в частотах встречаемости разных групп крови и генетических болезней могли возникнуть таким образом, но вероятность того, что этот процесс играл сколько-нибудь заметную роль в развитии расовых различий в интеллекте, также считается невысокой. Полагают, что за появление расовых различий в интеллекте ответственны два остающихся процесса: адаптация к различной окружающей среде и генетические мутации, появившиеся у одних рас и не появившиеся у других.
Многие из различий человеческих рас могут быть истолкованы как адаптация к климату. Морфологические различия развились в соответствии с правилом Аллена, согласно которому у биологических видов и пород в холодных регионах обычно развиваются более короткие конечности, потому что в результате этого снижается отношение поверхности тела к его объёму, а это уменьшает потерю тепла. Поэтому у аборигенов Восточной Азии и европейцев в умеренном и холодном климате конечности короче, чем у африканцев в тропическом и субтропическом климате. Тёмная кожа африканцев и австралийских аборигенов, живущих в тропиках и субтропиках, обеспечивает защиту от солнечных ожогов и рака кожи; отсутствие растительности на лице у мужчин в Восточной Азии предотвращает обморожение из-за примерзания волос к лицу; меньшие, чем у африканцев и австралийских аборигенов, ноздри аборигенов Восточной Азии и европейцев согревают и увлажняют вдыхаемый воздух (Coon, 1962; Krantz, 1980).
Требования к когнитивной сфере в северных широтах
Давлением отбора в сторону повышения интеллекта, действующим на народы, мигрировавшие из тропической и субтропической экваториальной Африки в Северную Африку, Азию, Европу и в Америку, явилась проблема выживания зимой и весной в умеренном и холодном климате.
222
Ричард Линн
Это была новая, предъявляющая повышенные когнитивные требования среда обитания, где было необходимо охотиться на крупных животных для пропитания и сохранять тепло, для чего требовалось строить убежища, добывать огонь и изготавливать одежду. Кроме того, как предположил Миллер (Miller, 2005), в умеренном и холодном климате женщины стали зависимыми от мужчин в части обеспечения пищей, потому что они были неспособны охотиться, тогда как в тропиках женщины были в состоянии собирать для себя растительную пищу. Вследствие этого женщинам должен был требоваться более высокий интеллект, чтобы выбирать себе в супруги мужчин, способных их прокормить. По всем этим причинам умеренный и холодный климат должен был оказывать давление отбора в сторону увеличения интеллекта. Чем холоднее были зимы, тем сильнее было это давление и тем выше развившийся интеллект. Это объясняет определённую связь между широтой или, точнее, суровостью зимы и интеллектом рас.
Теория о том, что расовые различия в интеллекте развились вследствие попадания людей, мигрировавших из Африки в зону умеренного и холодного климата Азии и Европы, в экологическую нишу, предъявляющую более высокие когнитивные требований, где был нужен более высокий интеллект, является ещё одним примером действия общего принципа, который работал в эволюции более высокого интеллекта у млекопитающих, колонизировавших ночную нишу, у птиц, при освоении ими воздушной среды, у обезьян и человекообразных обезьян, когда они стали кооперированными социальными животными, и у гоминидов, адаптировавшихся к жизни в открытой саванне. Новая ниша умеренного и холодного климата, которую колонизировали мигрировавшие из Африки расы, потребовала перехода от травоядного к преимущественно плотоядному образу жизни. Приматы, от которых произошли люди, обитали приблизительно 60 миллионов лет назад в тропическом и субтропическом поясах экваториальной Африки, где растительная пища доступна в течение всего года, и были травоядными. Гоминиды, которые эволюционировали в экваториальной Восточной Африке, оставались в значительной степени травоядными. Современные племена охотников-собирателей в тропических и субтропических широтах продолжают питаться в основном растительной пищей, многочисленные виды которой доступны в течение всего года (Lee, 1968; Tooby, de Vore, 1989).
Поскольку приматы адаптированы как травоядные животные к тропической и субтропической среде, им оказалось непросто выжить в умеренном климате, где растительная пища недоступна в течение нескольких месяцев зимой и весной. Впервые приматы столкнулись с проблемой выживания зимой и весной в умеренном климате в среднем миоцене от 16 до 14 миллионов лет назад. Это был тёплый период, тогда большая часть Ев
Расовые различия в интеллекте
223
разии стала субтропической. Два вида человекообразных обезьян, плио- питеки и дриопитеки, мигрировали из Африки в Евразию и «процветали» там. В конце этого периода, приблизительно 14 миллионов лет назад, в Евразии похолодало, и климат там стал умеренным. В Европе и в большей части Азии эти человекообразные обезьяны не смогли выживать зимой и вымерли. Единственной частью Азии, где эти ранние человекообразные обезьяны смогли выжить, был её тропический Юго-Восток и Индонезийский архипелаг, где они эволюционировали в орангутангов и гиббонов (Pickford, 1986).
Расовые различия в размере мозга
У рас, мигрировавших в регионы Северной Африки, Азии, Европы и Америки с умеренным и холодным климатом, развился более высокий интеллект, благодаря чему они выжили в этой среде, предъявлявшей повышенные когнитивные требования. Для обеспечения болеевысокого интеллекта у них развился больший мозг, так же, как это имело место ранее в эволюции млекопитающих, птиц и приматов при их адаптации к предъявляющим более высокие когнитивные требования нишам. Результаты исследований расовых различий в размере мозга для каждой расы были представлены в Главах с 3 по 12. Эти данные нельзя усреднить, так как различные методы измерения размера мозга дают различающиеся результаты. Основными методами являются: измерение длины, ширины и высоты головы живых людей и вычисление объема мозга или заполнение черепов свинцовой дробью или мелкими семенами, которые затем пересыпают в мерный контейнер для определения объема. Требуется большой набор размеров мозга, измеренных одним и тем же методом, набор должен включать все расы. Доступен всего лишь один такой набор данных. Это — средние размеры мозга 87 популяций со всего мира, основанные на измерении примерно 20 тыс. черепов, они были опубликованы Смитом и Билсом (Smith, Beals, 1990). В Таблице 16.1 эти данные распределены по десяти рассматриваемым нами расам. Жирным шрифтом выделены средние значения выборок из каждой расы.
224
Ричард Линн
Таблица 16.1. Размеры мозга (смЗ) десяти рас

Расовые различия в интеллекте
225
Расовые различия: зимние температуры, размер мозга и IQ
Развитие мозга большего размера для обеспечения более высокого интеллекта у рас, заселивших среду с более холодным климатом, показано в Таблице 16.2.
Таблица 16.2. Расовые различия в зимних температурах (градусы Цельсия), размере мозга (см3) и IQ

В колонке 1 указаны расы, ранжированные по суровости зимних температур, действию которых они подвергаются. В колонке 2 даны сегодняшние средние температуры самого холодного зимнего месяца, взятые из Атласа мира Энциклопедии Британика, усреднённые для регионов, в которых проживает раса. В колонке 3 приведены средние температуры самого холодного зимнего месяца во время главного Вюрмского оледенения, длившегося в интервале приблизительно 28-10 тыс. лет назад, во время которого зимние температуры упали примерно на 5°С в Северном полушарии, но не в Южном (Roberts, 1989; Foley, 1987). В колонке 4 представлены средние размеры мозга, взятые из Таблицы 16.1. Очевидно, что в общем существует соответствие между средней температурой самого холодного зимнего месяца и размером мозга. Для шести рас, перечисленных первыми,
8 Ричард Линн
226
Ричард Линн
имеет место уменьшение размера мозга с повышением средней температуры самого холодного зимнего месяца. Однако, в случае остальных четырех рас, этот линейный тренд делается нерегулярным. Африканцы населяют более тёплую климатическую зону, чем бушмены, но имеют больший размер мозга. Австралийские аборигены «продолжают тренд»: они живут в более тёплом климате и у них размер мозга меньше. Но у аборигенов Юго-Восточной Азии и аборигенов островов Тихого океана в тропической и субтропической зонах размер мозга больше, чем у аборигенов Южной Азии и Северной Африки, бушменов, африканцев и австралийских аборигенов.
В колонке 5 приведены величины IQ рас. Здесь также очевидно, что в целом имеется соответствие между IQ, средней температурой самого холодного зимнего месяца и размером мозга, но вновь имеются аномалии. Во- первых, арктические народы населяют самую холодную зону и имеют наибольший размер мозга, но их IQ составляет всего 91 балл. Во-вторых, у бушменов мозг предпоследний по размеру (1.270 см3), но самый низкий IQ (54 балла), в то время как у австралийских аборигенов наименьший мозг (1.225 см3), но немного более высокий IQ (62 балла), чем у бушменов. Не считая этих аномалий, наблюдается полное соответствие между различиями рас в размере мозга и в IQ. Для объяснения этих аномалий мы должны рассмотреть генетические законы, имеющие отношение к эволюции расовых различий в интеллекте. Этот вопрос рассматривается в Параграфе 8.
Сходным образом к вопросу о связи между зимней температурой и интеллектом народов всего мира подошли Темплер и Арикава (Templer, Arikawa, 2005). Они исследовали связь между средним значением IQ у 129 этносов и самыми низкими зимними температурами. Корреляция составила -0,68. Это говорит о том, что чем ниже зимняя температура, тем выше IQ этноса. Они также изучили зависимость между средним IQ этноса и цветом кожи, и выявили корреляцию в 0,92: чем светлее кожа, тем выше IQ.
Размер мозга и интеллект человека
Таким образом, существует общая зависимость между суровостью зимы, размером мозга и интеллектом рас. Объяснением этого является то, что чем холоднее зимы в климатической зоне, в которой обитает раса, тем сильнее давление отбора в сторону увеличения мозга для обеспечения более высокого интеллекта, необходимого для решения задачи выживания в течение суровой зимы. Это теория предполагает, что у человека размер мозга положительно связан с интеллектом. Ряд антропологов и психологов явно не желают признать существование этой связи. К примеру, Рей-
Расовые различия в интеллекте
227
дел и соавт. (Reidel et al., 1994, стр. 533) сообщают об исследовании, в котором было показано, что средний вес мозга австралийских аборигенов существенно ниже, чем у европейцев, но пишут, что «должно быть подчёркнуто, что это отличие объёма мозга не может быть интерпретировано как свидетельство об интеллектуальных возможностях или об интеллекте аборигенов. Ни одно исследование никогда не выявляло корреляции между нейроанатомическими данными и интеллектом у человека». Брэдшоу (Bradshaw, 1997, стр. 145) пишет о «больном вопросе о возможной зависимости между размером мозга и интеллектуальными способностями внутри нашего вида». Билс, Смит и Додд (Beals, Smith, Dodd, 1984) пишут о связи между размером мозга и интеллектом: «Никогда не было представлено ни одного убедительного примера наличия такой связи». Аналогично: «Накоплено даже больше данных против прямой связи между объёмом черепа и силой интеллекта» (Hennenberg, 1984); «Действительно, нет данных, показывающих, что размер мозга положительно коррелирует с интеллектом» (Lathan, 1974); «Нет данных о том, что больший мозг в чём- либо лучше маленького мозга» (Halpem, 2000, стр. 196) и «Никогда не было выполнено научно приемлемого исследования, подтверждающего гипотезу о том, что интеллект (например, оценки IQ) зависит от размера мозга» (Graves, 2002, стр. 81).
Вопреки этим утверждениям, положительная связь между размером мозга и интеллектом человека была показана в многочисленных исследованиях, выполнявшихся начиная с первого десятилетия двадцатого века. Эти исследования рассмотрены в обзоре Вернона и соавт. (Vernon et al., 2000), обобщившими результаты 54 исследований, в которых использовались внешние измерения головы. Во всех исследованиях была выявлена положительная связь, коэффициент корреляции по всем исследованиям составил 0,18. Они также сообщают об 11 исследованиях в группах нормальных испытуемых, у которых определяли размер мозга с помощью аксиальной компьютерной томографии и магнитного резонанса, обеспечивающих более точное измерение размера мозга; корреляция по всем этим исследованиям составила 0,40. В другом исследовании, опубликованном позже этого обзора, у 40 испытуемых была выявлена корреляция между размером мозга (измерявшимся методом магнитного резонанса) и интеллектом, равная 0,44 (Thompson et al., 2001). Вернон и соавторы заключают, что наиболее разумная интерпретация этой корреляции состоит в том, что размер мозга является детерминантой интеллекта. В большем мозге больше нейронов, и это обуславливает его повышенную способность к обработке информации. Не только у,человека размер мозга коррелирует с интеллектом. Такая же связь была найдена у крыс в исследовании Андерсона (Anderson, 1993): способность крыс заучивать путь через лабиринты положительно коррелировала с весом их мозга.
228
Ричард Линн
В корреляцию между размером мозга и IQ в 0,40, полученную Верноном и соавт. (Vernon et al., 2000), следует внести поправку на ошибку измерения тестами интеллекта («поправка на аттенюацию»). Поправка на ошибку измерения вносится посредством деления корреляции на квадратный корень произведения коэффициентов надежности двух показателей, корреляция между которыми вычисляется. Надежность тестов интеллекта составляет обычно около 0,90 (Bouchard, 1993, стр. 49; Mackintosh, 1998). Надежность измерения размера мозга не известна, но она, как предполагается, должна быть идеальной. Внесение поправки в корреляцию между размером мозга и IQ (0,40) на неполную надежность тестов интеллекта (0,90) даёт истинный коэффициент корреляции в 0,44.
Вклад расовых различий размера мозга в различия в интеллекте
Теперь мы рассмотрим, в какой степени расовые различия в размере мозга могут объяснять различия в интеллекте. Чтобы сделать это, мы должны выразить расовые различия в размере мозга в единицах стандартного отклонения (d)и умножить значения dна корреляцию между размером мозга и интеллектом. Это даёт различия IQ рас, объясняемые различиями в размере мозга. Для этих вычислений требуются средние значения и стандартные отклонения размера мозга рас. Значения стандартных отклонений имеются лишь для европейцев, африканцев, американских индейцев, аборигенов Южной Азии и аборигенов Восточной Азии, они были представлены Силсом, Смитом и Доддом (Seals, Smith, Dodd, 1984). Таким образом, вычисления могут быть сделаны только для этих рас. Полученные результаты представлены в Таблице 16.3.
Таблица 16.3. Расовые различия в IQ, предсказанные по различиям в размере мозга

Расовые различия в интеллекте
229
В колонке 1 приведены пары сравниваемых рас. В колонке 2 представлены различия в размере мозга между этими двумя расами, выраженными в d(т.е. в единицах стандартного отклонения), вычисленные по данным, приведённым в Таблице 16.1. В колонке 3 даны различия IQ двух рас (выраженные в d), предсказанные по различию размера мозга, их вычисляли умножением значений d,приведённых в колонке 2, на 0,44 (корреляция между размером мозга и интеллектом с поправкой на ошибку измерения). В колонке 4 даны фактические значения IQ рас в сравнении с 99 баллами европейцев.
В строке 1 представлены эти значения при сравнении европейцев и африканцев. Величина IQ африканцев, предсказанная по различию в размере мозга, должна составлять 91 балл. В действительности их IQ составляет 67 баллов, таким образом, различие размера мозга предсказывает 28 процентов различия IQ. В Главе 4 генотипный IQ африканцев был определён в 80 баллов, так что различие размера мозга объясняет приблизительно половину различия генотипного IQ. Другая половина должна быть приписана различиям в нейрофизиологических процессах.
В строке 2 даны значения для сравнения европейцев с американскими индейцами. Величина IQ американских индейцев, предсказанная исходя из различий размера мозга, составляет 97 баллов. В действительности их IQ равен 86 баллам. Таким образом, различие в размере мозга предсказывает приблизительно одну пятую различия в IQ. В строке 3 даны значения для сравнения европейцев с аборигенами Южной Азии и Северной Африки. Предсказанный по различию в размере мозга IQ аборигенов Южной Азии и Северной Африки в составляет 96 баллов. Их фактический IQ составляет 84 балла, так что различие размера мозга предсказывает четверть различия IQ. В строке 4 представлены значения для сравнения европейцев с аборигенами Восточной Азии. Исходя из различия в размере мозга, IQ аборигенов Восточной Азии должен составлять 109 баллов. Их действительный IQ равен 105, таким образом, в этот раз различие в размере мозга переоценивает различие в IQ на 4 балла. Есть два правдоподобных объяснения этого. Первое: аборигены Восточной Азии находятся в неблагоприятной среде, которая препятствует реализации их генотипного IQ; если это так, то IQ аборигенов Восточной Азии возрастёт приблизительно до 109 баллов, когда условия их среды обитания улучшатся до европейского уровня. Второе: большой мозг аборигенов Восточной Азии обслуживает когнитивные способности, не полностью представленные в тестах интеллекта. Наиболее вероятно, что это визуализационная способность.
Хотя вклады расовых различий размера мозга в различия IQ рас могут быть вычислены только для пар рас, представленных в Таблице 16.3, результаты, показывающие что расовые различия в размере мозга объясняют около четверти разницы в интеллекте, было бы, вероятно, разумным
230
Ричард Линн
распространить на различия всех рас. Остающаяся часть различий может быть отнесена на счёт неодинаковой среды обитания и различий в нейрофизиологических процессах.
Половые различия в интеллекте и размере мозга
Проблема, поднимаемая иногда в связи с существованием расовых различий в размере мозга и интеллекте, состоит в том, что мозг женщин значимо меньше мозга мужчин, а до сих пор почти повсеместно утверждалось, что различий в интеллекте между мужчинами и женщинами нет. Например: «Мозг женщин на 10% меньше, чем у мужчин, но их IQ в среднем такой же» (Butterworth 1999, стр. 293). Так как женщины с их меньшим средним размером мозга столь же интеллектуальны, как и мужчины, представляется, что размер мозга не влияет на интеллект. Этот вывод сделан Гулдом (Gould, 1996, стр. 132), который пишет, что это опровергает «миф, что групповые различия в размере мозга имеют какое-либо отношение к интеллекту». То, что средняя величина мозга у женщин меньше, было показано Анкни (Ankney, 1992) и Раштоном (Rushton, 1992). Анкни подсчитал, что средний мозг мужчин (с поправкой на больший размер тела) на 100 граммов тяжелее мозга женщин. Раштон вычислил на материале другого набора данных для 6.325 военнослужащих, что средний мозг мужчины (с поправкой на больший размер тела) имеет объём 1.442 см3, а средний мозг женщины — 1.332 см3, преимущество мужчин составляет 110 см3. Учитывая, что 1 см3 ткани мозга весит приблизительно 1 грамм, результаты Анкни и Раштона весьма сходны.
Таким образом, налицо парадокс, заключающийся в том, что размер мозга положительно связан с интеллектом, у мужчин средний размер мозга больше, чем у женщин, и все же, мужчины и женщины имеют равный интеллект. Я предложил разрешение этого парадокса в следующих работах — Lynn, 1994b, 1999; Lynn, Irwing, 2004. Оно в том, что до возраста 15 лет мужчины и женщины имеют приблизительно одинаковый интеллект, за исключением небольшого преимущества мужчин в способности к визуализации, но с 16-летнего возраста у мужчин начинает проявляться более высокий интеллект, достигающий преимущества от 3 до 6 баллов IQ у взрослых. Интеллект может быть определен четырьмя путями и в случае трёх из них наличие у мужчин более высоких средних показателей IQ вполне очевидно. Во-первых, интеллект можно определить как IQ тестов Векслера по всей шкале, который представляет собой среднее всех основных способностей, включая вербальное понимание, вербальное и невербальное рассуждение, визуализацию, перцепцтивные способности, непосредствен
Расовые различия в интеллекте
231
ную память и скорость восприятия. По данным пяти выборок стандартизации тестов среднее преимущество взрослых мужчин составляет 3,5 балла IQ (Lynn, 1994b, 1997, 1999). В более свежем исследовании Колома и соавт. (Colom et al., 2002) на данных испанской выборки стандартизации теста WAIS-III было выявлено почти такое же преимущество мужчин в 3,6 балла IQ. Среднее преимущество мужчин в 3,5 балла IQ среди взрослых по полномасштабному IQ тестов Векслера, полученное на данных шести выборок стандартизации, недооценивает истинное преимущество мужчин по двум причинам. Во-первых, тесты смещены в пользу женщин, так как вербальные способности, в которых преимущество мужчин сравнительно невелико, представлены в них чрезмерно (шестью субтестами), в то время как способность к визуализации, в которой преимущество мужчин больше, представлена недостаточно (только двумя субтестами: складывания кубиков и лабиринты). Во-вторых, при составлении тестов был изъят ряд пунктов, дающих преимущество мужчинам или женщинам, что должно вести к получению равных оценок IQ мужчинами и женщинами (Matarazo, 1972).
Второй подход к проблеме состоит в определении интеллекта как способности к невербальному рассуждению, как она измеряется Прогрессивными матрицами. Это определение принято Макинтошем (Mackintosh, 1996), и этот тест по широко распространённому мнению — один из лучших тестов фактора g(общего интеллекта). Корт (Court, 1983), Макинтош (Mackintosh, 1996) и Дженсен (Jensen, 1998) утверждали, что различия между средними оценками мужчин и женщин по Прогрессивным матрицам нет и, следовательно, нет различия между мужчинами и женщинами в способности к рассуждению или в g.Вопреки этим утверждениям, мета-анализ всех четырнадцати известных исследований у взрослых показал, что по Прогрессивным матрицам мужчины неизменно получают более высокий IQ, чем женщины, среднее различие составляет 5 баллов (Lynn, Irwing, 2004).
Третий подход к проблеме состоит в определении интеллекта как среднего из невербального рассуждения, вербального понимания и способности к визуализации, как это было предложено Густафсоном (Gustafsson, 1984). Собранные в тринадцати странах данные показывают, что при таком определении у взрослых средний IQ мужчин превышает IQ женщин на 4,9 балла (Lynn, 1999, стр. 4).
Наиболее удовлетворительными представляются следующие подходы определения интеллекта: (1) последовать за Макинтошем (Mackintosh, 1996) и определить интеллект как способность к невербальному рассуждению, или (2) определить его как среднее IQ невербального рассуждения, вербального понимания и способности к визуализации. Эти два определения дают сходные результаты. При первом определении преимущество взрослых мужчин равно 5 баллам IQ (Lynn, Irwing, 2004), а при втором 4,9 баллам (Lynn, 1999).
232
Ричард Линн
Четвёртый подход состоит в определении интеллекта как фактора g, измеренного батареей тестов, оценивающих все или большинство главных первичных способностей. Это определение было принято Дженсеном (Jensen, 1998, стр. 538). Он представляет результаты пяти исследований. Они таковы: IQ мужчин выше на 5,49 баллов по ASVAB (Батарея тестов на пригодность к военной службе для 18-23-летних), на 0,18 балла в американской выборке стандартизации теста WAIS (25-34-летние) и на 2,83 балла в американской выборке стандартизации теста WISC-R (5-16-летние), в то время как женщины получили более высокую оценку IQ на 0,03 балла по BAS (Британские шкалы способностей 14—17-летних) и на 7,91 балл по GATB (Батарея тестов общих способностей для 18-летних). На основании этих противоречивых результатов он заключает: «Половое различие в психометрическом g или вообще не существует, или же оно незначительно и его направленность неясна» (Jensen, 1998, стр. 340). Вывод Дженсена не решает проблему, состоящую в том, что у мужчин мозг в среднем больше, чем у женщин, и однако же, согласно его анализу, они имеют одинаковый средний IQ. Дженсен полагает, что решение этой проблемы может состоять в том, что мужчины и женщины имеют одинаковое число нейронов, но у женщин они более плотно «упакованы» в меньшем мозге (стр. 541). Исходя из фундаментальных биологических принципов, представляется невероятным, что половое различие такого рода могло развиться, и это было опровергнуто Пакенбергом и Гюндерсоном (Packenberg, Gunderson, 1997), определившими, что у мужчин больше нейронов, чем у женщин (22,8 миллиарда против 19,3 миллиардов), а различия в плотности нейронов в мозге мужчин и женщин нет.
В связи с заключением Дженсена об отсутствии половых различий в интеллекте возникает два вопроса. Во-первых, он не принимает во внимание свидетельства, представленные в моей статье 1994 г. (Lynn, 1994b) и подтверждённые последующими работами (Colom, Lynn, 2004; Lynn, Irwing, 2004), о том, что преимущество мужчин отсутствует или минимально до возраста 16 лет, поэтому результаты теста WISC-R у 5-16-летних и Британских шкал способностей 14-17-летних учитывать не следует. Во-вторых, несоответствие между преимуществом мужчин в 5,49 балла IQ по тесту ASVAB и преимуществом женщин в 7,91 балла по GATB настолько велико, что это говорит о каких-то методических погрешностях. Проблема заключается в том, что природа g,оцененного с помощью батареи тестов, зависит от типа тестов в батарее. Преимущественно вербальный тест, такой как тест Векслера, даёт вербальный g,по которому преимущество взрослых мужчин весьма невелико. Батарея GATB, по которой преимущество женщин в g составило 7,9 балла IQ, содержит ряд перцептивных и психомоторных тестов и, следовательно, даёт перцептивно-психомоторный g.Женщины выполняют эти тесты лучше мужчин и, таким образом,
Расовые различия в интеллекте
233
по верному наблюдению Ниборга (Nyborg, 2003, стр. 206), имеют более высокий g. Если изъять эти тесты и проанализировать данные тестов вербальных, числовых и пространственных способностей, преимущество женщин исчезает, как это было показано Дженсеном (Jensen, 1998, стр. 543). Более свежие исследования, использующие этот метод, вновь дали противоречивые результаты. Колом и соавторы, проанализировав с помощью того же метода большую испанскую выборку (Colom et al., 2000) и испанскую выборку стандартизации теста WAIS-III (Colom et al., 2002), нашли, что половые различия в g были незначительны. Однако, использовавший тот же метод Ниборг (Nyborg, 2003, стр. 209) выявил преимущество взрослых мужчин в 5,55 баллов IQ, а Колом и Линн (Colom, Lynn, 2004) при анализе испанской выборки стандартизации теста DAT выявили среди 18-летних преимущество мужчин в 4,3 балла IQ. Очевидно, что половые различия, полученное этим методом, очень вариабельны. Причина этого в том, что различные тесты дают разные g.
Более высокий IQ мужчин может быть приписан большему мозгу. В трёх исследованиях было показано, что средний размер мозга мужчин превышает размер мозга женщин (с поправкой на размер тела). Анкни выявил (при измерении в весовых единицах), что мозг мужчин больше приблизительно на 100 граммов (Ankney, 1992). Раштон (Rushton, 1992) обнаружил различие в объеме, равное 110 см3, а Тэн и соавт. (Tan et al., 1999) выявили различие среди студентов колледжа в Турции в 91см3. Анкни выразил различие между мужчинами и женщинами в 0,78d (единицы стандартного отклонения). Корреляция размера мозга с интеллектом составляет 0,44 (с поправкой на надежность теста). Отсюда, предсказанное преимущество мужчин в интеллекте вследствие большего среднего размера мозга будет равно величине 0,78, умноженной на 0,44, что составляет 0,34с?или 5,1 балла IQ. Это, в пределах диапазона ошибки, возникающей вследствие использования различных тестов, ошибок измерения и различных определений интеллекта, следует считать тем же самым значением, что и фактическое мужское преимущество в 4,9 балла (Lynn, 1999), в 5,0 баллов (Lynn, Irwing, 2004), в 5,49 баллов (по ASVAB), в 5,55 балла (в датской выборке — Nyborg, 2003, стр. 212) и в 4,3 балла (в испанской выборке — Colom, Lynn, 2004). Среднее значение четырёх оценок равно 5,0 баллам IQ, и это должно быть принято как наилучшая оценка преимущества взрослых мужчин. Это преимущество полностью объясняется большим размером мозга мужчин, как и можно было бы ожидать, потому что мужчины и женщины подвергаются одинаковым средовым воздействиям, вследствие чего различия между ними не могут быть объяснены средовыми факторами. С эволюционной точки зрения, объяснение того, что средний интеллект у мужчин выше, состоит, вероятно, в том, что у большинства социальных видов животных самцы конкурируют друг с другом за самок, и в
234
Ричард Линн
эволюции гоминидов интеллект стал играть существенную роль при достижении успеха в этом соревновании. Самки не конкурируют за самцов в сколь-нибудь подобной степени. Это должно было оказывать более сильное давление отбора в сторону увеличения интеллекта у мужчин, чем у женщин.
Генетические процессы в эволюции расовых различий в IQ
Следует полагать, что за развитие расовых различий в интеллекте ответственны два генетических процесса. Первый из них состоит в том, что между расами развились следующие различия в частотах аллелей высокого и низкого интеллекта: аллели высокого интеллекта больше распространены в расах с более высоким IQ и реже встречаются в расах с более низким IQ. Ранние люди, мигрировавшие из Африки и распространившиеся по всему миру, должны были нести аллели как высокого, так и низкого интеллекта, но те, кто колонизировал Азию и Европу, столкнулись с задачей выживания в течение холодной зимы, предъявлявшей повышенные требования к когнитивной сфере. Многие из несущих аллели низкого интеллекта должны были оказаться неспособными выжить в течение холодных зим и менее интеллектуальные индивиды и племена должны были вымирать, а выживали более интеллектуальные. Этот процесс должен был уменьшать частоту и, возможно, устранять аллели низкого интеллекта, увеличивая долю аллелей высокого интеллекта. Чем ниже зимние температуры, тем сильнее давление отбора в сторону элиминации индивидов с низким IQ, несущих аллели низкого интеллекта. Этот процесс объясняет некоторую связь между наиболее холодными зимними температурами, IQ и размером мозга, представленную в Таблице 16.2.
Параллельный генетический процесс должен был действовать в эволюции расовых различий в цвете кожи. Первые люди, эволюционировавшие в тропической Африке, как и народы, живущие там сегодня, должны были иметь чёрную или очень тёмную кожу в силу адаптивного преимущества тёмной кожи при ярком солнечном свете. Когда некоторые из этих ранних народов мигрировали в Северную Африку, Азию и Европу, у них вследствие мутаций должны были появиться аллели, обеспечивающие развитие более светлой кожи. Индивиды с такими мутациями должны были иметь преимущество в плане отбора, поскольку могли синтезировать витамин D под действием солнечного света, и в то же время они не страдали от заболевания раком кожи из-за чрезмерно яркого солнечного света в тропиках. В результате люди с более светлой кожей оставляли больше
Расовые различия в интеллекте
235
выживших потомков, так что в итоге аллели более светлого цвета кожи распространились в популяции и в конце концов заменили аллели тёмной кожи. Этот процесс произвел такой же широкий градиент цвета кожи, какой образовался в случае интеллекта. У арктических народов, аборигенов Восточной Азии и европейцев развилась наиболее бледная кожа, у аборигенов Южной Азии и Северной Африки, американских индейцев, аборигенов Юго-Восточной Азии и у аборигенов островов Тихого океана — менее светлая кожа, в то время как африканцы, бушмены и австралийские аборигены, подвергающиеся действию самого сильного солнечного излучения, сохранили самую тёмную кожу.
Второй генетический процесс, наличие которого было предположено Миллером (Miller, 1996, 2005), состоит в появлении нескольких новых аллелей высокого интеллекта вследствие мутаций у некоторых рас, эти аллели не появлялись у других рас и никогда не передавались каким-либо другим расам. Это предположение необходимо, чтобы объяснить некоторые из аномалий в общей связи между суровостью зимы и расовыми различиями в интеллекте. Общие правила заключаются в том, что новые мутантные аллели высокого интеллекта с наибольшей вероятностью могут появиться в больших популяциях и в популяциях, подвергающихся воздействию стресса. Наибольшая вероятность появления этих мутантных аллелей в больших популяциях объясняется тем, что мутация — случайное генетическое событие и, следовательно, оно более вероятно произойдет в многочисленных расах. Кроме того, в популяциях, подвергающихся стрессу, включая экстремальные температуры, также появляется больше мутаций (Plomin, DeFries, McCleam, 1990, стр. 91). Действия этих двух правил имело результатом то, что мутации более высокого интеллекта с наибольшей вероятностью должны были появиться и, как можно полагать, появлялись чаще у аборигенов Южной Азии, живших большими популяциями и подвергавшихся холодовому стрессу, и особенно у многочисленных аборигенов Восточной Азии и европейцев, подвергавшихся предельному холодовому стрессу, чем у африканцев, живших большими популяциями, но не подвергавшихся экстремальному холодовому стрессу, а также у малочисленных австралийских аборигенов и бушменов, не подвергавшихся экстремальному холодовому стрессу. Арктические народы подвергались экстремальному холодовому стрессу, но их популяции были очень небольшими, так что появление у них мутаций более высокого интеллекта было маловероятным. Возможно также, что при возникновении новых мутантных аллелей высокого интеллекта у аборигенов Южной Азии, и особенно у аборигенов Восточной Азии и европейцев, действовал также механизм «направленной мутации». Концепция «направленной мутации» заключается в том, что мутация произойдет с большей вероятностью, если это будет выгодно для организма. Эта теория первоначально была выдвинута
236
Ричард Линн
Кернсом, Овербахом и Миллером (Cairns, Overbaugh, Miller, 1988) и затем была поддержана рядом биологов (Lenski, Mittler, 1993). Более высокий интеллект был бы более полезен для аборигенов Южной Азии и, в особенности, для аборигенов Восточной Азии и европейцев, чем для африканцев.
Однажды появившись, новая мутантная аллель более высокого интеллекта будет давать преимущество перед отбором и распространится в группе из приблизительно 50-80 человек, какими люди жили на эволюционной стадии охотников-собирателей. Затем она должна распространиться довольно быстро в соседние группы, потому что охотники-собиратели обычно образуют союзы с соседними группами и обмениваются с ними брачными партнёрами, так что разумно предположить, что этот обычай существовал в течение многих тысячелетий эволюции рас. Эти союзы групп известны как демы. Новая мутантная аллель более высокого интеллекта, дававшая преимущество при отборе, должна была распространиться в деме достаточно быстро. Время от времени браки должны были происходить между демами, и таким путём новые мутантные аллели более высокого интеллекта распространялись от одного дема к другому и в конце концов по всей расе. Как было оценено Рухани (Rouhani, 1989), принявшим разумные допущения, что коэффициент отбора равен 0,01 и миграция между демами охотников-собирателей приблизительно в 500 человек за поколение составляет 5 процентов, обеспечивающие преимущество аллели должны были распространяться со скоростью 0,8 миль за поколение. Таким образом, через 25.000 лет, что составляет приблизительно 1.000 поколений, дающая преимущество аллель должна будет распространиться примерно на 800 миль. Таким образом, выгодная аллель, появившаяся вследствие мутации в области, скажем, Пекина 25.000 лет назад, ещё не должна была распространиться за пределы Китая и потребовалось бы ещё 50.000 лет или около того, чтобы она достигла арктических народов крайнего Северо-Востока Азии. Эта модель, однако, не принимает во внимание географические барьеры, обычно существовавшие между расами, такие как пустыня Гоби между аборигенами Восточной Азии и европейцами, и Сахара между африканцами районов Африки южнее Сахары и аборигенами Северной Африки, которые в значительной степени предотвращали ин- тербридинг между демами разных рас и, следовательно, передачу новых мутантных аллелей более высокого интеллекта от одной расы к другой.
Г Ж 17 МСОШХ РАЗЛИЧНА й НЛШМШ
Африканцы
Бушмены
Аборигены Южной Азии и Северной Африки
Аборигены Юго-Восточной Азии
Аборигены островов Тихого океана
Австралийские аборигены
Европейцы
Аборигены Восточной Азии
Арктические народы
Американские индейцы
Заключение
Теперь, когда общие принципы эволюции интеллекта и решающая роль климатических факторов в развитии расовых различий в интеллекте и размере мозга были изложены в Главах 15 и 16, мы в состоянии реконструировать, как и когда развились особенности интеллекта у каждой расы. Начнём с Homo erectus,живших в экваториальной Африке приблизительно от 1,7 миллионов до 200.000 лет назад. За этот период их мозг увеличился примерно с 885 см3 до 1.186 см3(Ruff, Trinkaus, Holliday, 1997). Причина этого увеличения мозга в том, что у всех млекопитающих интеллект находился под постоянным давлением отбора, т.е. более интеллектуальные индивиды оставляли больше выжившего потомства, а у эволюционирующих гоминидов этот процесс ускорился. В конце этого периода появился Homo sapiens— Человек разумный(Relethford, 1988), качество орудий которого даёт основание предположить, что эти люди находились на стадии конкретных операций по Пиаже, достигаемой современными европейскими 7-8-летними детьми, что указывает на то, что их IQ равнялся примерно 50 баллам (Глава 15, Параграф 6).
238
Ричард Линн
Африканцы
В последние 200.000 лет предки африканцев продолжали обитать в тропической и субтропической зонах Африки южнее Сахары. Окружающая среда не предъявляла высоких требований к их когнитивной сфере, поскольку приматы адаптировались к ней приблизительно за 60 миллионов лет. В период своей эволюции Homo erectusпитались в основном растительной пищей, но пополняли свой рацион, подбирая остатки туш животных, задранных львами, леопардами и гепардами (Lee, 1968; Tooby, de Vore, 1989). Эволюционировавшие африканцы жили, как и нынешние охотники-собиратели, в тропической и субтропической среде, питаясь преимущественно растительной пищей, многочисленные виды которой доступны в течение всего года, а также насекомыми и яйцами, лишь время от времени потребляя мясо животных, добываемое охотой.
Наличие под руками в течение всего года растительной пищи, насекомых и яиц означало, что африканские народы, эволюционировавшие в тропической и субтропической Африке, не были вынуждены охотиться на животных для получения мяса. В 1966 году была созвана конференция антропологов для обсуждения темы «Человек-охотник» о значении охоты для современных охотников-собирателей, на которой «единым мнением было, что в рационе современных тропических собирателей мясо имеет сравнительно небольшое пищевое значение» (Stanford, Bunn, 2001, стр. 4). В 1999 г. проводилась подобная конференция, на которой было «достигнуто согласие в том, что основу рациона гоминидов составляла в первую очередь растительная пища, как и у современных собирателей в тропиках» (Stanford, Bunn, 2001, стр. 356). Поэтому африканцам не было никакой нужды развивать интеллект, навыки, инструменты и оружие, необходимые для охоты на крупных млекопитающих. Кроме того, температура в экваториальной Африке меняется в течение года приблизительно от 32°С в самом жарком месяце до 17°С в самом холодном, так что африканские народы не сталкивались с предъявляющими требования к когнитивной сфере задачами изготовления игл и нитей для изготовления одежды и шатров, добычи огня и его поддержания или приготовления пищи и её сохранения для будущего потребления. Им было сравнительно легко выхаживать новорожденных, младенцев и малолетних детей, так как отсутствовала необходимость обеспечивать их одеждой, а с очень раннего возраста дети были способны ходить на поиски пищи самостоятельно.
Тем не менее, мозг африканцев увеличился за последние 200.000 лет или около того приблизительно с 1.186см3 до 1.276см3 и можно резонно предположить, что это повлекло за собой увеличение их интеллекта до его
Расовые различия в интеллекте
239
современной величины в 67 баллов. Это увеличение произошло из-за постоянного направленного отбора по интеллекту, то есть более интеллектуальные индивиды имели больше выживших потомков. Генетические процессы должны были состоять в увеличения частот аллелей более высокого IQ и, вероятно, в некотором количестве мутаций, обеспечивающих повышение интеллекта. Если мутации, обеспечивающие более высокий интеллект, появлялись, то они должны были распространяться в популяции, потому что высокий интеллект — приспособительная характеристика, но они должны были распространяться не так быстро и широко, как в расах, живущих в умеренном и холодном климате, потому что давление отбора в сторону повышения интеллекта было не столь сильным в мягком климате экваториальной Африки.
Уровень интеллекта, развившийся у африканцев, был достаточен для достижения небольшого прогресса в переходе от охотников-собирателей к оседлому сельскому хозяйству, но не достаточен для создания чего- либо, что можно было бы назвать цивилизацией с письменностью и арифметикой, календарём, городами с капитальными каменными зданиями и удовлетворяющего другим критериям, сформулированным Бейкером (Baker, 1974).
Бушмены
По-видимому, приблизительно 100.000 лет назад некоторые группы архаичных африканцев начали мигрировать к Югу, где они эволюционировали в бушменов, заселивших большую часть Южной Африки, но к нашему времени лишь несколько десятков тысяч бушменов выжило в пустыне Калахари. В течение последних 100.000 лет мозг бушменов увеличился приблизительно на десять процентов до сегодняшнего размера в 1.270 см3, а их IQ увеличился до 54 баллов. Климат в Южной Африке умеренно тёплый с немного более прохладными, чем в экваториальной Африке, зимами. Тем не менее бушмены были в состоянии выжить, питаясь в основном растительной пищей, насекомыми и яйцами, как они делают это сегодня. Как сообщал Стал (Stahl, 1984), бушмены потребляют приблизительно 90 видов растительной пищи, что составляет 70-85 процентов их рациона. Следовательно, перед их когнитивной сферой не вставала задача выживания в холодном и умеренном климате. Однако, исходя исключительно из климатической теории развития расовых различий в интеллекте, у бушменов должен был развиться более высокий интеллект, чем у африканцев. Этого не произошло, и IQ бушменов сегодня ниже, чем у африканцев (54 и 67 баллов, соответственно). Объяснение этого, вероятно, в том, что некоторые мутации более высокого интеллекта появились у африканцев в силу
240
Ричард Линн
их многочисленности, но не появились у бушменов вследствие их меньшей численности. Однако, мозг у бушменов лишь немного меньше, чем у африканцев (примерно 1.270 см3 против 1.276 см3). Это указывает на то, что мутантные аллели более высокого IQ, которые вероятно появились у африканцев и распространились в их популяции, скорее обуславливали неврологические процессы, а не увеличение размера мозга.
Аборигены Южной Азии и Северной Африки
Первые группы людей, мигрировавших из районов Африки южнее Сахары, колонизировали Северную Африку и Юго-Западную Азию в период примерно от 100.000 до 90.000 лет назад. В период приблизительно 90.000-60.000 лет назад они колонизировали всю Южную Азию. Здесь они были изолированы от африканцев расстоянием и пустыней Сахара и эволюционировали в аборигенов Южной Азии и Северной Африки. Первоначально они столкнулись с подобным сегодняшнему умеренным климатом, с температурой самого холодного зимнего месяца около 13°С.Примерно 70.000 лет назад в Северном полушарии начались ледниковые периоды, продлившиеся приблизительно 20.000 лет. Затем последовал более тёплый период (приблизительно от 50.000 до 28.000 лет назад), а после него второй, более суровый ледниковый период (главное Вюрмское оледенение), начавшийся приблизительно 28.000 лет назад и продлившийся примерно до 10.000 лет назад, после чего температуры повысились достаточно быстро до соответствующих сегодняшнему мягкому климату (Roberts, 1989; Foley, 1987). Во время главного Вюрмского оледенения зимние температуры в Северной Африке, Евразии и Северной Америке снизились примерно на 5°С. (Roberts, 1989) и температура самого холодного зимнего месяца в Северной Африке и Южной Азии снизилась приблизительно до 7°С.
Выживание во время ледниковых периодов в холодном и умеренном климате Северной Африки и Южной Азии и позже в субарктическом климате Европы и Северной Азии ставило перед людьми ряд требующих когнитивных усилий задач, которые должны были действовать как давление отбора в сторону увеличения интеллекта выше уровня, требовавшегося в тропическом и субтропическом климате Африки южнее Сахары. Основных проблем должно было быть пять. Во-первых: растительная пища была не доступна в течение зимы и весны, её не было в изобилии даже летом и осенью; насекомые и рептилии также не были доступны, так как в умеренном климате они впадают в спячку. Поэтому главным источником пищи стали крупные млекопитающие, такие как антилопы, олени, лошади и кабаны, на которых люди вынуждены были охотиться, чтобы обеспечить
Расовые различия в интеллекте
241
себе запасы продовольствия. Должно было быть нелегко охотиться на этих крупных млекопитающих на лугах, покрывавших большую часть Северного полушария во время последнего ледникового периода, потому что на них хорошая видимость на несколько тысяч ярдов, а у травоядных животных существует хорошая система предупреждения о приближающихся хищниках. Охотиться на открытых лугах сложнее, чем в лесах тропиков и субтропиков, где имеется масса укрытий для охотников. Люди, эволюционировавшие в экваториальной Африке, питались с основном растительной пищей и были не приспособлены к охоте на крупных млекопитающих, так что это должно было поставить перед ними новые когнитивные задачи. Крупные травоядные животные могут быстро бегать и их практически невозможно поймать с помощью простого преследования. Единственный способ добыть этих животных состоял в использовании естественных ловушек, куда животных можно было загнать и затем убить. Одной из наиболее часто используемых естественных ловушек были узкие ущелья, через которые можно гнать животных, которые могут там споткнуться и быть поражены копьями или дубинами поджидающих в засаде охотников. Другой были утёсы, к которым группа охотников могла подогнать стадо травоядных животных так, чтобы некоторые из них упали с края и разбились или были ранены достаточно тяжело для того, чтобы другие охотники могли добить их. Естественными ловушками были также болота и петли рек, в которые группа охотников могла загнать травоядных животных и затем их убить. Археологические раскопки показали, что все эти ловушки использовались ранними людьми в Евразии (Geist, 1978; Mellars, 1999). Выработка стратегии групповой охоты и заманивания в ловушку крупных травоядных животных описанными способами требовали развития когнитивных способностей.
Было показано, что у современных охотников-собирателей доля продуктов, получаемых охотой и собирательством, изменяется с географической широтой. Народы тропических и субтропических широт преимущественно собиратели, в то время как народы умеренных широт больше полагаются на охоту, а народы арктической и субарктической зон обеспечивают себя почти исключительно охотой и рыболовством, они вынуждены делать это из-за недоступности растительных продуктов, за исключением ягод и орехов летом и осенью (Lee, 1968). Когда люди мигрировали в области Северной Африки и Южной Азии, где был умеренный климат, многие из обладавших низким IQ не смогли выжить холодными зимами и это повысило IQ оставшихся в живых до 84 баллов.
Во-вторых: для успешной охоты на крупных травоядных животных требовалось изготавливать разнообразные изделия из камня, дерева и кости для того, чтобы сделать копья и орудия для разделки туш. Некоторых из этих животных можно было поразить копьями, которые нужно было
242
Ричард Линн
сделать, вставив в конец древка или привязав к нему предварительно заготовленный острый кусок камня. Когда эти охотники заваливали и убивали крупное травоядное животное, они должны были содрать с него шкуру и разделать тушу на части такого размера, чтобы их можно было перенести на стоянку для женщин и детей. У этих животных толстые шкуры и жёсткие связки, которые трудно разрезать, и людям требовались острые орудия, специально изготовленные для этих целей. В субарктическом климате убитые животные довольно быстро замерзают и тогда разрезать их становится невозможно, так что охотники должны были иметь хорошие режущие орудия для быстрого завершения работы до того, как туша затвердеет на морозе.
В холодном климате людям нужно больше разнообразных и более сложных орудий, чем в тропическом и субтропическом климате. Это было показано Торренсом, продемонстрировавшим связь между географической широтой и числом и сложностью орудий, используемых современными охотниками-собирателями (Torrence, 1983). Он обнаружил, что охотники-собиратели в тропических и субтропических широтах, например в бассейне Амазонки и на Новой Гвинее, обычно имеют от 10 до 20 разных орудий, тогда как народы в более холодных северных широтах (в Сибири, на Аляске и в Гренландии) имеют от 25 до 60 разных орудий. Кроме того, народы холодных северных широт изготавливают более сложные орудия, требующие сборки составных частей, такой как вставка острого куска камня или кости в древко копья или закрепление головки каменного топора на деревянной рукоятке.
В-третьих: ещё один ряд проблем, встававших перед людьми в Северном полушарии, был связан с сохранением тепла. Люди были вынуждены решать задачи добывания огня и устройства укрытий. Археологические раскопки показали, что во время ледниковых периодов люди в Китае и в Европе добывали огонь. Для этого они должны были научиться получать искры, ударяя один камень о другой, и зажигать этими искрами высушенную траву. Им был нужен запас сухой травы, сухой древесины и помёта животных, хранящийся в пещерах, чтобы зажечь огонь и затем поддерживать его горение. Это требовало интеллекта и предварительного планирования. У народов в Африке южнее Сахары и в Австралии также был огонь, но в тропиках и субтропиках им было легче его получать, потому что там возникали самопроизвольные лесные пожары, где можно было брать горящие головни и приносить их на стоянку для разведения костра. Разжигание огня и его поддержание были значительно более трудны в Евразии и Северной Америке, чем в тропическом и субтропическом Южном полушарии.
В-четвертых: другой проблемой, связанной с сохранением тепла, являлась необходимость изготовления одежды и шатров с помощью сшивания кусков шкур животных. Для этого требовалось высушивать и выде
Расовые различия в интеллекте
243
лывать шкуры крупных травоядных животных и изготавливать костяные иглы и нити, чтобы сшивать шкуры при изготовлении одежды и обуви. Нужно было делать одежду для младенцев и детей, так же как и для взрослых. Некоторые люди согревались, живя в пещерах, но там, где пещер не было, они использовали большие кости и сшитые вместе шкуры для выделки шатров, напоминающих юрты, до сих пор изготавливаемые в Монголии (Geist, 1978; Mellars et al., 1999).
В-пятых: наконец, проблемой для народов в умеренном и холодном климате являлось сохранение пищи. Это было необходимо, потому что, когда они убивали и расчленяли несколько крупных млекопитающих, они были не в состоянии съесть их всех в течение нескольких дней и должны были сохранить их для будущего потребления.
Некоторые животные, на которых можно охотиться, мигрируют и появляются в течение года в определённой местности только на короткие периоды времени. Это даёт возможность добыть их в большом количестве, слишком большом для немедленного потребления, но их можно сохранить впрок. Один из примеров — северные олени, которые регулярно мигрируют на длинные расстояния в определенное время года. Во многих случаях они следуют одними и теми же маршрутами в одно и то же время года, так что их появление могло быть предсказано ранними людьми, которые приобрели знание о сезонах и календаре посредством астрономических наблюдений. Другой мигрирующий вид — лосось, который мигрирует в больших количествах в определенное время года из моря в реки на нерест. Многие из этих рек весьма мелководны, и не слишком трудно добыть копьём большое количество лосося, плывущего вверх по течению. Также возможно поймать его сетями, изготовление которых было другой требующей когнитивных усилий задачей для народов Евразии. Эти народы были в состоянии предвидеть появление мигрирующих стад и рыбы и добывать их в больших количествах во время прохождения.
В очень холодном климате проблема хранения пищи для будущего потребления в течение части года могла быть решена путём постройки ледников для хранения туш. Другим решением было нарезать мясо на тонкие ломтики и высушить их. Если это сделано должным образом, то ломтики останутся съедобными в течение долгого времени, но если нет — они станут токсичными. Некоторые из менее интеллектуальных индивидов, неспособных сделать это правильно, должны были умирать от пищевого отравления. Это было ещё одним из многих видов давления отбора, действовавшего в сторону увеличения интеллекта народов, колонизировавших нишу умеренного и холодного климата. Как предположил Миллер (Miller, 1991), хранение пищи также требовало выработки правил нормирования её потребления и это должно было приводить к развитию счёта, чтобы распределять пищу справедливо.
244
Ричард Линн
Бинфорд показал (Binford, 1980,1985), что у современных охотников- собирателей имеется связь между тем, в какой мере они прибегают к хранению пищи и температурой среды их обитания. Чем холоднее среда обитания, тем больше пищи они запасают для будущего потребления. Он сообщает, что в целом только в более холодном климате, где сезон вегетации короче примерно 200 дней, охотники-собиратели хранят пищу.
В дополнение к этим пяти когнитивным проблемам выживания в Северном полушарии ещё одним фактором давления отбора в сторону увеличения интеллекта этих народов должно было быть действие полового отбора со стороны женщин. В Евразии и Северной Америке женщины должны были стать полностью зависимыми от мужчин, на протяжении большей части года обеспечивавших их и их детей пищей. В экваториальной Африке и в Южном полушарии, где растительная пища и насекомые доступны в течение всего года, женщины сравнительно независимы от мужчин. Даже женщины с младенцами и малолетними детьми могут взять их с собой на поиски продовольствия или оставить на попечение других женщин на несколько часов, пока они уходят собирать растительную пищу. В Северном полушарии женщинам с младенцами и маленькими детьми было бы труднее, а часто невозможно отправиться на охоту, длившуюся, вероятно, несколько дней, убивать и расчленять крупных млекопитающих и нести части их туш много миль обратно на стоянку. Это приводило к тому, что женщины в Северном полушарии должны были стать зависимыми от мужчин, снабжавших их пищей. Женщины, следовательно, предпочитали выбирать в мужья интеллектуальных мужчин, которые были хорошими охотниками и умели изготавливать хорошие инструменты и оружие. Результатом такого полового отбора со стороны женщин должно было быть то, что у умных мужчин было больше детей, и это должно было повышать групповой интеллект. Другим эффектом большей зависимости женщин от мужчин в Евразии было то, что мужчины и женщины стали крепче связаны психологически. Это объясняет, почему браки и внебрачные отношения у европейских и Восточно-азиатских народов более устойчивы, чем у африканцев (Lynn, 2002).
Выживание в холодном климате Северного полушария должно было потребовать увеличения общего интеллекта, определяемого как способность к решению общих проблем и к обучению, и большинства первичных когнитивных способностей, из которых состоит общий интеллект. Более высокая способность к рассуждению должна была потребоваться для решения всех, встающих в холодных северных широтах новых задач, таких как построение убежищ и добывание огня, изготовление одежды и производство более эффективных орудий для охоты, забоя и разделки туш крупных животных. Улучшенная вербальная способность должна была быть необходима для лучшего общения при обсуждении того, как решать
Расовые различия в интеллекте
245
эти задачи, для планирования будущих действий и для передачи приобретённых культурных знаний и навыков детям. Повысившаяся способность к визуализации была необходима для выработки плана групповых действий на охоте и выполнения его, для точного метания копий и другого метательного оружия и для изготовления более сложных инструментов и оружия из камня, кости и дерева. Отцы должны были показывать сыновьям, как откалывать кремень, чтобы сделать хорошие режущие инструменты и копья с острыми наконечниками, а эти навыки передаются в значительной степени путём наблюдения и имитации, подобно тому, как навыки ручных ремёсел изучаются сегодня учениками в большей степени путём наблюдения за квалифицированными мастерами, чем посредством устных объяснений. Охотой и изготовлением инструментов занимались преимущественно мужчины, и именно поэтому практически всегда обнаруживается, что способность к визуализации у мужчин выше, чем у женщин (Linn, Peterson, 1986). Давление отбора на народы Северного полушария в сторону развития лучшей кратковременной памяти и скорости восприятия должно было быть слабее, что объясняет, почему эти способности не возросли в такой степени у европейцев по сравнению с африканцами.
Давление отбора в сторону увеличения интеллекта в умеренном климате Северной Африки и Южной Азии и позже в субарктическом климате Европы и Северной Азии должно было действовать и на мужчин и на женщин. Давлением отбора в сторону увеличения интеллекта на мужчин должна была быть необходимость ходить на охоту на крупных млекопитающих и изготавливать орудия, требующиеся для этого, и для сдирания шкур и разделки туш. Это должно было требовать увеличения пространственного интеллекта и способности к рассуждению, которые в среднем выше у мужчин, чем у женщин (Linn, Petersen, 1986; Lynn, Irwing, 2004). Повышенный общий интеллект был нужен женщинам, чтобы добывать и поддерживать огонь, чтобы заготавливать пищу и сохранять её для будущего потребления; на них лежала ответственность за сохранение жизни младенцев и малолетних детей, которых они должны были держать в тепле. Генетические процессы, протекавшие у аборигенов Северной Африки и Южной Азии, должны были состоять в увеличении частот аллелей более высокого интеллекта и, вероятно, в появлении новых мутаций, обуславливающих более высокий интеллект, и в их распространении в популяции.
Самый вероятный сценарий: интеллект аборигенов Северной Африки и Южной Азии увеличился во время обоих ледниковых периодов, первый из которых длился приблизительно от 70.000 до 50.000 лет назад, а второй примерно от 28.000 до 10.000 лет назад. Об увеличении их интеллекта после завершения первого из этих двух ледниковых периодов можно судить по более сложным орудиям и другим изделиям (Stringer, McKie, 1996, стр. 185-187). Однако их интеллект не возрос до уровня, при котором они
246
Ричард Линн
были бы в состоянии совершить неолитический переход от охотников-собирателей к осёдлому сельскому хозяйству. Это дальнейшее повышение интеллекта должно было произойти во время второго большого ледникового периода (главное Вюрмское оледенение). Суровость климата во время этого периода должна была быть основным давлением отбора, которое увеличило размер мозга аборигенов Северной Африки и Южной Азии до 1.342 см3 и их IQ до 84 баллов. Этого оказалось достаточно, чтобы позволить им совершить неолитический переход к осёдлому сельскому хозяйству и затем построить ранние цивилизации в долинах рек Нил, Тигр, Евфрат и Инд, где они строили города, создавали письменность, арифметику, системы законов и всё, являющееся критериями цивилизации.
Аборигены Юго-Восточной Азии
Некоторые из народов Южной Азии мигрировали в Юго-Восточную Азию приблизительно 70.000 лет назад и эволюционировали в аборигенов Юго-Восточной Азии. Климат этого региона тропический и субтропический с температурой самого холодного зимнего месяца около 24°С. Эти народы достигли указанной области до начала ледниковых периодов, мало её затронувших. Поэтому давление отбора в сторону увеличения интеллекта на эти народы было невелико. Однако, их IQ (87 баллов) немного выше, чем у аборигенов Северной Африки и Южной Азии (84 балла), от которых они по большей части произошли. Наиболее вероятное объяснение этого состоит в том, что аборигены Юго-Восточной Азии имеют некоторую примесь аборигенов Восточной Азии, которые мигрировали к Югу и перемешались с местным населением. Миграция аборигенов Восточной Азии в Юго-Восточную Азию была существенной. Так, сегодня в Сингапуре 76 процентов населения китайцы, в Малайзии 30 процентов населения китайцы, значительные китайские меньшинства имеются также в Камбодже и Таиланде (Philiprs, 1996). Эти аборигены Восточной Азии перемешались с местными народами, породив расово-смешанное население Юго-Восточной Азии. Вследствие этой миграции и интербри- динга народы Юго-Восточной Азии генетически близки южным китайцам (Cavalli-Sforza, Menozzi, Piazza, 1994, стр. 78). Китайская примесь у аборигенов Юго-Восточной Азии дала им некоторые из аллелей высокого интеллекта и подняла их IQ до 87 баллов.
Такой IQ позволил аборигенам Юго-Восточной Азии совершить неолитический переход от охотников-собирателей к осёдлому сельскому хозяйству и затем построить сравнительно внушительные цивилизации в период приблизительно от 0 до 1.000 гг. нашей эры. Эти цивилизации воз
Расовые различия в интеллекте
247
никли несколько позже, чем в Северной Африке и Южной Азии, потому что долины рек в Юго-Восточной Азии были покрыты густыми лесами и не имели пойм, в которых производились излишки сельскохозяйственных продуктов, обеспечивавших первые цивилизации в Месопотамии, Египте и Китае. Однако, примерно с 1.000 г. нашей эры их IQ оказался недостаточным для того, чтобы они могли конкурировать в экономике или в науке и технике с европейцами и аборигенами Восточной Азии.
Аборигены островов Тихого океана
Лишь примерно 6.000 лет назад некоторые аборигены Юго-Восточной Азии начали мигрировать на острова Тихого океана, где они эволюционировали в аборигенов островов Тихого океана. Величина их IQ (85 баллов) не отличается значимо от 87 баллов аборигенов Юго-Восточной Азии, от которых они по большей части произошли, но он также выше, чем можно ожидать, исходя из их жизни в мягком климате с температурой самого холодного зимнего месяца около 24°С. Объяснение этого такое же, как и в случае аборигенов Юго-Восточной Азии, а именно — примесь генов аборигенов Восточной Азии, мигрировавших на Юг и смешивавшихся с местным населением. О присутствии существенной доли восточноазиатской родословной у аборигенов островов Тихого океана говорят их маленькие зубы, которые малы у аборигенов Восточной Азии, но велики у австралийских аборигенов (Brace, Hinton, 1981). В отличие от аборигенов Юго-Восточной Азии, аборигены островов Тихого океана достигли лишь незначительных успехов в неолитическом переходе к оседлому сельскому хозяйству и совсем не преуспели в создании цивилизаций. Объяснение этого состоит в том, что их популяции были очень маленькими, обычно численностью всего в несколько тысяч, и рассеянными по удаленным островам, разделённым огромными расстояниями. Лишь Маори имели большую территорию в Новой Зеландии, но они колонизировали эти острова только примерно в 800 г. нашей эры и у них не было достаточно времени, чтобы развиться в большую популяцию, совершить полный неолитический переход и начать строить цивилизацию.
Австралийские аборигены
Некоторые из народов Южной и Восточной Азии мигрировали на острова Индонезийского архипелага и приблизительно 65.000 лет назад достигли Новой Гвинеи. Примерно 60.000 лет назад некоторые из этих
248
Ричард Линн
народов переселились в Австралию, где эволюционировали в австралийских аборигенов (Bradshaw, 1997). Близко родственные им народы, выжившие в горных районах Новой Гвинеи, известны как аборигены Новой Гвинеи.
Предки австралийских аборигенов и новогвинейцев никогда не испытывали суровых зим, начавшихся в Южной Азии с натиском первого ледникового периода примерно 70.000 лет назад. К этому времени они должны были находиться в Юго-Восточной Азии, в Индонезии или на Новой Гвинее, лежащих на экваторе или очень близко к нему. Они не были затронуты и вторым ледниковым периодом в Северном полушарии. Таким образом, австралийские аборигены и новогвинейцы имеют морфологические черты людей, эволюционировавших в тропической и субтропической среде и никогда не подвергавшихся воздействию умеренного климата. Они сходны с африканцами своей тёмной кожей, широким носом, длинными ногами, стройным туловищем и большими зубами.
Подобно другим народам, эволюционировавшим в тропической и субтропической зонах, новогвинейцы и австралийские аборигены имели возможность круглый год питаться растительной пищей, насекомыми и яйцами. Когда в двадцатом столетии жизнь австралийских аборигенов изучалась в пустыне Западной Австралии, было обнаружено, что 70-80 процентов их рациона составляет растительная пища, а большая часть остального рациона состоит из яиц и насекомых. У них не было хорошо разработанных методов групповой охоты (Gould, 1969). По оценке, у людей новогвинейского племени Гадио рацион состоит на 96 процентов из растительной пищи и только на 4 процента из мяса (Dornstreich, 1973). Лёгкая доступность в течение всего года растительной пищи наряду с насекомыми и яйцами означала, что коренные народы тропиков и субтропиков Новой Гвинеи и Австралии никогда не были должны зависеть от мяса как источника пищи и не попадали под сильное давление отбора, которое должно было развить когнитивные способности, требующиеся для охоты на крупных животных. Они не имели нужды и в изготовлении одежды для сохранения тепла. Даже на острове Тасмания, расположенном к Югу от Австралии, температура в июле, самом холодном месяце года, составляет в среднем 45 градусов по Фаренгейту (около ТС — прим перев.) и «тасманийцы обычно ходят нагишом» (Coon, 1967, стр. 114). Это объясняет, почему их интеллект и размер мозга имеют низкие значения: IQ 62 балла и средний размер мозга 1.225 см3. Оба эти показателя немного ниже, чем у африканцев с их IQ 67 и средним размером мозга 1.280 см3. Самое вероятное объяснение этого в том, что у африканцев в их более крупных популяциях шансы на появление мутаций, обеспечивающих повышение интеллекта, были намного выше, чем у австралийских аборигенов. Численность новогвинейских аборигенов в горной местности Новой Г винеи составляет
Расовые различия в интеллекте
249
около четверти миллиона. Численность аборигенов в Австралии в восемнадцатом столетии, когда там впервые появились европейцы, оценивается приблизительно в 300.000. В такой маленькой популяции вероятность появления новых мутаций, обеспечивающих увеличение интеллекта, должна была быть низкой, а географическая изоляция аборигенов Австралии и Новой Гвинеи предотвращала приобретение этих мутаций от других рас.
Когда европейцы открыли Австралию в конце восемнадцатого столетия, они обнаружили, что аборигены находятся на примитивном уровне культурного развития. «В их культуре мезолита (каменного века) не было (и всё ещё нет в отдаленных областях) глиняной посуды, сельского хозяйства или металлов» (Cole, 1965, стр. 82). Они не делали посевов, чтобы вырастить себе пищу, и не держали стад животных (Elkin, 1967). Они не хранили пищу для будущего потребления. По описанию Блекли (Bleakley, 1961, стр. 78): «Аборигены, как кажется, не имеют понятия о хранении запасов на случай голода». Томас (Thomas, 1925, стр. 295) описал аборигенов как «кочевников, не знающих ни глиняной посуды, ни металлообработки, не имеющих никаких домашних животных (наиболее прирученный — динго) и до сих пор не знают земледелия, поддерживая своё существование змеями и ящерицами, страусами эму, личинками и простой растительной пищей». «Их главные каменные орудия — каменный топор на рукоятке и нож, а также микролиты (маленькие каменные пластинки), используемые в качестве наконечников копий, зубцов пил-ножей и так далее. Оружие состоит из дубин, копий, копьеметалок и бумерангов. Женщины используют особые палки-копалки для выкапывания ямса и других корней» (Cole, 1965, стр. 83). Они никогда не изобретали и не заимствовали лук и стрелы (Coon, 1967). Некоторые из британских исследователей и ранних антропологов, изучавших аборигенов в девятнадцатом столетии, пришли к заключению, что те имеют низкий уровень интеллекта: «Они всё еще дети в своём умственном развитии» (Wake, 1872, стр. 80). В их языках отсутствовали числительные, кроме одного и двух: «два или пара представляют диапазон их числительных» (Crawfurd, 1883, стр. 170). Их языки были также лишены абстрактных понятий и «были бедны собирательными существительными» (Curr, 1886, стр. 20), что указывает на неспособность формулировать общие понятия, а это является одной из основных характеристик интеллекта. Аборигены, однако, делали примитивные рисунки фигур людей, уцелевшие в горном убежище Джинмиун на Северных территориях, которые были датированны возрастом приблизительно в 58.000 лет (Bradshaw, 1997).
Даймонд (Diamond, 1997, стр. 309), приписывает неуспех австралийских аборигенов в одомашнивании животных или в развитии сельского хозяйства «отсутствию поддающихся приручению животных, скудностью поддающихся окультуриванию растений и малопригодными почвами и
250
Ричард Линн
климатом», но на той же странице он говорит нам, что ямс, таро и канна произрастают в диком виде в Северной Австралии и могут возделываться, а также имеются два вида местных диких трав, которые можно подвергнуть селекции, чтобы получить зерновые культуры. Кенгуру и динго могли бы быть одомашнены путем селекции в течение ряда поколений. Климат Австралии очень разнообразен, и, не считая пустынь центрального региона, потенциально пригоден для сельского хозяйства, которое было развито в течение девятнадцатого и двадцатого столетий европейцами.
Тасманийцы находились даже на ещё более низком уровне культурного развития, чем аборигены австралийского материка. Российский антрополог Владимир Кабо (Kabo, 1985, стр. 603) писал, что они — «единственное общество, которое сохранилось на уровне позднего палеолита вплоть до начала европейской колонизации». Капитан Уильям Блай посетил Тасманию в 1788 г. и описал тасманийцев как кочевых охотников-со- бирателей, имевших «небольшое число убогих вигвамов, в которых не было ничего, кроме нескольких шкур кенгуру, расстеленных на земле», «они переходили из одной области в другую, добывая по дороге пропитание сбором ягод, фруктов и семян различных кустарников. Кроме водорослей, они редко носили с собой какую-либо пищу» и «они обычно ходили нагими, но иногда накидывали на тело шкуру кенгуру» (Bowdler, Ryan, 1997, стр. 313-326). Они — единственный известный народ, никогда не научившийся добывать огонь (Gott, 2002). Они иногда могли получать огонь от самопроизвольных лесных пожаров, но когда те гасли, то должны были ждать нового лесного пожара или получить огонь от соседней группы. Они никогда не придумали, как насаживать острые камни на деревянное древко, чтобы сделать копьё или топор (Ryan, 1992).
Когда европейцы обнаружили новогвинейцев в семнадцатом и восемнадцатом столетиях, они застали их на стадии культурного развития немного более высокой, чем у австралийских аборигенов. Новогвинейцы были преимущественно охотниками-собирателями, но имели некоторое сельское хозяйство, заключавшееся в выращивании ямса и бананов, у них также были домашние куры и свиньи. Но, «пока европейцы не начали колонизировать их, все новогвинейцы были неграмотными, пользовались каменными орудиями и не были еще политически объединены в государства или (за редким исключением) в союзы племён (chiefdoms)» (Diamond, 1997, стр. 299). После европейской колонизации некоторые из аборигенов переселились в города и посёлки, а другие остались в своих поселениях, живя натуральны хозяйством. Европейцы построили и укомплектовали школы для проживающих в городах и посёлках, и школы-интернаты для живущих в своих селениях, хотя некоторые сельские дети не посещали школу. Келли (Kelly, 1977) описал образ жизни типичных племён, живущих в своих селениях и в посёлках на Папуа-Новой Гвинее в 1970-х гг.
Расовые различия в интеллекте
251
Они существовали в основном за счёт подсечно-огневого земледелия, которым занимались преимущественно женщины. Мужчины немного промышляли охотой, а некоторые из них работали за деньги на плантациях кофе, управляемых европейцами. Одежда менее развитых племён состояла из юбок, изготовленных из листьев и коры. Некоторые племена имели системы счёта, позволявшие им считать до тысячи, в то время как у других были только слова, обозначающие: «один», «больше одного» и «много». Главная причина того, что новогвинейцы были немного более развиты в сравнении с австралийскими аборигенами, состояла в том, что прибрежные области острова были заселены выходцами из Юго-Восточной Азии и с островов Меланезийского архипелага Тихого океана, которые принесли с собой таро (выращиваемый ими съедобный корень), а также одомашненных кур и свиней. Новогвинейцы переняли некоторые из этих культурных новшеств, но никогда не развивали чего-нибудь, что можно было бы назвать цивилизацией с городами, капитальными зданиями, обработкой металла, письменностью или арифметикой.
Европейцы
Некоторые из народов, колонизировавших Ближний Восток 100.000
лет назад, мигрировали к Северу и приблизительно 60.000 лет назад достигли Кавказа, откуда распространились на Украину и затем, примерно 40.000 лет назад, в Центральную и Западную Европу. Другие народы Юго-Западной Азии начали колонизировать Юго-Восточную Европу из Анатолии. Эти народы эволюционировали в европейцев с более светлой кожей и, на Севере Европы, со светлыми волосами и голубыми глазами. Европейцы были в значительной мере изолированы от аборигенов Южной Азии и Северной Африки на Юге Средиземным морем, а на Востоке Чёрными и Каспийским морями, высокими горами Кавказа и Гималаев, а также пустыней Кара-Кум в нынешней Туркмении. В течение последнего ледникового периода, который продлился примерно от 28.000 до 10.000 лет назад, зимы в Европе были значительно холоднее, чем в Южной Азии, температура самого холодного зимнего месяца снизилась примерно до - 5°С. Ландшафт в Европе стал похожим на существующий ныне в современной Аляске и в Сибири. Север Англии, Германии, России и вся Скандинавия были покрыты постоянным ледяным покровом, а остальная часть Европы представляла собой холодные луга и тундру с отдельными рощами деревьев в защищённых местах.
Эти холодные зимы должны были быть главным фактором давления отбора в сторону увеличения размера мозга и интеллекта европейцев,
252
Ричард Линн
повысившим размер их мозга в среднем до 1.369см3, a IQ до 99 баллов. Выразив увеличение размера мозга коэффициентом энцефализации (EQ) для учёта размера тела, Катлер (Cutler, 1976) оценил, что у «до-вюрмских» европейцев EQ составлял 7,3, а к концу Вюрмского оледенения EQ европейцев равнялся 8,1. Когда отступили ледяные пласты, покрывавшие Северную Европу примерно до 10.000 лет назад, европейцы с их возросшим интеллектом оказались в состоянии совершить неолитический переход к осёдлому сельскому хозяйству. Однако, несмотря на свой высокий IQ, они не были в состоянии создать ранние цивилизации, подобные созданным аборигенами Южной Азии и Северной Африки, потому что в Европе всё еще было холодно, она была покрыта лесами и её почвы были тяжёлыми, их было трудно пахать в отличие от лёгких почв, на которых создавались ранние цивилизации. В Европе также не было речных пойм, обеспечивающих высоко плодородные аллювиальные отложения, на которых ежегодно можно было получать излишки сельскохозяйственной продукции для обеспечения городской цивилизации и интеллектуального класса (Landes, 1998). Приблизительно с 2.500 г. до нашей эры европейцы преодолели эти трудности в относительно мягком климате Южной Европы, где они создали свои первые цивилизации на острове Крит и в Греции. Примерно с 700 г. до н.э. италики начали построение цивилизации, которая со временем стала Римской империей и к 200 г. нашей эры охватила всю Европу к Западу от Рейна и включала бассейн Дуная, Ближний Восток и Северную Африку. Эти первые европейские цивилизации Греции и Рима превзошли цивилизации Южной Азии и Северной Африки в науке, математике, технике, литературе, философии и искусствах. Западная Римская империя пала в 455 г. нашей эры и европейская культура находилась в упадке в последовавшие средние века, но приблизительно с 1.000 года нашей эры она возродилось, а примерно с 1.500 г. европейцы стали передовыми народами фактически во всех областях цивилизации, как это обстоятельно документировано Мюрреем (Murray, 2003).
Генетические процессы, посредством которых развился более высокий IQ европейцев, должны были состоять в изменениях частот аллелей в сторону увеличения пропорции аллелей высокого интеллекта и, вероятно, также в появлении новых мутаций, повышающих интеллект, и их быстрого распространения в популяции. Вероятность появления новых мутаций более высокого интеллекта у европейцев должна была быть увеличена стрессом, вызванным чрезвычайным холодом, действию которого подвергались европейцы.
Более низкие значения IQ в Юго-Восточной Европе (90-94 балла) следует отнести, вероятно, на счёт некоторого генного потока между аборигенами Южной Азии и европейцами через Дарданеллы и Эгейское море, что привело к возникновению градиента признаков южно-азиатских и ев
Расовые различия в интеллекте
253
ропейских гибридов на Балканах с IQ, промежуточным между европейцами (99 баллов) и аборигенами Южной Азии (84 балла). Такой же градиент признаков присутствует в Турции, где IQ составляет около 90 баллов — лишь незначительно ниже, чем на Балканах.
Аборигены Восточной Азии
Некоторые из народов Южной и Средней Азии начали колонизировать Северо-Восточную Азию в районе современного Китая от 60.000 до
лет назад, где они эволюционировали в аборигенов Восточной Азии и позже в арктические народы крайнего Северо-Востока. Архаические народы Восточной Азии были в значительной степени изолированы от европейцев пустыней Гоби на Западе и от аборигенов Южной Азии Гималаями на Юге. Зимы, которые они переживали, были намного более суровыми, чем в Южной Азии и несколько более суровыми, чем в Европе: во время главного Вюрмского оледенения минимальная зимняя температура снизилась приблизительно до -12°С. Причины сильных зимних холодов в Северо-Восточной Азии по сравнению с Европой состоят в том, что Северо-Восточная Азия представляет собой намного больший массив суши, чем Европа, и Европу согревают преобладающие западные ветры с Атлантики. Именно в ответ на холодные зимы у аборигенов Восточной Азии развились такие приспособительные особенности, как уплощённый нос для предотвращения обморожения, короткие ноги и широкое туловище для сохранения тепла, слой подкожного жира, который придаёт коже желтоватый оттенок, редкие волосы на лице у мужчин (поскольку густая борода замёрзла бы и привела к обморожению) и эпикантус, смягчающий ослепляющее действие света, отражённого от снега и льда. Суровые зимы должны были обеспечить жёсткий отбор по интеллекту и повысили IQ народов Восточной Азии до 105 баллов. Действовавшие генетические процессы представляли собой, вероятно, увеличение частот аллелей высокого интеллекта, а также появление новых мутаций более высокого интеллекта, возникающих случайно и под влиянием сильного холодового стресса. Появлением новых мутаций можно объяснить, почему у аборигенов Восточной Азии особенно высока способность к визуализации по сравнению с европейцами. Новые мутации, обеспечивающие повышенную способность к визуализации, могли появиться у аборигенов Восточной Азии и распространиться в популяции потому, что были полезны для охоты, изготовления орудий и ориентирования на больших пространствах лишённой выраженных примет местности.
Как и в случае европейцев, большая часть увеличения интеллекта аборигенов Восточной Азии произошла, вероятно, во время главного Вюрмского оледенения. Оно действовало как давление отбора в сторону увеличения мозга
254
Ричард Линн
и должно было увеличить их IQ до его современной величины в 105 баллов. Только после окончания Вюрмского оледенения их интеллект достиг уровня, который позволил им совершить неолитический переход к осёдлому сельскому хозяйству и затем построить цивилизацию в долине Жёлтой реки и в дальнейшем более развитые цивилизации в Китае, Японии и Корее. В период приблизительно 0-1.500 гг. нашей эры китайцы создали внушительные цивилизации, которые в некоторых отношениях превосходили существовавшие в Европе. Например, китайцы изобрели книгопечатание, бумагу, бумажные деньги, порох, магнитный компас и устройство каналов со шлюзами за несколько столетий до европейцев. В период с 1.500 г. до настоящего времени интеллектуальные достижения аборигенов Восточной Азии были, однако, менее внушительными, чем у европейцев, как было исчерпывающе документально подтверждено Мюрреем (Murray, 2003). Историки расценивают это как большую загадку, которая не имеет единодушно признанного решения. Одним из факторов может быть то, что у аборигенов Восточной Азии в большей степени, чем у европейцев, развился социальный конформизм (см. Allik, Realo, 2004), выразившийся, в частности, в низком уровне психотизма, как показано в моей работе (Lynn, 2002). Низкий уровень социального конформизма и некоторый пси- хотизм, по-видимому, благоприятствуют творческим достижениям, потому что они снижают тревогу по поводу социального неодобрения и, вероятно, облегчают генерацию оригинальных идей, необходимых на высших уровнях научного поиска. Другим фактором, по предложению Виде и Кам- пфа (Weede, Kampf, 2002), может быть исторический «несчастный случай», состоявший в том, что на протяжении большей части свой истории Китай был единым государством, деспотичные правители которого были в состоянии подавить свободы, включая свободу мысли, более эффективно, чем правители многочисленных европейских государств, которые были вынуждены конкурировать в части предоставления свобод своим народам.
Арктические народы
Когда-то, 50.000-40.000 лет назад, некоторые из архаичных народов Восточной Азии мигрировали на крайний Северо-Восток Азии, где они эволюционировали в арктические народы. Эти народы развились в отдельную расу, потому что были географически изолированы от аборигенов Восточной Азии на Юге высокими горами хребта Черского, Хингана и Саян, и приблизительно тысячей миль лесов к Северу от реки Амур. Среди всех рас арктические народы испытывали наиболее суровые зимы с самыми низкими зимними температурами около -15°С, снизившимися примерно до -20°С во время главного Вюрмского оледенения. В ответ на эти
Расовые различия в интеллекте
255
холодные зимы у арктических народов развились более выраженные формы морфологической Холодовой адаптации, присущие аборигенам Восточной Азии: уплощённый нос, короткие ноги и широкое туловище, слой подкожного жира, придающий коже желтоватый оттенок, и эпикантус. Эти суровые зимы, как можно ожидать, должны были действовать как сильное давление отбора в сторону увеличения интеллекта, но этого явно не произошло, так как IQ арктическихнародов составляет всего 91 балл.
Объяснение этому должно заключаться в малочисленности арктических народов, популяция которых в конце двадцатого столетия составляла лишь приблизительно 56 тысяч человек в сравнении с приблизительно 1,4 миллиардами аборигенов Восточной Азии. Хоть и невозможно точно оценить величины популяций во время главного Вюрмского оледенения, несомненно, что аборигены Восточной Азии были многократно более многочисленными, чем арктические народы. Результатом различия размеров популяций будет то, что мутации более высокого интеллекта, появившиеся и распространившиеся у аборигенов Восточной Азии, никогда не появлялись у арктических народов. Аборигены Восточной Азии, состоящие из китайцев, корейцев и японцев, должны были образовывать единую протяжённую популяцию обменивающихся генами демов, в которых распространялись мутантные аллели высокого интеллекта, но эти аллели не передались арктическим народам, изолированным высокими горными цепями и большими расстояниями. У арктических народов, однако, развился крупный мозг, приблизительно такого же размера, как у аборигенов Восточной Азии, так что удивительно, что они не обладают таким же интеллектом. Возможное объяснение этого в том, что у арктических народов развилась хорошая визуальная память, которая была им необходима, когда они отправлялись далеко на охоту и должны были запоминать ориентиры для того, чтобы вернуться домой в основном по лишённой примет местности, покрытой снегом и льдом. Увеличение этой способности должно было потребовать увеличения мозга, но она не измеряется тестами интеллекта. Другая возможность состоит в том, что одна или более новых мутантных аллелей, обеспечивающих большую эффективность лежащих в основе интеллекта нейрофизиологических процессов, могли появиться у аборигенов Восточной Азии и не появиться у арктических народов.
В том, что известно об интеллекте народов Северо-Восточной Азии, имеется ещё одна аномалия, относящаяся к IQ монголов в Монголии и тесно связанных с ними самодийцами северной Сибири. Интеллект этих народов не исследовался, но низкий уровень их культурного развития и техники даёт основание предположить, что он не столь высок, как у аборигенов Восточной Азии в Китае, Японии и Корее. Но эти народы также испытывали воздействие суровых зим в течение многих тысяч лет, что вызвало у них, как и у арктических народов, появление выраженных морфологических
256
Ричард Линн
особенностей, обеспечивающих холодовую адаптацию — эпикантус, короткие ноги и широкое туловище. Вероятное объяснение этой аномалии состоит в малочисленности этих народов (население современной Монголии составляет приблизительно 2,4 миллиона человек и всего несколько десятков тысяч самодийцев живёт в Северной Сибири) и в том, что они были изолированы от соседних народов пустыней Гоби и высокими горными цепями, так что новые мутации, обеспечивающие более высокий интеллект, у них не появились, а географическая изоляция должна была предотвратить приобретение ими этих мутаций от других рас.
Американские индейцы
Американские индейцы эволюционировали от народов, мигрировавших из Северо-Восточной Азии через Берингов пролив на Аляску и затем проложивших путь к Югу Америки. По поводу того, когда произошёл этот переход, идёт спор, но часто утверждается, что он произошёл приблизительно 12.000-11.000 лет назад. Вопреки этим утверждениям, имеются убедительные данные в пользу того, что он произошёл намного раньше, примерно 40.000 лет назад. Об этом свидетельствуют как данные археологии, так и генетический анализ. Археологические находки изделий индейцев датируются радиоуглеродным методом возрастом 24.000 лет в Мексике (Lorenzo, Mirambell, 1996), 30.000 лет в Калифорнии (Bada, Schroeder, Carter, 1974), 32.000 лет на Северо-Востоке Бразилии (Guidon, Delibrias, 1996), 35.000-43.000 лет — наскальная живопись в Национальном парке Сиерра да Капивара в Северо-Восточной Бразилии (Watanabe, Ayta, Mamaguchi, et al., 2003) и 33.000 лет в Монте Верде в Чили (Dillehay, Collins, 1998). Этим народам должно было потребоваться несколько тысяч лет для того, чтобы добраться от Аляски до Южной Америки. Таким образом, археологические данные свидетельствуют о том, что первые люди пересекли Берингов пролив, по крайней мере, 40.000 лет назад. Эти археологические данные подкрепляются генетическим анализом, согласно которому первая миграция в Америку произошла приблизительно 40.000 лет назад (Cavalli-Sforza, 2000).
Наиболее вероятным представляется, что приблизительно 50.000 лет назад в Северо-Восточной Азии жили архаичные восточноазиатские народы, некоторые из которых мигрировали к Северу на Камчатку и на полуостров Черского, а затем приблизительно 40.000 лет назад совершили переход через Берингов проливов на Аляску. Некоторые из этих народов передвигались на Юг, пока не колонизировали оба континента Америки и не эволюционировали в индейцев, а архаичные народы Восточной Азии, остававшиеся в Северо-Восточной Азии, эволюционировали в современ
Расовые различия в интеллекте
257
ных аборигенов Восточной Азии. Относительно недавнее общее происхождение этих двух рас очевидно из ряда общих генетических черт. Например, у обеих рас аллель резус-отрицательной группы крови встречается редко, группа крови Диего уникальна для этих двух рас, обе они имеют похожие жёсткие, прямые чёрные волосы, лопатовидные резцы и «кость инков» в черепе (Krantz, 1990).
Архаичные восточноазиатские предки американских индейцев, обитавшие в Северо-Восточной Азии около 60.000-50.000 лет назад, подвергались действию холодных зим, но не столь суровых, как во время главного Вюрмского оледенения примерно 28.000-10.000 лет назад (Roberts, 1994), когда предки индейцев колонизировали материки Америки. Следовательно, американские индейцы никогда не подвергались действию чрезвычайного холода и не имеют морфологических особенностей, обеспечивающих адаптацию к нему, которые развились у аборигенов Восточной Азии. Нос у них не вдавленный, но весьма выступающий, они не имеют присущих аборигенам Восточной Азии полного эпикантуса, коротких ног и широкого туловища. В этом отношении они сходны с айнами, коренными жителями Японии, небольшое число которых сохранилось на о-ве Хоккайдо, которые также не имеют присущих аборигенам Восточной Азии морфологических черт, обеспечивающих адаптацию к холоду, потому что климат японских островов был более морским и менее суровым, чем в материковой части Северо-Восточной Азии.
Таким образом, индейцы расселились по обоим континентам Америки примерно 33.000-30.000 лет назад. Те из них, кто находился в южной части Соединенных Штатов и в Центральной и Южной Америке, не испытывали суровых условий главного Вюрмского оледенения, и у них не развилось ни морфологических адаптаций к холоду ни высокого IQ, присущего аборигенам Восточной Азии. Более того, как только предки американских индейцев пересекли Берингов пролив и проложили путь на американские континенты, они обнаружили, что условия жизни там намного легче тех, к которым привыкли их предки в Северо-Восточной Азии. Они должны были встретить массу травоядных млекопитающих, таких как мамонты, антилопы, ленивцы, броненосцы и бизоны, незнакомых с охотниками. Обычно хищники и добыча развиваются вместе, в том смысле, что интеллект хищников становятся более высоким, чтобы поймать добычу, а интеллект добычи возрастает, чтобы ускользнуть от хищников. Но травоядные животные Америки не имели опыта общения с человеком-охот- ником и должны были представлять лёгкую добычу для искусных охотников, эволюционировавших в течение многих тысяч лет в более суровой среде Северо-Восточной Азии. Американские индейцы должны были встретить большое количество травоядных животных, которых было легко добыть, а поскольку они мигрировали к Югу, то должны были находиить
9 Ричард Линн
258
Ричард Линн
больше доступной растительной пищи, так что растительная пища стала играть существенную роль в их рационе (MacNeish, 1976; Hayden, 1991).
Развитие интеллекта американских индейцах может быть реконструировано следующим образом. Архаичные народы Восточной Азии, от которых они произошли, должны были иметь более высокий интеллект, чем аборигены Южной Азии, потому что они подвергались действию холодного климата Северо-Восточной Азии в течение приблизительно 20.000 лет, примерно от 60.000 до 40.000 лет назад. Предки американских индейцев провели ещё несколько тысяч лет на Аляске, в течение которых они подвергались действию сурового климата, что должно было ещё больше повысить их интеллект. Как только они оказались в Америке южнее Аляски, давление отбора в сторону какого-либо дальнейшего увеличения интеллекта должно было ослабнуть по причине мягкого климата и лёгкости выживания на континенте, ещё не затронутом человеком. Это объясняет их нынешний IQ в 86 баллов — немного выше 84 баллов аборигенов Южной Азии, но много ниже 105 баллов аборигенов Восточной Азии. Эта реконструкция даёт новое свидетельство в пользу того, что именно давление отбора, оказанное главным Вюрмским оледенением в период приблизительно от 28.000 до 10.000 лет назад, должно было увеличить интеллект аборигенов Восточной Азии на величину примерно в 19 баллов IQ в сравнении с американскими индейцами.
Слабое место этой реконструкции в том, что американские индейцы в северной части Северной Америки должны были подвергнуться действию суровых зим во время главного Вюрмского оледенения и можно было бы ожидать, что это должно было повысить их интеллект. Наиболее вероятное объяснение того, почему это не произошло, заключается в малочисленности популяции индейцев Северной Америки. Самая ранняя заслуживающая доверия оценка размера их популяции относится к 400 г. до н.э. и определяет их число в Северной Америке приблизительно в 1 миллион (Biraben, 1980). Таким образом, вероятность мутаций, обеспечивающих повышение интеллекта индейцев в северной части Северной Америки, была весьма мала, и, возможно, таких мутаций не было или их было меньше, чем у существенно более многочисленных аборигенов Восточной Азии и европейцев.
Американские индейцы имеют такой же профиль интеллекта, как и аборигены Восточной Азии и арктические народы: высокая способность к визуализации и более слабая вербальная способность. Вероятное объяснение этого общего профиля состоит в том, что одна или больше мутаций, обеспечивающих повышение способности к визуализации, появились в предковой популяции архаичных народов Восточной Азии приблизительно 50.000 лет назад и передались впоследствии аборигенам Восточной Азии, арктическим народам и американским индейцам, которые эволюциони
Расовые различия в интеллекте
259
ровали от этой предковой популяции. Генетические исследования показали, что имеются гены, определяющие выраженность способности к визуализации в дополнение к генам, определяющим выраженность вербальной способности и фактора g (Plomin, DeFries, McCleam, 1990).
Со своим IQ в 86 баллов американские индейцы были в состоянии совершить неолитический переход от охотников-собирателей к оседлому сельскому хозяйству и затем построить цивилизации майя, ацтеков и инков. Причина того, что эти цивилизации были созданы в Центральной и Южной, а не в Северной Америке, состоит, вероятно, в том, что численность населения в Центральной и Южной Америке была намного большей (приблизительно 11 миллионов против лишь 2 миллионов в Северной Америке в 500 г. нашей эры — Biraben, 1990). Однако, несмотря на свои дольно впечатляющие цивилизации, американские индейцы оказались неравными соперниками для европейцев, которым, начиная с шестнадца- того-семнадцатого столетий, не составляло большого труда побеждать их в битвах, захватывать большую часть их земель и истреблять их в больших количествах.
Заключение
Показатели интеллекта рас, рассмотренные в Главах 3-12, могут быть объяснены как возникшие благодаря различиям окружающей среды, в которой они эволюционировали, в особенности под влиянием ледниковых периодов в Северном полушарии, оказывавших давление отбора в сторону повышения интеллекта, требующегося для выживания в течение холодных зим; и, в дополнение к этому, возникновению мутаций более высокого интеллекта, появившихся в более многочисленных расах и под влиянием максимального холодового стресса. Различия в IQ между расами объясняют различия в успехах в осуществлении неолитического перехода от стадии охотников-собирателей к осёдлому сельскому хозяйству, в построении ранних цивилизаций и в развитии зрелых цивилизаций в течение последних двух тысяч лет. Точка зрения сторонников средовой теории, заключающаяся в том, что в течение около 100.000 лет народы в разных частях света, разделённые географическими барьерами, эволюционировавшие в десять различных рас с явными генетическими различиями в морфологии, группах крови и частотах генетических болезней, все же имеют идентичные генотипы по интеллекту, является настолько невероятной, что те, кто защищают её, должны или быть полностью невежественны в основах эволюционной биологии или же руководствоваться политическим предписанием отрицать важность расы. Либо и то, и другое.
ПРИПОЯМ
muштшшл12
ААВ. Американский армейский бета, разработанный для тестирования личного состава армии во время первой мировой войны. Невербальный тест общего интеллекта, на котором основываются субтесты действия тестов Векслера.
AFQT. Вооружённых сил квалификационный тест. Преимущественно вербальный тест общего интеллекта.
АН. Alice Heim. Тест способности к вербальному и невербальному рассуждению.
АР. Alexander Passalong тест. Невербальный тест интеллекта и способности к визуализации, представляет собой ряд неглубоких коробочек в которых находятся цветные квадратные и прямоугольные кубиков. Задача состоит в том, чтобы перерасположить кубики таким образом, чтобы все красные оказались на одном конце, а синие на другом.
Артура Пойнта действия шкала. Невербальный тест общего интеллекта.
BAS. Британские способностей шкалы. Тест общего интеллекта, вербальных и невербальных способностей
BG. Bender Gestalt. Бендер зрительно-моторный гештальт тест. Рисуночный тест общего интеллекта.
ВТВС. Боэма тест базисных понятий. Тест общего интеллекта, оценивающий вербальное понимание пространственных и количественных понятий.
BTBC-R. Боэма тест базисных понятий пересмотренный. Пересмотренная версия ВТВС.
ССАТ. Канадский когнитивных способностей тест. Тест вербального, количественного и невербального рассуждения.
CEFT. Детский встроенных фигур тест. Детская версия EFT: Тест на способность найти простую фигуру, встроенную в боульшую фигуру.
CF. Кэттелла культурно-беспристрастный тест. Невербальный тест общего интеллекта.
CITO. Голландский тест, оценивающий числовое рассуждение и вербальное понимание.
12 Ниже приводятся краткие описания тестов, аббревиатуры которых встречаются в таблицах. При переводах названий некоторых тестов мы руководствовались вариантами, данными в книге А.Анастази и С.Урбина «Психологическое тестирование».
Расовые различия в интеллекте
261
СММ. Колумбийская шкала умственной зрелости. Тест вербального и невербального рассуждения на общий интеллект.
СРМ. Цветные прогрессивные матрицы. Тест невербального рассуждения для детей 5-11 лет.
СРМТ. Тест на визуализацию.
СТММ. Калифорнийский тест умственной зрелости. Тест невербального рассуждения на общий интеллект.
DAM. Гудинаф нарисуй человечка тест. Рисуночный тест общего интеллекта.
EFT. Встроенных фигур тест. Тест на способность найти простую фигуру, включенную в боульшую фигуру. Коррелирует со шкалой действия (0,65) и с вербальной шкалой (0,30) теста Векслера для детей (Witkin et al., 1962).
EPVT. Английский картиночный словарный тест.
FF. Фергюссона форм доски. Тест на визуализацию, в котором требуется вставить плоские фигуры различной формы в ячейки.
GALO. Голландский тест общего интеллекта.
GFT. Готтшальдта фигуры тест. Тест на визуализацию.
GMRT. Групповой вращения в уме тест. Тест на визуализацию.
GSAT. Общих академических способностей тест. Южно-африканский тест на рассуждение, визуализацию, вербальную и другие способности.
ITPA. Иллинойский тест психолингвистических способностей. Измеряет 12 аудиторных (вербальных) и визуальных языковых способностей.
JAT. Юношей способностей тест. Южно-африканский тест с 10 субтестами, оценивающими рассуждение, вербальные, пространственные и др. способности.
КАВС. Кауфмана оценочная батарея для детей. Тест общего интеллекта, имеющий сходство с тестами Векслера.
KAIT. Кауфмана подростков и взрослых интеллекта тест. Тест общего интеллекта, имеющий сходство с тестами Векслера.
LPT. Способности к обучению тест. Тест общего интеллекта.
LT. Лордж-Торндайк. Тест общего интеллекта.
Матричных аналогий тест. Невербального рассуждения тест.
MFFT. Подбора знакомых фигур тест.
МН. Моррея дом. Вербальный тест общего интеллекта.
MHV. Милл Хилл словарь. Показатель вербальной способности.
MMFT. Подбора знакомых фигур тест. Тест преимущественно на визуализацию.
MMSE. Умственного состояния мини-обследование. Тест общего интеллекта.
NFER. Британской национальной организации для исследований в области образования тест. Тест невербального рассуждения и вербальной способности.
262
Ричард Линн
ОТ. Отиса тест. Преимущественно вербальный тест общего интеллекта.
РАТ. Прогрессивных достижений тест. Вербальный тест общего интеллекта.
PIPS. Тихоокеанский младенцев действия тест. Невербальный тест общего интеллекта.
PNL. Пинтнера неязыковой тест. Невербальный тест общего интеллекта.
РРМА. Первичный тест на музыкальный слух. Тест музыкальных способностей.
PPVT. Пибоди словарный тест в картинках. Набор из 4 картинок различных объектов, которые должны быть названы.
QT. Квинслендский тест. Невербальный тест общего интеллекта.
RACIT. Голландский тест, включающий ряд субтестов, измеряющих рассуждение, вербальную, пространственную и др. способности.
SA. Стэнфордский тест достижений. Вербальный тест на понимание значений слов, правописание и арифметику.
SB. Стэнфорд-Бине. Преимущественно вербальный тест общего интеллекта.
Сишор. Тест музыкальных способностей.
SON-R. Снайдерса-Оомана невербального интеллекта тест. Невербальный тест общего интеллекта.
SOT. «Спиральный омнибус» тест. Тест на рассуждение.
SRAT. Научных исследований объединения тест. Тест общего интеллекта.
STAS. Стэнфордский тест учебных навыков. Тест для ряда учебных предметов.
TOSCA. Тест академических способностей. Вербальный и числовой тест общего интеллекта.
WAIS. Векслера интеллекта взрослых шкала. Измеряет общий и вербальный интеллект, визуализацию.
WB. Векслер-Бельвю. Измеряет общий интеллект, вербальную способность и визуализацию.
WCST. Висконсинский карточек сортировки тест. Невербальный тест общего интеллекта.
WISC. Векслера интеллекта шкала для детей. Измеряет общий и вербальный интеллект, визуализацию.
WPPSI. Векслера интеллекта шкала для дошкольников и младших школьников. Измеряет общий и вербальный интеллект, визуализацию у 4-6 летних детей.
WRAT. Широкого диапазона достижений тест. Тест общего интеллекта.
3DW. Австрийский тест общего интеллекта.
СПИСОК ШШТУРИ
Abbink, J. G. (2002). Ethnic trajectories in Israel. Anthropos, 97, 3-19.
Abdel-Khalek, A. M. (1988). Egyptian results on the Standard Progressive Matrices. Personality and Individual Differences, 9, 193-195.
Abdel-Khalek, A. М., and Lynn, R. (2005). Sex differences on a standardisation of the Standard Progressive Matrices in Kuwait. Personality and Individual Differences (to appear).
Abell, S. C., Wood, W., and Leibman, S. J. (2001). Children’s human figure drawings as measures of intelligence: the comparative validity of three scoring systems. Journal of Psychoeducational Assessment, 19, 204-215.
Abul-Hubb, D. (1972). Application of Progressive Matrices in Iraq. In L. J. Cronbach and P. J. Drenth (Eds.). Mental Tests and Cultural Adaptation. The Hague: Mouton.
Adcock, C. J., McCleary, J. R., Ritchie, J. E., and Somerset, H. C. (1954). An analysis of Maori scores on the Wechsler-Bellvue. Australian Journal of Psychology, 6, 16-29.
Afzal, M. (1988). Consequences of consanguinity on cognitive behavior. Behavior Genetics, 18, 583-594.
Agrawal, N., Sinha, S. N., and Jensen, A. R. (1984). Effects of inbreeding on Raven matrices. Behavior Genetics, 14, 579-585.
Ahmed, R. A. (1989). The development of number, space, quantity and reasoning
concepts in Sudanese schoolchildren. In L. L. Adler (Ed.). Cross Cultural Research in Human Development. Westport, CT: Praeger.
Albalde Paz, E., and Munoz, C. J. (1993). El test PMS de Raven у los escolares de Galicia. Universidade da Coruna: Servicio de Publicaciones.
Alexander, R. D. (1989). Evolution of the human psyche. In P. Mellors and C. Stringer (Eds.). The Human Revolution. Edinburgh: University of Edinburgh Press.
Al-HeetiK., Ganem, A., Al-Kubaldl, A., and Al-Nood, Y. (1997). Standardization of Raven’s Coloured Progressive Matrices Scale on primary school children ages 6-11 in Yemen schools. Indian Psychological Review, 48, 49-56.
Allik, J., and Realo, A. (2004). Individualism collectivism and social capital. Journal of Cross-Cultural Psychology, 35, 29-49.
Alvi, S. A., Khan, S. B., Vegeris, S. L., and Ansari, Z. A. (1986). A cross-cultural study of psychological differentiation. International Journal of Psychology, 21, 659-670.
Alzobaie, A. J. (1964). The Cattell Culture-Free Test as tried on Iraqi students. Journal of Educational Research, 57, 476-479.
Amir, Y. (1975). Perceptive articulation in three Middle Eastern cultures. Journal of Cross-Cultural Psychology, 6, 333-344.
Anderson, B. (1993). Evidence from the rat for a general factor that underlies cognitive performance and that relates to brain size: intelligence. Neuroscience Letters, 153, 98102.
Ankney, C. D. (1992). Sex differences in relative brain size: the mismeasure of intelligence in women, too? Intelligence, 16, 329-336.
A. P. E. (1960). Australia. Encyclopedia Britannica. Chicago: Benton.
Ardila, A., Pineda, D., and Rosselli, M. (2000). Correlation between intelligence test scores and executive function measures. Archives of Clinical Neuropsychology, 15, 31-36.
Arthur, G. (1941). An experience in testing Indian school children. Mental Hygiene, 25, 188-195.
Ausubel, D. P. (1961) Maori Youth. New York: Holt, Rinehart & Winston.
264
Ричард Линн
Avenant, Т. J. (1988). The Establishment of an Individual Intelligence Scale for Adult South Africans. Report No. P-91. Pretoria: Human Sciences Research Council.
Avolio, B. J., and Waldman, D. A. (1994). Variations in cognitive, perceptual and
psychomotor abilities across the working life span: examining the effects of race, sex, experience, education and occupational type. Psychology and Aging, 9, 430-442.
Baare, W. R C., Pol, H. E. H., Boosma, D. L, Postuma, D., and Geus, E. J. C. (2001). Quantitative genetic modelling of variation in human brain morphology. Cerebral Cortex, 11, 816-824.
Backman, М. E. (1972). Patterns of mental abilities: ethnic, socioeconomic and sex differences. American Educational Research Journal, 9, 1-12.
Bada, J. L., Schroeder, R. A., and Cater, G. F. (1974). New evidence for the antiquity of man in North America deduced from aspartic and recemization. Science, 184, 791-793.
Badri, М. B. (1965a). The use of finger drawing in measuring the Goodenough quotient of culturally deprived Sudanese children. Journal of Psychology, 59, 333-334.
Badri, М. B. (1965b). Influence of modernization on Goodenough quotients of Sudenese children. Perceptual and Motor Skills, 20, 931-932.
Bagley, C., Iwawaki, S., and Young, L. (1983). Japanese children: group-oriented but not field dependent? In C. Bagley and G. K. Verma (Eds.). Multicultural Childhood, Education, Ethnicity and Cognitive Styles. Aldershot, UK: Gower.
Baker, D. B., and Jones, D. P. (1993). Creating gender equality: cross-national gender stratification and mathematical performance. Sociology of Education, 66, 91-103.
Baker, J. R. (1974). Race. Oxford, UK: Oxford University Press.
Barnabus, I. P., Kapur, М., and Rao, S. (1995). Norm development and reliability of Coloured Progressive Matrices Test. Journal of Personality and Clinical Studies, 11, 17-22.
Bart,W., Kamal, A., and Lane, J. F. (1987). The development of propor-tional reasoning in Qatar. Journal of Genetic Psychology, 148,95-103.
Beals, K. L., Smith, C. L., and Dodd, S. M. (1984). Brain size, cranial morphology, climate and time machines. Current Anthropology. 25, 301-330.
Beaton, A. E., Mullis, I. V. S., Martin, М. O., Gonzales, E. J., Kelly, D. L., and Smith, T. A. (1996). Mathematics Achievement in the Middle School Years. Chestnut Hill, MA: Boston College, TIMSS International Study Center.
Beaton, A. E., Martin, М. O., Mullis, I. V. S., Gonzales, E. J., Smith, T. A., and Kelly, D. L., (1996). Science Achievement in the Middle School Years. Chestnut Hill, MA: Boston College, TIMSS International Study Center.
Beaumont, P. B., De Villiers, H., and Vogel, J. C. (1978). Modern man in sub-Saharan Africa prior to 49,000 years B. P.: A review and evaluation with particular reference to border cave. South African Journal of Science, 74, 409-419.
Beck, L. R., and St. George, R. (1983). The alleged cultural bias of PAT: reading comprehension and reading vocabulary tests. New Zealand Journal of Educational Studies, 18, 32-47.
Beiser, M. and Gotowiec, A. (2000). Accounting for native/non-native differences in IQ scores. Psychology in the Schools, 37, 237-252.
Benton, D., and Cook, R. (1991). Vitamin and mineral supplements improve intelligence scores and concentration of six year old children. Personality and Individual Differences, 12, 1151-1158.
Benton, D., and Roberts, G. (1988). Effect of vitamin and mineral supplementation on intelligence in a sample of school children. The Lancet, 1, 140-143.
Веге, M. (1924). A comparative study of the mental capacity of children of foreign parentage. Columbia University Contributions to Education, no. 154.
Berlioz, L. (1955). Etude des progressive matrices faite sur les Africains de Douala. Bulletin du Centre Etude Recherce Psychotechnique, 4, 33-44.
Berry, J. W. (1966). Temne and Eskimo perceptual skills. International Journal of Psychology, 1, 207-229.
Расовые различия в интеллекте
265
Berry, J. W. (1971). Ecological and cultural factors in spatial perceptual development. Canadian Journal of Behavioral Science, 3, 324-336.
Bhatnagar, J. (1970). Immigrants at School. London: Cornmarket.
Bhogle, S., and Prakash, I. J. (1992). Performance of Indian children on the Colored Progressive Matrices. Psychological Studies, 37, 178-181.
Biesheuvel, S. (1949). Psychological tests and their application to non-European peoples. In G. B. Jeffrey (Ed.). Yearbook of Education. New York: Evans.
Binford, L. R. (1980). Yellow smoke and dogs’ tails: hunter gatherer settlement systems and archeological site formation. American Antiquity, 45, 4-20.
Binford, L. R. (1985). Human ancestors: changing views of their behavior. Journal of Anthropological Archaeology, 4, 292-327.
Binnie-Dawson, J. L. (1984). Bio-social and endocrine bases of spatial ability.
Psychologia, 27, 129-151.
Biraben, J. N. (1980). An essay concerning mankind’s evolution. Population, 4,1-13.
Black Peoples. (1978). Cause for Concern: West Indian Pupils in Redbridge. Ilford: Black Peoples Progressive Association.
Blatchford, P., Burke, J., Farquhar, C., Plewis, L, and Tizard, B. (1985). Educational achievement in the infant school: the influence of ethnic origin, gender and home on entry skills. Educational Research, 27, 52-60.
Bleakley, J. W. (1961). The Aborigines of Australia. Brisbane: Jacaranda Press.
Bleichrodt, N., and Bom, M. P. (1994). A meta-analysis of research on iodine and its relationship to cognitive development. In J. B. Stanbury (Ed.). The Damaged Brain of Iodine Deficiency. New York: Cognizant Communication Corp.
Bleichrodt, N., Drenth, P. J. D., and Querido, A. (1980). Effects of iron deficiency on motor and psychomotor abilities. American Journal of Physical Anthropology, 53, 55-67.
Bleichrodt, N., Garcia, I., Rubio, C., Morreale, D. E., and De Escobar, M. (1987). Developmental disorders with severe iodine deficiency. In B. S. Hetzel, J. T. Dunn and J. B. Stanbury (Eds.). The Prevention and Control of Iodine Deficiency. Amsterdam: Elsevier.
Blumenbach, J. F. (1776). De generis humani uarietate nativa liber. Goettingen: Vandenhoek.
Blumenschine, R. J. (1989). Man the scavenger. Archaeology. July, 26-32.
Bodmer, W. F. and Cavalli-Sforza, L. L. (1976). Genetics, Evolution and Man. San Francisco: Freeman.
Bohannon, A. D., Fillenbaum, G. G., and Pieper, C. F. (2002). Relationship of гасе/ ethnicity and blood pressure to change in cognitive function. Journal of the American Geriatrics Society, 50, 424—429.
Boissiere, М., Knight, J. B. and Sabot, R. H. (1985). Earnings, schooling, ability and cognitive skills. American Economic Review, 75, 1016-1030.
Boivin, M. J. and Giordani. B. (1993). Improvements in cognitive performance for school children in Zaire following an iron supplement and treatment for intestinal parasites. Journal of Pediatric Psychology, .249-264.
Boivin, M. J., Giordani, B., and Bomfeld, B. (1995). Use of the tactual performance test for cognitive ability testing with African children. Neuropsychology, 9, 409-417.
Boivin, M. J., Giordani, B., Crist, C. L., and Chounramany, C. (1996). Validating a cognitive ability testing protocol with Lao children for community development applications. Neuropsychology, 11, 588-599
Borkowski, J. G. and Krause, A. (1983). Racial differences in intelli-gence: the importance of the executive system. Intelligence, 7, 379-395.
Bouchard, T. J. (1993). The genetic architecture of human intelligence. In P. A. Vernon (Ed.). Biological Approaches to the Study of Human Intelligence. Norwood, NJ: Ablex.
Bouchard, T. J. (1998). Genetic and environmental influences on adult intelligence and special mental abilities. Human Biology, 70, 257-279.
Bourdier, G. (1964). Utilisation et nouvel etalonnage du P. M. 47. Bulletin de Psychologies 235, 39-41.
266
Ричард Линн
Bowd, A. D. (1973). A cross-cultural study of the factorial composition of mechanical aptitude. Canadian Journal of Behavioral Science, 5, 13-23.
Bowdler, S. and Ryan, L. (1987). Southeast Tasmania. In: D. J. Mulvaney and J. P. White (Eds.). Australians to 1788. Sydney, Australia: Fairfax, Syme and Weldon.
Boyd, W. (1950). Genetics and the Races of Man. Boston: Little, Brown.
Brace, C. L. and Hinton, R. J. (1981). Oceanic tooth size variation as a reflection of biological and cultural mixing. Current Anthropology, 22, 549-569.
Brace, C. L. (1999). An anthropological perspective on «race» and intelligence: the non- clinal nature of human cognitive capabilities. Journal of Anthropological Research, 55, 245-264.
Bradshaw, J. L. (1997). Human Evolution: A Neuropsychological Perspective. Hove, UK: Psychology Press.
Brandon, P. R., Newton, B. J., and Hammond, O. W. (1987). Children’s mathematics achievement in Hawaii: differences favoring girls. American Educational Research Journal, 24, 437-461.
Brandt, T. (1978). Growth dynamics of low birth weight infants with emphasis on the perinatal period. In: Human Growth, vol. 2. F. Falkner and J. M. Tanner (Eds.), pp 557-617. New York: Plenum Press.
Broca, P. (1861). Sur le volume et la forme du cerveau suivant les individus et suivant les races. Bulletin Societe de Anthropologie Paris, 2, 139-207, 301-321,441-446.
Brody, N. (1992). Intelligence. San Diego, GA: Academic.
Brody, N. (2000). Comment. In: Novaris Foundation Symposium. The Nature of Intelligence. New York: Wiley.
Brody, N. (2003). Jensen’s genetic interpretation of racial differences in intelligence: critical evaluation. In H. Nyborg (Ed.). The Scientific Study of General Intelligence. Amsterdam: Elsevier.
Broer, M. (1996). Rasch-homogene Leistungstests (3DW, WMT). im Kulturvergleich Chile-Osterreich: Erstellung einer spanischen Version ein-er Testbatterie und deren interkulturellen Validierung in Chile [Rasch-ho-mogeneous achievement tests (3DW, WMT) in a cross-cultural comparison of Chile and Austria: development of a Spanish test-battery version and its cross-cultural validation in Chile]. Unpublished M. Sc. thesis, University of Vienna.
Broman, S. H., Nichols, P. L., and Kennedy, W. A. (1975). Preschool IQ. New York: J. Wiley.
Broman, S. H., Nichols, P. L., Shaughnessy, P., and Kennedy, W. (1987). Retardation in Young Children. Hillsdale, New Jersey: Lawrence Erlbaum.
Brown, R.T., Reynolds, C. R., and Whitaker, J. S. (1999). Bias in mental testing since Bias in Mental Testing. School Psychology Quarterly, 14, 208-238
Browne, D. B. (1984). WISC-R scoring patterns among Native Americans of the northern plains. White Cloud Journal, 3, 3-16.
Bruce, D. W, Hengeveld, М., and Radford, W. C. (1971). Some Cognitive Skills in Aboriginal Children in Victorian Primary Schools. Victoria: Australian Council for Educational Research.
Bruner, F. G. (1908). The hearing of primitive peoples. Archives of Psychology, no. 11.
Buj, V. (1981). Average IQ values in various European countries. Personality and Individual Differences, 2, 168-169.
Bunn, H. T, and Stanford, С. B. (2001). Conclusions. In С. B. Stanford and H. T. Bunn (Eds.). Meat Eating and Human Evolution. Oxford, UK: Oxford University Press.
Burg, B., and Belmont, I. (1990). Mental abilities of children from different cultural backgrounds. Journal of Cross-Cultural Psychology, 21, 90-108.
Butterworth, B. (1999). The Mathematical Brain. London: Macmillan.
Byrne, R. W. (2002). The primate origins of human intelligence. In R. J. Sternberg and J. K. Kaufman (Eds.) The Evolution of Intelligence. Mahway, NJ: Lawrence Erlbaum.
Cairns, J., Overbaugh, J. and Miller, S. (1988). The Origin of Mutants. Nature, 335, 142-145.
Расовые различия в интеллекте
267
Callan, V. J. (1986). Australian Minority Groups. Sydney: Harcourt Brace Javanovich.
Carr, A. (1993). Twenty years a growing: a research note on gains in the intelligence test scores of Irish children over two decades. Irish Journal of Psychology, 14, 576-582.
Carroll, J. B. (1993). Human Cognitive Abilities. Cambridge: Cambridge University Press.
Cashdan, E. (2001). Ethnic diversity and its environmental determinants: effects of climate, pathogens, and habitat diversity. American Anthropologist, 103, 968-992.
Cattell, R. B. (1951). The fate of national intelligence — test of a thirteen year prediction. Eugenics Review, 17, 136-148.
Cattell, R. B. (1971). Abilities: Their Structure, Growth and Action. Boston: Houghton Mifflin.
Cavalli-Sforza, L. L. (2000). Genes, Peoples and Languages. New York: North Point.
Cavalli-Sforza, L. L., and Bodmer, W. (1971). The Genetics of Human Populations. San Francisco: Freeman.
Cavalli-Sforza, L. L., Menozzi, P., and Piazza, A. (1994). The History and Geography of Human Genes. Princeton, NJ: Princeton University Press.
Cazden, С. B., and John, V. P. (1971). Learning in American Indian children. In M. L. Wax, S. Diamond, and F. O. Gearing (Eds.). Anthropological Perspectives on Education. New York: Basic Books.
Ceci, S. J. (1991). How much does schooling influence general intelligence and its
cognitive components? A reassessment of the evidence. Developmental Psychology, 27, 703-722.
Chagnon, N. A. (1983). Yanomamo: The Fierce People. New York: Holt, Rinehart &C Winston.
Chaim, H. H. (1994). Is the Raven Progressive Matrices Valid for Malaysians? Unpublished.
Chakraborty, R., Kamboh, M. L, Nwankwo, М., and Ferrell, R. E. (1992). Caucasian genes in American blacks: new data. American Journal of Human Genetics, 50, 145-155.
Chambers, С. M. and Grantham-McGregor, S. M. (1986). Research note:pattems of mental development among middle class Jamaican children. Journal of Child Psychology and Psychiatry, 27, 117-123.
Chan, J., Eysenck, H. J., and Lynn, R. (1991). Reaction time and intelligence among Hong Kong children. Perceptual and Motor Skills, 72, 427-433.
Chan, J. and Lynn, R. (1989). The intelligence of six year olds in Hong Kong. Journal of Biosocial Science, 21, 461-464.
Chan, J. and Vernon, P. E. (1988). Individual differences among the peoples of China. In Irvine, S. H. and Berry, J. W. Human Abilities in Cultural Context. Cambridge, UK: Cambridge University Press.
Chandra, S. (1975). Some patterns of response on the Queensland Test. Australian Psychologist, 10, 185-191.
Chavaz, A., Matrinez, B., and Soberanes, B. (1995). Effect of malnutrition on infant development. In N. S. Scrimshaw (Ed.). Longitudinal Community Based Studies of the Impact of Early Malnutrition on Child Health and Development. Boston, MA: INFDC.
Chen, C., Lee, S., and Stevenson, H. W. (1996). Long term prediction of academic achievement of American, Chinese and Japanese adolescents. Journal of Educational Psychology, 98, 750-759.
Cheung, P. (2003). Congo’s Pygmies accuse rebels and army of cannibalism; demand U.
N. genocide tribunal. Associated Press, May 22, 2003.
Chisholm, J. S. (1989). Biology, culture and the development of temperament — a Navajo example. In J. K. Nugent, B. L. Lester, and T. B. Brazelton (Eds ) The Cultural Context of Infancy. Norwood, NJ: Ablex.
Chopra, S. L. (1966). Family size and sibling position as related to measured intelligence and academic achievement. Journal of Social Psychology, 70 133-137.
Chorley, M. J., Chorley, K., Seese, N., Owen, M. J., Daniels, J., McGuffln, P., Thompson, L. A., Detterman, D. K., Benbow, C., Lubinski, D., Eley, Т., and Plomin, R. (1998). A
268
Ричард Линн
quantitative trait locus associated with cognitive ability in children. Psychological Science, 9, 159-166.
Chovioto, L., Aghini-Lombardi, R, and Vitti, P. (1994). The impact of iodine deficiency on neurological and cognitive development: the European experience. In J. B. Stan- bury (Ed.). The Damaged Brain of Iodine Deficiency. New York: Cognizant Communication Corporation.
Claassen, N. C. W. (1990). The comparability of General Scholastic Aptitude Test scores across different populations groups. South African Journal of Psychology, 20, 80-92.
Clark, E. A., and Hanisee, J. (1982). Intellectual and adaptive performance of Asian children in adoptive American settings. Developmental Psychology, 18, 595-599.
Clay, М. M. (1971). The Polynesian language skills of Maori and Samo-an school entrants. International Journal of Psychology, 6, 135-145.
Codd, J. A. (1972). Cultural factors in the cognitive development of Maori children. Delta, 11,26-36.
Codwell, J. E. (1947). A study of the kind and amount of change in motor function as the amount of Negro blood increases or decreases in the Negro-white hybrid. Ph. D. thesis, University of Michigan.
Cohen, M. N. (2002). An anthropologist looks at race and IQ testing. In J. M. Fish (Ed.). Race and Intelligence. Mahwah, NJ: Lawrence Erlbaum.
Cole, S. (1965). Races of Man. London: Her Majesty’s Stationery Office.
Coleman, J. S. (1966). Equality of Educational Opportunity. Washington, D.C.: U. S.
Office of Education.
Colom, R., Garcia, L. R, Juan-Espinoza, М., and Abad, R (2002). Null differences in general intelligence: evidence from the WAIS-R. Spanish Journal of Psychology, 5, 29-35.
Colom, R., Juan-Espinosa, М., Abad, R, and Garcia, L. R (2000). Negligible sex differences in general intelligence. Intelligence, 28, 57-68.
Colom, R., and Lynn, R. (2004). Testing the developmental theory of sex differences in intelligence on 12-18 year olds. Personality and Individual Differences, 2004, 36, 75-82.
Coombs, L. M. (1958). The Indian Child Goes to School. Washington, D.C.: U. S. Department of the Interior, Bureau of Indian Affairs.
Coon, C. S., Gam, S. М., and Birdsell, J. B. (1950). Races. Springfield, 111: Thomas.
Coon, C. S. (1962). The Origin of Races. New York, Knopf.
Coren, S. (1994). The Intelligence of Dogs: Canine Consciousness and Capability. New York: Free Press.
Costenbader, V., and Ngari, S. M. (2000). A Kenya standardisation of the Coloured Progressive Matrices. School Psychology International, 22, 258-268.
Cottereau-Reiss, P., and Lehalle, H. (1998). Comparaison des performances d’enfants Kanak et d’enfants francais dans une situation de jugements de morphismes: structuration spatiale et moulin a vent. Archives de Psychologies 66, 3-21.
Counter, S. A., Buchanan, L. H., Rosas, H. D., and Ortega, F. (1998). Neurocognitive effects of chronic lead intoxication in Andean children. Journal of Neurological Sciences, 160, 47-53.
Court, J. H. (1983). Sex differences in performance on Raven’s Progressive Matrices: a review. Alberta Journal of Educational Research, 29, 54-74.
Сох, М. V., Perara, J., and Fan, X. U. (1998). Children’s drawing ability in the UK and China. Psychologia, 41, 171-182.
Сох, М. V., Koyasu, М., Hranamu, H., and Perara,]. (2001). Children’s human figure drawings in the UK and Japan: the effects of age, sex and culture. British Journal of Developmental Psychology, 19, 275-292.
Craig,]. D. (1974). A Study of the Education and Abilities of Young Australian Male Adults. Australian Department of Labor Research Report. Canberra: Government Printing Service.
Cra vioto, J. (1966). Malnutrition and behavioural development in the pre-school child. In National Academy of Sciences. Pre-school Child Malnutrition. Washington, D.C.
Расовые различия в интеллекте
269
Cravioto, J., Birch, Н. G., Licardie, Е., Resales, L., and Vega, L. (1969). Ecology of growth and development in a Mexican pre-industrial community. Report 1: Method and findings from birth to one month of age. Monographs of the Society for Research in Child Development, vol. 35, no. 129.
Crawfurd, J. (1863). On the antiquity of man, from the evidence of language. Transactions of the Ethnographic Society of London, 1, 170-181.
Cropley, A. J., and Cardey, R. M. (1975). Contact with the dominant culture and
cognitive competence in Canadian Indians and whites. Canadian Journal of Behavioral Science, 7, 328-238.
Cundick, B.P., Gottfredson, D.K., and Willson, L. (1974) Changes in scholastic achievement and intelligence of Indian children enrolled in a foster placement program. Developmental Psychology, 10, 815-820.
Curr, E. M. (1887). The Australian Race. Melbourne: Government Printer.
Cutler, R. G. (1976). Evolution of longevity in primates. Journal of Human Evolution, 5, 169-202.
Dague, P., Garelli, М., and Lebettre, A. (1964). Recherches sur l'echelle de maturite mentale de Colombia. Revue de Psychologie Applique, 14, 71-96.
Daley, Т. C., Whaley, S. E., Sigman, M. D., Espinosa, M. P., and Neuman, C. (2003). IQ on the rise: the Flynn effect in rural Kenyan children. Pychological Science, 14, 215-219.
Dan, L., Yu, J., Vandenberg, S. G., Yuemei, Z., and Caihong, T. (1990). Report on Shanghai norms of the Chinese translation of the Wechsler Intelligence Scale for Children-Revised. Psychological Reports, 67, 531-541.
Dasen, P. R., de Lacey, P. R., and Seagrim, G. N. (1973). Reasoning ability in adopted and fostered Aboriginal children. In G. E. Kearney, P. R. de Lacey, and G. R. Davidson (1973). The Psychology of Aboriginal Australians. New York: Wiley.
Davidson, G. R. (1974). Linguistic determinants of choice reaction time among
Aborigines and white Australians. Journal of Cross-Cultural Psychology, 5,199-210.
Davies, М., and Hughes, A. G. (1927). An investigation into the comparative intelligence and attainments of Jewish and non-Jewish school children. British Journal of Psychology, 18, 134-146.
Dawkins, A., and Snyder, R. (1977). Disadvantaged junior high school students compared with norms of the Seashore tests. Journal of Research on Music Education, 20, 438-444.
Dawkins, R. (1988). The Blind Watchmaker. London, UK: Penguin.
Dawkins, R., and Krebs, J. R. (1979). Arms races within and between races. Proceedings of the Royal Society, 205B, 489-511.
Darwin, C. (1868). The Variation of Animals and Plants under Domestication. London: Murray.
Dawson, I., Colder, R. Y, and Jonas, E. G. (1982). Birthweight by ges-tational age and its effect on perinatal mortality in white and in Pun-jabi births. British Journal of Obstetrics and Gynaecology, 89, 896-899.
Dayi, E., Okuyan, М., and Tan, U. (2002). Predictability of hand skill and cognitive abilities from craniofacial width in right and left-handed men and women: relation of skeletal structure to cerebral function.International journal of Neuroscience, 112, 383-412.
Deary, I. J. (2000). Looking Down on Human Intelligence. Oxford, UK: Oxford University Press.
De Jong, M. J. (1988). Ethnic origin and educational careers in Holland Netherlands Journal of Sociology, 24, 65-75.
De Jong, M. J., and van Batenburg, T. A. (1984). Etnische heromst, intelligentic en schoolkeuzeadvies. Pedagogische Studien, 61, 362-371.
De Lacey, P. R. (1971a). Classificatory ability and verbal intelligence among high-contact aboriginal children and low socio-economic status white Australian children. Journal of Cross-Cultural Psychology, 3, 393-396.
De Lacey, P. R. (1971b). Verbal intelligence, operational thinking and environment in part —Aboriginal children. International Journal of Psychology, 23, 145-149.
270
Ричард Линн
De Lacey, P. R. (1972). A relationship between classificatory ability and verbal intelligence. International Journal of Psychology, 1, 243-246.
De Lacey, P. R. (1976). Lifeways and cognitive performance in Australia and the United States. In G. E. Kearney and D. W. McElwain (Eds.). Aboriginal Cognition. Canberra: Australian Institute of Aboriginal Studies.
De Lemos, М. M. (1969). The development of the concept of conservation in Aboriginal children. International Journal of Psychology, 4, 255-269.
De Lemos, М. M. (1979). Aboriginal Students in Victoria. ACER Research Monograph No. 3. Melbourne: ACER.
De Lemos, М. M. (1989). Standard Progressive Matrices: Australian Manual Camberwell, Vic.: Australian Council for Educational Research.
De Maeyer, E., and Adiels-Tegman, M. (1985). The prevalence of anaemia in the world. World Health Statistical Bulletin, 38, 302-316.
Dennis, W. (1957). Performance of Near Eastern children on the Draw-a-Man test. Child Development, 28, 427-430.
Dennis, W., and Najarian, P. (1963). Development under environmental handicap. In W. Dennis (Ed.). Readings in Child Psychology. Englewood Cliffs: Prentice-Hall.
Dent, G. R. (1937). An investigation into the applicability of certain perforomance and other mental tests to Zulu children. In E. G. Malherbe (Ed.). Report of New Education Conference. Cape Town, p. 456.
De Silva, H. A., and Gunatilake, S. B. (2002). Mini Mental State examination in
Sinhalese: a sensitive test to screen for dementia in Sri Lanka. International Journal of Geriatric Psychiatry, 17, 134-139.
Diamond, J. (1997) Guns, Germs and Steel. New York; Random House.
Dickenson, L., Hobbs, A., Kleinberg, S. М., and Martin, P. J. (1975). The Immigrant School Leaver. Windsor, UK: National Foundation for Educational Research.
Dillehay, T. D., and Collins, М. B. (1988). Early cultural evidence from Monte Verde in Chile. Nature, 332, 150-152.
Dixon, E. J. (1999). Bones, Boats and Bison. Albuquerque, NM: University of New Mexico Press.
Dobbing, J., and Smart, J. L. (1974). Vulnerability of developing brain and behaviour. British Medical Bulletin, 30, 164-168.
Dodge, P. R., Palkes, H., Fierro-Benitez, R., and Ramirez, I. (1969). Effect on intelligence of iodine in oil administration to young adult Andean children. In J. B. Stanbury (Ed.). Endemic Goitre. Washington, D.C.: Pan American Health Organization.
Dolan, С. V, and Hamaker, E. (2001). Investigating black-white differences in psychometric IQ: multi-group factor analysis and a critique of the method of correlated vectors. In F. Columbus (Ed). Advances in Psychological Research, vol. 6, Huntington: Nova Science.
Domstreich, M. D. (1973). Food habits of early man and the balance between hunting and gathering. Science, 179, 306.
Drennan, M. R. (1937). A Short Course on Physical Anthropology. Cape Town: Mercantile-Atlas.
Driessen, G. W. (1997). Islamic primary schools in The Netherlands: the pupil’s
achievement levels, behavior and attitudes and their parents’ cultural backgrounds. Netherlands1 Journal of Social Sciences, 33, 42-75.
Drinkwater, B. A. (1976). Visual memory skills of medium contact Aboriginal children. Australian Journal of Psychology, 28, 37-44.
Du Bois, W. E. B. (1939). Black Folk: Then and Now. New York: Henry Holt.
Du Chateau, P. (1967). Ten point gap in Maori aptitudes. National Education, 49, 157-158.
Dunbar, R. I. (1992). Neocortex size as a constraint on group size in primates. Journal of Human Evolution, 20, 469^493.
Dunn, J. T. (1994). Societal implications of iodine deficiency and the value of its prevention. In J. B. Stanbury (Ed.). The Damaged Brain of Iodine Deficiency. New York: Cognizant Communication Corporation.
Расовые различия в интеллекте
271
Dunn, L. М. (1988). Bilingual Hispanic Children on the U. S. Mainland. Honolulu: Dunn Educational Services.
Dunbrow, E. H., Schaefer, B. A., and Jimerson, S. (2002). Diverging a paths in rural Caribbean children. School Psychology International, 155-168.
Dyck, M. J. (1996). Cognitive assessment in a multicultural society: comment on Davidson. Australian Psychologist, 31, 66-69.
Edwards, L. D. (1970). Malnutrition and disease in pre-school Aboriginal children in the Walgett area of N. S. W. Medical Journal of Australia, 2, 1007-1010.
Edwards, L. D., and Craddock, L. J. (1973). Malnutrition and intellectual de velopment. Medical journal of Australia, 5 May, 880-884.
Eells, W. C. (1933). Mental ability of the native races of Alaska. Journal of Applied Psychology, 17,417-438.
Elkin, A. P. (1964). The Australian Aborgines. Sydney: Angus and Robertson.
El-Mneizel, A. F. (1987). Development and psychometric analysis of a Jordanian adaptation of the Kaufman Assessment Battery for Children. Ph.D. dissertation, University of Alabama.
Evans, E. М., Schweingruber, H., and Stevenson, H. W. (2002). Gender differences in interest and knowledge acquisition: the United States, Taiwan, and Japan. Sex Roles, 47, 153-167.
Eyferth, K. (1961). Leistungen verschiedener Gruppen von Besatzungskindem im
Hamburg Wechsler Intelligenz Test fur Kinder. Archive fur die Gesamte Psychologie, 113, 222-241.
Eysenck, H. J. (1971). Race, Intelligence and Education. London: Temple Smith.
Eysenck, H. J. (1981). Intelligence: The Battle for the Mind. London: Pan. Eysenck, H. J. (1995). Genius. Cambridge, UK: Cambridge University Press.
Eysenck, H. J. (1998). Intelligence: A New Look. New Brunswick, NJ: Transaction.
Eysenck, H. J., and Schoenthaler, S. J. (1997). Raising IQ with vitamins and minerals. In R. J. Sternberg and E. Grigorenko (Eds.). Intelligence, Heredity and Environment. Cambridge, UK: Cambridge University Press.
Fahmy, M. (1964). Initial exploring of the intelligence of Shilluk children. Vita Humana, 1, 164-177.
Fahrmeier, E. D. (1975). The effect of school attendance on intellectual development in Northern Nigeria. Child Development, 46, 281-285.
Falconer, D. S. (1960). Introduction to Quantitative Genetics. London: Longman.
Farron, O. (1966). The test performance of coloured children. Educational Research, 8, 42-57.
Fatouros, M. (1972). The influence of maturation and education on the development of abilities. In L. J. Cronbach and P. J. Drenth (Eds). Mental Tests and Cultural Adaptation. The Hague: Mouton.
Faverge, J. М., and Falmagne, J. C. (1962). On the interpretation of data in intercultural psychology. Psychologia Africana, 9, 22-96.
Feldman, D. H. (1971). Map understanding as a possible crystallizer of cogni tive structures. American Educational Research Journal, 8,485-500.
Fergusson, D. М., Lloyd, М., and Horwood, L. J. (1991). Family ethnicity, social background and scholastic achievement — an eleven year longitudinal study. New Zealand Journal of Educational Studies, 26, 49-62.
Fergusson, D. М., and Horwood, L. J. (1997). Sex differences in educational achievement in a New Zealand birth cohort. New Zealand Journal of Educational Studies, 32, 83-95.
Fernandez, M. (2001). A study of the intelligence of children in Brazil. Mankind Quarterly, 42, 17-21.
Femandez-Ballesteros, R., Juan-Espinoza, М., Colom, R., and Calero, M. D. (1997). Contextual and personal sources of individual differences in intelligence. In J. S. Carlson (Ed.). Advances in Cognition and Educational Practice. Greenwich, CT: JAI Press.
272
Ричард Линн
Pick, М. L. (1929). Intelligence test results of poor white, native (Zulu), coloured and Indian school children and the social and educational implications. South African Journal of Science, 26, 904-920.
Fierro-Benitez, R. (1994). Impact of iodine deficiency on development in the Andean world. In J. B. Stanbury (Ed.). The Damaged Brain of Iodine Deficiency. New York: Cognizant Communication Corporation.
Fierro-Benitez, R., Cazar, R., and Sandoval, H. (1989). Early correction of iodine
deficiency and late effects on psychomotor capabilities and migration. In De Long, G. R., Robbins, J., and Condliffe, P. G. (Eds.). Iodine and the Brain. New York: Plenum.
Fierro-Benitez, R., Ramirez, L, Estrella, E., and Stanbury, J. B. (1972). Effect of iodine correction in early fetal life on intelligence quotient. In J. B. Stanbury and R. L. Kroc, R. L. (1972). Human Development and the Thyroid Gland: Relation to Endemic Cretinism. New York: Plenum.
Fierro-Benitez, R., Ramirez, I., Estrella, E., and Stanbury, J. B. (1974). Effect of iodine in intellectual development in an area of endemic goitre. In J. R. Dunn and G. A. Medeiros-Neto (Eds.). Endemic Goiter and Cretinism: Continuing Threats to World Health. Washington, D.C.: Pan American Health Organization.
Fish, J. M. (ed.) (2002). Race and Intelligence. Mahvvah, NJ: Lawrence Erlbaum.
Flaherty, M. (1997). The validity of tests of visuo-spatial skills in cross-cultural studies. Irish Journal of Psychology, IS, 439-412.
Flaherty, М., and Connolly, M. (1996). Visual memory skills in Japanese and Caucasians. Perceptual and Motor Skills, 82, 1319-1329.
Flaughter, R. L. (1971). Project Access research report no. 2. Princeton, NJ: Educational Testing Service.
Flores, М. B., and Evans, G. T. (1972). Some differences in cognitive abilities between selected Canadian and Filipino students. Multivariate Behavioral Research, 7, 175-191.
Flynn, J. R. (1980). Race, IQ and Jensen. London: Routledge & Kegan Paul.
Flynn, J. R. (1984). The mean IQ of Americans: massive gains 1932 to 1978. Psychological Bulletin, 95, 29-51.
Flynn, J. R. (1987). Massive IQ gains in 14 nations: what IQ tests really measure. Psychological Bulletin, 101, 171-191.
Flynn, J. R. (1991). Asian Americans: Achievement Beyond IQ. Hillsdale, NJ: Lawrence Erlbaum.
Flynn, J. R. (1998). WAIS-111 and WISC-III IQ gains in the United States from 1992 to 1995: how to compensate for obsolete norms. Perceptual and Motor Skills, 86, 1231-1239.
Foley, R. (1987). Another Unique Species. New York: Wiley.
Fowler, H. L. (1940). Psychological tests on natives in the north west of Western Australia. Australian Journal of Science, 2, 124-127.
Fox, E., Sitompul, A., and Van Schaik, C. P. (1999). Intelligent tool use in wild Sumatran orangutans. In S. T. Parker, H. L. Miles, and R. W. Mitchell (Eds.). The Mentality of Gorillas and Orangutans. Cambridge:Cambridge University Press.
Fraser, A. (1995). The Gypsies. Cambridge, MA: Blackwell.
Freedman, L., Blumer, W. E, and Lofgren, M. (1991). Endocranial capacity of Western Australian Aboriginal crania: comparisons and association with stature and latitude. American Journal of Physical Anthropology, 84, 399-405.
Frydman, М., and Lynn, R. (1989). The intelligence of Korean children adopted in Belgium. Personality and Individual Differences, 10, 1323-1326.
Gaddes, W. H., McKenzie, A., and Baensley, R. (1968). Psychometric intelligence and spatial imagery in two northwest Indian and two white groups of children. Journal of Social Psychology, 75, 35-42.
Caller, J. R., Ramsey, R, and Forde, V. (1986). A follow-up study of the influence of early malnutrition on subsequent development. Nutrition and Behavior, 3, 211-222.
Caller, J. R., Ramsey, R, Solimano, G., Lowell, W. E. and Mason, E. (1983). Influence of early malnutrition on subsequent behavioural development. Journal of the American Academy of Child Psychiatry, 22, 8-15.
Расовые различия в интеллекте
273
Galton, R (1869). Hereditary Genius. London: Macmillan.
Galton, R (1888). Head growth in students at the University of Cambridge. Nature, 38, 14-15.
Garrett, H. E. (1945). Facts and interpretations regarding race differences. Science, 102, 404-406.
Garrett, H. E. (1961). The equalitarian dogma. Perspectives in Biology and Medicine, 4, 480-484.
Garth, T. R. (1931). Racial Psychology. New York: Macmillan.
Garth, T. R., and Smith, O. D. (1937). The performance of full-blood Indians on language and non-language intelligence tests. Journal of Abnormal and Social Psychology, 32, 376-381.
Geary, D. C., Hamson, С. O., Chen, G-R, Liu, R, Hoard, М. K., and Salthouse, T. A.
. Computational and reasoning abilities in arithmetic: cross-generational change in China and the United States. Psychonomic Bulletin and Review, 4, 425-430.
Geary, D. C., Liu, R, Chen, G-R, Salts, S. J., and Hoard, М. K. (1999). Contributions of computational fluency to cross-national differences in arithmetical reasoning abilities. Journal of Educational Psychology, 91, 716-719.
Geelhoed, G. W., and Downing, D. (1994). Goiter and cretinism in the Uele Zaire endemia. In J. B. Stanbury (Ed.). The Damaged Brain of Iodine Deficiency. New York:
Cognizant Communication Corporation.
Geist. V. (1978). Life Strategies, Human Evolution and Environmental Design. New York: Springer-Verlag.
Georgas, J. G., and Georgas, C. (1972). A children’s intelligence test for Greece. In
L. J. Cronbach and P. J. D. Drenth (Eds.). Mental Tests and Cultural Adaptation. The Hague: Mouton.
Georgas, J., Weiss, L. G., van der Vijver, R J., and Saklofske, D. H. (2003). A cross- cultural analysis of the WISC-III. In J. Georgas, L. G. Weiss, R van der Vijver and D. H. Saklofske (Eds.). Culture and Children’s Intelligence. Amsterdam: Academic Press.
Gill. P.. and Byrt, E. (1973). The Standardization of Ravens Progressive A Matrices and the Mill Hill Vocabulary Scale for Irish School Children Aged 6-12 Years. University College, Cork: MA Thesis.
Ginsburg, H. P., Choi, E., Lopez, L. S., Netley, R., and Chao-Yuan, C. (19975 Happy birthday to you: early mathematical thinking of Asian, South American and U. S. children. In T. Nunes and P. Bryant (Eds.). Learning and Teaching Mathematics: An International Perspective. Hove, UK Psychology Press.
Giordani, B., Boivin, M. J., Opel, B., Nseyila, D. N., and Lauer, R. E. (1996). Use of the K-ABC with children in Zaire. International Journal of Disability, Development and Education, 43, 5-24.
Glewwe, P., and Jaccoby, H. (1992). Estimating the Determinants of Cognitive Achievement in Low Income Countries. Washington, D.C.: World Bank.
Globerson, T. (1983). Mental capacity and cognitive functioning: developmental and social class differences. Developmental Psychology, 19, 225-230.
Goa, Y., Qian, M. and Wang, D. (1998). Changes in intelligence over 10 years (in Chinese). Chinese Journal of Clinical Psychology, 6, 185-186.
Gobineau, A. de (1853). essai sur Tinegalite des races humaines. Paris: Didot.
Godman, A. (1964). The attainments and abilities of Hong Kong primary IVpupils: a first study. Hong Kong: Hong Kong University Press.
Goodall, J. (1986). The Chimpanzees of Gombe: Patterns of Behavior. Cambridge, MA: Cambridge University Press.
Goodenough, F. L. (1926a). The Measurement of Intelligence by Drawings. New York: World Books.
Goodenough, F. L. (1926b). Racial differences in the intelligence of school children. Journal of Experimental Psychology, 9, 388-397.
Goodenough, F. L., and Harris, D. B. (1950). Studies in the psychology of children’s drawings: 11, 1928-49. Psychological Bulletin, 27, 396-433.
274
Ричард Линн
Goosens, G. (1952а). Etalonnage du Matrix 1947 de J. C. Raven. Revue Beige de Psychologie et de Pedagogie, 14, 74-80.
Goosens, G. (1952b) Une application du test d’intelligence de R.B
Gordon, E. E. (1980). Developmental music aptitudes among inner city primary children.
Bulletin Council for Research in Music Education, 63, 25-30.
Gott, B. (2002). Fire making in Tasmania: absence of evidence is not evidence of absence.
Current Anthropology, 43, 650-655.
Gottfredson, L. S. (1997). Editorial: Mainstream science on intelligence. Intelligence, 24, 13-24.
Gottfredson, L. S. (2005). What if the hereditarian hypothesis is true? Psychology, Public Policy, and Law, 11, 311-319.
Gould, R. A. (1969). Subsistence behaviour among the Western Desert Aborigines of Australia. Oceania, 39, 253-274.
Gould, S. J. (1978). Morton’s ranking of races by cranial capacity. Science, 200, 503-509. Gould, S. J. (1981; 1996). The Mismeasure of Man. New York, Norton. Grantham-McGregor, S, М., and Hawke, W. A. (1971). Developmental assessment of Jamaican infants. Developmental Medicine and Child Neurology, 13, 582-589. Grantham-McGregor, S. М., Powell, C., Walker, S. P., and Himes, J. H. (1991).
Nutritional supplementation, psychosocial stimulation and the mental development of stunted children: the Jamaica study. The Lancet, 338, 1-5.
Grantham-McGregor, S. М., Powell, C., Walker, S. P., Chang, S., and Fletcher, P. (1994). The long-term follow-up of severely malnourished children who participated in an intervention program. Child Development, 65, 428-439.
Grantham-McGregor, S. М., Walker, S. P., and Powell, C. (1994). Methodological approaches used in Kingston, Jamaica, to determine the effect of nutrition and stimulation on child development. In J. B. Stanbury (Ed.). The Damaged Brain of Iodine Deficiency. New York: Cognizant Communication.
Graves, J. L. (2002). An anthropologist looks at race and IQ testing. In J. M. Fish (Ed.).
Race and Intelligence. Mahwah, NJ: Lawrence Erlbaum.
Greenfield, P. M. (1998). The cultural evolution of IQ. In U. Neisser (Ed.). The Rising Curve: Long Term Gains in IQ and Related Matters. Washington, D.C.: American Psychological Association.
Grieve, K. W, and Viljoen, S. (2000). An exploratory study of the use of the Austin maze in South Africa. South African Journal of Psychology, 30, 14-18.
Grigorenko, E. L., and Sternberg, R. J. (2001). Analytical, creative and practical
intelligence as predictors of self-reported adaptive functioning: a case study in Russia. Intelligence, 29, 57-73.
Grine, R E., and Kay, R. R (1988). Early hominid diets from quantitative image analysis of dental microwear. Nature, 333, 765-768.
Groves, C. P. (1991). Genes, genitals and genius: the evolutionary ecology of race. In P. O’Higgins (Ed.). Human Biology: An Integrative Science. Perth, Australia: Center for Human Biology.
Guenole, N., Englert, P., and Taylor, P. J. (2003). Ethnic group differences in cognitive ability test scores within a New Zealand applicant sample. New Zealand Journal of Psychology, 32, 49-54.
Guidon, N., and Delibrias, G. (1986). Carbon-14 dates point to man in the Americas 32,000 years ago. Nature, 321, 769.
Gupta, G. C., and Gupta, S. (1966). Norms for Raven’s Colored Progressive Matrices. Manus, 13, 87-89.
Gupta, S. (1991). Effects of time of day and personality on intelligence test scores.
Personality and Individual Differences, 12, 1227-1231.
Gustafsson, J. E. (1984). A unifying model of the structure of mental abilities. Intelligence, 8, 179-203.
Расовые различия в интеллекте
275
Guthke, J., and Al-Zoubi, А. (1987). Kulturspezifische Differenzen in den Coloured Progressive Matrices (CPM) und in einer Lemtestvariante der CPM. Psychologie in Erziehung und Unterricht, 34, 306-311.
Hadidjaja, P., Bonang, E., Suyardi, A., Abidin, A. N., Ismid, I. S., and Margono, S. S. (1998). The effect of intervention methods on nutritional status and cognitive function of primary school children infected with ascaris lumbri-coides. American Journal of Tropical Medicine, 59, 791-795.
Halpem, D. (2000). Sex Differences in Cognitive Abilities. Mahwah, N. J.: Lawrence Erlbaum.
Halsey, A. H. (1972). Educational Priority. Volume 1. London: HMSO.
Hamers, J. H. М., Hessels, M. G. P., and Pennings, A. H. (1996). Learning potential of ethnic minority children. European Journal of Psychological Assessment, 12, 183-192.
Hammer, М. E, Redd, A. J., and Wood, E. T. (2000). Jewish and Middle Eastern non- Jewish populations share a common pool of Y-chromosome biallelic haplotypes. Proceedings of the National Academy of Sciences, 97, 6769-6774.
Hanushek, E. A., and Kimko, D. D. (2000). Schooling, labor force quality, and the growth of nations. American Economic Review, 90, 1184-1208.
Harker, R. K. (1978). Achievement and ethnicity: environmental deprivation or cultural difference. New Zealand Journal of Educational Studies, 13, 107-124.
Harris, S. A. (1977). Milingimbi Aboriginal Learning Contexts. Ph.D. dissertation, University of New Mexico, Albuquerque, New Mexico.
Hart, J. A. (1965). A study of Cognitive Capacity of a Group of Australian Aboriginal Children. M. A. thesis, University of Queensland.
Harvey, P. H., and Glutton-Brock, Т. H. (1985). Life history variation in primates. Evolution, 39, 559-581.
Haught, B. F. (1934). Mental growth of the southwest Indians. Journal of Applied Psychology, 18,419-433.
Havighurst, R. J., and Hilkevitch, R. R. (1944). The intelligence of Indian children measured by a performance scale. Journal of Abnormal and Social Psychology, 39,419-433.
Hayden, B. (1981). Subsistence of Modem Hunter Gathers. In R. O. S. Harding and G. Telela (Eds.). Omnivorous Primates. New York: Columbia Univ. Press.
Heim, A. W. (1968). AH4 Group Test of General Intelligence Manual. Slough,UK:
National Foundation for Educational Research.
Helms-Lorenz, М., Van de Vijver, F. J. R., and Poortinger, Y. P. (2003). Cross-cultural differences in cognitive performance and Spearman’s hypothesis. Intelligence, 31, 9-29.
Henneberg, M. (1983). Trends in cranial capacity and cranial index in sub-Saharan Africa during the Holocene. American Journal of Human Biology, 5, 473-479.
Henneberg, M. (1984). Comment on Beals Dodd and Smith. Current Anthropology, 25, 321-322.
Henneberg, М., Budnik, A., Pazacka, М., and Puch, A. E. (1985). Head size, body size and intelligence: introspecific correlations in Homo sapiens. Homo, 36, 207-218.
Herrnstein, R. J., and Murray, C. (1994). The Bell Curve: Intelligence and Class Structure in American Life. New York: Free Press.
Hertzig, М., Birch, H. G., Richardson, S. A., and Tizard, J. (1972). Intellectual levels of school children malnourished during the first two years of life. Pediatrics, 49, 814-824.
Hetzel, B. S. (1994). Historical development of the concepts of brain-thyroid relationships. In J. B. Stanbury (Ed.). The Damaged Brain of Iodine Deficiency. New York:
Cognizant Communication Corporation.
Heyneman, S. P., and Jamison, D. T. (1980). Student learning in Uganda. Comparative Education Review, 24, 207-220.
Hirszfeld, L., and Hirszfeld, H. (1919). Essai d’application des methods au probleme des races. Anthropologies 29, 505-537.
Ho, H. — Z., Baker, L. A., and Decker, S. N. 1988. Covariation between intelligence and speed of cognitive processing: genetic and environmental influences. Behavior Genetics, 18, 247-261.
10*
276
Ричард Линн
Но, К. — С, Roessmann, U., Straumfjord, J. V., and Monroe, G. (1980). Analysis of brain weight: 1 & 11. Adult brain weight in relation to sex, race and age. Archives of Pathology and Laboratory Medicine, 104, 635-639; 640-645.
Holding, P. A., Taylor, H. G., Kazungu, S. D., and Mkala, T. (2004). Assessing cognitive outcomes in a rural African population: development of a neuro-psychological battery in Kilifi district, Kenya. Journal of the International Neuropsychological Society, 10, 246260.
Horn, J. L. (1991). Measurement of intellectual capabilities: a review of theory. In K. S. McGrew, J. K. Werder, and R. W. Woodcock (Eds.). Woodcock-Johnson Technical Manual. Chicago: Riverside.
Houghton, V. (1966). Intelligence testing of West Indian and English children. Race, 8, 147-156.
Howell, R. J., Evans, L., and Downing, L. N. (1958). A comparison of test scores from the 16-17 year age group of Navajo Indians with standardisation norms from the WAIS. Journal of Social Psychology, 47, 355-359.
Hsu, С. — C. (1971). Chinese children’s responses to Raven’s Colored Progressive Matrices. Journal of the Formosan Medical Association, 70, 579-593.
Hsu, С. — C., See, R., and Lin, C-C. (1973). Assessment of learning potential of Chinese children with Raven’s Standard Progressive Matrices. Journal of the Formosan Medical Association, 72, 658-670.
Hsu, С. — C. (1976). The learning potential of first graders in Taipei city as measured by Raven’s Coloured Progressive Matrices. Acta Pediatrica Sinica, 17, 262-274.
Humphreys, L. G. (1988). Trends in level of academic achievement of blacks and other minorities. Intelligence, 12, 231-260.
Hunkin, V. (1950). Validation of the Goodenough Draw-a-Man test for African children. Journal for Social Research, 1, 52-63.
Hunter, B., and Schwab, R. G. (1998). The determinants of indigenous educational outcomes. Canberra: Australian National University,
Hunter, J. E., and Hunter, R. F. (1984). Validity and utility of alternative predictors of job performance. Psychological Bulletin, 96, 72-98.
Hunter, W. S., and Sommermier, E. (1922). The relation of the degree of Indian blood to score on the Otis Intelligence Test. Journal of Comparative Psychology, 2, 257-275.
ILEA. (1967). The Education of Immigrant Pupils in Primary Schools. London: Inner London Education Authority.
Isaac, G. (1978). The food sharing behavior of protohuman hominoids. Scientific American. 238, 90-109.
Ishikuma, T, Moon, S., and Kaufman, A. S. (1988). Sequential-simultaneous analysis of Japanese children’s performance on the Japanese McCarthy scales. Perceptual and Motor Skills, 66, 355-362.
Jahoda, G. (1970). Supernatural beliefs and changing cognitive structures among Ghanaian university students. Journal of Cross-Cultural Psychology, 1,115-130.
Jaworowska, A., and Szustrowa, T. (1991). Podrecznik Do Testu Matryc Ravena. Warsaw: Pracownia Testow Psychologicznych.
Jencks, C, (1972). Inequality. London: Penguin.
Jencks, G,and Phillips,M. (1998). The Black-WhiteTest Score Gap. Washington, D.C.: Brookings Institution.
Jensen, A. R. (1969). How much can we boost IQ and scholastic achievement? Harvard Educational Review, 39, 1-123.
Jensen, A. R. (1972). Genetics and Education. London: Methuen.
Jensen, A. R. (1973). Educability and Group Differences. London: Methuen.
Jensen, A. R. (1974). How biased are culture-loaded tests? Genetic Psychology Monographs, 90, 185-244.
Jensen, A. R. (1977). Cumulative deficit in IQ of blacks in the rural south. Developmental Psychology, 13, 184-191.
Расовые различия в интеллекте
277
Jensen, A. R. (1980). Bias in Mental Testing. London: Methuen.
Jensen, A. R. (1981). Obstacles, problems and pitfalls in differential psychology. In:
S. Scarr (Ed). Race, Social Class and Individual Differences in IQ. Hillsdale, NJ: Lawrence Erlbaum.
Jensen, A. R. (1982). Reaction time and psychometric g. In H. J. Eysenck (Ed.). A Model for Intelligence. Berlin: Springer-Verlag.
Jensen, A. R. (1993). Spearman’s hypothesis tested with chronometric information- processing tasks. Intelligence, 17, 47-77.
Jensen, A. R. (1998). The g Factor. Westport, CT: Praeger.
Jensen, A. R., and Figueroa, R. A. (1975). Forward and backward digit span interaction with race and IQ:predictions from Jensen’s theory. Journal of Educational Psychology, 67, 882-893.
Jensen, A. R., and Inouye, A. R. (1980). Level I and Level II abilities in Asian, white and black children. Intelligence. 4, 41-49.
Jensen, A. R., and Reynolds, C. R. (1982). Race, social class and ability patterns on the WISC-R. Personality and Individual Differences, 3, 423-438.
Jensen, A. R., and Rohwer, W. D. (1970). An experimental analysis of learning abilities in culturally disadvantaged children. Washington, D.C.: Office of Economic Opportunity.
Jensen, A. R., Saccuzzo, D. P., and Larson, G. E. (1988). Equating the standard and advanced forms of the Progressive Matrices. Educational and Psychological Measurement, 48, 1091-1095.
Jensen, A. R., and Whang, P. A. (1993). Reaction times and intelligence: a comparison of Chinese and Anglo-American children. Journal of Biosocul Science, 25, 397-411.
Jensen, A. R., and Whang, P. A. (1994). Speed of accessing arithmetic facts in long term memory: a comparison of Chinese-American and Anglo-American children. Contemporary Educational Psychology, 19, 1-12.
Jerison, H. (1973). Evolution of the Brain and Intelligence. New York, Academic Press.
Jerison, H. (2000). The evolution of intelligence. In R. J. Sternberg (Ed.). Handbook of Intelligence. Cambridge: Cambridge University Press.
Jinabhai, С. C., Taylor, М., Rangongo, N. J., Mkhize, S., Anderson, S., Pillay, B. J., and Sullivan, K. R. (2004). Investigating the mental abilities of rural primary school children in South Africa. Ethnicity and Health, 9, 17-36.
Johnson, D. L., Johnson, C. A., and Price-Williams, D. (1967). The Draw-a-Man test and Raven Progressive Matrices performance of Guatemalan boys and Ladino children. Revista Interamericana de Psicologial,M3-\51.
Johnson, G. B. (1948). Musical talent and the Negro. Musical Supervisors Journal, 15, 81-96.
Jordheim, G. D., and Olsen, I. A. (1963). The use of a non-verbal test of intelligence in the trust territory of the Pacific. American Anthropologist, 65, 1122-1125.
Joseph, A., and Murray, V. F. (1951) Chamorros and Carolinians of Saipan. Westport, CT: Greenwood.
Jurgens, H. W., Aune, I. A., and Pieper, U. (1990). International Data on Anthropometry. Geneva, Switzerland: International Labor Office.
Kabo, V. (1985). The origins of the food-producing economy. Current Anthropology, 26, 601-616.
Kagitcibasi, C. (1972). Application of the D-48 test in Turkey. In L. J. Cronbach and P. J.
Drenth (Eds.). Mental Tests and Cultural Adaptation. The Hague: Mouton.
Kaniel, S., and Fisherman, S. (1991). Level of performance and distribution of errors in the Progressive Matrices test: a comparison of Ethiopian immigrant and native Israeli adolescents. International Journal of Psychology, 26, 25-33.
Kaplan, G. J., Fleshman, J. K., Bender, T. R., and Clark, P. S. (1973). Long term effects of otitis media: a ten-year cohort study of Alaskan Eskimo children. Pediatrics, 52, 577-585.
Kaszycka, K. A., and Strkalj, G. (2002). Anthropologist’s attitudes towards the concept of race: the Polish sample. Current Anthropology, 43, 329-335.
278
Ричард Линн
Kaszycka, К. A., and Strzalko, J. (2003). Race — still an issue for physical anthropology? Results of Polish studies seen in the light of the U. S. findings. American Anthropologist, 105, 114-122.
Kaufman, A. S., and Doppelt, J. E. (1976). Analysis of WISC-R standardization data in terms of the stratification variables. Child Development, 47, 165-171.
Kaufman, A. S., and Kaufman, N. L. (1983). KABC Interpretive Manual. Circle-Pines: MN: American Guidance Service.
Kaufman, A. S., and Lichtenberger, E. (2002). Assessing Adolescent and Adult Intelligence. Boston: Allyn & Bacon.
Kaufman, A. S., McLean, J. E., Ishikuma,T., and Moon, S. B. (1989). Integration of literature on the intelligence of Japanese children and analysis of the data from a sequential-simultaneous perspective. School Psychology International, 10, 173-183.
Kaufman, A.S., and Wang, J. J. (1992). Gender, race and educational differences on the K-BIT at ages 4-90. Journal of PsychCoeducational Assessment, 10, 219-229.
Kaufman, J. C, McLean, J. E., Kaufman, A. S., and Kaufman, N. L. (1994). White-black and white-Hispanic differences on fluid and crystallized abilities by age across the 11 to 94 year range. Psychological Reports, 75, 1279-1288.
Kearins, J. M. (1981). Visual spatial memory in Australian Aboriginal children of desert regions. Cognitive Psychology, 3, 434-460.
Keith, A. (1922). The dawn of national life. In J. A. Hammerton (Ed.). Peoples of All Nations. London: Amalgamated Press.
Kelly, M. (1977). Papua New Guinea and Piaget — an eight-year study. In P. R. Dasen (Ed). Piagetian Psychology. New York: Gardner Press.
Kendall, I. M. (1976). The predictive validity of a possible alternative to the Classification Test Battery. Psychologia Africana, 16, 131-146.
Kendall, I. M. (1977). Some observations concerning reasoning styles of black South African workers: perceptual versus conceptual considerations. Psychologia Africana, 17, 1-29.
Kennedy, W. A., and Lindner, R. S. (1964). A normative study of the Goodenough Draw- a-Man Test on Southeastern Negro elementary school children. Child Development, 35, 33-62.
Kennedy, W. A., Van der Reit, V, and White, J. C. (1963). A normative study of intelligence and achievement of Negro schoolchildren in the south-eastern United States. Monographs of the Society for Research in Child Development, 28, no. 6.
Kimura, D. (2002). Sex hormones influence human cognitive pattern Neuroendocrinology Letters, Special Issue Supplement 4, vol. 23.
Klaatsch, H. (1908). The skull of the Australian aboriginal. Reports of the Pathology Laboratory Lunacy Department NSW, 1, 43-167.
Kleinfeld, J. (1971). Visual memory in village Eskimo and urban Caucasian children. Arctic, 24, 132-138.
Klekamp,J.,Reidel, A., Harper, C, and Kretschmann, H. J. (1 987). Mor-phometric study of the postnatal growth of the visual cortex of Austra-lian aborigines and Caucasians. Journal of Brain Research, 35, 541-548.
Kline, C. L., and Lee, N. (1972). A transcultural study of dyslexia: analysis of language disabilities in Chinese children simultaneously learning to read and write in English and in Chinese. Journal of Social Education, 6, 9-26.
Klingelhofer, E. L. (1967). Performance of Tanzanian secondary school pupils on the Raven Standard Progressive Matrices test. Journal of Social Psychology, 72, 205-215.
Knapp, P. A., and Seagrim, G. N. (1981). Visual memory in Australian Aboriginal children and children of European descent. International Journal of Psychology, 16, 213-231.
Kohler, W. (1927). The Mentality of Apes. New York: Vintage.
Kozulin, A. (1998). Profiles of immigrant students’ cognitive performance on Raven’s Progressive Matrices. Perceptual and Motor Skills, 87, 1311-1314.
Расовые различия в интеллекте
279
Kramer, R. A., Allen, L., and Gergen, P. J. (1995). Health and social characteristics and children’s cognitive functioning: results from a national cohort. American Journal of Public Health, 85, 312-318.
Krantz, G. S. (1980). Climatic Races and Descent Groups. North Quincy, Mass: Christopher.
Kuhnen, U., Roeder, В. H. U., Ahah, A. A., Upmeyer, B. S. A., and Zakaria, S. (2001). Cross-cultural variations in identifying embedded figures. Journal of Cross-Cultural Psychology, 32, 365-371.
Kunce, J., Rankin, L. S., and Clement, E. (1967). Maze performance and personal, social and economic adjustment of Alaskan natives. Journal of Social Psychology, 73, 37-45.
Kurth, von E. (1969). Erhohung der Leistungsnormen bei den farbigen progressiven Matrizen. Zeitschrift fiir Psychologic, 177, 85-90.
Kuttner, R. E. (1962). Prehistoric technology and human evolution. Mankind Quarterly, 3, 71-87.
Kyostio, О. K. (1972). Divergence among school beginners caused by different cultural influences. In L. J. Cronbach and P. J. Drenth (Eds.). Mental Tests and Cultural Adaptation. The Hague: Mouton.
Lai, T. J., Guo, Y. L., Guo, N. W., and Hsu, С. C. (2001). Effects of prenatal exposure to polychlorinated biphenyls on cognitive development in children: a longitudinal study in Taiwan. British Journal of Psychiatry, 178, 49-52.
Lancer, I., and Rim, Y. (1984). Intelligence, family size and sibling age spacing.
Personality and Individual Differences, 5, 151-157.
Landes, D. S. (1998). The «Wealth and Poverty of Nations: Why Some Are So Rich and Some So Poor. New York: W. W. Norton & Company.
Landman, J. (1988). Appendix to the Manual of the junior South African Individual Scales. Pretoria: Human Sciences Research Council.
Laosa, L. М., Swartz, J. D., and Diaz-Guerrero, R. (1974). Perceptual-cognitive and personality development of Mexican and Anglo-American children as measured by human figure drawings. Developmental Psychology, 10, 131-139.
Laroche, J. L. (1959). Effets de repetition du Matrix 38 sur les resultats d’enfants Katangais. Bulletin du Centre d'Etudes et Recherches Psychotechniques, 1, 85-99.
Laros, J. A., and Telegren, P. J. (1991). Construction and Validation of the Son-R NonVerbal Test of Intelligence. Groningen: Wolters-Noordhoff.
Lathan, М. C. (1974). Protein-calorie malnutrition in children and its relation to psychological development and behavior. Physiological Reviews, 54, 541-565.
Latouche, G. L., and Dormeau, G. (1956). La foration professionelle rapide en Afrique Equatoriale Framaise. Brazzaville: Centre d’Etudes des Problems du Travail.
Lee, R. B. (1968). What hunters do for a living. In Man the Hunter. In R. B. Lee and I. Devore (Eds.), Chicago, Aldine Press.
Lenski, R., and Mittler, J. E. (1993). The directed mutation controversy and neo- Darwinism. Science, 259, 188-194.
Lesser, G. S., Fifer, G., and Clark, D. H. (1965). Mental abilities of children from different social-class and cultural groups. Monographs of the Society for Research in Child Development, 30, no.4.
Levin, M. (1994). Comments on the Minnesota Transracial Adoption study. Intelligence, 19, 13-20.
Levin, M. (1997). Why Race Matters. Westport, CT: Praeger. Levinson, В. M. (1957). The intelligence of applicants for admission to Jewish day schools. Jewish Social Studies, 19, 129-140.
Lewis, B. (1990) Race and Slavery in the Middle East. New York: Oxford University Press.
Li, D., Jin,Y., Vandenberg, S. G., Zhu,Y., and Tang, C. (1990). Report on Shanghai norms for the Chinese translation of the Wechsler Intel-ligence Scale for Children — Revised. Psychological Reports, 67, 531-541.
Li, X., Sano, H., and Merwin, J. C. (1996). Perception and reasoning abilities among
American, Japanese and Chinese adolescents. Journal of Adolescent Research, 11, 173-193.
280
Ричард Линн
Lieberman, L., and Reynolds, L. T. (1996). Race: the deconstruction of a scientific concept. In L. T. Reynolds and L. Lieberman (Eds.). Race and Other Misadventures: Essays in Honor of Ashley Montagu. Dix Hills: General Hall.
Lieblich, A., and Kugelmas, S. (1981). Patterns of intellectual ability of Arab school children in Israel. Intelligence, 5, 311-320.
Lieblich, A., Ninio, A., and Kugelmas, S. (1972). Effects of ethnic origin and parental SES on WPPSI performance of pre-school children in Israel. Journal of Cross Cultural Psychology, 3, 159-168.
Lim, Т. K. (1994). Gender-related differences in intelligence: application of confirmatory factor analysis. Intelligence, 19, 179-192.
Linn, М. C., and Petersen, A. C. (1986). A meta-analysis of gender differences in spatial ability: implications for mathematics and science achievement. In J. S. Hyde and М. C, Linn (Eds.). The Psychology of Gender. Baltimore: Johns Hopkins University Press.
Linnaeus, C. (1758). Systerna naturae sistens animate Sveciae regni. Holmae: Salvius.
Lipovechaja, N. G., Kantonistowa, N. S., and Chamaganova, T. G. (1978). The role of heredity and of intellectual function. Medidskie Probleing Formirouaniga Livenosti, 27, 48-59.
Little, A. (1975). Performance of children from ethnic minority backgrounds in primary schools. Oxford Review of Education, 1, 117-135.
Liu, J., Raine, A., Venables, P. H., Dalais, C., and Mednick, S. A. (2003). Malnutrition at age 3 years and lower cognitive ability at age 11 years. Archives of Pediatric and Adolescent Medicine, 157, 593-600.
Livingstone, F. B. (1962). On the non-existence of the human races. Current Anthropology, 3, 279-281.
Llanos, Z. M. (1974). El funcionamiento intelectuel de los nicos en las zones marginales de Lima. Montevideo, Uruguay: Institute Americano de Nino.
Lloyd, E, and Pidgeon, D. A. (1961) An investigation into the effects of couching on verbal and non-verbal tests with European, Indian and African children. British Journal of Educational Psychology, 31, 145-151.
Loehlin, J. C., Lindzey, G., and Spuhler, J. N. (1975). Race Differences in Intelligence. San Francisco, CA: Freeman.
Lombardi, T. P. (1970). Psycholinguistic abilities of Papago Indian school children. Exceptional Children, 36, 485-493.
Lorenzo, J. L., and Mirambell, L. (1986). Tlapacoya: 35,000: Anos de Historic del Lago de Chaleo. Mexico City: Instituto Nacional de Antropologia e Historia.
Lovegrove, M. N. (1966). The scholastic achievement of European and Maori children. New Zealand Journal of Educational Studies, 1, 15-39.
Lubinski, D. (2000). Scientific and social significance of assessing individual differences. Annual Review of Psychology, 51, 405 -444.
Lucas, A., Morley, R., Cole, T. J., Lister, G., and Leeson-Payne, C. (1992). Breast milk and subsequent intelligence quotient in children born pre-term. The Lancet, 339, 261-264.
Lucas, A., Morley, R., Cole, T. J. (1998). Randomised trial of early diet in pre-term babies and later intelligence quotient. British Medical Journal, 317, 1481-1487.
Lynn, R. (1977a). The intelligence of the Japanese. Bulletin of the British Psychological Society, 30, 69-72.
Lynn, R. (1977b). The intelligence of the Chinese and Malays in Singapore. Mankind Quarterly, 18, 125-128.
Lynn, R. (1979). The social ecology of intelligence in the British Isles. British Journal of Social and Clinical Psychology, 18, 1-12.
Lynn, R. (1980). The social ecology of intelligence in the France. British Journal of Social and Clinical Psychology, 19, 325-331.
Lynn, R. (1987). The intelligence of the Mongoloids: a psychometric, evolutionary and neurological theory. Personality and Individual Differences, 8, 813-844.
Расовые различия в интеллекте
281
Lynn, R. (1988). Educational Achievement in Japan. London: Macmillan.
Lynn, R. (1990a). The role of nutrition in secular increases of intelligence. Personality and Individual Differences, 11, 273-285.
Lynn, R. (1990b). Differential rates of secular increase of five major primary abilities.
Social Biology, 38, 137-141.
Lynn, R. (1991 a). Race differences in intelligence: a global perspective. Mankind Quarterly, 31, 254-296.
Lynn, R. (1991b). The evolution of race differences in intelligence. MankindQuarterly, 32, 99-173.
Lynn, R. (1991c). Intelligence in China. Social Behavior and Personality, 19, 1-4.
Lynn, R. (1992). Sex differences on the Differential Ability Test in British and American adolescents. Educational Psychology, 12, 101-106.
Lynn, R. (1993). Nutrition and intelligence. In R A. Vernon (Ed.), Biological Approaches to the Study of Human Intelligence. Norwood, NJ: Ablex.
Lynn, R. (1994a). The intelligence of Ethiopian immigrant and Israeli adolescents.
International Journal of Psychology, 29, 55-56.
Lynn, R. (1994b). Sex differences in brain size and intelligence: a para-dox resolved.
Personality and Individual Differences, 17, 257-271.
Lynn, R. (1994c). Some reinterpretations of the Minnesota Transracial Adoption Study.
Intelligence, 19, 21-28.
Lynn, R. (1996). Racial and ethnic differences in intelligence in the United States on the Differential Ability Scale. Personality and Individual Differences, 20, 271-273.
Lynn, R. (1997). Intelligence in Taiwan. Personality and Individual Differnces, 22, 585-586. Lynn, R. (1998a). Sex differences on the Scottish standardisation sample of the WAIS-R.
Personality and Individual Differences, 24, 289-290.
Lynn, R. (1998b). In support of the nutrition theory. In U. Neisser (Ed.). The Rising Curve: Long Term Gains in IQ and Related Matters. Washington, D.C.: American Psychological Association.
Lynn, R. (1998c). Sex differences in intelligence: a rejoinder to Mackintosh. Journal of Biosocial Science, 30, 529-532.
Lynn, R. (1998d). New data on black infant precocity. Personality and Individual Differences, 25, 801-804.
Lynn, R. (1998e). Has the black-white intelligence difference in the United States been narrowing over time? Personality and Individual Differences, 25, 999-1002.
Lynn, R. (1999). Sex differences in intelligence and brain size: a developmental theory.
Intelligence, 27, 1-12.
Lynn, R. (2001). Intelligence in Russia. Mankind Quarterly, 42, 151-154.
Lynn, R. (2002). Racial and ethnic differences in psychopathic personality. Personality and Individual Differences, 32, 273-316.
Lynn, R. (2002a). Skin color and intelligence in African Americans. Population and Environment, 23, 365-375.
Lynn, R. (2004). The intelligence of American Jews. Personality and Individual Differences, 36, 201-207.
Lynn, R., Allik, J., Pullmann, H., and Laidra, J. (2002). A study of the IQ in Estonia.
Psychological Reports, 95, 611-612.
Lynn, R., Backhoff, E., and Contreras, L. A. (2005). Ethnic and racial differences on the Standard Progressive Matrices in Mexico. Journal of Biosocial Science, 37, 107-113. Lynn, R., Chan, J. W. C., and Eysenck, H. J. (1991). Reaction times and intelligence in Chinese and British children. Perceptual and Motor Skills, 72, 443-452.
Lynn, R., and Chan, P. W. (2003). Sex differences on the Progressive Matrices: some data from Hong Kong. Journal of Biosocial Science, 35, 145-154.
Lynn, R., and Dziobon, J. (1980). On the intelligence of the Japanese and other Mongoloid peoples. Personality and Individual Differences, 1, 95-96.
282
Ричард Линн
Lynn, R., and Hampson, S. L. (1986). The rise of national intelligence: evidence from Britain, Japan and the USA. Personality and Individual Differences, 7, 23-332.
Lynn, R., and Hampson, S. (1986a). The structure of Japanese abilities: an analysis in terms of the hierarchical model of intelligence. Current Pychological Research and Reviews, 4, 309-322.
Lynn, R., and Hampson, S. (1986b). Intellectual abilities of Japanese children: an
assessment of 2-8 year olds derived from the McCarthy Scales of Children’s Abilities. Intelligence, 10, 41-58.
Lynn, R., and Hampson, S. (1987). Further evidence on the cognitive abilities of the Japanese: data from the WPPSI. International Journal of Behavioral Development, 10, 23-36.
Lynn, R., Hampson, S., and Bingham, R. (1987a). Japanese, British and American
adolescents compared for Spearman’s g and for the verbal, numerical and visuo-spatial abilities. Psychologia, 30, 137-144.
Lynn, R., Hampson, S. L., and Iwawaki, S. (1987b). Abstract reasoning and spatial abilities among American, British and Japanese adolescents. Mankind Quarterly, 27, 397-434.
Lynn, R., Hampson, S., and Lee, M. (1988). The intelligence of Chinese children in Hong Kong. School Psychology International, 9, 29-32.
Lynn, R., Hampson, S., and Magee, M. (1984). Home background, intelligence,
personality and education as predictors of unemployment in young people. Personality and Individual Differences, 5, 549-558.
Lynn, R., and Harland, E. P. (1998). A positive effect of iron supplementation on the IQs of iron deficient children. Personality and Individual Differences, 24, 883-887.
Lynn, R., and Hattori, K. (1990). The heritability of intelligence in Japan. Behavior Genetics, 20, 545-546.
Lynn, R., and Holmshaw, M. (1990). Black-white differences in reaction times and intelligence. Social Behavior and Personality, 18, 299-308.
Lynn, R., and Irwing, P. (2004). Sex differences on the Progressive Matrices: a metaanalysis. Intelligence, 32, 481-98.
Lynn, R., and Kazlauskaite, V. (2002). A Study of the IQ in Lithuania. Perceptual and Motor Skills, 95,611-612.
Lynn, R., and Owen, K. (1994). Spearman’s hypothesis and test score differences between whites, Indians and blacks in South Africa. Journal of General Psychology, 121, 27-36.
Lynn, R., and Pagliari, C. (1994). The intelligence of American children is still rising. Journal of Biosocial Science, 26, 65-68.
Lynn, R., Pagliari, C., and Chan, J. (1988). Intelligence in Hong Kong measured for Spearman’s g and the visuospatial and verbal primaries. Intelligence, 12, 423-433.
Lynn, R., Plaspalanova, E., Stetinsky, D., and Tzenova, B. (1998). Intelligence in Bulgaria. Psychological Reports, 82, 912-914.
Lynn, R., Pullmann, H., and Allik, J. (2003). A new estimate of the IQ in Estonia. Psychological Reports, 97, 662-664.
Lynn, R., and Shigehisa, T. (1991). Reaction times and intelligence: a comparison of Japanese and British children. Journal of Biosocial Science, 23, 409-416.
Lynn, R., and Song, J. M. (1993). Sex differences in reaction times, decision times and movement times in British and Korean children. Journal of Genetic Psychology, 154, 209-313.
Lynn, R., and Song, J. M. (1994). General intelligence, visuospatial and ver bal abilities of Korean children. Personality and Individual Differences, 16, 363-364.
Lynn, R., Wilson, R. G., and Gault, A. (1989). Simple musical tests as measures of Spearman’s g. Personality and Individual Differences, 10, 25-28.
Lynn, R., and Vanhanen, T. (2002). IQ and the Wealth of Nations. Westport, CT: Praeger.
Mabey, C. (1981). Black British literacy: a study of reading attainment of London black children from 8 to 15 years. Educational Research, 23, 83-95.
Расовые различия в интеллекте
283
MacArthur, R. S. (1965). Mackenzie District Norming Project. Ottawa: Department of Northern Affairs.
MacArthur, R. S. (1967). Some cognitive abilities of Eskimo, white and Indian-Metis children aged 9 to 12 years. Canadian Journal of Behavioral Science, 1, 50-59.
MacArthur, R. S., Irvine, S. H., and Brimble, A. R. (1964). The Northern Rhodesia Mental Ability Survey. Lusaka: Rhodes Livingstone Institute.
MacDonald, K. (1994). A People That Shall Dwell Alone. Westport, CT: Praeger.
Mackintosh, N. J. (1977). Stimulus control: attention factors. In W. K. Honig and J. E. R. Staddon (Eds.). Handbook of Operant Behavior. Englewood Cliffs: Prentice-Hall.
Mackintosh, N. J. (1996). Sex differences and IQ. Journal ofBiosocial Science, 28, 559-572.
Mackintosh, N. J. (1998). IQ and Human Intelligence. Oxford: University Press.
Mackintosh, N. J., and Mascie-Taylor, C. G. N. (1985). The IQ question. In Education for All. Cmnd paper 4453. London: HMSO.
MacLean, A. W., and McGhie, A. (1980). The AH4 group test of intelligence applied in a Canadian high school sample. Canadian Journal of BehavioralScience, 12, 217-291.
MacNeish, R. S. (1976). Early man in the New World. American Scientist, 64, 316-327.
Maity, H. (1926). A report on the application of the Stanford adult tests to a group of college students. Indian Journal of Psychology, 1, 214-222.
Majumdar, P. K., and Nundi, P. C. (1971). Raven’s Standard Progressive Matrices in two different populations. Journal of the Indian Academy of Applied Psychology, 8, 30-33.
Manley, D. R. (1963). Mental ability in Jamaica. Social and Economic Studies, 12, 51-77.
Mann, V. A., Sasanuma, S., Sakuma, N., and Masaki, S. (1990). Sex differences in cognitive abilities: a cross cultural perspective. Neuropsychologia, 28, 1063-1077.
Marinkovich, R. I., Sparosvich, H. E, Santana, М. C. D., Game, J. H., Gomez, С. C., and Marinkovich, D. I. (2000). Estudio de la capacidad intellectuel (test de Matrices Progressivas de Raven) en escolares Chilerios de 5 a 18 acos. Revista de Psicologia General у Aplicada, 53, 5-30.
Martin, W. A. (1969). Word fluency: a comparative study. Journal of Genetic Psychology, 114, 253-262.
Martinelli, V., and Lynn, R. (2005). Sex differences on verbal and non-verbal abilities among primary school children in Malta. Unpublished.
Martorell, R., Malina, R. М., Castillo, R. O., Mendoza, F. S., and Pawson, I. G. (1988). Body proportions in three ethnic groups: children and youths 2-17 years in NHANES 11 and NHANES. Human Biology, 60, 205-222.
Martorell, R. (1997). Undernutrition during pregnancy and early childhood: consequences for cognitive and behavioural development. In М. E. Young (Ed.). Early Child Development: Investing in Our Children’s Future, Amsterdam: Elsevier.
Matarazo, J. D. (1972). The Measurement and Appraisal of Adult Intelligence. Baltimore: Williams and Wilkins.
Maugham, B., and Rutter, M. (1986). Black pupils’ progress in secondary schools: examination attainments. British Journal of Developmental Psychology, 4, 19-29.
McElwain, D. W, and Kearney, G. E. (1970). Queensland Test Handbook. Hawthorne, Australia: Australian Council for Educational Research.
McElwain, D. W, and Kearney, G. E. (1973). Intellectual development. In G. E. Kearney, P. R. de Lacey and G. R. Davidson (Eds.). The Psychology of Aboriginal Australians. New York: Wiley.
McFie, J., and Thompson, J. A. (1970). Intellectual abilities of immigrant children. British Journal of Educational Psychology, 40, 348-351.
McGrew, K. S., and Flanagan, D. P. (1998). The Intelligence Test Desk Reference. Oston: Allyn and Bacon.
McGurk, F. C. F. (1953a). On white and Negro test performance and socio-economic factors. Journal of Abnormal and Social Psychology, 37, 448-450.
McGurk, F. C. F. (1953b). Socio-economic status and culturally weighted test scores of Negro subjects. Journal of Abnormal and Social Psychology, 37, 276-277.
284
Ричард Линн
McIntyre, G. А. (1938). The Standardisation of Intelligence Tests in Australia. Melbourne: University Press.
McShane, D. A., and Plas, J. M. (1984). The cognitive functioning of American Indian children: moving from the WISC to the WISC-R. School Psychology Review, 17, 39-51.
Mehrotra, К. K. (1968). A comparative study of the WISC and Raven’s Progressive Matrices. Psychological Studies, 13, 47-50.
Mehryar, A. H., Shapurian, R., and Bassiri, T. (1972). A preliminary report on a Persian adaptation of Heim’s AH4 test. Journal of Psychology, 80, 167-180.
Mehryar, A. H., Tashakkori, A., Yousefi, R, and Khajavi, F. (1987). The application of the Goodenough-Harris Draw-a-Man test to a group of Iranian children in the city of Shiraz. British Journal of Educational Psychology, 57, 401-406.
Mellars, P., and Stringer, C. (1989). The Human Revolution. Edinburgh: Edinburgh University Press.
Mercer, J. R., and Lewis, J. F. (1984). System of Multicultural Pluralistic Assessment: Manual. San Antonio, TX: Psychological Софоration.
Michael, R. T. (2003). Children’s cognitive skill development in Britain and the United States. International Journal of Behavioral Development, 27, 396—408.
Miller, E. M. (1991). Climate and intelligence. Mankind Quarterly, 32, 127-131.
Miller, E. M. (1996). The evolution of Australoid and Amerindian intelligence. Mankind Quarterly, 37, 149-186.
Miller, E. M. (2005). Geographical Centrality As an Explanation for Racial Differences in Intelligence. Mankind Quarterly (to appear)
Miron, M. (1977). A validation study of a transferred group intelligence test. International Journal of Psychology, 12, 193-205.
Misawa, G., Motegi, М., Fujita, K., and Hattori, K. (1984). A comparative study of intellectual abilities of Japanese and American children on the Columbia Mental Maturity Scale. Personality and Individual Differences, 5, 173-181.
Modiano, N. (1962). Mental testing among Tzeltal and Tzotzil children. Proceedings of the 35 th International Congress of Americanists. Mexico City.
Mohan, V. (1972). Raven’s Standard Progressive Matrices and a verbal test of general mental ability. Journal of Psychological Researches, 16, 67-69.
Mohan, V., and Kumar, D. (1979). Performance of neurotics and stables on the Standard Progressive Matrices. Intelligence, 3, 355-368.
Mohanty, A. K., and Babu, N. (1983). Bilingualism and metalinguistic ability among Kond tribals in Orissa, India. Journal of Social Psychology, 121, 15-22.
Montagu, A. (1945a). Man’s Most Dangerous Myth: The Fallacy of Race. New York; Columbia University Press.
Montagu, A. (1945b). Intelligence of northern Negroes and southern whites in the First World War. American Journal of Psychology, 58, 161-188.
Montagu, A. (1999). (Ed.). Race and IQ. Expanded Edition. New York: Oxford University Press.
Montie, J. E., and Pagan, J. F. (1988). Racial differences in IQ: item analysis of the Stanford-Binet at 3 years. Intelligence, 12, 315-332.
Moon, S. B. (1988). A Cross Cultural Study of the Kaufman Assessment Battery for Children with Korean Children. Ph.D. thesis, University of Alabama.
Morant, G. M. (1927). A study of Australian and Tasmanian skulls based on previously studied measurements. Biometrika, 19, 417-440.
Morton, S. G. (1849). Observations on the size of the brain and families of man. Proceedings of the Academy of Natural Sciences Philadelphia, 4, 221-224.
Movies, E. W, and Wolins, M. (1973). Group care and intellectual development. Developmental Psychology, 4, 370-380.
Murray, C. (1998). Income Inequality and IQ. Washington, D.C.: AEI Press.
Murray, C. (2003). Human Accomplishment. New York: Harper Collins.
Расовые различия в интеллекте
285
Murray, L. S. (1983). Nutritional Status and Development of St. Lucian Preschool Children. M.Sc. Thesis. UWI, Mona.
Nagoshi, С. Т., and Johnson, R. C. (1986). The ubiquity of g. Personality and Individual Differences, 7, 201-208.
Nardi, N. (1948). Studies in intelligence of Jewish children. Jewish Education, 19, 41-45.
Nathawat, S. S., and Puri, P. (1995). A comparative study of MZ and DZ twins on Level I and Level II mental abilities and personality. Journal of the Indian Academy of Applied Psychology, 21, 545-546.
National Center for Education Statistics. (2003). National Assessment of Educational Progress. Washington, D.C.
National Center for Health Statistics. (1979). Caloric and selected nutrient values for persons 1-74 years of age. Vital Health Statistics, No. 209. Hyattsville, MD.
Nei, М., and Roychoudhury, A. K. (1993). Evolutionary relationships of human populations on a global scale. Molecular Biology and Evolution, 10, 927-943.
Neisser,U. (1998). Intelligence: knowns and unknowns. American Psychologist, 51, 77—
101.
Nell, V. (2000). Cross-Cultural Neuropsychological Assessment. Mahwah, NJ: Lawrence Erlbaum.
Nelson, W. E. (1933). Mitchell-Nelson Textbook of Pediatrics. Philadelphia: Saunders.
Nettle, D. (2003). Intelligence and class mobility in the British population. British Journal of Psychology, 94, 551-561.
Nisbett, R. E. (1998). Race, genetics and IQ. In C. Jencks and M. Phillips (Eds.). The Black-White Test Score Gap. Washington, D.C.: Brookings Institution Press.
Nissen, S., Machover, S., and Kinder, E. F. (1935). A study of performance tests given to a group of Native African Negro children. British Journal of Psychology, 25, 308-355.
Nkaya, H. N., Huteau, М., and Bonnet, J-P. (1994). Retest effect on cognitive
performance on the Raven Matrices in France and in the Congo. Perceptual and Motor Skills, 78, 503-510.
Norman, R. D., and Midkiff, K. L. (1955). Navaho children on Raven Progressive
Matrices and Goodenough Draw-a-Man tests. Southwestern Journal of Anthropology, 11, 129-136.
Notcutt, B. (1950). The measurement of Zulu intelligence. Journal of Social Research, 1, 195-206.
Nurcombe, B., de Lacey, P., and Walker, S. L. (1999). Children of the Dispossessed. Stamford, CT: Ablex.
Nurcombe, B., and Moffitt, P. (1973). Cultural Deprivation and Language Deficit. In G.
Kearney, P. R. de Lacey, and G. R. Davidson (Eds.). The Psychology of Aboriginal Australians. Sydney: John Wiley.
Nyborg, H. (2003). Sex differences in g. In H. Nyborg (Ed.). The Scientific Study of General Intelligence. Amsterdam: Elsevier.
Nyborg, H., and Jensen, A. R. (2000). Black-white differences on various psychometric tests: Spearman’s hypothesis tested on American armed services veterans. Personality and Individual Differences, 28, 593-599.
Office of the Surgeon General. (1968). Supplement to Health of the Army. Washington, D.C.: Dept, of the Army.
Ombredane, A., Robaye, E, and Robaye, E. (1952). Analyse des resultats d’une application experimentale du matrix 38 a 485 noirs Baluba. Bulletin Centre d’Etudes et Recherches Psychotechniques, 7, 235-255.
Ortar, G. (1952). Standardization of the Wechsler Test for Intelligence for children in Israel. Megamot, 4, 87-100.
Osborne, R. T. (1980). Twins: Black and White. Athens, GA: Foundation for Human Understanding.
Osborne, R. T, and Gregor, A. J. (1966). The heritability of visualization, perceptual speed and spatial orientation. Perceptual and Motor Skills, 23, 379-390.
286
Ричард Линн
Osborne, R. Т., and McGurk, F. С. (1982). The Testing of Negro Intelligence. Athens, GA: Foundation for Human Understanding.
Osmon, D.C., and Jackson, R. (2002). Inspection time and IQ: fluid or perceptual aspects of intelligence. Intelligence, 30, 119-127.
Owen, K. (1992). The suitability of Raven’s Progressive Matrices for various groups in South Africa. Personality and Individual Differences, 13, 149-159.
Packenberg, B., and Gundersen, H. J. (1997). Neurocortical nerurone number in humans: effects of age and sex. Journal of Comparative Neurology, 384, 312-320.
Paine, P., Dorea, J. G., Pasquali, L., and Monteiro, A. M. (1992). Growth and cognition in Brazilian school children: a spontaneously occurring intervention study. International Journal of Behavioral Development, 15, 169-183.
Pakkenberg, H., and Voigt, J. (1964). Brain weight of Danes. Acta Anatoma (Basel), 5, 297-307.
Pal, S., Shyam, R., and Singh, R. (1997). Genetic analysis of general intelligence g: a twin study. Personality and Individual Differences, 22, 779-780.
Parker, S. Т., and Gibson, K. R. (1977). A development model of the evolution of
language and intelligence in early hominids. The Behavioral Brain Sciences, 2, 367-408
Parker, S.Т., and McKinney, M. L. (1999). Origins of Intelligence: The Evolution of
Cognitive Development in Monkeys, Apes and Humans. Baltimore, MD: Johns Hopkins University Press.
Parra, E. J., Marcini, A., and Akey, J. (1998). Estimating African American admixture proportions by use of population-specific alleles. American Journal of Human Genetics, 63, 1839-1851.
Paul, S. M. (1985). The Advanced Raven’s Progressive Matrices: normative data for an American university population and an examinatioA of the relationship with Spearman’s g. Journal of Experimental Education, 54, 95-100.
Payne. J. (1974). Educational Priority: EPA Surveys and Statistics, vol. 2, London: HMSO.
Pearson, R. (1974). Introduction to Anthropology. New York: Holt, Rinehart and Winston.
Pearson, R. (1985). Anthropological Glossary. Malabar, FL: Kreiger.
Peoples, С. E., Fagan, J. E, and Drotar, D. (1995). The influence of race on 3 year old children’s performance on the Stanford-Binet Fourth Edition. Intelligence, 21, 69-82.
Persaud, G. (1972). The Performance of Two Samples of Primary School Children on Two Culture Free and Two Culture Bound Tests of Intelligence. University of Stockholm, Sweden: Institute of Applied Psychology.
Peters, L., and Ellis, E. N. (1970). An Analysis of WISC Profile Scores of Chinese-Canadian Children. Vancouver, BC: Board of School Trustees.
Peterson, J., and Lanier, L. H. (1929). Studies in the comparative abilities of whites and Negroes. Mental Measurement Monographs, No. 5.
Petrogiannis, K. S., Bardos, A. N., and Randou, E. (1999). Performance of Greek and American students on the Matrix Analogies Test. School Psychology International, 20, 233-238.
Philip’s. (1996). World Atlas. London: Chancellor.
Phillips, C. J. (1979). Educational under-achievement in different ethnic groups. Educational Research 21, 116-130.
Pickford, M. (1986). Major events in primate paleontology: possible support for climatic forcing models of evolution. Antropologia Contemporarea, 9, 89-94.
Piddington, М., and Piddington, R. (1932). Report of field work in Northwestern Australia. Oceania, 2, 327-328.
Pieke, F. N. (1988). The social position of the Dutch Chinese: an outline. China Information, 3, 12-23.
Pind, J., Gunnarsdottir, E. K., and Johannesson, H. S. (2003). Raven’s Standard Progressive Matrices: new school age norms and a study of the test’s validity. Personality and Individual Differences, 34, 375-386.
Plomin, R. (1994). Genetics and Experience: The Interplay between Nature and Nurture. Thousand Oaks, CA: Sage.
Расовые различия в интеллекте
287
Plomin, R., and Buss, D. (1973). Reflection-impulsivity and intelligence. Psychological Reports, 33, 726.
Plomin, R., DeFries, J. C., and McCleam, G. E. (1990). Behavioral Genetics. New York: Freeman.
Pollitt, E., Gorman, K. S., Engle, P. L., Martorell, R., and Rivera,J. (1993). Early Supplementary Feeding and Cognition. Monographs Society for Research in Child Development, 58, No. 235.
Pollitt, E., Hathirat, P., Kotchabhakdi, N., Missell, L., and Valyasevi, A. (1989). Iron deficiency and educational achievement in Thailand. American Journal of Clinical Nutrition, 50, 687-697.
Pons, A. L. (1974). Administration of Tests Outside the Cultures of their Origin. 26th Congress of the South African Psychological Association.
Popper, K. R. (1959). The Logic of Scientific Discovery. London: Methuen.
Poortinga, Y. (1971). Cross-cultural comparison of maximum performance tests: some methodological aspects and some experiments with simple auditory and visual stimuli. Psychologia Africana, Monograph Supplement No. 6.
Poortinga, Y. (1972). A comparison of African and European students in simple auditory and visual tasks. In L. J. Cronbach and P. D. Drenth (Eds.). Mental Tests and Cultural Adaptation. The Hague: Mouton.
Poortinga, Y, and Foden, I. M. (1975). A comparative study of curiosity in black and white South African students. Psychologia Africana, Monograph supplement No. 8.
Portenier, L. G. (1947). Abilities and interests of Japanese-American high school seniors. Journal of Social Psychology, 25, 53-61.
Porteus, S. D. (1917). Mental tests with delinquents and Australian Aboriginal children. Psychological Review, 24, 32-42.
Porteus, S. D. (1930). Race and social differences in performance tests. Genetic Psychology Monographs, 8, no. 2.
Porteus, S. D. (1931). The Psychology of a Primitive People. London: Edward Arnold.
Porteus, S. D. (1933a). Mentality of Australian Aborigines. Oceania, 4, 30-36.
Porteus, S. D. (1933b). Correspondence — the psychology of a primitive people. Oceania, 4, 30-36.
Porteus, S.D. (1937) Ethnic groups and the Maze Test. In R.E.Kuttner (Ed) Race and Modem Science. New York: Social Science Press.
Porteus, S. D. (1965). Porteus Maze Test. Palo Alto, С A: Pacific Books.
Porteus, S. D., and Babcock, H. (1926). Temperament and Race. Boston, MA: Badger.
Porteus, S. D., Brochner, S., Russell, J., and David, K. (1967). Age a factor in Australid mentality. Perceptual and Motor Skills, 25, 3-16
Porteus, S. D., and Gregor, A. J. (1963). Studies in intercultural testing.Perceptual and Motor Skills, 16, 705-724.
Post, R. H. (1962). Population differences in red and green colour visiondeficiency: review and a query on relaxation selection. Eugenics Review, 9, 131-146.
Pnfitera, A., Lawrence, L. G., and Saklofske, D. H. (1998). The WISC-III in context. In A. Prifitera and D. H. Saklofske (Eds.). (1998). WISC-III Clinical Use and Interpretation. San Diego, С A: Academic.
Prince, J. R. (1968). The effect of western education and science conceptualisation in New Guinea. British Journal of Educational Psychology, 38, 64-74.
Proctor, В. E., Kranzler, J. H., Rosenbloom, A. L., Martinez, V., and Guevara-Aguire, J. (2000). An initial investigation of validation of the Matrix Analogies Test-Expanded Form in Ecuador. Psychological Reports, 86, 445-453.
Pumfrey, P. D. (1983). The reading attainments of British children of parents of West Indian origin. Reading, 17, 111-124.
Rabinowitz, М. B., Wang, J-D., and Soong, W-T. (1991). Dentine lead and child intelligence in Taiwan. Archives of Environmental Health, 46, 351-360.
288
Ричард Линн
Rahman, A., Macbool, Е., and Zuberi, Н. S. (2002). Lead-associated deficits in stature, mental ability and behavior in children in Karachi. Annals of Tropical Paediatrics, 22, 301-311.
Raine, A., Reynolds, C, Venables, P. H., and Mednick, S. A. (2002). Stimulation seeking and intelligence: a prospective longitudinal study. Journal of Personality and Social Psychology, 82, 663-674.
Rao, S. N., and Reddy, I. K. (1968). Development of norms for Raven’s Coloured
Progressive Matrices on elementary school children. Psychological Studies, 13, 105-107.
Raudenbush, S. W, and Kasim, R. M. (1998). Cognitive skill and economic inequality: findings from the national adult literacy survey. Harvard Educational Review, 68, 33-68.
Raveau, F. H. М., Elster, E., and Lecoutre, J. P. (1976). Migration et acculturation differentiale. International Review of Applied Psychology, 25, 145-163.
Raven, J. (1981). Irish and British Standardisations. Oxford: Oxford Psychologists Press.
Raven, J. (1986). Manual for Raven’s Progressive Matrices and Vocabulary Scales.
London: Lewis.
Raven, J. (1998). Manual for Raven’s Progressive Matrices. Oxford: Oxford Psychologists Press.
Raven, J., and Court, J. H. (1989). Manual for Ravens Progressive Matrices and Vocabulary Scales. London: Lewis.
Raven, J. C. (1939). The RECI series of perceptual tests: an experimental survey. British Journal of Medical Psychology, 18, 16-34.
Raven, J. C., Court, J. H., and Raven,J. (1994). Advanced Progressive Matrices. Oxford: Oxford Psychologists Press.
Raven,J. C., Court, J. H., and Raven, J. (1995). Coloured Progressive Matrices. Oxford: Oxford Psychologists Press.
Raven,J. C., Court,J. H., and Raven,J. (1996). Standard Progressive Matrices. Oxford: Oxford Psychologists Press.
Raven,J. C., Court,J. H., and Raven,J. (1998). Advanced Progressive Matrices. Oxford: Oxford Psychologists Press.
Raven, ]., and Court, J. H. (1999). Manual for Raven’s Progressive Matrices and Vocabulary Scales. London: Lewis.
Raven, J., Raven, J. C., and Court, J. H. (2000). Standard Progressive Matrices. Oxford: Oxford Psychologists Press.
Razrin, G. (1971). Mind in Evolution. New York: Houghton Mifflin.
Redmond, М., and Davies, F. R. (1940). The Standardisation of Two Intelligence Tests. Wellington: New Zealand Council for Educational Research.
Reed, Т. E. (1969). Caucasian genes in American Negroes. Science, 165, 762-768.
Reed,T. E. (1971). The population variance of the proportion of genetic admixture in human intergroup hybrids. Proceedings of the National Academy of Science, 68, 3168— 3169.
Reid, N., and Gilmore, A. (1983). Pupil performance on TOSCA: Some additional information. New Zealand Journal of Educational Studies, 18, 13-81.
Reid, N., and Gilmore, A. (1989). The Raven’s Standard Progressive Matrices in New Zealand. Psychological Test Bulletin, 2, 25-35.
Reidel, A., Klekamp, ]., Harper, C., and Kretschmann, H. J. (1994). Morphometric study on the postnatal growth of the cerebral cortex of Australian aborigines and Caucasians. Journal of Brain Research, 35, 531-540.
Relethford, J. H. (1988). Genetics of modern human origins and diversity. Annual Review of Anthropology, 27, 1-23.
Resing, W. С. М., Bleichrodt, N., and Drenth, P. J. D. (1986). Het gebruit van de RAKIT bij allochtoon etnische groepen. Nederlands Tijdschrift voor de Psychologies 41, 179— 188.
Reuning, H. (1968). Psychological studies of Kalahari Bushmen. In L. J. Cronbach and P. J. Drenth (Eds.). Mental Tests and Cultural Adaptation.The Hague: Mouton.
Расовые различия в интеллекте
289
Reuning, Н. (1972). Testing Bushmen in the central Kalahari. In S. H. Irvine and J. W. Berry (Eds.). Human Abilities in Cultural Context. Cambridge, UK: Cambridge University Press.
Reynolds, C. R., Chastain, R. L., Kaufman, A. S., and McLean, J. E. (1987). Demographic characteristics and IQ among adults: analysis of WAIS-R standardization sample as a function of the stratification variables. Journal of School Psychology, 25, 323-342.
Reynolds, C. R., and Jensen, A. R. (1983). WISC-R subscale patterns of abilities of blacks and whites matched on full scale IQ. Journal of Educational Psychology, 15, 207-214.
Reynolds, C. R., Willson, V. L., and Ramsey, M. (1999). Intellectual differences among Mexican Americans, Papagos and whites, independent of g. Personality and Individual Differences, 27, 1181-1188.
Richardson, K., and Spears, D. (1972). From biology. In K. Richardson and D. Spears (Eds). Race, Culture and Education. Harmondsworth, UK: Penguin Books.
Richter, L. М., Griesel, R. D., and Wortley, М. E. (1989). The Draw-a-Man Test: A 50 year perspective on drawings done by black South African children. South African Journal of Psychology, 19, 1-5.
Rimoldi, H. J. (1948). A note on Raven’s Progressive Matrices Test. Educational and Psychological Measurement, 8, 347-352.
Risso,W. L. (1961). El test de Matrice Progressivas у el test Domino. Proceedings of the 1961 Conference of the Psychological Society of Uruguay.
Ritchie, J. E. (1966). Some observations on Maori and Pakeha intelligence test performance. Journal of the Polynesian Society, 66, 351-356.
Roberts, N. (1989). Pleistocene environments in time and space. In R. Foley (Ed.). Hominid Evolution and Community Ecology: Prehistoric Human Adaptation in Biological Perspective. London: Academic Press.
Robin, R. W, and Shea, J. D.C. (1983). The Bender Gestalt visual motor test in Papua New Guinea. International Journal of Psychology, 18, 263-270.
Rodd,W. G. (1959). A cross cultural study of Taiwan’s schools. Journal of Social Psychology, 50, 30-36.
Rohrer, J. H. (1942). The test intelligence of Osage Indians. Journal of Social Psychology, 16, 99-105.
Roth, P. L., Bevier, C. A., Bobko, P., Switzer, E S., and Tyler, P. (2001). Ethnic group differences in cognitive ability in employment and educational settings: a metaanalysis. Personnel Psychology, 54, 297-330.
Rouhani, S. (1989). Molecular genetics and the pattern of human evolution: plausible and implausible models. In P. Mellars and C. Stringer (Eds.). The Human Revolution. Cambridge, UK: Cambridge University Press.
Rowe, D. C. (2002). IQ, birth weight, and number of sexual partners in white, African American, and mixed race adolescents. Population and Environment, 23, 513-524.
Ruff, С. B., Trinkaus, E. and Holliday, T. W. (1997). Body mass and encephalization in Pleistocene Homo. Nature, 387, 173-176.
Rushton, J. P. (1988). Race differences in behaviour: a review and evolutionary analysis. Personality and Individual Differences, 9, 1009-1024.
Rushton, J. P. (1992a). Cranial capacity related to sex, rank and race differences in a stratified sample of 6,325 U. S. military personnel. Intelligence, 16, 401-413.
Rushton, J. P. (1992b). Life history comparisons between Orientals and whites at a Canadian university. Personality and Individual Differences, 13, 439-442.
Rushton, J. P. (1994). Sex and race differences in cranial capacity from International Labor Office data. Intelligence, 19, 281-294.
Rushton, J. P. (1997). Cranial size and IQ in Asian Americans from birth to seven. Intelligence, 25, 7-20.
Rushton, J. P. (2000). Race, Evolution and Behavior. Port Huron, MI: Charles Darwin Research Institute.
290
Ричард Линн
Rushton, J. P. (2003). Race differences in g and the «Jensen Effect.» In H. Nyborg (Ed.). The Scientific Study of General Intelligence. Amsterdam: Elsevier.
Rushton, J. P., and Jensen, A. R. (2005). Thirty years of research on race differences in cognitive ability. Psychology, Public Policy, and Law, 11, 235-294.
Rushton,J. P., and Osborne, R.T. (1995). Genetic and environmental contributions to cranial capacity in black and white adolescents. Intelligence, 20, 1-13.
Rushton, J. P., and Skuy, M. (2000). Performance on Raven’s Matrices by African and white university students in South Africa. Intelligence, 28, 251-266.
Rushton, J. P., Skuy, М., and Fridjhon, P. (2002). Jensen effects among African, Indian and white engineering university students in South Africa on Raven’s Standard Progressive Matrices. Intelligence, 30, 409-423.
Rushton, J. P., Skuy, М., and Fridjhon, P. (2003). Performance on Raven’s Advanced Progressive Matrices by African, Indian and white engineering university students in South Africa. Intelligence, 31, 123-138.
Rushton, J. P., Skuy, М., and Bons, T. A. (2004). Construct validity of Raven’s Advanced Progressive Matrices by African and Non-African engineering university students in South Africa. International Journal of Selection and Assessment, 12, 220-229.
Ryan, L. (1982). The Aboriginal Australians. St. Lucia: University of Queensland Press.
Saco-Politt, C. (1989). Ecocultural context of developmental risk: birth in the high altitudes (Peru). In J. K. Nugent, B. L. Lester, and Т. B. Bra-zelton (Eds.). The Cultural Context of Infancy. Norwood, NJ: Ablex.
Sadek, A. A. M. (1972). A Factor Analytic Study of Musical Abilities of Egyptian
Students Taking Music as a Special Subject. Ph.D. dissertation, University of London.
Sahin, N., and Duzen, E. (1994). Turkish standardisation of Raven’s SPM. Proceedings of the 23rd International Congress of Applied Psychology, Madrid.
Salkind, N.J.,Kojima,H.,and Zelniker,T. (1978). Cognitive tempo in American, Japanese and Israeli children. Child Development, 49, 1024-1027.
Sandiford, P. and Kerr, R. (1926). The intelligence of Chinese and Japanese children. Journal of Educational Psychology, 17, 361-367.
Scarr, S. (1981). Race, Social Class, and Individual Differences in IQ. Hillsdale, NJ: Lawrence Erlbaum.
Scarr, S. (1981). Comments and replies. In S. Scarr. Race, Social Class, and Individual Differences in IQ. Hillsdale,NJ: Lawrence Erlbaum.
Scarr, S. (1995). Inheritance, intelligence and achievement. Planning for Higher Education, 23,1-9.
Scarr, S., Carparulo, B. K.,Ferdman, В. М., Tower, R. B., and Caplan, J. (1983).
Developmental status and school achievements of minority and non-minority children from birth to 18 years in a British Mid-lands town. British Journal of Developmental Psychology, 1, 31-48.
Scarr, S., and McCartney, K. (1983). How people make their own environments: a theory of genotype-environment effects. Child Development, 54, 424—435.
Scarr, S., and Weinberg, R. A. (1976). IQ test performance of black children adopted by white families. American Psychologist, 31, 726-739.
Scarr, S., and Weinberg, R. A. (1978). The influence of family background on intellectual attainment. American Sociological Review, 43, 674-692.
Scott, L. H. (1981). Measuring intelligence with the Goodenough Draw-a-Man test. Psychological Bulletin, 89, 483-505.
Scottish Council for Research in Education. (1933). The Intelligence of Scottish Children. London: University of London Press.
Scottish Council for Research in Education. (1949). The Trend of Scottish Intelligence. London: University of London Press.
Seagrim, G. N., and Lendon, R. J. (1980). Furnishing the Mind. New York: Academic.
Seshadri, S., and Gopaldas, T. (1989). Impact of iron supplementation on cognitive functions in preschool and school-aged children: the Indian experience. American Journal of Clinical Nutrition, 50, 675-686.
Расовые различия в интеллекте
291
Seyfort, В., Spreen, О., and Lahmer, V. (1980). A critical look at the WISC-R with native Indian children. Alberta Journal of Educational Research, 16, 14-24.
Sheldon, W. H. (1924). The intelligence of Mexican children. School and Society, 19, 139-142.
Shigehisa, Т., and Lynn, R. (1991). Reaction times and intelligence in Japanese children. International Journal of Psychology, 26, 195-202.
Shockley, W. B. (1968). Proposed research to reduce racial aspects of the environment- heredity uncertainty. Science, 160, 443.
Shockley, W. B. (1969). Human quality problems and research taboos. New Concepts and Directions in Education, 28, 67-99.
Shockley, W. B. (1971). Hardy-Weinberg law generalized to estimate hybrid variance for Negro populations. Proceedings of the National Academy of Sciences, 68, 1390.
Shuey, A. M. (1966). The Testing of Negro Intelligence. New York: Social Science Press.
Sidles, C., and MacAvoy, J. (1987). Navajo adolescents’ scores on a primary language questionnaire and the Raven Progressive Matrices. Educational and Psychological Measurment, 47, 703-709.
Sijtsma, K., and Resing, W. С. M. (1991). Scalability of an intelligence test for different ethnic groups. In: N. Bleichrodt and P. J. D. Drenth (Eds.). Contemporary Issues in Cross-Cultural Psychology. Amsterdam: Swets and Zeitlinger.
Simeon, D. Т., and Gratham-McGregor, S. (1989). Effects of missing breakfast on the cognitive functions of school children of differing nutritional backgrounds. American Journal of Clinical Nutrition, 49, 646-653.
Simeon, D. Т., and Gratham-McGregor, S. (1990). Effects of nutritional deficiencies on intelligence and behavior. Nutrition Research Reviews, 3, 1-24.
Simmons, K. (1942). Cranial capacities by both plastic and water techniques with cranial linear measurements of the Reserve Collection: white and Negro. Human Biology, 14, 473-498.
Simoes, М. M. R. (1989). Un estudo exploratorio com о teste das matrizes progressivas de Raven para criancas. Proceedings of the Congress of Psychology, Lisbon.
Simon, J. A., Schreiber, G. B., Crawford, P. B., Frederick, М. М., and Sabry, Z. I. (1993). Income and racial patterns of dietary vitamin С intake among black and white girls. Public Health Reports, 108, 760-764.
Sinha, U. (1968). The use of Raven’s Progressive Matrices in India. Indian Educational Review, 3, 75-88.
Skandinaviska Testforlaget. (1970). Manual of the Swedish WISC. Stockholm: Skandinaviska Testforlaget.
Skuy, М., Gewer, A., Osrin, Y., Khunou, D., Fridjhon, P., and Rushton, J. P. (2002). Effects of mediated learning experiences on Raven’s matrices scores of African and non-African students in South Africa. Intelligence, 30, 221-232.
Skuy, М., Schutte, E., Fridjhon, P., and O’Carroll, S. (2001). Suitability of published neuropsychological test norms for urban African secondary school students in South Africa. Personality and Individual Differences, 30, 1413-1425.
Smith, C. L., and Beals, K. L. (1990). Cultural correlates with cranial capacity. American Anthropologist, 92, 193-200.
Smith, S. (1942). Language and non-verbal test performance of racial groups in Honolulu before and after a fourteen year interval. Journal of General Psychology, 26, 51-93.
Soewondo, S., Husaini, М., and Pollitt, E. (1989). Effects of iron defi-ciency on attention and learning processes in preschool children: Ban-dung, Indonesia. American Journal of Clinical Nutrition, 50, 667-674.
Sonke, C. J. (2000). Cross-cultural differences on simple cognitive tasks: a psychophysiolo- gical investigation. Tilberg: University Press.
Sorokin, B. (19 54). Standardisation of the Progressive Matrices test. Unpublished report.
Sowell, T. (1978). Three black histories. In T. Sowell and L. D. Collins (Ed.). Essays and Data on American Ethnic Groups. Washington, D.C.: The Urban Institute.
292
Ричард Линн
Sowell, Т. (1986). Education: Assumptions versus History. Stanford, CA: Hoover Institution Press.
Spearman, C. (1923). The Nature of Intelligence and the Principles of Cognition. London: Macmillan.
Spicher,P. (1993). Nouvel etalonnage du SPM. Fribourg, Switzerland: University of Fribourg.
Stahl, A. B. (1984). Hominid dietary selection before fire. Current Anthropology, 25, 151-68.
Stams, G. J., Juffer, R, Rispens, J., and Hoksergen, R. A. C. (2000). The development and adjustment of 7 year old children adopted in infancy. Journal of Child Psychology and Psychiatry, 41, 1025-1037.
Stanford, С. B., and Bunn, H. T. (2001). Introduction. In С. B. Stanford and H. T. Bunn (Eds.). Meat Eating and Human Evolution. Oxford, UK: Oxford University Press.
Stein, Z., Susser, М., Saenger G., and Marolla, F. (1972). Nutrition and mental performance. Science, 178, 708-713.
Sternberg, R. J., Grigorenko, E. L., Ngorosho, D., Tantufuye, E., Mbise, A., Nokes, C., Jukes, М., and Bundy, D. A. (2002). Assessing intellectual potential in rural Tanzanian school children. Intelligence, 30, 141-162.
Sternberg, R. J., Nokes, C., Geissler, P. W., Prince, R., Okatcha, E, Bundy, D. A., and Grigorenko, E. L. (2001). The relationship between academic and practical intelligence: a case study in Kenya. Intelligence, 29, 401-418.
Sternberg, R. J., Powell, C., McGrane, P., and Grantham-McGregor, S. (1997). Effects of a parasitic infection on cognitive functioning. Journal of Experimental Psychology : Applied, 3, 67-76.
Stevenson, H. W., and Azuma, H. (1983). IQ in Japan and the United States: methodological problems in Lynn’s analysis. Nature, 306, 291-292.
Stevenson, H. W., Stigler, J. W., Lee, S., Lucker, G. W. Kitanawa, S. and Hsu, C. (1985). Cognitive performance and academic achievement of Japanese, Chinese and American children. Child Development, 56, 718-734.
Stewart, L. H., Dole, A. A., and Harris, Y. Y. (1967). Cultural differences in abilities during high school. American Educational Research Journal, 4, 19-30.
Stewart, R., Johnson, J., Richards, М., Brayne, C., and Mann, A. (2002). The distribution of Mini-Mental State Examination scores in an older UK African-Caribbean population compared to MRC CFA study norms. International Journal of Geriatric Psychiatry, 17, 745-751.
St. George, A. (1974). Cross-cultural ability testing. Unpublished.
St. George, R. (1970). The psycholinguistic abilities of children from different ethnic backgrounds. Australian Journal of Psychology, 22, 85-89.
St. George, R. (1983). Some psychometric properties of the Queensland Test of Cognitive Abilities with New Zealand, European and Maori children. New Zealand Journal of Psychology. 12, 57-68.
St. George, R., and Chapman,J. W. (1983).TOSCA results from a New Zealand sample. New Zealand Journal of Educational Studies, 18, 178-183.
St. George, R., and St. George, S. (1975). The intellectual assessment of Maori and European children. In R D. K. Ramsay (Ed.). The Family at School in New Zealand. London: Pitman.
St. John, J.,Krichev, A., and Bauman, E. (1976). North Western Ontario Indian children and the WISC. Psychology in the Schools. 13, 407-411.
Storfer, M. D. (1990). Intelligence and Giftedness. Jossey-Bass, San Erancisco.
Strauss, M. A. (1954). Sub-cultural variation in Ceylonese mental ability: a study in national character. Journal of Social Psychology, 39, 129-141.
Stringer, С. B., and Andrews, R (1988). Genetic and fossil evidence for the origin of modem humans. Science, 239, 1263-1268.
Stringer, С. B., and McKie, R. (1996). African Exodus. London: Pimlico.
Strum, S. C. (1981) Processes and products of change: baboon predatory behavior at Gilgil, Kenya. In R. Harding and G. Telela (Eds.). Omnivorous Primates. New York, Columbia University Press.
Расовые различия в интеллекте
293
Sugishita, М., and Omura, К. (2001). Learning Chinese characters may improve visual memory. Perceptual and Motor Skills, 93, 579-594.
Sundberg, N., and Ballinger, T. (1968). Nepalese children’s cognitive development as revealed by drawings of man, woman and self. Child Development, 39, 969-985.
Sung, Y. H., and Dawis, R. V. (1981). Level and factor structure differences in selected abilities across race and sex groups. Journal of Applied Psychology, 66,613-624.
Symonds, P. M. (1924). The intelligence of Chinese in Hawaii. School and Society, 19, 442.
Takeuchi, М., and Scott, R. (1992). Cognitive profiles of Japanese and Canadian kindergarten and first grade children. Journal of Social Psychology, 132, 505-512.
Tamaoka, K., Saklofske, D. H., and Ide, M. (1993). The non-verbal reasoning ability of Japanese children measured by Naglieri’s matrix analogies test — short form. Psychologia, 36, 53-60.
Tan, U., Tan, М., Polat, P., Ceylan, Y, Suma, S., and Okur, A. (1999). Magnetic resonance imaging brain size/IQ relations in Turkish university students. Intelligence, 27, 83-92.
Tarnopol, L., and Tarnopol, M. (1980). Arithmetic ability in Chinese and Japanese children. Focus on Learning Problems in Mathematics, 2, 29-48.
Taschinski, R. (1985). Eine Untersuchung zur Kulturfaimws der Progressiven Matrizen von Raven gegeniiber tiirkischen Kindem in Deutschland. Psychologic in Erziehung und Unterricht, 32, 229-239.
Taylor, J. М., and Radford, E. J. (1986). Psychometric testing as an unfair labour practice. South African Journal of Psychology, 16, 79-86.
Taylor, L. J., and Skanes, G. R. (1976a). Level 1 and level 11 abilities in Inuit and white children from similar environments. Journal of Cross-Cultural Psychology, 7, 157-167.
Taylor, L. J., and Skanes, G. R. (1976b). Cognitive abilities of Inuit and white children from similar environments. Canadian Journal of Behavioral Science, 8, 1-8.
Taylor, L. J., and Skanes, G. R. (1977). A cross-cultural examination of some of Jensen’s hypotheses. Canadian Journal of Behavioral Science, 9, 315-322.
Teasdale, G. R., and Katz, F. M. (1968). Psycholinguistic abilities of children from different ethnic and socio-economic backgrounds. Australian Journal of Psychology, 20, 133-159.
Teasdale, T. W., and Owen, D. R. (1994). Thirty year secular trend in the cognitive abilities of Danish male school leavers at a high educational level. Scandinavian Journal of Psychology, 35, 328-335.
Teasdale, T. W., and Owen, D. R. (2000). Forty-year secular trends in cognitive abilities. Intelligence, 28, 115-120.
Teeter, A., Moore, C., and Petersen, J. (1982). WISC-R verbal and performance abilities of Native American students referred for school learning problems. Psychology in the Schools, 19, 39-44.
Telford, C. W. (1932). Test performance of full and mixed-blood North Dakota Indians. Journal of Comparative Psychology, 14, 123-145.
Templer, D. I., and Arikawa, H. (2005). Temperature, skin color, per capita income and IQ: an international perspective. Intelligence (to appear).
Te Nijenhuis, J. (1997). Comparability of Test Scores for Immigrants and Majority Group Members in the Netherlands. Enschede, the Netherlands: Ipskamp.
Te Nijenhuis, J., Tolboom, E. A., Resing, W. C., and Bleichrodt, N. (2004). Does cultural background influence the intellectual performance of children of immigrant groups? European Journal of Psychological Assessment, 20, 10-26.
Te Nijenhuis,J., and van der Flier, H. (2001). Group differences in mean intelligence for the Dutch and third world immigrants. Journal of Biosocial Science, 33, 469-475.
Terman, L. M. (1916). The Measurement of Intelligence. Boston, MA: Houghton Mifflin.
Tesi, G., and Young, В. H. (1962). A standardisation of Raven’s Progressive Matrices. Archive de Psicologia Neurologia e Psichologia, 5, 455-464.
Tesser, P. Т. М., Merens, J. G., and van Prag, C. (1999). Rapportage minder heden 1999. Rijswijk: Social en Cultured Planburewau.
294
Ричард Линн
Themstrom, A., and Thernstrom, S. (2003). No Excuses: Closing the Racial Gap in Learning. New York: Simon and Schuster.
Thomas, N. W. (1925). Australia: its native races and their customs. In: J. A. Hammerton (Ed.). Peoples of All Nations. London: Amalgamated Press.
Thomas, R. М., and Shah, A. (1961). The Draw-a-Man Test in Indonesia. Journal of Educational Psychology, 32, 232-235.
Thompson, P. М., Cannon, T. D., Narr, K. L., van Erp, T, and Poutanen, V-P. (2001). Genetic influences on brain structure. Nature Neuroscience, 4, 1253-1258.
Thorndike, R. L., Hagen, E. P., and Sattler, J. M. (1986). Stanford-Binet Intelligence Scale: Fourth Edition Manual. Chicago: Riverside.
Thurstone, L. L. (1938). Primary Mental Abilities. Chicago: University of Chicago Press.
Tizard, B. (1972). IQ and race. Nature, 247, 316.
Tobias, P. V. (1970). Brain-size, grey matter and race — fact or fiction? American Journal of Physical Anthropology, 32, 3-26.
Tooby,J., and Devore, I. (1989). The reconstruction of hominid behavioral evolution through strategic modelling. In W. G. Kinzey (Ed.). The Evolution of Human Behavior: Primate Models. Albany, NY: State University of New York Press.
Torrence, R. (1983). Time budgeting and hunter-gatherer technology. In G. Bailey (Ed.). Hunter-Gatherer Economy in Prehistory: A European Perspective. Cambridge, Cambridge University Press.
Turner, G. H., and Penfold, D. J. (1952). The scholastic aptitude the Indian children of the Caradoc reserve. Canadian Journal of Psychology, 6, 31-44.
Tzriel, D., and Caspi, N. (1992). Cognitive modifiability and cognitive performance of deaf and hearing preschool children. Journal of Special Education, 26, 235-252.
Ucman, P. (1972). A normative study of the Goodenough-Harris test on a Turkish sample. In L. J. Cronbach and P. J. D. Drenth (Eds.). Mental Tests and Cultural Adaptation. The Hague: Mouton.
UNICEF. (1996). The State of the World’s Children. Oxford, U. K.: Oxford University Press.
United States Bureau of the Census. (1989). The Hispanic population of the United States. Current Population Reports, series P-20, No. 444.
United States Department of Health, Education and Welfare. (1970). Intellectual Maturity of Children. Washington, D.C.
United States Department of Health, Education and Welfare. (1971). Intellectual Development of Children. Washington, D.C.
United States National Aeronautics and Space Administration. (1978). Anthropometric Source Book, vol. 2, Handbook of Anthropometric Data. Washington, D.C.
Valencia, R. R. (1979). Comparison of performance of Chicano and Anglo-Saxon third grade boys on Raven’s Colored Progressive Matrices. Psychology in the Schools, 16, 448-453.
Valentine, M. (1959). Psychometric testing in Iran. Journal of Mental Science, 105, 93-107.
Van de Vijver, F. J. R., and Willemse, G. R. (1991). Are reaction time tasks better suited for cultural minorities than paper and pencil tests? In N. Bleichrodt and P. J. D.
Drenth (Eds.). Contemporary Issues in Cross-Cultural Psychology. Amsterdam: Swets and Zeitlinger.
Vandenburg, S.G. (1962).The hereditary abilities study: hereditary components in a psychological test battery. American Journal of Human Genetics, 14, 220-237.
Vejleskov, H. (1968). An analysis of Raven Matrix responses in fifth grade children. Scandinavian Journal of Psychology, 9, 177-186.
Verhagen, P. (1956). Utilite actuelle des tests pour l»etude psychologique des autochtones Congolais. Revue de Psychologie Appliquee, 6, 139-151.
Verive, J. М., and McDaniel, M. A. (1996). Short-term memory tests in personnel selection: low adverse impact and high validity. Intelligence, 23, 15-32.
Vernon, P. A. (1989). The heritability of measures of speed of informa-tion-processing. Personality and Individual Differences, 10, 573-576.
Расовые различия в интеллекте
295
Vernon, P. A., Wickett, J. С., Bazana, P. G., and Stelmack, R. М. (2000). The
neuropsychology and neurophysiology of human intelligence. In R. J. Sternberg (Ed.). Handbook of Intelligence. Cambridge, UK: Cambridge University Press.
Vernon, P. E. (1969). Intelligence and Cultural Environment. London: Methuen.
Vernon, P. E. (1979). Intelligence: Heredity and Environment. San Francisco: Freeman. Vernon, P. E. (1982). The Abilities and Achieve-ments of Orientals in North America. New York: Academic Press.
Vernon, P. E. (1984). The abilities and achievements of ethnic groups in Canada with special reference to Canadian natives and Orientals. America. In S. M. Samuda, J. W. Berry, and M. Laferriere (Eds.). Multiculturalism in Canada. Toronto: Allyn and Bacon.
Vincent, P. (1966). The measured intelligence of Glasgow Jewish schoolchildren. Jewish Journal of Sociology, 8, 92-108.
Vitti, P., Aghini-Lombardi, E, and Antonangeli, L. (1992). Mild iodine deficiency in fetal/ neonatal life and neuropsychological performances. Ada Medica Austriaca, 19, 57-67.
Wagner, K. (1937). The cranial capacity of the Oceanic races. Norske Videnskaps-akademi I Oslo. 1 Mat. Naturv. Klasse No 2.
Wake, C. S. (1872). The mental characteristics of primitive man as exemplified by the Australian Aborigines. Journal of the Anthropological Institute, 1, 74-84.
Waldman, I. D., Weinberg, R. A., and Scarr, S. (1994). Racial-group differences in IQ in the Minnesota Transracial adoption study. Intelligence, 19, 29-44.
Waldron, L. A., and Gallimore, A. J. (1973). Pictorial depth perception in Papua New Guinea, Torres Straits and Australia. Australian Journal of Psychology, 25, 89-92.
Walters, R. H. (1958). The intelligence test performance of Maori children: a cross- cultural study. Journal of Abnormal and Social Psychology, 58, 107-114.
Watanabe, S., Ayta, W. E. E, and Hamaguchi, H. (2003). Some evidence of a date of first humans to arrive in Brazil. Journal of Archeological Science, 30, 351-254.
Weede, E., and Kampf, S. (2002). The impact of intelligence and institutional improvements on economic growth. Kyklos, 55, 361-380.
Wein,N., and Stevenson, B. (1972). Pre-school education programme — Dominica: Pilot evaluation of 3 and 4 year olds. Jamaica: Bernard Van Leer Foundation UWI, Mona.
Weinberg, R. A., Scarr, S., and Waldman, I. D. (1992). The Minnesota transracial
adoption study: a follow-up of the IQ test performance at adolescence. Intelligence, 16, 117-135.
Werner, E. E., Bierman, J., and French, E (1971). The Children of Kauai: A Longitudinal Study from the Prenatal Period to Age Ten. Honolulu: University of Hawaii Press.
Werner, E. E., Simonian, K., and Smith R. S. (1968). Ethnic and socio-economic status differences in abilities and achievements among preschool and school-aged children in Hawaii. Journal of Social Psychology, 75, 43-59.
West, A. М., Mackintosh, N. J., and Mascie-Taylor, C. G. N. (1992). Cognitive and educational attainment in different ethnic groups. Journal of Biosocial Science, 24, 539-554.
West, L. W, and MacArthur, R. S. (1964). An evaluation of selected intelligence tests for two samples of Metis and Indian children. Alberta Journal of Educational Research, 10, 17-27.
Weyl, N. (1967a). White Rhodesians: an unrecognised intellectual elite. Mankind Quarterly, 7, 207-210.
Weyl, N. (1967b). Personal communication.
Wilgosh, L., Mulcahy, R., and Walters, B. (1986). Assessing intellectual performance of culturally different Inuit children with the WISC-R. Canadian Journal of Behavioral Science, 18, 270-277.
Williams, W. M. (1998). Are We Raising Smarter Children Today? School- and Home- Related Influences on IQ. In U. Neisser (Ed.) The Rising Curve: Long term changes in IQ and related measures. Washington, D.C.: American Psychological Association Books.
296
Ричард Линн
Winkelmann, W. von (1972). Normen fur den Mann-Zeichen-Test von Ziler und die Coloured Progressive Matrices von Raven fur 5-7 jahrige Kinder. Psychologische Beitrage, 17, 80-94.
Winick, М., Meyer, К. K. and Harris, R. C. (1975). Malnutrition and environmental enrichment by early adoption. Science, 190, 1173-1175.
Wober, M. (1969). The meaning and stability of Raven’s matrices test among Africans. International Journal of Psychology, 4, 229-235.
Woodworth, R. S. (1910). Race differences in mental traits. Science, 31,171-186.
Workman, P. L. (1968). Gene flow and the search for natural selection in man. Human Biology, 40, 260-279.
Wright, S. C.,Taylor, D. М., and Ruggiero, К. M. (1996). Examining the potential for academic achievement among Inuit children. Journal of Cross-Cultural Psychology, 27, 733-753.
Wycherley, R., and Benjamin, L. (1998). WAIS-111 UK Manual. London: Psychological Corporation.
Wynn, T. (1989). The Evolution of Spatial Competence. Urbana, II: University of Illinois Press.
Yates, A. J., and Forbes, A. R. (1967). Raven’s Advanced Progressive Matrices: Provisional Manual for Australia and New Zealand. Hawthorn, Victoria: Australian Council for Educational Research.
Yee, L. Y., and La Forge, R. (1974). Relationship between mental abilities, social class, and exposure to English in Chinese fourth graders. Journal of Educational Psychology, 66, 826-834.
Yeung, К. T. (1922). The intelligence of Chinese children in San Francisco and vicinity. Journal of Applied Psychology, 5, 267-274.
Yousefi, E, Shahim, S., Razavieh, A., Mehryar, A. H., Hosseini, A. A., and Alborzi, S. (1992). Some normative data on the Bender Gestalt test performance of Iranian children. British Journal of Educational Psychology, 62, 410-416.
Yule, W., Berger, М., Rutter, М., and Yule, B. (1975). Children of West Indian
immigrants — 11. Intellectual performance and reading attainment. Journal of Child Psychology and Psychiatry, 16, 1-17.
Zaaiman, H. (1998). Selecting Students for Mathematics and Science. Pretoria: Sigma Press.
Zahimic, C., Girboveanu, М., Onofrei, A., Turcu, A., Voicu, C., Voicu, М., and Visan,О. M. (1974). Etolonarea matricelor progressive colorate Raven. Revista de Psihologie, 20, 313-321.
Zeidner, M. (1987a). Test of the cultural bias hypothesis: some Israeli findings. Journal of Applied Psychology, 72, 38-48.
Zeidner, M. (1987b). Validity of college admission indices for Jews and Arabs in Israel. Personality and Individual Differences, 8, 587-588.
Zhou, Z., and Boehm, A. E. (2001). American and Chinese children’s knowledge of basic relational concepts. School Psychology International, 22, 5-21.
Zindi, E (1994). Differences in psychometric performance. The Psychologist, 1, 549-552.
Zsembik, B. A., and Peek, М. K. (2001). Race differences in cognitive functioning among older adults. Journal of Gerontology: Social Sciences, 56B, 266-274.
ЖПСРЖ ЭШОЧЖ
на книгу Ричарда Линна «Расовые различия в интеллекте: эволюционный анализ». Вашингтон, 2006.
Книга Р. Линна «Расовые различия в интеллекте: эволюционный анализ» представляет собой фундаментальный научный труд, в котором а) проанализированы данные примерно 550 исследований этнических особенностей мышления и б) сделана попытка объяснить возникновение этих различий в рамках эволюционного подхода. Исследования этнических особенностей мышления имеют давнюю традицию, они проводились не только за рубежом, но и в нашей стране, сошлемся на известную работу А.Р. Лурии «Об историческом развитии познавательных процессов», вышедшую в 1974 г. в издательстве «Наука». Существование этнических различий мышления является, в свете данных многочисленных исследований, бесспорным. Спорным, однако, является вопрос об источниках этих различий.
Одни специалисты считают, что этнические различия познавательных процессов, в частности, мышления, всецело обусловлены средовыми факторами (питанием, культурными традициями, уровнем образования и т.д.), в то время как другие придают значение генетическим факторам. В число последних входит и автор экспертируемой книги Р. Линн. Принятие второй позиции имеет своим вероятным следствием поиск различий между генетически различающимися популяциями. И действительно, в рассматриваемой книге автор сосредотачивается на анализе различий между десятью геногеографическими кластерами, которые он именует более традиционным термином «раса». Принятая автором классификация человеческих популяций была предложена рядом биологов на основе результатов геногеографических исследований и имеет значительное сходство с традиционной антропологической классификацией человеческих рас. Между выделяемыми в геногеографических исследованиях кластерами, или расами, существуют, как показывает сопоставительный аналих данных исследований, в которых использовались разные методы, различия в интеллектуальных способностях (вербальных способностях, способности к визуализации и т.д.).
Автор экспертируемой книги, Р. Линн, объясняет возникновение данных различий тем, что предки народов, принадлежащих к разным расам, проживали в природных зонах, предъявляющих не одинаковые требования к выживанию. В тех природных зонах, где выжить было трудно, более высокий уровень развития той или иной способности повышал вероятность выживания. Это обуславливало естественный отбор особей, обла
298
Ричард Линн
дающих данной способностью. Так, высокое развитие у арктических народов (эскимосов и др.) визуализационной способности объясняется тем, что в условиях однообразной, бедной приметами местности люди, обладающие этой способностью, имели больше шансов выжить. Большое значение Р. Линн придает также фактору многочисленности популяции, способствующей распространению полезных мутаций. Данное объяснение полностью укладывается в рамки весьма влиятельного направления в психологии, известного под названием функционализма, объясняющего возникновение и развитие психических функций с точки зрения теории эволюции Ч. Дарвина: их приспособительной полезностью и связанными с этим преимуществами в ходе естественного отбора.
Придавая большое значение фактору наследственности, Р. Линн отнюдь не игнорирует средовые факторы. Более того, несомненным достоинством его книги являются расчеты вклада в этнические различия интеллекта средовых факторов, например, некачественного питания, особенно недостатка в пище йода. На основе этих расчетов можно принимать решения о том, на какие средовые факторы следует воздействовать, чтобы уменьшить интеллектуальные различия между конкретными популяциями, и предсказывать эффективность таких воздействий.
Межпопуляционные различия в интеллекте связываются в экспедируемой книге с особенностями материальной и духовной культуры народов. Автора, однако, нельзя обвинить в столь распространенном на Западе «европоцентризме». Напротив, им подчеркивается тот большой вклад, который некоторые неевропейские народы внесли в мировую культуру. Фиксируя интеллектуальные и культурные различия между этносами, Р. Линн не делает утверждений о якобы вытекающем отсюда праве одних народов диктовать нормы жизни другим, тем более об их праве господствовать.
В книге Линна, как и во всяком научном исследовании, просматривается, подчас, и определенная увлеченность автора идеей расовых различий интеллекта, эмоциональная окрашенность некоторых утверждений и т.п. Мы упомянули данный аспект работы автора в связи с тем, что современная российская общественность, как ни странно, и к месту и не к месту является чрезвычайно чувствительной к расовым проблемам. Такая настороженность должна бы быть более понятна в таких странах как США (в связи с не так давно, но, слава Богу, уже ушедшим в прошлое расизмом и геноцидом коренного населения) или Германия (с приснопамятным немецким национал-социализмом), но не в России. Хотя я сам и не являюсь сторонником активного развития расовой теории интеллекта, однако, сообразуясь с принципом «сон разума рождает чудовищ», считаю необходимым ввести исследования, проведенные Р. Линном, в поле проблем, рассматриваемых российской научной общественностью.
Расовые различия в интеллекте
299
В целом, книга Р. Линна «Расовые различия в интеллекте: эволюционный анализ» является работой, имеющей несомненную научную и практическую ценность. Она не содержит призывов к межнациональной розни, утверждений о праве одних народов навязывать свои культурные образцы другим и пр. В книге, на наш взгляд, не содержится антиправовых и экстремистских высказываний и оценок. Данная книга может быть рекомендована к изданию в РФ для ознакомления научной общественности с достижениями западной науки.
Директор центра этнофункциональных исследований
при Государственной академии славянсю
А?
доктор психологических наук, профест^
А.В. Сухарев
300
Ричард Линн
О некоторых работах Р. Линна после 2006 года
Необходимо сказать несколько слов о работах Ричарда Линна, опубликованных после 2006 года — года издания его книги «Расовые различия в интеллекте: эволюционный анализ» на языке оригинала — и являющихся интересным и важным дополнением к ее материалу1.
В одной из них, озаглавленной «Расовые различия в интеллекте: креативность и творческие достижения»2, сделана попытка разрешить парадокс: почему народы Восточной Азии (китайцы, японцы и корейцы), обладая более высоким, чем у европейцев, интеллектом, значительно уступают им в творческих достижениях. В качестве показателя творческих достижений нации в данном исследовании берется суммарное количество Нобелевских премий в области науки, литературы и экономики, и Филдсовских премий в области математики, полученных с 1901 по 2006 гг., на 1 миллион человек популяции. По этому показателю европейцы, отставая от народов Восточной Азии по IQ, явно лидируют. Для объяснения этого несоответствия привлекается такая личностная характеристика, как «открытость к опыту» (один из т. наз. «большой пятерки» личностных факторов). Приводятся ее национальные оценки, полученные в ряде исследований в различных странах. Как оказалось, медиана исследований в странах Восточной Азии на 0,6 стандартного отклонения (имеется в виду стандартное отклонение для данных США) ниже медианы исследований в европейских странах. Причем данное превосходство европейцев больше превосходства жителей Восточной Азии над ними в IQ. Таким образом, относительно низкие творческие достижения народов Восточной Азии могут быть объяснены их недостаточной «открытостью к опыту».
В другой работе3 прогнозируется динамика интеллекта во всем мире в ближайшие десятилетия: согласно расчетам, к 2050 г. общемировое снижение интеллекта должно составить 1,27 единиц IQ. Наблюдавшееся в XX веке повышение интеллекта, получившее название «эффект Флинна», было вызвано, надо полагать, улучшением условий жизни. В настоящее время этот фактор, похоже, «исчерпал свои возможности». А с прекращением его действия выступает на передний план другой фактор — фактор отрицательной связи между рождаемостью и интеллектом (эта связь в большей мере характерна для женщин). А этот фактор, понятно, действует в сторону снижения интеллекта. И действительно, согласно результатам четырех исследований, проведенных в Норвегии, Австралии, Дании и Великобритании, повышение интеллекта остановилось или даже началось его снижение.
1 Некоторые из упоминаемых здесь работ профессор Р. Линн любезно предоставил в распоряжение публикатора русского перевода своей книги.
2Lynn, R. (2007). Race differences in intelligence: creativity and creative achievement. The Mankind Quarterly, 48(2), 157-168.
3Lynn, R. & Harvey, J.J. (2008). The decline of the world’s IQ. Intelligence, 36, 112-120.
Расовые различия в интеллекте
301
В двух статьях приводятся данные о способностях народов, не охваченных приведенными в книге исследованиями, а именно монголов4 и тибетцев5. Эти данные были собраны китайскими исследователями и сравнивались ими с аналогичными данными китайцев. Но, так как имеется оценка IQ китайцев относительно британских норм, результаты монголов оказалось возможным выразить в «британской метрике», обычно используемой для сопоставления национальных IQ. Оценка IQ монголов в этой метрике равна 100 — в точности как в Великобритании. Эта величина IQ монголов очень хорошо предсказывается из данных об их генетической близости с народами, интеллект которых был измерен раньше. Что касается интеллекта тибетцев, то он, в британской метрике, оказался равным 92,4 баллам (на 12,6 баллов ниже, чем у китайцев). Уступили тибетцы китайцам и в результатах испытаний по математике. Как и в случае с монголами, результаты тестирований согласуются с антропологическими данными о расовой принадлежности тибетцев.
В двух работах6 рассматривается связь между национальным IQ и результатами двух независимых обследований достижений в образовании в ряде стран, проведенными в 2003 и в 2006 годах (программы TIMSS и PISA). В первом исследовании коэффициенты корреляции между IQ и показателями успеваемости варьировали от 0,85 до 0,93, после поправки на аттеньюацию от 0,92 до 1. Во втором исследовании коэффициент корреляции между IQ и суммарным показателем учебных достижений оказался равным 0,84, а поправка на аттеньюацию повысила его значение до 0,935;. Во втором исследовании рассматривалась также связь IQ с рядом других переменах, в частности, с заинтересованностью учащихся в изучении различных наук.
Наконец, в одной статье7 сделан подробный обзор данных о социоэкономи- ческих и этнических различиях в интеллекте, собранных отечественными исследователями до того, как работы такого рода были в СССР запрещены (в 1930-х годах). Эти данные рассмотрены в сопоставлении с результатами других исследований и с материалами обследования достижений в образовании по программе PISA; согласованность сопоставляемых данных оказалась очень высокой. Отечественному читателю, интересующемуся историей исследований IQ в СССР, можно также порекомендовать обратиться к работам Н. С. Курека8.
Исследования межпопуляционных различий в интеллекте продолжают проводиться, и можно надеяться, что вскоре будут получены данные об интеллекте народов, обойденных, на сегодняшний день, вниманием психологов.
д. филол. н. А. А. Григорьев
4Lynn, R. (2007). The IQ of Mongolians. Mankind Quarterly, 48, 91-97.
5 Lynn, R. (2008). The IQ and math ability of Tibetans and Han Chinese. Mankind Quarterly, 48, 505-510.
6 Lynn, R. & Mikk, J. (2007). National differences in intelligence and educational attainment. Intelligence, 35, 115-121; Lynn, R. & Mikk, J. (2009). National IQs predict attainment in math, science and reading across 56 nations. Intelligence, 37, 305-310.
7 Grigoriev, A., & Lynn, R. (2009). Studies of socioeconomic and ethnic differences in intelligence in the former Soviet Union in the early twentieth century. Intelligence, 37, 447-452.
8Курек H.C. (1997). Педология и психотехника о нравственном, интеллектуальном и физическом уровнях развития населения СССР в 20-е годы. Психологический журнал, 18, 3, 149-159; Курек Н.С. (2004). История ликвидации педологии и психотехники в СССР. СПб.: АЛЕТЕЙЯ.
Издательская Группа Профит Стайл
ИЗДАТЕЛЬСКИЙ ОТДЕЛ РАССМОТРИТ РУКОПИСИ ПО РАЗЛИЧНЫМ ЭЗОТЕРИЧЕСКИМ НАУКАМ
Приглашаются для совместной работы
Авторы и Переводчики Тел. 8-903-559-24-73 E-mail: gedas-maskvoje@mail.ru
ОТДЕЛ РЕАЛИЗАЦИИ ПРИГЛАШАЕТ К СОТРУДНИЧЕСТВУ:
❖ Распространителей эзотерической литературы;
♦> Книготорговые организации;
Тел./факс в Москве (495) 958-10-68, т. (499) 342-84-96
www.profit-style.ru* * * e-mail: proflt-l@mail.ru

Издательство «БЕЛЫЕ АЛЬВЫ» предлагает книги серии «Библиотека расовой мысли»
Андрей Савельев. Образ врага. Расология и политическая антропология. Серия «Библиотека расовой мысли». — М.: Белые альвы, 2007. — 608 с.: ил.
Философия вождизма. Хрестоматия по вождеведению под ред. В. Б. Авдеева. Серия «Библиотека расовой мысли» / Перев. с нем. А. М. Иванова. — М.: Белые альвы, 2006. — 608 с.: ил.
Ганс Ф. К. Гюнтер. Избранные работы по расологии. Изд. второе до- полн. и проилл. / Перевод с нем. А. М. Иванова. Предисловие Вл. Авдеева, Ан. Иванова, Юр. Ригера. Серия «Библиотека расовой мысли». — М.: Белые альвы, 2005. — 576 с.: ил.
Ганс Ф. К. Гюнтер. Расовые типы Европы. Постер (плакат). Приложение к серии «Библиотека расовой мысли»/ Под ред. В. Б. Авдеева.
Эрнст Крик. Преодоление идеализма. Основы расовой педагогики/ Пер. с нем. А. М. Иванова. Предисловие В. Б. Авдеева и А. М. Иванова. Серия «Библиотека расовой мысли». — М.: Белые альвы, 2004. — 432 с.: ил.
Авдеев В. Б. Расология: Наука о наследственных качествах людей. Изд
2-е, дополн. Серия «Библиотека расовой мысли». — М.: Белые альвы, 2007. — 672 с.: ил.
Карл Штрац. Расовая женская красота. Изд. 2-е, исправл. /Пер. с нем. В. Комаровского. Предисловие В. Б. Авдеева. Серия «Библиотека расовой мысли». — М.: Белые альвы, 2009. — 368 с.
Раса и мировоззрение. Хрестоматия. Сборник оригинальных философских работ / Пер. с нем. А. М. Иванова. Под ред. В. Б. Авдеева. Предисловие В. Б. Авдеева. — М.: Белые альвы, 2009. — 480 с.: ил.
Заказ книг
Интернет-магазин: shop.influx.ru
Книжный клуб: тел. (495) 235-87-97
Ричард Линн
Расовые различия в интеллекте. Эволюционный анализ
Идея проекта: Авдеев В. Б.
Перевод с английского: Румянцев Д. О.
Научный консультант: доктор филологических наук Григорьев А. А. Верстка: Странников В.Ю.
Книжный Дом «Профит Стайл»,
Адрес: г. Москва, ул. М. Калужская, д. 8.
Тел./факс в Москве: (495) 958-10-68 т. (499) 342-84-96 e-mail: profit-l@mail.ruwww.profit-style.ru
Подписано в печать 27.10.09. Формат 70x100 7)6 Гарнитура «Таймс». Печать офсетная.
Бумага офсетная. Печ. л. 19.0.
Тираж 2000 экз. Заказ № 6505.
Отпечатано в полном соответствии с качеством предоставленных материалов в ОАО «Дом печати — ВЯТКА». 610033, г. Киров, ул. Московская, 122.
Факс: (8332) 25-58-83, 53-53-80 http://www.gipp.kirov.ru e-mail: pto@gipp.kirov.ru

Последние комментарии
4 часов 5 минут назад
4 часов 8 минут назад
2 дней 10 часов назад
2 дней 14 часов назад
2 дней 16 часов назад
2 дней 17 часов назад