Биохимия гормонов [Алла Анатольевна Масловская] (fb2) читать постранично, страница - 3
- Биохимия гормонов 1.08 Мб, 31с. скачать: (fb2) - (исправленную) читать: (полностью) - (постранично) - Алла Анатольевна Масловская
[Настройки текста] [Cбросить фильтры]
- 1
- 2
- 3
- 4
- 5
- . . .
- последняя (13) »
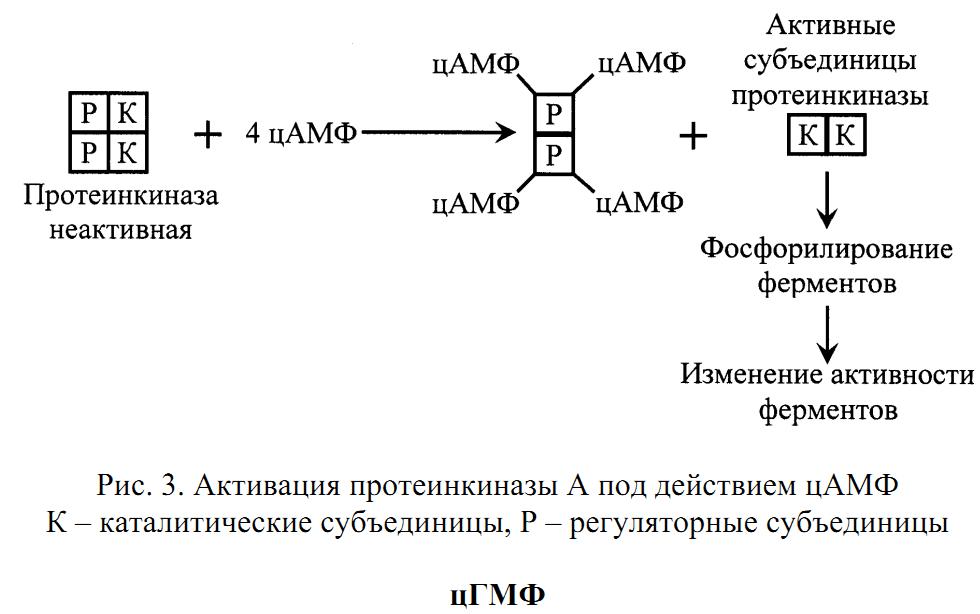
отсоединение (диссоциация) каталитических субъединиц, которые фосфорилируют белки (ферменты), изменяя их активность.
Разрушается цАМФ фосфодиэстеразой.
 Циклический ГМФ образуется из ГТФ под действием гуанилатциклазы по аналогии с синтезом цАМФ. Циклический ГМФ активирует специфическую цГМФ-зависимую протеинкиназу или протеинкиназу G, которая фосфорилирует ферменты, что сопровождается изменением их активности. Разрушается цГМФ, как и цАМФ, фосфодиэстеразой.
Циклический ГМФ образуется из ГТФ под действием гуанилатциклазы по аналогии с синтезом цАМФ. Циклический ГМФ активирует специфическую цГМФ-зависимую протеинкиназу или протеинкиназу G, которая фосфорилирует ферменты, что сопровождается изменением их активности. Разрушается цГМФ, как и цАМФ, фосфодиэстеразой.


 Такая иодированная молекула тиреоглобулина секретируется в просвет фолликула, в коллоид. Когда в щитовидную железу приходит сигнал в виде ТТГ (тиреотропного гормона), клетки фолликула захватывают капельки коллоида вместе с тиреоглобулином, ферменты лизосом протеазы гидролизуют белок до аминокислот, и готовые Т3 и Т4 поступают в кровь.
В крови тиреоидные гормоны связываются с белком- переносчиком и в таком виде транспортируются в ткани-мишени. Концентрация Т4 в крови в 10 раз больше, чем Т3, поэтому Т4 называют главной формой тиреоидных гормонов в крови. Но Т3 в 10 раз
Такая иодированная молекула тиреоглобулина секретируется в просвет фолликула, в коллоид. Когда в щитовидную железу приходит сигнал в виде ТТГ (тиреотропного гормона), клетки фолликула захватывают капельки коллоида вместе с тиреоглобулином, ферменты лизосом протеазы гидролизуют белок до аминокислот, и готовые Т3 и Т4 поступают в кровь.
В крови тиреоидные гормоны связываются с белком- переносчиком и в таком виде транспортируются в ткани-мишени. Концентрация Т4 в крови в 10 раз больше, чем Т3, поэтому Т4 называют главной формой тиреоидных гормонов в крови. Но Т3 в 10 раз
Циклический ГМФ образуется из ГТФ под действием гуанилатциклазы по аналогии с синтезом цАМФ. Циклический ГМФ активирует специфическую цГМФ-зависимую протеинкиназу или протеинкиназу G, которая фосфорилирует ферменты, что сопровождается изменением их активности. Разрушается цГМФ, как и цАМФ, фосфодиэстеразой.
Кальций
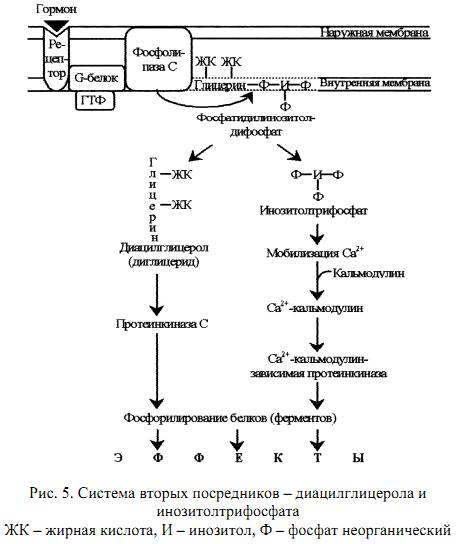
Концентрация Са2+ во внеклеточной жидкости в 10 000 раз больше, чем в цитоплазме. Такая концентрация кальция оказалась бы потенциально летальной для клетки. Клетка борется с избытком Са2+ в цитоплазме путём откачивания его наружу с помощью Са2+- АТФазы, расположенной в плазматической мембране. Под действием гормонального сигнала концентрация Са2+ в клетке возрастает в 10 и более раз за счет притока Са2+ из внеклеточной жидкости, а также выхода Са2+ из ЭПР и митохондрий. В клетке Са2+ связывается с белком кальмодулином и активирует специфическую Са2+-кальмодулин-зависимую протеинкиназу. В ГЛ 2+ ответ на гормональный сигнал и повышение концентрации Са в клетке специфическая протеинкиназа катализирует фосфорилирование множества внутриклеточных ферментов, регулируя, тем самым, их активность.Инозитолтрифосфат и диглицерид
Эти вторые посредники являются производными фосфолипида мембран, фосфатидилинозитола. Указанный фосфолипид фосфорилируется в мембране под действием фермента фосфатидилинозитолкиназы при участии 2 молекул АТФ (рис. 4. . Образующийся фосфатидилинозитолдифосфат расщепляется под действием специфического мембраносвязянного фермента фосфолипазы С, в результате чего образуются два вторых посредника: диглицерид (диацилглицерол, ДАГ) и инозитолтрифосфат (ИФ3.) Биологические эффекты этих двух вторых посредников реализуются по-разному. ДАГ активирует Са2+-зависимую протеинкиназу С, которая катализирует фосфорилирование внутриклеточных ферментов, изменяя их активность. ИФ3 связывается со специфическим рецептором на ЭПР, способствуя выходу из него Са2+ в цитоплазму. Далее Са2+ связывается с кальмодулином и активирует Са2+-кальмодулин-зависимую протеинкиназу, которая катализирует фосфорилирование ферментов, изменяя их активность (рис. 5.)
Механизм действия стероидных гормонов и тироксина
Рецепторы к этим гормонам находятся внутри клетки, в цитоплазме. Гормон легко и быстро проникает через мембрану внутрь клетки, взаимодействует с рецептором с образованием гормон-рецепторного комплекса. Этот комплекс переносится в ядро, где связывается с ядерным хроматином в определенном участке ДНК. С этого участка ДНК начинается синтез специфических мРНК, которые затем выходят в цитоплазму и служат матрицей для синтеза ферментов или других белков, необходимых клетке и обеспечивающих ответ клетки на действие гормона (рис. 6. . Таким образом, под действием пептидных гормонов и адреналина в клетке изменяется активность ферментов; стероидные гормоны и тироксин приводят к изменению количества ферментов.
Гормоны щитовидной железы
В щитовидной железе синтезируются тиреоидные гормоны – тироксин (Т4. и трииодтиронин (Т3. . Для синтеза этих гормонов необходим иод, который активно захватывается из крови клетками фолликулов щитовидной железы. Тироксин и трииодтиронин являются производными аминокислоты тирозина. Тироксин содержит в своей молекуле 4 атома иода, в состав трииодтиронина входит 3 атома иода. В клетках эпителия фолликулов щитовидной железы находится белок тиреоглобулин. Это гликопротеин, содержащий много остатков аминокислоты тирозина (около 3 % от массы белка). Синтез тиреоидных гормонов идет из тирозина и атомов иода именно в составе молекулы тиреоглобулина и включает 2 стадии. На апикальных мембранах клеток фолликулов сначала происходит иодирование тирозина с образованием моноиодтирозина (МИТ) и дииодтирозина (ДИТ). Следующим этапом является конденсация МИТ и ДИТ с образованием Т3 и Т4 (рис. 7. .
Такая иодированная молекула тиреоглобулина секретируется в просвет фолликула, в коллоид. Когда в щитовидную железу приходит сигнал в виде ТТГ (тиреотропного гормона), клетки фолликула захватывают капельки коллоида вместе с тиреоглобулином, ферменты лизосом протеазы гидролизуют белок до аминокислот, и готовые Т3 и Т4 поступают в кровь.
В крови тиреоидные гормоны связываются с белком- переносчиком и в таком виде транспортируются в ткани-мишени. Концентрация Т4 в крови в 10 раз больше, чем Т3, поэтому Т4 называют главной формой тиреоидных гормонов в крови. Но Т3 в 10 раз- 1
- 2
- 3
- 4
- 5
- . . .
- последняя (13) »
Последние комментарии
22 часов 53 минут назад
23 часов 11 минут назад
23 часов 20 минут назад
23 часов 22 минут назад
23 часов 24 минут назад
23 часов 42 минут назад