Следы былых биосфер, или Рассказ о том, как устроена биосфера и что осталось от биосфер геологического прошлого [Андрей Витальевич Лапо] (fb2) читать онлайн
- Следы былых биосфер, или Рассказ о том, как устроена биосфера и что осталось от биосфер геологического прошлого 1.3 Мб, 225с. скачать: (fb2) - (исправленную) читать: (полностью) - (постранично) - Андрей Витальевич Лапо
[Настройки текста] [Cбросить фильтры]
[Оглавление]
Следы былых биосфер, или Рассказ о том, как устроена биосфера и что осталось от биосфер геологического прошлого
Предисловие
Более 100 лет назад, в 1875 г., впервые появилось понятие о сфере жизни и был введен в обиход термин «биосфера». Полвека спустя наш великий соотечественник академик В. И. Вернадский вложил в него строго научное содержание. Своими работами В. И. Вернадский доказал планетное значение жизни, ее основную роль в механизме земной коры, гидросферы и нижних слоев атмосферы. Значение этих работ огромно и не может быть переоценено. В настоящее время они интенсивно развиваются как в нашей стране, так и за рубежом. «Следы былых биосфер, или Рассказ о том, как устроена биосфера и что осталось от биосфер геологического прошлого» — одно из самых удачных изложений проблем биосферы, сделанных за последние годы. Его автор, геолог А. В. Лапо, излагает и анализирует взгляды как самого В. И. Вернадского, так и целой плеяды других ученых, зачастую воскрешая талантливые, но забытые ныне работы отечественных исследователей. Это свидетельствует о большой эрудиции автора и о его бережном и уважительном отношении к первоисточникам. Однако самое ценное в книге — это синтез достижений современной науки по широкому кругу вопросов, касающихся проблем биосферы, ее развития и взаимодействия различных типов вещества, в биосфере. Чрезвычайно важным представляется выделение автором на первое место основной функции живого вещества — энергетической, которую выполняют хлорофиллоносные растения. Хотя, по существу, все функции живого вещества в той или иной мере воздействуют на среду, это не снимает необходимости выделения особой средообразующей функции, как это справедливо делается автором. Книга насыщена обильным фактическим материалом, показывающим огромную роль живого вещества в геологических процессах, что непременно должно учитываться при проектировании инженерных сооружений. Многочисленные примеры такого рода читатель найдет в книге. Автор обсуждает и малоразработанный вопрос о вертикальном расчленении биосферы на экогоризонты, опираясь на предложенное А. И. Перельманом деление на две зоны: фитосферу и редусферу, которые Н. Б. Вассоевич заменил терминами «фотобиосфера» и «мелабиосфера», а А. В. Лапо предложил последнюю называть, на мой взгляд, более удачно — меланобиосфера. В целом определения этих сфер достаточно обоснованны, но жизнь на суше вносит в это деление существенные поправки. Еще в палеозое основными компонентами биоценозов, осуществляющими роль трансформатора солнечной энергии в химическую, стали высшие растения, которые в большинстве случаев состоят из двух неразрывно связанных частей: верхних — фотосинтезирующих и нижних — всасывающих (корневые системы), расположенных в почве. Таким образом, жизнь высших растений связана с двумя сферами — фотобиосферой и меланобиосферой. Роющие животные — грызуны, а также некоторые хищники проводят свою жизнь в обеих сферах. Книга А. В. Лапо, впервые изданная в 1979 г. на русском языке и в 1982 г. — на английском, вызвала многочисленные отклики в печати. Второе издание значительно переработано и дополнено с учетом пожеланий, высказанных в рецензиях. Удачно использованы многие новейшие публикации, а также материалы научных конференций, посвященных 120‑летию со дня рождения В. И. Вернадского, в организации которых в Ленинграде деятельное участие принимал автор «Следов былых биосфер». Книга А. В. Лапо, вновь предлагаемая читателям, не только популяризирует достижения современной науки, но и содействует научному рассмотрению биосферы в экологическом аспекте. Пожелаем же доброго пути новому изданию этой увлекательной книги.Е. М. ЛАВРЕНКО, почетный президент Всесоюзного ботанического общества, академик
Вступление. Странная судьба
Отец умел понимать все.В наше время необычайно расширился круг знаний о явлениях и процессах, происходящих в окружающем мире. Вместе с тем человечество впервые стало ощущать не только масштабы своей деятельности, но и свою зависимость от состояния окружающей среды. Поэтому в центре внимания современного естествознания оказались всеобщая взаимосвязь и взаимообусловленность природных явлений, а ведущей тенденцией развития науки стал синтез достижений частных наук. Наиболее ярко эта тенденция к синтезу проявилась во взаимопроникновении наук о живой и неживой природе. Возникло учение о биосфере — особая наука, не сводимая ни к географии, ни к биологии, но использующая их достижения и, в свою очередь, влияющая на развитие геологии. Основоположником этой науки был гениальный русский ученый Владимир Иванович Вернадский (1863—1945). «В. И. Вернадский не просто обогатил обширную область знания, которую ныне называют науками о Земле, — пишут А. Л. Яншин, С. Р. Микулинский и И. И. Мочалов, — но и настолько преобразовал их теоретические основы, что без его учения о биосфере и биогеохимических процессах, о роли живого вещества в эволюции нашей планеты сегодня нельзя представить этих наук»[1]. В этой книге рассказывается о биосфере и о роли жизни в геологических процессах, а также об ученых, которые разрабатывали эти проблемы, в первую очередь об академике Вернадском. Удивительной фигурой предстает Владимир Иванович Вернадский в естествознании XX в. Казалось, он не ведал о делении науки на отрасли (а в современном естествознании их более тысячи). Вернадский был натуралистом в широком смысле этого слова, может быть, последним в истории науки. Он не был по образованию ни биологом, ни географом, но биологи ставят его в один ряд с Дарвином и Павловым и помещают его произведения в свои хрестоматии, а географы считают одним ил создателей современной географии. Труды Вернадского не просто основополагающие в какой-то отрасли — Вернадский закладывал новые науки. Их по крайней мере четыре: радиогеология, биогеохимия, учение о биосфере и науковедение. Вместе с В. М. Гольдшмидтом и А. Е. Ферсманом Вернадского считают также основоположником геохимии. Называют имя Вернадского и как основоположника генетической минералогии, учения о симметрии и диссимметрии, учения о естественных производительных силах. Ломоносов XX века… Будучи глубоким теоретиком, Вернадский обладал и несравненным организационным даром. Он был инициатором создания Комиссии по изучению естественных производительных сил России, одним из организаторов Академии наук УССР и ее первым президентом. По инициативе Владимира Ивановича в системе Академии наук СССР были созданы Институт географии, Институт минералогии и геохимии им. М. В. Ломоносова, Радиевый, Керамический и Оптический институты, Биогеохимическая лаборатория, Комиссия по изучению вечной мерзлоты и Комиссия по истории знаний, Комитет по метеоритам, Комиссии по изотопам, урану и многие другие. Многие из этих лабораторий и комиссий сейчас превратились в академические институты. Наконец, Вернадскому принадлежит идея создания Международной комиссии по определению геологического возраста Земли. Основанные Вернадским биогеохимия и учение о биосфере — новые науки, но они активно воздействуют на ранее сформировавшиеся отрасли естествознания. В «Ботаническом журнале» как-то появилась статья «Жизненные формы растений в аспекте учения о биосфере», а на одном из научных заседаний известный советский биолог, президент Гидробиологического общества, член-корреспондент АН СССР Г. Г. Винберг выступил с докладом «Идеи Вернадского в современной лимнологии». Лимнология же — наука об озерах, которой В. И. Вернадский никогда специально не занимался. Странная судьба — идеи Вернадского способствуют развитию смежных отраслей, в которых сам Владимир Иванович не работал. Дело, видимо, в том, что Вернадский не только закладывал основы наук и формулировал плодотворные идеи. Альберт Эйнштейн как-то сказал: «Наука — это попытка привести хаотическое многообразие нашего чувственного опыта в соответствие с некоторой единой системой мышления». Вернадский создал такую систему. Эта система мышления, или, если хотите, глобальная концепция, основана на определяющей роли жизни в геологических процессах. Она была создана Вернадским для геологического прошлого, но — странная судьба! — она стала актуальной, даже злободневной для нас, людей конца второго тысячелетия. Именно эта концепция является методологической основой разработки программы по охране окружающей среды. «Странная судьба, — писал Владимир Иванович жене в 1889 г. — Больше всего меня прельщали, с одной стороны, вопросы исторической жизни человечества и, с другой — философская сторона математических наук. И я не пошел ни по той, ни по другой отрасли. Не пошел по истории, потому что хотел раньше получить подготовку естественноисторическую и потом перейти на историю, не пошел по математике, так как не верил и не верю в свои математические способности…»[2] И все-таки он очень рано выбрал свой путь. Володя Вернадский на свое семнадцатилетие просил отца подарить ему английское издание одной из книг Дарвина (он уже тогда читал на нескольких языках). Отец подарил ему что-то другое — видимо, считая, что сыну рано читать такие серьезные книги, — но Володя так расстроился, что отец вынужден был уступить. Эта книга с надписью «Любимому сыну» сейчас хранится в Кабинете-музее В. И. Вернадского в Москве. Еще в молодые годы Вернадский сформулировал вопросы, над которыми думал всю жизнь. Сохранилась рукопись доклада, сделанного Вернадским на заседании студенческого Научно-литературного общества Петербургского университета в декабре 1884 г. Там есть такие слова: «Но что такое жизнь? И мертва ли материя, которая находится в вечном непрерывном законном движении, где происходит бесконечное разрушение и созидание, где нет покоя? Неужели только едва заметная пленка на бесконечно малой точке мироздания — Земле обладает коренными, особенными свойствами, а всюду и везде царит смерть?.. Покуда можно только предлагать эти вопросы. Их решение дастся рано или поздно наукой». «Феномен Вернадского» сформировала энциклопедичность интересов Владимира Ивановича. Его ближайшими друзьями были люди разных специальностей. Хорошо знавших его людей поражало умение В. И. Вернадского не только слушать, но и «разговаривать» собеседника, причем это были не пустые разговоры. С Д. С. Рождественским Вернадский говорил о физике ядра, с Н. Я. Марром — о яфетической теории, с. Д. Д. Плетневым — о научных основах кардиологии, с Н. И. Вавиловым — об окраске растений и о сортах пшеницы. Постоянным собеседником Вернадского был Иван Петрович Павлов, с которым (бывает же!) они жили на одной площадке академического дома. Поистине Вернадский «умел понимать все». В год своего 80‑летия на вопрос анкеты «Что наиболее характерного и наиболее ценного усматриваете Вы в организации Вами вашего труда как ученого?» Вернадский ответил: «Я думаю, что скорее всего — систематичность и стремление понять окружающее. Кроме того, я придаю огромное значение вопросам этики»[3]. Откроем «Биогеохимические очерки» В. И. Вернадского. Над авторским предисловием — посвящение: «Биогеохимические очерки» я посвящаю моей жене Наталье Егоровне, урожденной Старицкой, с которой мы прожили более пятидесяти лет вместе, моей помощнице в работе, всегда неуклонно относившейся к жизни, как к делу любви к людям и к свободному исканию истины». Нужно ли говорить, что такое понимание смысла жизни — главный этический принцип самого Владимира Ивановича? Вернадский не был бы Вернадским, если бы он не умел в высшей степени организовывать свой труд. Умение Владимира Ивановича использовать малейшие «кусочки времени» (это его выражение) отмечают многие современники. Он придавал этому огромное значение и передавал умение беречь время своим детям. И еще одна характерная деталь (из той же анкеты): «Я владею (для чтения) всеми славянскими, романскими и германскими языками»[4]. Странная судьба! «Мы обращаемся к нему не реже, а даже, пожалуй, чаще, чем при его жизни; в особенности в ходе научной работы, где его мысли и оценки всегда сопутствуют нам» — так писал о Вернадском один из его современников. Труды Владимира Ивановича поражают не только глубоким знанием предмета, но и обилием фактического материала и (теперь мы понимаем — почему!) исторических экскурсов. Некоторые из трудов Вернадского таковы, что написать их под силу, кажется, только целому коллективу. А писал их один человек, причем человек немолодой (основные труды Вернадского о биосфере написаны, когда ему было уже за 60). Не всегда это давалось легко. «Если ты наблюдаешь лиц, работающих научно, ты замечаешь, как часто они находятся в раздраженном состоянии вследствие хода и сути работы, или как часто они не могут заставить себя работать, потому что вся их воля ушла в мелкой, напряженной работе и им надо опять собраться с силами…» — это из другого письма жене[5]. У него был ясный взгляд философа и ребенка, быстрая походка и стройная, в старости немного сутуловатая фигура. Фотографироваться Вернадский не любил, и фотографий его осталось немного. Почему-то мало в воспоминаниях современников и описаний его внешности. Лучшее дал академик Д. В. Наливкин. Относится оно к 1914 г., когда Владимиру Ивановичу было немного за 50: «Узкое, точеное лицо, высокий выпуклый лоб ученого, темные волосы с сединой, каскадами поднимавшиеся над ним, поражали и удивляли. Но и они были только фоном для глаз, необычайно чистых, ясных и глубоких. Казалось, что в них светился весь облик, вся душа этого необыкновенного человека»[6]. Живой облик Вернадского удачно передают живописные портреты, выполненные хорошо знавшей его художницей Е. С. Зарудной-Кавос (1861—1917). Один из них впервые публикуется в этой книге. Вернадский умер в 1945 г. на 82‑м году жизни. При жизни Вернадского понимали не всегда. Но по отношению к Вернадскому и историческая перспектива становится обратной: достаточно сравнить оценку его творческого наследия в разные годы. В 1927 г. классический труд Вернадского «Биосфера» был оценен рецензентом в таких выражениях: «Геологи спокойно слушали академика Вернадского ранее, молчаливо выслушивают и еще одно его антиисторическое выступление… Довольно интересная книжка Вернадского требует все же к себе критического отношения. Изложена она несколько тяжеловатым стилем. Теряет она также и на том, что недостаточно выпячены и сконцентрированы основные важнейшие мысли. Однако интерес, который книжка представляет, все же должен побудить изучать ее». Бывали рецензии и похлеще, а геологи, действительно, «спокойно слушали» академика Вернадского. Работы Вернадского о роли жизни в геологических процессах в 30‑е годы цитировались редко, а в некрологе Вернадского даже не упоминалось о том, что он создал учение о биосфере. Вопросами биогеохимии в то время занималась только организованная Вернадским Биогеохимическая лаборатория — БИОГЕЛ (сейчас из нее вырос Институт геохимии и аналитической химии АН СССР, который носит имя Владимира Ивановича). Грандиозность идей Вернадского была осознана только после его смерти. Академик Александр Евгеньевич Ферсман (1883—1945), один из ближайших сподвижников Вернадского, прозванный друзьями за свою потрясающую энергию (и за фигуру) «шаровой молнией», писал в 1945 г.: «Десятилетиями, целыми столетиями будут изучаться и углубляться его гениальные идеи, а в трудах его — открываться новые страницы, служащие источником новых исканий… Еще не время углубиться в его огромный архив и многочисленные записи его биографии; еще много лет придется поработать и его ученикам, и историкам естествознания, чтобы выявить основные пути его научного творчества, разгадать сложные, еще непонятные построения его текста. Эта задача лежит на будущих поколениях…»[7] Перед Вернадским уже преклонялись, но его еще не понимали. Сейчас пришло время понимания. Создана Комиссия по разработке научного наследия Вернадского, выпущено собрание его избранных трудов. Из печати продолжают выходить книги, не опубликованные при жизни Вернадского: «Химическое строение биосферы и ее окружения» (1965), два тома «Размышлений натуралиста» (1975, 1977), «Живое вещество» (1978), третий выпуск «Проблем биогеохимии» (1980), труды по истории науки (1981), «Страницы автобиографии» (1981), переписка с Б. Л. Личковым (1979, 1980) и А. Е. Ферсманом (1985)… Написанные 50—70 лет назад, они по сей день остаются новыми по своим идеям, по глубине проработки научного материала, по широте его обобщения. Ежегодно в день рождения Вернадского в институте его имени проводятся мемориальные чтения, на которых выступают с докладами крупнейшие советские и зарубежные ученые. Академия наук СССР присуждает золотые медали и премии имени Вернадского. В 1984 г. на 27‑м Международном геологическом конгрессе в Москве я был свидетелем любопытного эпизода, когда известный советский микробиолог, член-корреспондент АН СССР Георгий Александрович Заварзин начал свой доклад с демонстрации слайда с цитатой из «Биосферы» В. И. Вернадского, а кончил его — после 40 минут изложения собственного фактического материала! — следующей фразой (цитирую свою запись): «Я очень сожалею, что мне не удалось добавить ничего нового к словам Вернадского, опубликованным в 1926 году». В современном естествознании Вернадский остается лидером. Завидная судьба…Н. В. Толль-Вернадская
Глава первая. Биосфера
Века и тысячелетия прошли, пока человеческая мысль могла отметить черты единого связного механизма в кажущейся хаотической картине природы.В. И. Вернадский, 1926
 Небольшая книга Вернадского «Биосфера» впервые вышла в Ленинграде в 1926 г. Издана она была тиражом всего 2 тыс. экз. Казалось бы, чисто научный труд, — но им зачитывались Михаил Пришвин и Николай Заболоцкий, а Геннадий Гор писал: «Есть книги, воздействие которых на читателя ни с чем не сравнимо. Такой необыкновенной книгой оказалась для меня «Биосфера»…»
В этой книге, впоследствии неоднократно переиздававшейся на русском[8] и других языках, биосфера Земли впервые была показана как единая динамическая система, управляемая жизнью. «Развитое В. И. Вернадским представление о биосфере как о пронизанной, преобразованной и постоянно преобразуемой организмами оболочке земного шара — одно из крупнейших обобщений естествознания нашего века», — свидетельствует наш современник, академик М. С. Гиляров.
Термин «биосфера» появился в научной литературе в 1875 г. Его автором был Эдуард Зюсс (1831—1914), с которым В. И. Вернадский был знаком лично, — известный австрийский геолог, «обобщитель геологических фактов», как называл его академик В. А. Обручев. Зюсс писал: «Одно кажется чужеродным на этом большом, состоящем из сфер небесном теле (Земле. — А. Л.), а именно — органическая жизнь… На поверхности материков можно выделить самостоятельную биосферу…»
Сконструировав новый термин, которому было суждено такое блестящее будущее, и, по существу, вводя в науку новое понятие, Э. Зюсс не дал ему никакого определения. Слово «биосфера» стало использоваться в геологической и географической литературе от случая к случаю, причем каждый понимал его по-своему.
В научной, популярной и даже учебной литературе в качестве автора термина «биосфера» иногда указывается знаменитый французский естествоиспытатель Жан-Батист Ламарк (1744—1829). Это неверно: он ввел термин «биология» (а не «биосфера»). При этом, опередив свое время больше чем на столетие, Ламарк вплотную подошел к понятию биосферы и к осознанию планетарной роли жизни: «Сложные минеральные вещества всех видов, образующие внешнюю кору земного шара и встречающиеся там в виде отдельных скоплений, рудных тел, параллельных пластов и т. д. и образующие низменности, холмы, долины и горы, являются исключительно продуктами животных и растений, которые существовали на этих участках поверхности земного шара» (курсив Ламарка). Эти пророческие слова Ламарка были надолго забыты.
Вкладом принципиальной важности в разработку современного понимания биосферы явились и труды знаменитого немецкого естествоиспытателя-энциклопедиста Александра Гумбольдта (1769—1859). Именно Гумбольдт впервые сумел взглянуть на Землю как на единое целое. Этот глобальный (как мы теперь говорим) подход Гумбольдт развил в своем фундаментальном труде «Космос», над которым работал всю свою долгую жизнь. В нем, в частности, развивалась идея о повсеместном распространении жизни («всеоживленности» Земли), ее неразрывной связи с неорганическим миром (мысль, совершенно новая для XIX в.!) и даже был употреблен термин «жизнесфера» — правда, единожды и без всяких комментариев. Вернадский высоко ценил труды Гумбольдта: «Его постановка проблемы… приближается к геохимическим концепциям нашего времени. Для него живое вещество есть неразрывная и закономерная часть поверхности планеты, неотделимая от ее химической среды»[9].
Наконец, непосредственным предшественником Вернадского и его учителем был основоположник современного генетического почвоведения Василий Васильевич Докучаев (1846—1903), который первым осознал опасность дробления науки о природе на множество частных дисциплин. По мнению Докучаева, естествознание достигло больших успехов в изучении таких объектов природы, как живые организмы, минералы, горные породы и т. д. Однако изучались в то время именно отдельные объекты, «но не их соотношения, не та генетическая, вековечная и всегда закономерная связь, какая существует между силами, телами и явлениями, между мертвой и живой природой. А между тем именно эти соотношения, эти закономерные взаимодействия и составляют сущность познания естества, ядро истинной натурфилософии — лучшую и высшую прелесть естествознания».
Докучаев сконцентрировал свое внимание на почве: естественном теле, где происходит взаимодействие всех трех выделявшихся тогда «царств природы»: минералов, растений и животных. На примере почвы он всесторонне рассмотрел взаимодействие биотических и абиотических факторов в естественных телах. Поэтому, хотя термин «биосфера» в литературу ввел не Докучаев, а Зюсс, именно Докучаева считают предтечей современного учения о биосфере — естественном теле более высокого ранга, чем почва.
Творчески развив идеи своих предшественников — Ламарка, Гумбольдта, Докучаева — и использовав, по существу, «бесхозный» термин Зюсса, Вернадский создал принципиально новый подход к явлениям жизни. «Формирование интегративного подхода на уровне биосферы, как и само введение этого уровня организации живой материи, — целиком заслуга В. И. Вернадского», — пишет А. М. Гиляров.
Обыденно говоря, биосфера — это то, что нас окружает, та «природа», в которой мы живем. Вернадский в разных своих работах дал несколько определений биосферы, везде подчеркивая две ее отличительные особенности. Первая из них следующая: «Биосфера представляет оболочку жизни — область существования живого вещества»[10], вторая: «Биосфера может быть рассматриваема как область земной коры, занятая трансформаторами, переводящими космические излучения в действенную земную энергию — электрическую, химическую, механическую, тепловую и т. д.»[11].
В своих работах В. И. Вернадский подчеркивал «всюдность» жизни. В самом деле, заселенными оказываются самые невероятные, казалось бы, местообитания: термальные источники, температура воды в которых достигает точки кипения, а в некоторых случаях даже несколько превышает ее — и вечные снега Гималаев, где на высоте 8300 м обитают по крайней мере 9 видов бактерий; безводные пустыни (в славящихся своей сухостью африканских пустынях живут, например, более 500 видов насекомых) — и сверхсоленые озера, где процветают цианобактерии, архебактерии и один из видов креветок, а жгутиковые и диатомовые водоросли хоть и влачат жалкое существование, но все же существуют! Заселено организмами даже… Мертвое море — в нем обитает несколько видов архебактерий и водорослей (не следует ли это название писать в кавычках: так называемое «Мертвое море»?).
Для учителя В. И. Вернадского — В. В. Докучаева — биотический и абиотический факторы были равноправными партнерами формирования почвы. Вернадский же, перейдя на глобальный уровень, показал, что ведущим фактором, преобразующим лик Земли, является жизнь. Ее особенность заключается не только в ускорении химических реакций — некоторые реакции вне организмов вообще не происходят при нормальных температурах и давлениях. Хорошо об этом сказал Л. С. Берг: «Организмы осуществляют нечто с физической точки зрения невероятное». Так, жиры и углеводы окисляются в организме при температуре около 37°, а вне его — при температурах 400÷500°C. Синтез аммиака из молекулярного азота в промышленных условиях осуществляют при температуре 500° и давлении 300—350 атм. А микроорганизмы без особых затруднений проводят эту реакцию при обычной температуре и атмосферном давлении. Значит, в живых организмах должны развиваться какие-то особые катализаторы, ускоряющие протекание химических процессов. Такие белковые катализаторы, действительно, были найдены в живых организмах и получили название «ферментов». Действуют они очень специализированно; фермент и вещество, подверженное его действию, подходят друг к другу, как ключ к замку. Для протекания химических процессов достаточно ничтожных количеств ферментов. Во всем мире не наберется и десятка килограммов нитрогеназы — удивительного фермента, используемого организмами для синтеза азотистых соединений из атмосферного азота. Академик И. П. Павлов называл ферменты «возбудителями жизни».
Биосфера Земли представляет собой глобальную открытую систему со своими «входом» и «выходом». Ее «вход» — это поток солнечной энергии, поступающий из космоса, «выход» — те образованные в процессе жизнедеятельности организмов вещества, которые в силу каких-либо причин ускользнули из биотического круговорота (иногда — на многие миллионы лет). Образно говоря, это выход в «геологию».
На языке современной науки биосферу называют саморегулируемой кибернетической системой, обладающей свойствами гомеостата. 50 лет назад таких терминов еще не существовало, и Вернадский говорил в этом смысле об «организованности биосферы» (а в более ранних работах — о ее «механизме»).
Одно из наиболее характерных проявлений организованности биосферы Вернадский видел в наличии озонового экрана, находящегося за пределами биосферы и поглощающего губительные для жизни ультрафиолетовые лучи. Состав газовой оболочки нашей планеты полностью регулируется жизнью.
Другой пример саморегулирования представляет собой Мировой океан. Реки ежегодно вносят в океан 1,5 млн. т карбоната кальция, а солевой состав океанической воды существенно не меняется. Почему? Организмы используют эти карбонаты для построения своих скелетов, а после их отмирания карбонаты осаждаются на дно. Так путем создания «кальциевых покровов» нашей планеты стабилизируется состав океанских вод. Этот механизм действует в биосфере уже многие миллионы лет.
Таким образом, саморегулирование биосферы Земли обеспечивается живыми организмами. Это позволяет считать биосферу централизованной кибернетической системой. Под таким названием объединяют системы, в которых один элемент (или подсистема) играет доминирующую роль в функционировании системы в целом. Этот элемент называют ведущей частью системы или ее центром. Живые организмы в биосфере и играют роль такого центра.
Согласно закону необходимого разнообразия Эшби кибернетическая система только тогда обладает устойчивостью для блокирования внешних и внутренних возмущений, когда она имеет достаточное внутреннее разнообразие. Земля как планета характеризуется значительным разнообразием природных условий. Это определяется ее шарообразной формой, ее движением вокруг Солнца и вокруг своей оси, что, в свою очередь, обусловливает широтное и сезонное изменение интенсивности поступления солнечной энергии; значительное разнообразие природных условий создается и расчлененным рельефом Земли. Амплитуда абсолютных отметок поверхности Земли составляет больше 19 км — от +8848 м — гора Джомолунгма (Эверест) до −11022 м — Марианская впадина в Тихом океане. Но основное разнообразие биосферы Земли создается живыми организмами.
Интереснейшее обобщение (до сих пор недооцененное) о создании организмами неоднородности среды было сделано в начале нашего века русским микробиологом, профессором Михаилом Андреевичем Егуновым (1864—1937). «Всякая среда, населенная живыми организмами, есть среда биоанизотропная. Биоанизотропия есть явление общее; биоизотропии нет, — писал Егунов. — Это вытекает из того, что между средой и каждым организмом происходит непрерывный обмен веществ, и поэтому в каждый данный момент различные точки среды отличаются друг от друга по физико-химическому составу. Диффузия никогда не может вполне выравнять эти различия, пока существует причина, производящая их».
Считают, что в современной биосфере представлено около 2 млн. видов живых организмов (а за все время существования биосферы их было не меньше миллиарда), причем каждый из этих видов, в свою очередь, включает в себя миллионы и миллиарды особей, дисперсно распределенных в пространстве. Недавно подсчитали, например, что в Приангарье на площади 0,23 км² живет 535 (!) видов беспозвоночных, каждый из которых, естественно, по-своему взаимодействует с окружающей средой. Деятельность живых организмов и создает удивительное разнообразие окружающей нас «природы» — биосферы. Пожалуй, мы не могли до последнего времени оценить это разнообразие — нам просто не с чем было сравнивать нашу биосферу. И лишь теперь, когда мы увидели на экранах телевизоров, на страницах иллюстрированных изданий пейзажи иных планет, лишенных жизни, — лишь теперь мы можем в полной мере оценить «внутреннее разнообразие» биосферы. Оно служит своеобразной гарантией сохранения жизни на нашей планете.
Нам сейчас трудно представить себе масштабы геологических катастроф, происходивших в истории Земли. Самые страшные из известных нам землетрясений сопровождались сбросами амплитудой 4—6 м. Тектонические явления такого масштаба расцениваются как национальное бедствие — вспомним страшное перуанское землетрясение 1970 г., унесшее около 100 тыс. жизней. А в геологических отложениях известны сбросы амплитудой в несколько километров! Неисчислимые бедствия несут и вулканические извержения. Наиболее крупное из них произошло в начале этого века на Аляске. Высота утеса, образованного застывшей лавой, достигает здесь 100 метров! А при извержении на острове Кракатау в 1883 г. выпал слой пепла толщиной от 2 до 60 м.
Сколько их было в истории биосферы, геологических катастроф! Тем не менее все крупные катастрофы, когда-либо случавшиеся в биосфере (включая оледенения), всегда носили локальный, а не глобальный характер. Правда, в последнее время на этот счет появились некоторые сомнения. Причиной этого послужило то обстоятельство, что в ряде районов земного шара (Дания, Италия, Испания, Новая Зеландия, впадины Тихого и Атлантического океанов) в пограничных отложениях между мелом и палеогеном был обнаружен тонкий (всего несколько сантиметров) прослой, резко обогащенный иридием. Поскольку иридий является одним из характерных элементов метеоритов, было высказано предположение, что прослой, обогащенный иридием, образовался за счет вещества астероида, который взорвался при столкновении с нашей планетой. С этим событием стали связывать массовые вымирания многих групп организмов (в первую очередь динозавров), характерные для данного отрезка геологической истории.
Для проверки этой гипотезы в Палеонтологическом институте АН СССР была проведена фундаментальная работа по детальному анализу смены фауны на границе мезозоя с кайнозоем. Это исследование показало, что вымирание представителей различных групп органического мира происходило достаточно постепенно и для разных групп не одновременно. Что же касается вымирания динозавров, то оно предшествовало появлению в разрезе иридиевого прослоя. Директор Палеонтологического института академик Леонид Петрович Татаринов сделал вывод, что вообще «ни в одном случае гипотеза о глобальной катастрофе как основной причине смены биоты в истории Земли не получила подтверждения на достаточно достоверном палеонтологическом материале».
А как же иридий? Ученые университета штата Мэриленд (США) недавно показали, что иридий может иметь и земное происхождение. Исследуя продукты выброса вулкана Килауэа, расположенного на одном из островов Гавайского архипелага, они обнаружили необычайно высокую концентрацию в них иридия. При этом было доказано, что иридий накапливался не в излившихся при извержении лавах, а поступал с вулканическим пеплом и газами в атмосферу, что и обеспечивало широкое его площадное рассеивание. Правда, масштабы поступления иридия при извержении Килауэа не были особенно значительными. Однако нельзя забывать, что в конце мезозоя происходили грандиозные излияния базальтов. Не исключено, что именно они привели к резкому накоплению иридия сначала в атмосфере, а затем — и к его концентрации в четко ограниченном прослое. Кстати, прослой с иридием присутствует не везде: например, в детально изученных рубежных отложениях мела и палеогена в Крыму его так и не нашли, как ни искали.
Таким образом, весь материал, накопленный геологией, показывает непрерывность развития органического мира Земли в течение всей геологической истории и правильность вывода В. И. Вернадского об отсутствии в земной коре азойных (сформировавшихся в отсутствие жизни) отложений. Внутреннее разнообразие биосферы обеспечило ее устойчивость даже по отношению к самым значительным катастрофическим потрясениям. Она — эта устойчивость — определяется исключительным разнообразием населяющих биосферу живых организмов и почти безгранично большим их количеством, взаимозаменяемостью составляющих ее экосистем, дублированием отдельных звеньев биогеохимических циклов, жизненной устойчивостью и активностью особей и т. д.
Характерной особенностью биосферы является мозаичность ее строения. Жизнь, будучи организована в планетарном масштабе, функционирует в пределах отдельных «квантов» биосферы. В 1935 г. английским ученым Артуром Дж. Тэнсли (1871—1955) они были названы экосистемами. Согласно современному определению Д. В. Панфилова, «экосистемы — это комплексы взаимосвязанных популяций разных видов живых существ и изменяемой ими абиотической среды, обладающие способностью к саморегуляции и самовозобновлению всех главных компонентов их биоты». Размеры экосистемы очень различны: «от кочки до оболочки», по шутливому выражению географа и писателя Юрия Константиновича Ефремова. Их протяженность на суше, например, варьирует от нескольких метров (песчаные дюны, микродепрессии в степях и полупустынях, небольшие блюдечки озер в тундрах) до нескольких километров (солончаки, однородные участки степей, лесов и т. д.). Экосистемой глобального масштаба является биосфера Земли. Подобно биосфере, экосистемы всех рангов биоцентричны.
В советской научной литературе иногда употребляется также термин «биогеоценоз», предложенный в 1940 г. академиком Владимиром Николаевичем Сукачевым (1880—1967). Согласно его определению, биогеоценоз — это участок биосферы, через который не проходит ни одна существенная биоценотическая, микроклиматическая, гидрологическая, почвенная, геоморфологическая или геохимическая граница. Впоследствии биогеоценоз стали определять как экосистему в пределах фитоценоза, считая, что категории экосистемы и биогеоценоза совпадают на уровне растительного сообщества. По существу же, эти понятия близки и различаются главным образом в деталях. При этом понятие экосистемы является более гибким, и в дальнейшем изложении мы будем пользоваться главным образом этим термином.
Несмотря на свою относительную замкнутость, экосистемы тесно связаны между собой. Тому имеется много доказательств. Самое простое: мы круглый год потребляем кислород, а выделяется он растениями только во время вегетационного периода, который лишь в тропическом поясе продолжается круглогодично. В остальное время мы расходуем кислород, поставляемый растениями противоположного полушария. Есть и более печальные свидетельства глобальной интегрированности биосферного круговорота: в теле антарктических пингвинов найден ДДТ (который, конечно, никогда не применялся в Антарктиде), а в молоке европейских женщин радиоактивный стронций появлялся регулярно через четыре месяца после каждого испытания атомного оружия на атоллах Тихого океана.
Основными элементами, участвующими в биосферном круговороте, являются водород, кислород, углерод, азот, кальций, калий, кремний, фосфор, сера, стронций, барий, железо, марганец, цинк, молибден, медь и никель. Круговороты химических элементов, вызванные деятельностью живого вещества, получили название биогеохимических циклов. Их исследованию в настоящее время уделяется большое внимание в связи с проблемами охраны окружающей среды.
Гетерогенность строения биосферы, ее «мозаичность» определяются и наличием в ней регионов с различной биогеохимической специализацией или, как их первоначально называли, биогеохимических провинций. Это понятие было введено в 1936 г. В. И. Вернадским и А. П. Виноградовым в их совместном докладе «Геохимические провинции и заболевания» и в дальнейшем развито последним. По определению Виноградова, под биогеохимическими провинциями понимаются области на поверхности Земли, различающиеся по содержанию (в их почвах, водах и т. п.) химических элементов (или соединений), с которыми связаны определенные биологические реакции со стороны местной флоры и фауны. Впоследствии членом-корреспондентом ВАСХНИЛ Виктором Владиславовичем Ковальским (1899—1984) была составлена карта биогеохимического районирования СССР, первая редакция которой была опубликована в 1954 г., а последняя — в 1982 г.
Глобальный биогеохимический круговорот в биосфере не является замкнутым. Степень воспроизводства отдельных циклов достигает 90—98%. В масштабе геологического времени неполная замкнутость биогеохимических циклов приводит к дифференциации элементов и накоплению их в атмосфере, гидросфере или метабиосфере Земли. Эти несколько процентов вещества, ускользающие из круговорота, и составляют тот «выход в геологию», о котором мы уже упоминали.
Однако «геология» стоит не только на «выходе», но и на «входе» биогеохимического круговорота. Одним из первых обратил на это внимание знаменитый немецкий геолог Иоганнес Вальтер (1860—1937), который писал: «Биосфера образует своеобразную переходную зону между атмосферой, гидросферой и литосферой. Углекислота воздуха в виде твердых ископаемых углей так же могла принимать участие в составе земной коры, как и когда-то растворенная в воде известь…» Развивая это положение на материале современных данных, известный советский геохимик, лауреат Золотой медали им. В. И. Вернадского, член-корреспондент АН СССР Александр Борисович Ронов пришел к выводу о значительной «открытости» биогеохимического круговорота и о необходимости постоянного поступления в него углекислоты из недр. А. Б. Ронов сформулировал следующий «геохимический принцип сохранения жизни»: «Жизнь на Земле и других планетах при прочих равных условиях возможна лишь до тех пор, пока эти планеты активны и происходит обмен энергией и веществом между их недрами и поверхностью».
Непрерывному круговороту в биосфере подвергаются только вещества, в то время как для энергии можно говорить лишь о направленном потоке. Поступающая в биосферу солнечная энергия частично расходуется на синтез органического вещества. Биосфера — это «фабрика макромолекул»: фотоавтотрофные организмы, поглощая солнечную энергию, путем фотосинтеза превращают низкомолекулярные, бедные энергией неорганические вещества в высокомолекулярные, богатые энергией органические соединения и снабжают ими все живое.
Передаваясь с одного трофического уровня на другой, энергия постепенно рассеивается. После окончательного разложения органических остатков энергия частично накапливается в земной коре в виде алюмосиликатов, которые академик Н. В. Белов (1891—1982) назвал «геохимическими аккумуляторами». Говоря о пронизывании внешней оболочки Земли солнечной энергией, Н. В. Белов привел пример с бриллиантом. Когда на бриллиант падает свет, одна часть его отражается от граней драгоценного камня, а другая попадает внутрь и способна вырваться наружу только после многократных отражений от внутренних граней. Такого же рода странствиям (в масштабе геологического времени) подвергается и солнечная энергия, аккумулированная живым веществом.
Для иллюстрации различий превращений энергии и вещества в биосфере известные советские исследователи и популяризаторы науки — безвременно скончавшийся Петр Петрович Второв (1939—1979) и Николай Николаевич Дроздов, которого мы часто видим на экранах телевизоров в качестве ведущего передачи «В мире животных», — использовали наглядный образ — водяную мельницу. Колесо ее вертится и вертится, оставаясь на месте, и символизирует запас вещества в биосфере. Однако для того, чтобы колесо вертелось, нужен постоянный приток воды. Подобно этому, поток солнечной энергии, поступающий из космоса, крутит «колесо жизни» на нашей планете.
Насколько же быстро крутится это колесо? Во времена Вернадского это не было известно, однако сейчас на такой вопрос уже можно ответить.
Обновление всего живого вещества биосферы Земли осуществляется в среднем за 8 лет. При этом вещество наземных растений (фитомасса суши) обновляется примерно за 14 лет. В океане циркуляция вещества происходит во много раз быстрее: вся масса живого вещества обновляется за 33 дня, в то время как фитомасса океана — каждый день! Процесс полной смены вод в гидросфере осуществляется за 2800 лет. В атмосфере смена кислорода происходит за несколько тысячлет, а углекислого газа — за 6,3 года. Эти цифры показывают, что геохимический эффект деятельности живого вещества в биосфере проявляется не только в течение геологического времени (миллионы и миллиарды лет), но ясно выражен даже в пределах времени исторического (тысячи лет и менее).
Наряду с этим некоторые другие вещества, участвующие в биогеохимическом круговороте, имеют значительно меньшие скорости миграции. Так, время, необходимое для фотосинтетического разложения всей массы воды Мирового океана, исчисляется 5—6 млн. лет (выше мы говорили о круговороте воды без ее химического разложения). Миллионолетиями измеряется также продолжительность общепланетарных циклов углерода, азота и фосфора.
В 1926 г. Вернадский впервые поставил вопрос о границах биосферы; он вернулся к нему в специальной статье в 1937 г.[12] В то время, однако, трудно было дать на него определенный ответ. Непросто это сделать и сейчас.
Н. Б. Вассоевич обратил внимание на важную особенность строения биосферы, которую ранее подчеркивал Вернадский: на существование в биосфере «поля устойчивости жизни» и «поля существования жизни». В первом случае (в «поле устойчивости») существуют «условия, которые выдерживает жизнь, не прекращая своих функций, т. е. те, при которых организм хотя и страдает, но выживает», а во втором (в «поле существования») — «условия, при которых организм может давать потомство, т. е. увеличивать живую массу — увеличивать действенную энергию планеты… Пределы биосферы обусловлены прежде всего полем существования жизни»[13].
Рассмотрим, какие из реально существующих на Земле физико-химических условий ограничивают развитие жизни, а значит, и определяют границы биосферы.
Во-первых, это достаточное количество углекислого газа и кислорода. Установлено, что на Гималаях зона распространения зеленой растительности ограничена высотой 6200 м, где парциальное давление углекислого газа вдвое ниже, чем на поверхности моря. Однако и выше жизнь окончательно не замирает — там встречаются некоторые виды пауков и насекомых. Питаются они пыльцой и другими органическими остатками, заносимыми ветром.
Во-вторых, это достаточное количество воды (причем обязательно — в жидкой фазе!), которое могло бы обеспечить нормальный ход процессов жизнедеятельности. На поверхности Земли участки, где жизнь была бы ограничена этим фактором, встречаются чрезвычайно редко.
В-третьих, благоприятный термический режим, исключающий как слишком высокие температуры (вызывающие свертывание белков), так и слишком низкие (прекращающие работу ферментов). Чемпионами по выживанию являются прокариоты (речь о них впереди). Некоторые их виды живут на снегу, другие — в горячих источниках при температуре выше 100°.
В-четвертых, это наличие «прожиточного минимума» элементов минерального питания — фактор, в значительной мере ограничивающий развитие жизни на больших площадях в океанах, но крайне редко сводящий ее к нулю.
В-пятых, сверхсоленость водной среды, превышающая концентрацию солей в морской воде примерно в 10 раз. Искусственные садки, где производится выпаривание рассолов, стерильны. Лишены жизни и подземные воды с концентрацией солей свыше 270 г/л.
Указанные выше факторы ограничивают развитие жизни лишь на очень небольших территориях земного шара. В качестве примеров известный американский эколог Роберт Э. Риклефс приводит следующие участки: склоны горы Мак-Кинли на Аляске, где из-за круглогодично сохраняющихся минусовых температур вода существует только в твердой фазе, и заповедник Уайт-Сандс (штат Нью-Мексико, США), где чистые гипсовые пески, дающие название заповеднику (по-английски оно обозначает «Белые пески»), совершенно лишены элементов минерального питания. Впрочем, их безжизненность является проблематичной и уж во всяком случае эфемерной. Так, другие участки, которые Р. Э. Риклефс считает безжизненными, — Долина Смерти (штат Калифорния, США) и вулканический остров Суртсей, образовавшийся в 1963 г. вблизи Исландии, — уже заселены прокариотами.
Но, может быть, безжизненными являются океанские пучины?
До середины XIX в. господствовало твердое убеждение, что в океане на значительной глубине жизнь отсутствует. «Нуль животной жизни» проводили на глубине 540 м. Нижележащие слои водной толщи считали безжизненными и называли азойными (от греч. отрицания «а» и «зоон» — животное). В ходе последующих исследований, однако, «нуль животной жизни» приходилось опускать все ниже и ниже, пока в 40—50‑е годы нашего века результаты экспедиций на судах «Витязь» (СССР) и «Галатея» (Дания) окончательно не доказали, что океан заселен до максимальных своих глубин. Правда, живыми глубоководные организмы никто не видел — до того момента, когда 23 января 1960 г. батискаф «Триест» не коснулся дна Марианской впадины. В точке погружения глубина океана достигала 10 919 м; температура воды за бортом составляла 2,4°C, а давление — приблизительно 1100 атм.
Репортаж ведет один из участников погружения Жак Пикар: «Дно было совершенно ровным, если не считать нескольких комочков. Но — самое главное — за миг до соприкосновения с дном в световой круг вплыла рыба… В одну секунду, вместившую годы труда и подготовки, мы разрешили проблему, десятки лет мучившую океанографов… Мы добыли доказательства того, что ни давление, ни тьма, ни холод, вместе взятые, не в силах остановить жизнь». Миф о «нуле животной жизни» окончательно канул в Лету. А наиболее обычными обитателями максимальных глубин океана оказались все же не рыбы, а голотурии — своеобразные, довольно примитивные беспозвоночные. По описанию Джералда Даррелла, это — «самые противные среди всех обитателей моря». Их называют «морскими огурцами», хотя некоторые из них скорее напоминают тыквы или раздувшиеся сосиски, покрытые толстой бородавчатой кожей бурого цвета. Наверно, можно согласиться с Дарреллом, что они не первые красавцы на нашей планете, но ведь и жизнь в вечном мраке при давлении больше 1000 атм тоже далеко не мед…
А как обстоят дела в полярных морях? Обитаема ли их водная толща, изолированная от внешнего мира толстым панцирем льда? Исследования, проведенные в 1937 г. одним из членов папанинской четверки — Петром Петровичем Ширшовым (1905—1953), впоследствии академиком, — показали, что воды Северного Ледовитого океана заселены и в самых высоких широтах. А 40 лет спустя на другом конце Земли — в прибрежных водах Антарктиды, покрытых вечными льдами, — американскими учеными было сделано еще одно поразительное открытие.
Исследования проходили в море Росса, толщина льда на котором достигала 420 м. Морские воды, как оказалось, там были изолированы ледяным панцирем от солнечного света и прямого воздействия атмосферы по крайней мере в течение 120 тыс. лет. Естественно было предположить, что при таких условиях жизнь в море невозможна. Но, как сказал Козьма Прутков, «глядя на мир, нельзя не удивляться» — ледяная темница оказалась обитаемой! Телевизионные камеры и другие инструменты исследования, спущенные в прорубь, обнаружили в водной толще довольно разнообразное сообщество организмов, состоявшее из диатомовых водорослей, фораминифер, ракообразных и бактерий. Среди них были встречены и совершенно необычные, ранее не известные науке виды.
Итак, вся поверхность суши (за исключением очень ограниченных участков) и океанские пучины соответствуют «полю существования» жизни в понимании В. И. Вернадского и, следовательно, относятся к биосфере. Что же касается верхних слоев атмосферы, то здесь распространен только так называемый «аэропланктон» — бактерии, дрожжевые грибки, споры плесневых грибов, мхов и лишайников, а также вирусы, водоросли, цисты простейших и т. д. Жизнеспособные микроорганизмы найдены на высоте до 77 км. Большинство микроорганизмов, однако, гибнет на меньшей высоте в первые минуты или даже секунды после того, как попадает (не по своей воле!) в воздушную стихию. Для остающихся в живых микроорганизмов воздушная среда также не является благоприятной, и они впадают в состояние анабиоза. Поэтому вся атмосфера в целом не отвечает определению экосистемы — здесь нет ни саморегуляции системы, ни самовозобновления организмов. Это то «поле устойчивости жизни», о котором писал В. И. Вернадский, — зона, примыкающая к биосфере сверху. Американский ученый Дж. Э. Хатчинсон[14] назвал ее парабиосферой (от греч. «пара» — у, возле, при).
Где же проходят реальные границы биосферы?
Конкретизируя определение В. И. Вернадского, к биосфере можно отнести те зоны Земли, где существуют аборигенные сообщества живых организмов. Удовлетворяют ли этому условию приземные слои атмосферы? Видимо, да — ведь здесь стабильно обитают насекомые и птицы. Среди насекомых есть активные хищники, которые живут охотой в воздухе, а на землю опускаются только от случая к случаю и на ночлег. В свою очередь, среди птиц много насекомоядных, охотящихся за своей добычей налету. Пример тому — стрижи, которым длинные крылья затрудняют взлет. Это обстоятельство заставляет их большую часть жизни проводить в беспосадочном полете. В воздухе они не только кормятся, но и пьют, купаются, играют свадьбы…
Значит, зону обитания насекомых и птиц следует причислять к биосфере, а верхнюю ее границу проводить по рекордным их залетам (пока рекордсменом является безвременно погибший белоголовый сип, столкнувшийся с самолетом на высоте 12,5 км). Выше располагается парабиосфера.
Сложнее обстоит дело с нижней границей. Владимир Иванович предполагал, что вся осадочная оболочка Земли заселена бактериями. Более поздние работы, однако, не подтвердили это предположение. Оказалось, что распределение микроорганизмов в подземных водах — а, значит, и нижняя граница биосферы в пределах континентов — определяется температурой вод, режимом их циркуляции и концентрацией в них минеральных солей. Живые бактерии выявляются в подземных водах с температурой до 100°, хотя наиболее активная их жизнедеятельность ограничена «только» 80°. Предельная концентрация минеральных солей уже указывалась: 270 г/л. При глубоком бурении в Поволжье и Западной Сибири активная и разнообразная по составу анаэробная микрофлора была найдена на глубине 1—3 км, а иногда и глубже. В то же время, если минерализация вод превышает указанный предел или если залежь «запечатана», бактерии отсутствуют и на меньших глубинах. Такие лишенные жизни (или «азойные») зоны были обнаружены в Ангаро-Ленском бассейне уже на глубине около 500 м, а в Волго-Камском — на глубине около 1200 м.
Что же касается донных осадков Мирового океана и внутриконтинентальных водоемов, то здесь нижняя граница распространения живых организмов определяется содержанием в среде кислорода. Показано, например, что в зоне сероводородного заражения Черного моря осадки оказываются стерильными уже на глубине 5 см от поверхности дна. В Тихом и Индийском океанах микрофлора обитает в донных осадках до глубины 10—12 м, а в Каспийском море — не менее чем на 114 м ниже поверхности морского дна. Существует предположение (правда, пока не доказанное), что заселенным может оказаться 200—250‑метровый слой донных осадков.
Мощность биосферы по вертикали, таким образом, составляет в океане всю толщу океанских вод и значительно варьирующую по толщине донную пленку жизни, а на континентах — тонкий наземный и мощный подземный слой. Вся дневная поверхность нашей планеты относится к зоне биосферы. Единственные (да и то сомнительные) исключения составляют те немногие случаи, о которых мы упоминали выше.
Распределение живых организмов в биосфере показано на рис. 1.
Небольшая книга Вернадского «Биосфера» впервые вышла в Ленинграде в 1926 г. Издана она была тиражом всего 2 тыс. экз. Казалось бы, чисто научный труд, — но им зачитывались Михаил Пришвин и Николай Заболоцкий, а Геннадий Гор писал: «Есть книги, воздействие которых на читателя ни с чем не сравнимо. Такой необыкновенной книгой оказалась для меня «Биосфера»…»
В этой книге, впоследствии неоднократно переиздававшейся на русском[8] и других языках, биосфера Земли впервые была показана как единая динамическая система, управляемая жизнью. «Развитое В. И. Вернадским представление о биосфере как о пронизанной, преобразованной и постоянно преобразуемой организмами оболочке земного шара — одно из крупнейших обобщений естествознания нашего века», — свидетельствует наш современник, академик М. С. Гиляров.
Термин «биосфера» появился в научной литературе в 1875 г. Его автором был Эдуард Зюсс (1831—1914), с которым В. И. Вернадский был знаком лично, — известный австрийский геолог, «обобщитель геологических фактов», как называл его академик В. А. Обручев. Зюсс писал: «Одно кажется чужеродным на этом большом, состоящем из сфер небесном теле (Земле. — А. Л.), а именно — органическая жизнь… На поверхности материков можно выделить самостоятельную биосферу…»
Сконструировав новый термин, которому было суждено такое блестящее будущее, и, по существу, вводя в науку новое понятие, Э. Зюсс не дал ему никакого определения. Слово «биосфера» стало использоваться в геологической и географической литературе от случая к случаю, причем каждый понимал его по-своему.
В научной, популярной и даже учебной литературе в качестве автора термина «биосфера» иногда указывается знаменитый французский естествоиспытатель Жан-Батист Ламарк (1744—1829). Это неверно: он ввел термин «биология» (а не «биосфера»). При этом, опередив свое время больше чем на столетие, Ламарк вплотную подошел к понятию биосферы и к осознанию планетарной роли жизни: «Сложные минеральные вещества всех видов, образующие внешнюю кору земного шара и встречающиеся там в виде отдельных скоплений, рудных тел, параллельных пластов и т. д. и образующие низменности, холмы, долины и горы, являются исключительно продуктами животных и растений, которые существовали на этих участках поверхности земного шара» (курсив Ламарка). Эти пророческие слова Ламарка были надолго забыты.
Вкладом принципиальной важности в разработку современного понимания биосферы явились и труды знаменитого немецкого естествоиспытателя-энциклопедиста Александра Гумбольдта (1769—1859). Именно Гумбольдт впервые сумел взглянуть на Землю как на единое целое. Этот глобальный (как мы теперь говорим) подход Гумбольдт развил в своем фундаментальном труде «Космос», над которым работал всю свою долгую жизнь. В нем, в частности, развивалась идея о повсеместном распространении жизни («всеоживленности» Земли), ее неразрывной связи с неорганическим миром (мысль, совершенно новая для XIX в.!) и даже был употреблен термин «жизнесфера» — правда, единожды и без всяких комментариев. Вернадский высоко ценил труды Гумбольдта: «Его постановка проблемы… приближается к геохимическим концепциям нашего времени. Для него живое вещество есть неразрывная и закономерная часть поверхности планеты, неотделимая от ее химической среды»[9].
Наконец, непосредственным предшественником Вернадского и его учителем был основоположник современного генетического почвоведения Василий Васильевич Докучаев (1846—1903), который первым осознал опасность дробления науки о природе на множество частных дисциплин. По мнению Докучаева, естествознание достигло больших успехов в изучении таких объектов природы, как живые организмы, минералы, горные породы и т. д. Однако изучались в то время именно отдельные объекты, «но не их соотношения, не та генетическая, вековечная и всегда закономерная связь, какая существует между силами, телами и явлениями, между мертвой и живой природой. А между тем именно эти соотношения, эти закономерные взаимодействия и составляют сущность познания естества, ядро истинной натурфилософии — лучшую и высшую прелесть естествознания».
Докучаев сконцентрировал свое внимание на почве: естественном теле, где происходит взаимодействие всех трех выделявшихся тогда «царств природы»: минералов, растений и животных. На примере почвы он всесторонне рассмотрел взаимодействие биотических и абиотических факторов в естественных телах. Поэтому, хотя термин «биосфера» в литературу ввел не Докучаев, а Зюсс, именно Докучаева считают предтечей современного учения о биосфере — естественном теле более высокого ранга, чем почва.
Творчески развив идеи своих предшественников — Ламарка, Гумбольдта, Докучаева — и использовав, по существу, «бесхозный» термин Зюсса, Вернадский создал принципиально новый подход к явлениям жизни. «Формирование интегративного подхода на уровне биосферы, как и само введение этого уровня организации живой материи, — целиком заслуга В. И. Вернадского», — пишет А. М. Гиляров.
Обыденно говоря, биосфера — это то, что нас окружает, та «природа», в которой мы живем. Вернадский в разных своих работах дал несколько определений биосферы, везде подчеркивая две ее отличительные особенности. Первая из них следующая: «Биосфера представляет оболочку жизни — область существования живого вещества»[10], вторая: «Биосфера может быть рассматриваема как область земной коры, занятая трансформаторами, переводящими космические излучения в действенную земную энергию — электрическую, химическую, механическую, тепловую и т. д.»[11].
В своих работах В. И. Вернадский подчеркивал «всюдность» жизни. В самом деле, заселенными оказываются самые невероятные, казалось бы, местообитания: термальные источники, температура воды в которых достигает точки кипения, а в некоторых случаях даже несколько превышает ее — и вечные снега Гималаев, где на высоте 8300 м обитают по крайней мере 9 видов бактерий; безводные пустыни (в славящихся своей сухостью африканских пустынях живут, например, более 500 видов насекомых) — и сверхсоленые озера, где процветают цианобактерии, архебактерии и один из видов креветок, а жгутиковые и диатомовые водоросли хоть и влачат жалкое существование, но все же существуют! Заселено организмами даже… Мертвое море — в нем обитает несколько видов архебактерий и водорослей (не следует ли это название писать в кавычках: так называемое «Мертвое море»?).
Для учителя В. И. Вернадского — В. В. Докучаева — биотический и абиотический факторы были равноправными партнерами формирования почвы. Вернадский же, перейдя на глобальный уровень, показал, что ведущим фактором, преобразующим лик Земли, является жизнь. Ее особенность заключается не только в ускорении химических реакций — некоторые реакции вне организмов вообще не происходят при нормальных температурах и давлениях. Хорошо об этом сказал Л. С. Берг: «Организмы осуществляют нечто с физической точки зрения невероятное». Так, жиры и углеводы окисляются в организме при температуре около 37°, а вне его — при температурах 400÷500°C. Синтез аммиака из молекулярного азота в промышленных условиях осуществляют при температуре 500° и давлении 300—350 атм. А микроорганизмы без особых затруднений проводят эту реакцию при обычной температуре и атмосферном давлении. Значит, в живых организмах должны развиваться какие-то особые катализаторы, ускоряющие протекание химических процессов. Такие белковые катализаторы, действительно, были найдены в живых организмах и получили название «ферментов». Действуют они очень специализированно; фермент и вещество, подверженное его действию, подходят друг к другу, как ключ к замку. Для протекания химических процессов достаточно ничтожных количеств ферментов. Во всем мире не наберется и десятка килограммов нитрогеназы — удивительного фермента, используемого организмами для синтеза азотистых соединений из атмосферного азота. Академик И. П. Павлов называл ферменты «возбудителями жизни».
Биосфера Земли представляет собой глобальную открытую систему со своими «входом» и «выходом». Ее «вход» — это поток солнечной энергии, поступающий из космоса, «выход» — те образованные в процессе жизнедеятельности организмов вещества, которые в силу каких-либо причин ускользнули из биотического круговорота (иногда — на многие миллионы лет). Образно говоря, это выход в «геологию».
На языке современной науки биосферу называют саморегулируемой кибернетической системой, обладающей свойствами гомеостата. 50 лет назад таких терминов еще не существовало, и Вернадский говорил в этом смысле об «организованности биосферы» (а в более ранних работах — о ее «механизме»).
Одно из наиболее характерных проявлений организованности биосферы Вернадский видел в наличии озонового экрана, находящегося за пределами биосферы и поглощающего губительные для жизни ультрафиолетовые лучи. Состав газовой оболочки нашей планеты полностью регулируется жизнью.
Другой пример саморегулирования представляет собой Мировой океан. Реки ежегодно вносят в океан 1,5 млн. т карбоната кальция, а солевой состав океанической воды существенно не меняется. Почему? Организмы используют эти карбонаты для построения своих скелетов, а после их отмирания карбонаты осаждаются на дно. Так путем создания «кальциевых покровов» нашей планеты стабилизируется состав океанских вод. Этот механизм действует в биосфере уже многие миллионы лет.
Таким образом, саморегулирование биосферы Земли обеспечивается живыми организмами. Это позволяет считать биосферу централизованной кибернетической системой. Под таким названием объединяют системы, в которых один элемент (или подсистема) играет доминирующую роль в функционировании системы в целом. Этот элемент называют ведущей частью системы или ее центром. Живые организмы в биосфере и играют роль такого центра.
Согласно закону необходимого разнообразия Эшби кибернетическая система только тогда обладает устойчивостью для блокирования внешних и внутренних возмущений, когда она имеет достаточное внутреннее разнообразие. Земля как планета характеризуется значительным разнообразием природных условий. Это определяется ее шарообразной формой, ее движением вокруг Солнца и вокруг своей оси, что, в свою очередь, обусловливает широтное и сезонное изменение интенсивности поступления солнечной энергии; значительное разнообразие природных условий создается и расчлененным рельефом Земли. Амплитуда абсолютных отметок поверхности Земли составляет больше 19 км — от +8848 м — гора Джомолунгма (Эверест) до −11022 м — Марианская впадина в Тихом океане. Но основное разнообразие биосферы Земли создается живыми организмами.
Интереснейшее обобщение (до сих пор недооцененное) о создании организмами неоднородности среды было сделано в начале нашего века русским микробиологом, профессором Михаилом Андреевичем Егуновым (1864—1937). «Всякая среда, населенная живыми организмами, есть среда биоанизотропная. Биоанизотропия есть явление общее; биоизотропии нет, — писал Егунов. — Это вытекает из того, что между средой и каждым организмом происходит непрерывный обмен веществ, и поэтому в каждый данный момент различные точки среды отличаются друг от друга по физико-химическому составу. Диффузия никогда не может вполне выравнять эти различия, пока существует причина, производящая их».
Считают, что в современной биосфере представлено около 2 млн. видов живых организмов (а за все время существования биосферы их было не меньше миллиарда), причем каждый из этих видов, в свою очередь, включает в себя миллионы и миллиарды особей, дисперсно распределенных в пространстве. Недавно подсчитали, например, что в Приангарье на площади 0,23 км² живет 535 (!) видов беспозвоночных, каждый из которых, естественно, по-своему взаимодействует с окружающей средой. Деятельность живых организмов и создает удивительное разнообразие окружающей нас «природы» — биосферы. Пожалуй, мы не могли до последнего времени оценить это разнообразие — нам просто не с чем было сравнивать нашу биосферу. И лишь теперь, когда мы увидели на экранах телевизоров, на страницах иллюстрированных изданий пейзажи иных планет, лишенных жизни, — лишь теперь мы можем в полной мере оценить «внутреннее разнообразие» биосферы. Оно служит своеобразной гарантией сохранения жизни на нашей планете.
Нам сейчас трудно представить себе масштабы геологических катастроф, происходивших в истории Земли. Самые страшные из известных нам землетрясений сопровождались сбросами амплитудой 4—6 м. Тектонические явления такого масштаба расцениваются как национальное бедствие — вспомним страшное перуанское землетрясение 1970 г., унесшее около 100 тыс. жизней. А в геологических отложениях известны сбросы амплитудой в несколько километров! Неисчислимые бедствия несут и вулканические извержения. Наиболее крупное из них произошло в начале этого века на Аляске. Высота утеса, образованного застывшей лавой, достигает здесь 100 метров! А при извержении на острове Кракатау в 1883 г. выпал слой пепла толщиной от 2 до 60 м.
Сколько их было в истории биосферы, геологических катастроф! Тем не менее все крупные катастрофы, когда-либо случавшиеся в биосфере (включая оледенения), всегда носили локальный, а не глобальный характер. Правда, в последнее время на этот счет появились некоторые сомнения. Причиной этого послужило то обстоятельство, что в ряде районов земного шара (Дания, Италия, Испания, Новая Зеландия, впадины Тихого и Атлантического океанов) в пограничных отложениях между мелом и палеогеном был обнаружен тонкий (всего несколько сантиметров) прослой, резко обогащенный иридием. Поскольку иридий является одним из характерных элементов метеоритов, было высказано предположение, что прослой, обогащенный иридием, образовался за счет вещества астероида, который взорвался при столкновении с нашей планетой. С этим событием стали связывать массовые вымирания многих групп организмов (в первую очередь динозавров), характерные для данного отрезка геологической истории.
Для проверки этой гипотезы в Палеонтологическом институте АН СССР была проведена фундаментальная работа по детальному анализу смены фауны на границе мезозоя с кайнозоем. Это исследование показало, что вымирание представителей различных групп органического мира происходило достаточно постепенно и для разных групп не одновременно. Что же касается вымирания динозавров, то оно предшествовало появлению в разрезе иридиевого прослоя. Директор Палеонтологического института академик Леонид Петрович Татаринов сделал вывод, что вообще «ни в одном случае гипотеза о глобальной катастрофе как основной причине смены биоты в истории Земли не получила подтверждения на достаточно достоверном палеонтологическом материале».
А как же иридий? Ученые университета штата Мэриленд (США) недавно показали, что иридий может иметь и земное происхождение. Исследуя продукты выброса вулкана Килауэа, расположенного на одном из островов Гавайского архипелага, они обнаружили необычайно высокую концентрацию в них иридия. При этом было доказано, что иридий накапливался не в излившихся при извержении лавах, а поступал с вулканическим пеплом и газами в атмосферу, что и обеспечивало широкое его площадное рассеивание. Правда, масштабы поступления иридия при извержении Килауэа не были особенно значительными. Однако нельзя забывать, что в конце мезозоя происходили грандиозные излияния базальтов. Не исключено, что именно они привели к резкому накоплению иридия сначала в атмосфере, а затем — и к его концентрации в четко ограниченном прослое. Кстати, прослой с иридием присутствует не везде: например, в детально изученных рубежных отложениях мела и палеогена в Крыму его так и не нашли, как ни искали.
Таким образом, весь материал, накопленный геологией, показывает непрерывность развития органического мира Земли в течение всей геологической истории и правильность вывода В. И. Вернадского об отсутствии в земной коре азойных (сформировавшихся в отсутствие жизни) отложений. Внутреннее разнообразие биосферы обеспечило ее устойчивость даже по отношению к самым значительным катастрофическим потрясениям. Она — эта устойчивость — определяется исключительным разнообразием населяющих биосферу живых организмов и почти безгранично большим их количеством, взаимозаменяемостью составляющих ее экосистем, дублированием отдельных звеньев биогеохимических циклов, жизненной устойчивостью и активностью особей и т. д.
Характерной особенностью биосферы является мозаичность ее строения. Жизнь, будучи организована в планетарном масштабе, функционирует в пределах отдельных «квантов» биосферы. В 1935 г. английским ученым Артуром Дж. Тэнсли (1871—1955) они были названы экосистемами. Согласно современному определению Д. В. Панфилова, «экосистемы — это комплексы взаимосвязанных популяций разных видов живых существ и изменяемой ими абиотической среды, обладающие способностью к саморегуляции и самовозобновлению всех главных компонентов их биоты». Размеры экосистемы очень различны: «от кочки до оболочки», по шутливому выражению географа и писателя Юрия Константиновича Ефремова. Их протяженность на суше, например, варьирует от нескольких метров (песчаные дюны, микродепрессии в степях и полупустынях, небольшие блюдечки озер в тундрах) до нескольких километров (солончаки, однородные участки степей, лесов и т. д.). Экосистемой глобального масштаба является биосфера Земли. Подобно биосфере, экосистемы всех рангов биоцентричны.
В советской научной литературе иногда употребляется также термин «биогеоценоз», предложенный в 1940 г. академиком Владимиром Николаевичем Сукачевым (1880—1967). Согласно его определению, биогеоценоз — это участок биосферы, через который не проходит ни одна существенная биоценотическая, микроклиматическая, гидрологическая, почвенная, геоморфологическая или геохимическая граница. Впоследствии биогеоценоз стали определять как экосистему в пределах фитоценоза, считая, что категории экосистемы и биогеоценоза совпадают на уровне растительного сообщества. По существу же, эти понятия близки и различаются главным образом в деталях. При этом понятие экосистемы является более гибким, и в дальнейшем изложении мы будем пользоваться главным образом этим термином.
Несмотря на свою относительную замкнутость, экосистемы тесно связаны между собой. Тому имеется много доказательств. Самое простое: мы круглый год потребляем кислород, а выделяется он растениями только во время вегетационного периода, который лишь в тропическом поясе продолжается круглогодично. В остальное время мы расходуем кислород, поставляемый растениями противоположного полушария. Есть и более печальные свидетельства глобальной интегрированности биосферного круговорота: в теле антарктических пингвинов найден ДДТ (который, конечно, никогда не применялся в Антарктиде), а в молоке европейских женщин радиоактивный стронций появлялся регулярно через четыре месяца после каждого испытания атомного оружия на атоллах Тихого океана.
Основными элементами, участвующими в биосферном круговороте, являются водород, кислород, углерод, азот, кальций, калий, кремний, фосфор, сера, стронций, барий, железо, марганец, цинк, молибден, медь и никель. Круговороты химических элементов, вызванные деятельностью живого вещества, получили название биогеохимических циклов. Их исследованию в настоящее время уделяется большое внимание в связи с проблемами охраны окружающей среды.
Гетерогенность строения биосферы, ее «мозаичность» определяются и наличием в ней регионов с различной биогеохимической специализацией или, как их первоначально называли, биогеохимических провинций. Это понятие было введено в 1936 г. В. И. Вернадским и А. П. Виноградовым в их совместном докладе «Геохимические провинции и заболевания» и в дальнейшем развито последним. По определению Виноградова, под биогеохимическими провинциями понимаются области на поверхности Земли, различающиеся по содержанию (в их почвах, водах и т. п.) химических элементов (или соединений), с которыми связаны определенные биологические реакции со стороны местной флоры и фауны. Впоследствии членом-корреспондентом ВАСХНИЛ Виктором Владиславовичем Ковальским (1899—1984) была составлена карта биогеохимического районирования СССР, первая редакция которой была опубликована в 1954 г., а последняя — в 1982 г.
Глобальный биогеохимический круговорот в биосфере не является замкнутым. Степень воспроизводства отдельных циклов достигает 90—98%. В масштабе геологического времени неполная замкнутость биогеохимических циклов приводит к дифференциации элементов и накоплению их в атмосфере, гидросфере или метабиосфере Земли. Эти несколько процентов вещества, ускользающие из круговорота, и составляют тот «выход в геологию», о котором мы уже упоминали.
Однако «геология» стоит не только на «выходе», но и на «входе» биогеохимического круговорота. Одним из первых обратил на это внимание знаменитый немецкий геолог Иоганнес Вальтер (1860—1937), который писал: «Биосфера образует своеобразную переходную зону между атмосферой, гидросферой и литосферой. Углекислота воздуха в виде твердых ископаемых углей так же могла принимать участие в составе земной коры, как и когда-то растворенная в воде известь…» Развивая это положение на материале современных данных, известный советский геохимик, лауреат Золотой медали им. В. И. Вернадского, член-корреспондент АН СССР Александр Борисович Ронов пришел к выводу о значительной «открытости» биогеохимического круговорота и о необходимости постоянного поступления в него углекислоты из недр. А. Б. Ронов сформулировал следующий «геохимический принцип сохранения жизни»: «Жизнь на Земле и других планетах при прочих равных условиях возможна лишь до тех пор, пока эти планеты активны и происходит обмен энергией и веществом между их недрами и поверхностью».
Непрерывному круговороту в биосфере подвергаются только вещества, в то время как для энергии можно говорить лишь о направленном потоке. Поступающая в биосферу солнечная энергия частично расходуется на синтез органического вещества. Биосфера — это «фабрика макромолекул»: фотоавтотрофные организмы, поглощая солнечную энергию, путем фотосинтеза превращают низкомолекулярные, бедные энергией неорганические вещества в высокомолекулярные, богатые энергией органические соединения и снабжают ими все живое.
Передаваясь с одного трофического уровня на другой, энергия постепенно рассеивается. После окончательного разложения органических остатков энергия частично накапливается в земной коре в виде алюмосиликатов, которые академик Н. В. Белов (1891—1982) назвал «геохимическими аккумуляторами». Говоря о пронизывании внешней оболочки Земли солнечной энергией, Н. В. Белов привел пример с бриллиантом. Когда на бриллиант падает свет, одна часть его отражается от граней драгоценного камня, а другая попадает внутрь и способна вырваться наружу только после многократных отражений от внутренних граней. Такого же рода странствиям (в масштабе геологического времени) подвергается и солнечная энергия, аккумулированная живым веществом.
Для иллюстрации различий превращений энергии и вещества в биосфере известные советские исследователи и популяризаторы науки — безвременно скончавшийся Петр Петрович Второв (1939—1979) и Николай Николаевич Дроздов, которого мы часто видим на экранах телевизоров в качестве ведущего передачи «В мире животных», — использовали наглядный образ — водяную мельницу. Колесо ее вертится и вертится, оставаясь на месте, и символизирует запас вещества в биосфере. Однако для того, чтобы колесо вертелось, нужен постоянный приток воды. Подобно этому, поток солнечной энергии, поступающий из космоса, крутит «колесо жизни» на нашей планете.
Насколько же быстро крутится это колесо? Во времена Вернадского это не было известно, однако сейчас на такой вопрос уже можно ответить.
Обновление всего живого вещества биосферы Земли осуществляется в среднем за 8 лет. При этом вещество наземных растений (фитомасса суши) обновляется примерно за 14 лет. В океане циркуляция вещества происходит во много раз быстрее: вся масса живого вещества обновляется за 33 дня, в то время как фитомасса океана — каждый день! Процесс полной смены вод в гидросфере осуществляется за 2800 лет. В атмосфере смена кислорода происходит за несколько тысячлет, а углекислого газа — за 6,3 года. Эти цифры показывают, что геохимический эффект деятельности живого вещества в биосфере проявляется не только в течение геологического времени (миллионы и миллиарды лет), но ясно выражен даже в пределах времени исторического (тысячи лет и менее).
Наряду с этим некоторые другие вещества, участвующие в биогеохимическом круговороте, имеют значительно меньшие скорости миграции. Так, время, необходимое для фотосинтетического разложения всей массы воды Мирового океана, исчисляется 5—6 млн. лет (выше мы говорили о круговороте воды без ее химического разложения). Миллионолетиями измеряется также продолжительность общепланетарных циклов углерода, азота и фосфора.
В 1926 г. Вернадский впервые поставил вопрос о границах биосферы; он вернулся к нему в специальной статье в 1937 г.[12] В то время, однако, трудно было дать на него определенный ответ. Непросто это сделать и сейчас.
Н. Б. Вассоевич обратил внимание на важную особенность строения биосферы, которую ранее подчеркивал Вернадский: на существование в биосфере «поля устойчивости жизни» и «поля существования жизни». В первом случае (в «поле устойчивости») существуют «условия, которые выдерживает жизнь, не прекращая своих функций, т. е. те, при которых организм хотя и страдает, но выживает», а во втором (в «поле существования») — «условия, при которых организм может давать потомство, т. е. увеличивать живую массу — увеличивать действенную энергию планеты… Пределы биосферы обусловлены прежде всего полем существования жизни»[13].
Рассмотрим, какие из реально существующих на Земле физико-химических условий ограничивают развитие жизни, а значит, и определяют границы биосферы.
Во-первых, это достаточное количество углекислого газа и кислорода. Установлено, что на Гималаях зона распространения зеленой растительности ограничена высотой 6200 м, где парциальное давление углекислого газа вдвое ниже, чем на поверхности моря. Однако и выше жизнь окончательно не замирает — там встречаются некоторые виды пауков и насекомых. Питаются они пыльцой и другими органическими остатками, заносимыми ветром.
Во-вторых, это достаточное количество воды (причем обязательно — в жидкой фазе!), которое могло бы обеспечить нормальный ход процессов жизнедеятельности. На поверхности Земли участки, где жизнь была бы ограничена этим фактором, встречаются чрезвычайно редко.
В-третьих, благоприятный термический режим, исключающий как слишком высокие температуры (вызывающие свертывание белков), так и слишком низкие (прекращающие работу ферментов). Чемпионами по выживанию являются прокариоты (речь о них впереди). Некоторые их виды живут на снегу, другие — в горячих источниках при температуре выше 100°.
В-четвертых, это наличие «прожиточного минимума» элементов минерального питания — фактор, в значительной мере ограничивающий развитие жизни на больших площадях в океанах, но крайне редко сводящий ее к нулю.
В-пятых, сверхсоленость водной среды, превышающая концентрацию солей в морской воде примерно в 10 раз. Искусственные садки, где производится выпаривание рассолов, стерильны. Лишены жизни и подземные воды с концентрацией солей свыше 270 г/л.
Указанные выше факторы ограничивают развитие жизни лишь на очень небольших территориях земного шара. В качестве примеров известный американский эколог Роберт Э. Риклефс приводит следующие участки: склоны горы Мак-Кинли на Аляске, где из-за круглогодично сохраняющихся минусовых температур вода существует только в твердой фазе, и заповедник Уайт-Сандс (штат Нью-Мексико, США), где чистые гипсовые пески, дающие название заповеднику (по-английски оно обозначает «Белые пески»), совершенно лишены элементов минерального питания. Впрочем, их безжизненность является проблематичной и уж во всяком случае эфемерной. Так, другие участки, которые Р. Э. Риклефс считает безжизненными, — Долина Смерти (штат Калифорния, США) и вулканический остров Суртсей, образовавшийся в 1963 г. вблизи Исландии, — уже заселены прокариотами.
Но, может быть, безжизненными являются океанские пучины?
До середины XIX в. господствовало твердое убеждение, что в океане на значительной глубине жизнь отсутствует. «Нуль животной жизни» проводили на глубине 540 м. Нижележащие слои водной толщи считали безжизненными и называли азойными (от греч. отрицания «а» и «зоон» — животное). В ходе последующих исследований, однако, «нуль животной жизни» приходилось опускать все ниже и ниже, пока в 40—50‑е годы нашего века результаты экспедиций на судах «Витязь» (СССР) и «Галатея» (Дания) окончательно не доказали, что океан заселен до максимальных своих глубин. Правда, живыми глубоководные организмы никто не видел — до того момента, когда 23 января 1960 г. батискаф «Триест» не коснулся дна Марианской впадины. В точке погружения глубина океана достигала 10 919 м; температура воды за бортом составляла 2,4°C, а давление — приблизительно 1100 атм.
Репортаж ведет один из участников погружения Жак Пикар: «Дно было совершенно ровным, если не считать нескольких комочков. Но — самое главное — за миг до соприкосновения с дном в световой круг вплыла рыба… В одну секунду, вместившую годы труда и подготовки, мы разрешили проблему, десятки лет мучившую океанографов… Мы добыли доказательства того, что ни давление, ни тьма, ни холод, вместе взятые, не в силах остановить жизнь». Миф о «нуле животной жизни» окончательно канул в Лету. А наиболее обычными обитателями максимальных глубин океана оказались все же не рыбы, а голотурии — своеобразные, довольно примитивные беспозвоночные. По описанию Джералда Даррелла, это — «самые противные среди всех обитателей моря». Их называют «морскими огурцами», хотя некоторые из них скорее напоминают тыквы или раздувшиеся сосиски, покрытые толстой бородавчатой кожей бурого цвета. Наверно, можно согласиться с Дарреллом, что они не первые красавцы на нашей планете, но ведь и жизнь в вечном мраке при давлении больше 1000 атм тоже далеко не мед…
А как обстоят дела в полярных морях? Обитаема ли их водная толща, изолированная от внешнего мира толстым панцирем льда? Исследования, проведенные в 1937 г. одним из членов папанинской четверки — Петром Петровичем Ширшовым (1905—1953), впоследствии академиком, — показали, что воды Северного Ледовитого океана заселены и в самых высоких широтах. А 40 лет спустя на другом конце Земли — в прибрежных водах Антарктиды, покрытых вечными льдами, — американскими учеными было сделано еще одно поразительное открытие.
Исследования проходили в море Росса, толщина льда на котором достигала 420 м. Морские воды, как оказалось, там были изолированы ледяным панцирем от солнечного света и прямого воздействия атмосферы по крайней мере в течение 120 тыс. лет. Естественно было предположить, что при таких условиях жизнь в море невозможна. Но, как сказал Козьма Прутков, «глядя на мир, нельзя не удивляться» — ледяная темница оказалась обитаемой! Телевизионные камеры и другие инструменты исследования, спущенные в прорубь, обнаружили в водной толще довольно разнообразное сообщество организмов, состоявшее из диатомовых водорослей, фораминифер, ракообразных и бактерий. Среди них были встречены и совершенно необычные, ранее не известные науке виды.
Итак, вся поверхность суши (за исключением очень ограниченных участков) и океанские пучины соответствуют «полю существования» жизни в понимании В. И. Вернадского и, следовательно, относятся к биосфере. Что же касается верхних слоев атмосферы, то здесь распространен только так называемый «аэропланктон» — бактерии, дрожжевые грибки, споры плесневых грибов, мхов и лишайников, а также вирусы, водоросли, цисты простейших и т. д. Жизнеспособные микроорганизмы найдены на высоте до 77 км. Большинство микроорганизмов, однако, гибнет на меньшей высоте в первые минуты или даже секунды после того, как попадает (не по своей воле!) в воздушную стихию. Для остающихся в живых микроорганизмов воздушная среда также не является благоприятной, и они впадают в состояние анабиоза. Поэтому вся атмосфера в целом не отвечает определению экосистемы — здесь нет ни саморегуляции системы, ни самовозобновления организмов. Это то «поле устойчивости жизни», о котором писал В. И. Вернадский, — зона, примыкающая к биосфере сверху. Американский ученый Дж. Э. Хатчинсон[14] назвал ее парабиосферой (от греч. «пара» — у, возле, при).
Где же проходят реальные границы биосферы?
Конкретизируя определение В. И. Вернадского, к биосфере можно отнести те зоны Земли, где существуют аборигенные сообщества живых организмов. Удовлетворяют ли этому условию приземные слои атмосферы? Видимо, да — ведь здесь стабильно обитают насекомые и птицы. Среди насекомых есть активные хищники, которые живут охотой в воздухе, а на землю опускаются только от случая к случаю и на ночлег. В свою очередь, среди птиц много насекомоядных, охотящихся за своей добычей налету. Пример тому — стрижи, которым длинные крылья затрудняют взлет. Это обстоятельство заставляет их большую часть жизни проводить в беспосадочном полете. В воздухе они не только кормятся, но и пьют, купаются, играют свадьбы…
Значит, зону обитания насекомых и птиц следует причислять к биосфере, а верхнюю ее границу проводить по рекордным их залетам (пока рекордсменом является безвременно погибший белоголовый сип, столкнувшийся с самолетом на высоте 12,5 км). Выше располагается парабиосфера.
Сложнее обстоит дело с нижней границей. Владимир Иванович предполагал, что вся осадочная оболочка Земли заселена бактериями. Более поздние работы, однако, не подтвердили это предположение. Оказалось, что распределение микроорганизмов в подземных водах — а, значит, и нижняя граница биосферы в пределах континентов — определяется температурой вод, режимом их циркуляции и концентрацией в них минеральных солей. Живые бактерии выявляются в подземных водах с температурой до 100°, хотя наиболее активная их жизнедеятельность ограничена «только» 80°. Предельная концентрация минеральных солей уже указывалась: 270 г/л. При глубоком бурении в Поволжье и Западной Сибири активная и разнообразная по составу анаэробная микрофлора была найдена на глубине 1—3 км, а иногда и глубже. В то же время, если минерализация вод превышает указанный предел или если залежь «запечатана», бактерии отсутствуют и на меньших глубинах. Такие лишенные жизни (или «азойные») зоны были обнаружены в Ангаро-Ленском бассейне уже на глубине около 500 м, а в Волго-Камском — на глубине около 1200 м.
Что же касается донных осадков Мирового океана и внутриконтинентальных водоемов, то здесь нижняя граница распространения живых организмов определяется содержанием в среде кислорода. Показано, например, что в зоне сероводородного заражения Черного моря осадки оказываются стерильными уже на глубине 5 см от поверхности дна. В Тихом и Индийском океанах микрофлора обитает в донных осадках до глубины 10—12 м, а в Каспийском море — не менее чем на 114 м ниже поверхности морского дна. Существует предположение (правда, пока не доказанное), что заселенным может оказаться 200—250‑метровый слой донных осадков.
Мощность биосферы по вертикали, таким образом, составляет в океане всю толщу океанских вод и значительно варьирующую по толщине донную пленку жизни, а на континентах — тонкий наземный и мощный подземный слой. Вся дневная поверхность нашей планеты относится к зоне биосферы. Единственные (да и то сомнительные) исключения составляют те немногие случаи, о которых мы упоминали выше.
Распределение живых организмов в биосфере показано на рис. 1.
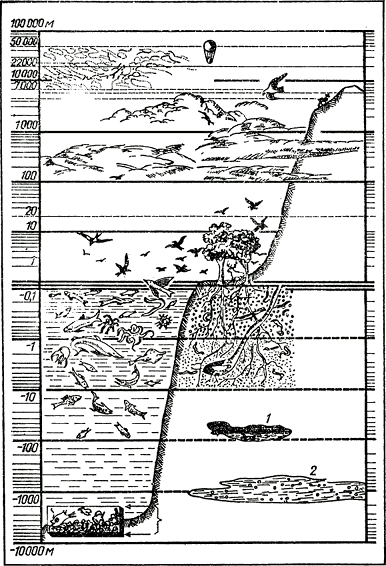 Рис. 1. Распределение живых организмов в биосфере (по Кадару, 1965): 1 — обитатели пещер; 2 — микроорганизмы подземных местообитаний; еще ниже — донная пленка жизни
Рис. 1. Распределение живых организмов в биосфере (по Кадару, 1965): 1 — обитатели пещер; 2 — микроорганизмы подземных местообитаний; еще ниже — донная пленка жизни
Таковы, по современным взглядам, границы биосферы Земли. Однако Вернадский главную книгу своей жизни назвал: «Химическое строение биосферы Земли и ее окружения». Владимир Иванович не определил, что он понимает под «окружением» биосферы. В значительной мере это было сделано Николаем Брониславовичем Вассоевичем. Развивая идеи В. И. Вернадского, Н. Б. Вассоевич предлагал выделять «мегабиосферу» (от греч. «мега» — великий) — многослойную оболочку Земли, сформированную в результате деятельности живого вещества. Верхней ее границей является предел распространения биогенной атмосферы, нижней — земные оболочки, не затронутые влиянием жизни. Мегабиосфера включает в себя: а) анобиосферу — верхнюю часть атмосферы Земли выше границы распространения форм жизни в состоянии анабиоза; б) парабиосферу; в) биосферу; г) метабиосферу, соответствующую «области былых биосфер» Вернадского. «Область былых биосфер» В. И. Вернадский понимал как оболочку Земли, когда-либо подвергавшуюся воздействию жизни. Владимир Иванович писал, что земная кора «захватывает в пределах нескольких десятков километров ряд геологических оболочек, которые когда-то были на поверхности Земли биосферами. Это биосфера, стратисфера, метаморфическая (верхняя и нижняя) оболочка, гранитная оболочка. Происхождение их всех из биосферы становится нам ясным только теперь. Это — былые биосферы»[15]. Таким образом, благодаря постоянному притоку солнечной энергии, интенсивной работе приемника этой энергии — живого вещества — и неполной замкнутости биогеохимического круговорота биосфера непрерывно создает вокруг себя концентрические планетарные оболочки: снаружи парабиосферу и анобиосферу, внутри метабиосферу. Характерной особенностью биосферы как динамической системы является ее неравновесность, являющаяся следствием работы живого вещества и притока солнечной энергии. Еще в 1935 г. младший современник Вернадского, известный биолог Эрвин Бауэр (1890—1942), проживший последние 17 лет своей жизни в Советском Союзе, писал: «Все и только живые системы никогда не бывают в равновесии и исполняют за счет своей свободной энергии постоянно работу против равновесия, требуемого законами физики и химии при существующих внешних условиях… Мы обозначим этот принцип как принцип устойчивого неравновесия живых систем». Этот принцип показывает, что живые организмы представляют собой открытые неравновесные системы, которые отличаются от неживых тем, что эволюционируют в сторону понижения энтропии. Сейчас его называют принципом Бауэра. Благодаря деятельности живого вещества, принцип устойчивого неравновесия действует и в биосфере в целом. Не менее важной отличительной особенностью биосферы является ее обводненность. «Вода без жизни в биосфере неизвестна; ничтожные по весу такие ее нахождения являются редкими — временными — минералами; таковы воды, богатые свободной серной или соляной кислотой вулканов, может быть, некоторые рассолы — и только»[16]. Одновременно вода является средой, по существу, и для всех химических процессов, происходящих в биосфере. Именно поэтому химически чистая вода в биосфере представляет собой столь же большую редкость, как и вода, лишенная жизни. Вернадскому очень правилось определение жизни как «одушевленной воды», данное французским биологом Р. Дюбуа, и он неоднократно приводил его в своих произведениях. Поэтические строки о воде принадлежат и другому великому французу — Антуану де Сент-Экзюпери: «Вода, у тебя нет ни вкуса, ни цвета, ни запаха, тобой наслаждаются, не ведая, что ты такое. Нельзя сказать, что ты необходима для жизни: ты — сама жизнь…» Наконец, еще одна характерная особенность биосферы — это ее неразрывная связь с космосом (в наибольшей степени — с Солнцем). Впервые ее заметили еще в прошлом веке. Так, в 1852 г. швейцарский астроном Рудольф Вольф (1816—1896) рассчитал зависимость магнетизма Земли от цикличности пятнообразования на Солнце. Однако датой рождения гелиобиологии (науки о влиянии Солнца на биологические процессы) обычно считают 1915 г., когда в Московском археологическом институте был прочитан доклад «Периодическое влияние Солнца на биосферу Земли». В этом докладе восемнадцатилетний Александр Леонидович Чижевский (1897—1964), впоследствии известный ученый, сформулировал идею, развитию которой посвятил всю жизнь. В наши дни она может показаться даже тривиальной — что же, собственно говоря, удивительного в том, что жизнь на Земле определяется Солнцем? Сейчас выявлен огромный диапазон явлений, подчиненных «ритму Солнца» — от вспышек массового размножения саранчи до средней успеваемости школьников. Чижевский же был первопроходцем, и ему не удалось дожить до признания своих идей… Подведем некоторые итоги. Биосфера вырисовывается как пронизанная и регулируемая жизнью, ею сформированная наружная оболочка Земли, развитие которой определяется главным образом постоянным притоком солнечной энергии[17]. «Основным признаком биосферы является участие во всех ее процессах живого вещества» (Вернадский). Биосфера Земли характеризуется присутствием жидкой воды и широким развитием низкотемпературных реакций, регулируемых действием ферментов. Биосфера продуцирует наружу газовую оболочку, внутрь планеты — «область былых биосфер», или метабиосферу. Говорят, что все в жизни познается в сравнении. Нашу, земную биосферу нам сравнивать не с чем — других биосфер во Вселенной пока не обнаружено, и шансы на их существование все уменьшаются. Впрочем, недавно ученые из Главной астрономической обсерватории АН УССР Л. О. Колоколова и А. Ф. Стеклов выдвинули гипотезу о наличии на спутниках Юпитера биосферы иного типа, чем земная, — биосферы, развивающейся под слоем льда. Такой гипотетический тип биосферы украинские ученые предложили называть «эндобиосферой» (от греч. «эндон» — внутри). Отдаленным ее земным аналогом может быть экосистема антарктического моря Росса, о которой мы уже упоминали. Установлено, что спутники Юпитера — Европа, Ганимед и Каллисто — покрыты ледяным панцирем толщиной 50—250 км, под которым, видимо, располагается водный океан глубиной 100—1000 км. В этом-то подледном океане и предполагается развитие жизни, представленной микроскопическими существами, приспособленными к существованию при огромных давлениях. Недавние исследования, произведенные космическими аппаратами, показали, что в океане юпитерианской Европы существуют полыньи размером в десятки квадратных километров. В этом случае энергии, получаемой Европой от Солнца, как будто должно хватать на фотосинтез, если, конечно, есть кому его осуществлять. Подледные океаны подобного типа существуют также на Марсе, Титане и, возможно, Тритоне. Так что в принципе существование эндобиосфер не исключено и на этих планетах. Но это — гипотеза, а пока наша биосфера — экзобиосфера! — остается в Солнечной системе уникальной. И мы должны ее сохранить. Наш очерк о биосфере будет неполным, если мы хотя бы в краткой форме не остановимся на современном этапе ее развития, который В. И. Вернадский называл ноосферой (от греч. «ноо» — разум). Конечно, как и в учении о биосфере, в ноосферной концепции В. И. Вернадский тоже имел предшественников. Одним из них был самобытный русский философ Николай Федорович Федоров (1828—1903), первый (да, по существу, и единственный) наставник К. Э. Циолковского. Тезис Н. Ф. Федорова «Природа в нас начинает не только сознавать себя, но и управлять собою»[18] по идейной сути близок к хорошо известному пониманию Ф. Энгельсом человека как позвоночного, «в котором природа приходит к осознанию самой себя»[19]. Н. Ф. Федоров ставит вопрос об управлении («регуляции») природы человеком: «И в чем же должно состоять наше, существ разумных и чувствующих, общее дело: в эксплуатации ли и утилизации природы, т. е. в ее истощении… или же в регуляции природы… Повиноваться природе для разумного существа значит управлять ею, неразумною силою, ибо природа в разумных существах приобрела себе главу и правителя»[20]. Первое употребление термина «ноосфера» (как и «биосфера») принадлежит не Вернадскому — он вообще не любил придумывать новые слова. Его ввели в 1927 г. два младших современника В. И. Вернадского, слушавшие его лекции по геохимии в Сорбонне, — философ, математик, палеонтолог и антрополог Эдуард Леруа (1870—1954) и его друг, палеонтолог и антрополог Пьер Тейяр де Шарден (1881—1955). Под ноосферой французские ученые понимали ту стадию эволюции природы, когда появился человеческий разум. Развернутое обоснование эта трактовка получила в книге П. Тейяра де Шардена «Феномен человека», впервые опубликованной в 1959 г., через 14 лет после смерти В. И. Вернадского. В ней Тейяр определял ноосферу как «новый покров», «мыслящий пласт, который, зародившись в конце третичного периода, разворачивается над миром растений и животных — вне биосферы и над ней» (курсив мой. — А. Л.). Иное содержание вкладывал в этот термин В. И. Вернадский. Его статья «Несколько слов о ноосфере» появилась в суровое время — в 1944 г. — и оказалась последней прижизненной публикацией ученого. По существу, она представляет собой научное завещание Вернадского. В этой статье были сформулированы представления, к которым Владимир Иванович пришел исподволь, всей логикой своего творчества: «Человечество, взятое в целом, становится мощной геологической силой. И перед ним, перед его мыслью и трудом, становится вопрос о перестройке биосферы в интересах свободно мыслящего человечества как единого целого. Это новое состояние биосферы, к которому мы, не замечая этого, приближаемся, и есть ноосфера»[21]. Ноосфера, по Вернадскому, это биосфера, разумно управляемая человеком. В поэтической форме эту идею развил Николай Заболоцкий:
Два мира есть у человека:
Один, который нас творил,
Другой, который мы от века
Творим по мере наших сил.
Так связан, съединен от века
Союзом кровного родства
Разумный гений человека
С творящей силой естества…
Глава вторая. Живое вещество
Скажи, прошу тебя, на что мы созданы?На что сотворены медведь, сова, лягушка?На что сотворены и Ванька и Петрушка?Д. И. Фонвизин. 1763
 В книге «Химическое строение биосферы Земли и ее окружения» Вернадский поставил вопрос о типах вещества, слагающих биосферу[26]:
«Вещество ее (биосферы. — А. Л.) состоит из семи глубоко разнородных природных частей, геологически не случайных.
Во-первых, из совокупности живых организмов, живого вещества, рассеянного в мириадах особей…
Во-вторых, мы имеем дело с веществом, создаваемым и перерабатываемым жизнью… с биогенным веществом, источником чрезвычайно мощной потенциальной энергии (каменный уголь, битумы, известняки, нефть и т. д.). Живые организмы в нем после его образования геологически малодеятельны.
В-третьих, мы имеем вещество, образуемое процессами, в которых живое вещество не участвует: косное вещество, твердое, жидкое и газообразное…
Четвертая часть — это биокосное вещество, которое создается одновременно живыми организмами и косными процессами, представляя динамические равновесные системы тех и других. Таковы вся океаническая и почти вся другая вода биосферы, нефть, почва, кора выветривания и т. д. Организмы в них играют ведущую роль…
В-пятых, вещество, находящееся в радиоактивном распаде в форме немногих относительно прочных радиоактивных элементов… Мы здесь имеем дело с химическими элементами сложного изотопного состава, проникающими все вещество биосферы и идущими вглубь на глубину, нам неизвестную…
С другой стороны, все вещество биосферы и, по-видимому, только биосферы, проникнуто шестой формой вещества — рассеянными атомами, которые непрерывно создаются из всякого рода земного вещества под влиянием космических излучений…
Наконец, седьмым типом земного вещества является вещество космического происхождения…» (курсив везде Вернадского. — А. Л.).
Во второй главе книги мы рассмотрим типы вещества, распространенные в биосфере, обращая особое внимание на живое вещество.
Н. Б. Вассоевич, рассматривая классификацию типов вещества по Вернадскому, отметил, что логически она не является безупречной, так как выделенные категории вещества частично перекрывают друг друга (например, вещество космического происхождения одновременно является и косным). С другой стороны, печальный опыт человечества, приобретенный уже после смерти В. И. Вернадского, научил нас, что радиоактивные изотопы (например, стронций‑90) накапливаются живыми организмами и, следовательно, могут входить в состав как живого, так и косного вещества. Что же касается «биокосного вещества», то его нельзя рассматривать в качестве особого типа вещества, поскольку оно, по определению Вернадского, состоит из двух веществ — живого и косного. Следовательно, по своему характеру это не единое вещество, а динамическая система, что подчеркивает и сам Вернадский (этот вопрос подробнее мы рассмотрим в главе пятой).
Свою классификацию видов вещества в биосфере В. И. Вернадский строил по нескольким параметрам:
а) по характеру самого вещества (живое или косное);
б) по характеру исходного вещества (выделяются биогенное вещество и косное вещество, в образовании которого жизнь не участвует);
в) по признаку радиоактивности (выделяется вещество, находящееся в радиоактивном распаде);
г) по степени дисперсности молекулярной структуры (выделяется вещество, представленное рассеянными атомами);
д) по признаку земного или внеземного происхождения (выделяется вещество космического происхождения).
Попытаемся в матричной форме представить генетическую классификацию типов вещества биосферы. В качестве классификационных выберем три основных параметра: а) характер вещества: живое или неживое; б) градации по исходному материалу: образовавшееся из живого вещества — биогенное и образовавшееся из неживого вещества — абиогенное; в) признак земного или внеземного происхождения (табл. 1). Радиоактивность и степень дисперсности молекулярной структуры в качестве параметров классификации опускаем.
Таблица 1
Типы вещества биосферы Земли
В книге «Химическое строение биосферы Земли и ее окружения» Вернадский поставил вопрос о типах вещества, слагающих биосферу[26]:
«Вещество ее (биосферы. — А. Л.) состоит из семи глубоко разнородных природных частей, геологически не случайных.
Во-первых, из совокупности живых организмов, живого вещества, рассеянного в мириадах особей…
Во-вторых, мы имеем дело с веществом, создаваемым и перерабатываемым жизнью… с биогенным веществом, источником чрезвычайно мощной потенциальной энергии (каменный уголь, битумы, известняки, нефть и т. д.). Живые организмы в нем после его образования геологически малодеятельны.
В-третьих, мы имеем вещество, образуемое процессами, в которых живое вещество не участвует: косное вещество, твердое, жидкое и газообразное…
Четвертая часть — это биокосное вещество, которое создается одновременно живыми организмами и косными процессами, представляя динамические равновесные системы тех и других. Таковы вся океаническая и почти вся другая вода биосферы, нефть, почва, кора выветривания и т. д. Организмы в них играют ведущую роль…
В-пятых, вещество, находящееся в радиоактивном распаде в форме немногих относительно прочных радиоактивных элементов… Мы здесь имеем дело с химическими элементами сложного изотопного состава, проникающими все вещество биосферы и идущими вглубь на глубину, нам неизвестную…
С другой стороны, все вещество биосферы и, по-видимому, только биосферы, проникнуто шестой формой вещества — рассеянными атомами, которые непрерывно создаются из всякого рода земного вещества под влиянием космических излучений…
Наконец, седьмым типом земного вещества является вещество космического происхождения…» (курсив везде Вернадского. — А. Л.).
Во второй главе книги мы рассмотрим типы вещества, распространенные в биосфере, обращая особое внимание на живое вещество.
Н. Б. Вассоевич, рассматривая классификацию типов вещества по Вернадскому, отметил, что логически она не является безупречной, так как выделенные категории вещества частично перекрывают друг друга (например, вещество космического происхождения одновременно является и косным). С другой стороны, печальный опыт человечества, приобретенный уже после смерти В. И. Вернадского, научил нас, что радиоактивные изотопы (например, стронций‑90) накапливаются живыми организмами и, следовательно, могут входить в состав как живого, так и косного вещества. Что же касается «биокосного вещества», то его нельзя рассматривать в качестве особого типа вещества, поскольку оно, по определению Вернадского, состоит из двух веществ — живого и косного. Следовательно, по своему характеру это не единое вещество, а динамическая система, что подчеркивает и сам Вернадский (этот вопрос подробнее мы рассмотрим в главе пятой).
Свою классификацию видов вещества в биосфере В. И. Вернадский строил по нескольким параметрам:
а) по характеру самого вещества (живое или косное);
б) по характеру исходного вещества (выделяются биогенное вещество и косное вещество, в образовании которого жизнь не участвует);
в) по признаку радиоактивности (выделяется вещество, находящееся в радиоактивном распаде);
г) по степени дисперсности молекулярной структуры (выделяется вещество, представленное рассеянными атомами);
д) по признаку земного или внеземного происхождения (выделяется вещество космического происхождения).
Попытаемся в матричной форме представить генетическую классификацию типов вещества биосферы. В качестве классификационных выберем три основных параметра: а) характер вещества: живое или неживое; б) градации по исходному материалу: образовавшееся из живого вещества — биогенное и образовавшееся из неживого вещества — абиогенное; в) признак земного или внеземного происхождения (табл. 1). Радиоактивность и степень дисперсности молекулярной структуры в качестве параметров классификации опускаем.
Таблица 1
Типы вещества биосферы Земли
| Характер вещества | Градации по исходному веществу | Типы вещества земного происхождения | Типы вещества внеземного происхождения |
|---|---|---|---|
| Живое | Биогенное | ЖИВОЕ ВЕЩЕСТВО. Синонимы: биос, биота | Неизвестно |
| Абиогенное | Неизвестно | Неизвестно | |
| Неживое | Биогенное | БИОГЕННОЕ ВЕЩЕСТВО: а) необиогенное вещество; б) палеобиогенное вещество Синоним: органогенное вещество | Неизвестно |
| Абиогенное | АБИОГЕННОЕ ВЕЩЕСТВО ЗЕМНОГО ПРОИСХОЖДЕНИЯ. Синоним: косное вещество | АБИОГЕННОЕ ВЕЩЕСТВО ВНЕЗЕМНОГО ПРОИСХОЖДЕНИЯ. Синоним: вещество космического происхождения |
…как только они загнивать начинают, от влаги
Тотчас в них черви являются, ибо материи тельца
Вследствие новых условий, меняя свой старый порядок,
Так сочетаются, что из них твари живые родятся…
И внял я неба содроганье,
И горний ангелов полет,
И гад морских подводный ход,
И дольной лозы прозябанье…
Слышно, как лезут с роптаньем и писком.
Слышно, как стену их лапки скребут.
Слышно, как камень их зубы грызут.
Вдруг ворвались неизбежные звери;
Сыплются градом сквозь окна, сквозь двери,
Спереди, сзади, с боков, с высоты…
Пусть вымерли все наши предки —
Бессмертные живые клетки
Наследье бережно хранят —
| Макросистема живых организмов[39] | Роль живых организмов в биосфере | ||||||
|---|---|---|---|---|---|---|---|
| Надцарства | Царства | Подцарства | Автотрофы | Гетеротрофы | Миксотрофы | ||
| Фототрофы | Хемотрофы | Биотрофы | Сапротрофы и некротрофы | ||||
| Доядерные (прокариоты) | Дробянки | Бактерии (эубактерии) | + | + | + | + | + |
| Архебактерии | + | + | − | + | + | ||
| Цианеи (цианобактерии, или синезеленые водоросли) | + | + | − | − | + | ||
| Ядерные (эукариоты) | Растения | Низшие растения (водоросли) | + | − | − | − | + |
| Высшие растения | + | − | Очень редко | ? | + | ||
| Грибы | Миксомицеты (слизевики) | − | − | + | + | − | |
| Грибы (высшие грибы) | − | − | + | + | − | ||
| Животные | Простейшие | − | − | + | Очень редко | + | |
| Многоклеточные животные | − | − | + | + | + | ||
А грибы были грибами,
Они ни на кого не похожи —
Толстое тело коровы,
Поставленное на четыре окончанья,
Увенчанное храмовидной головою
И двумя рогами (словно луна в первой четверти),
Тоже будет непонятно,
Тоже будет непостижимо,
Если забудем о его значеньи
На карте живущих всего мира.
Н. Заболоцкий
Глава третья. Сгущения и пленки жизни
Все живое представляет неразрывное целое, закономерно связанное не только между собою, но и с окружающей косной средой биосферы.В. И. Вернадский. 1926
 Из жизненного опыта нам хорошо известно, что жизнь размещена в биосфере очень неравномерно, и площади, густо населенные живыми организмами, чередуются с пустынными пространствами. Такая неравномерность распределения живого вещества наблюдается в биосфере повсеместно — на суше и на море, на земле и под землей.
В. И. Вернадский выделял две формы концентраций жизни: жизненные пленки, прослеживаемые на огромных площадях (например, планктонная пленка жизни, покрывающая всю верхнюю часть водной толщи океана), и сгущения жизни, имеющие более локальное распространение (например, сгущения стоячих водоемов). Мощность концентраций жизни обычно измеряется единицами или десятками, значительно реже — одной-двумя сотнями метров, т. е. по отношению к биосфере в целом — ничтожными величинами. Остальная часть биосферы представляет собой зону разрежения живого вещества.
Пленки и сгущения жизни являются областями наибольшей биогенной миграции атомов и трансформации энергии в биосфере. Развивая идеи Вернадского, А. И. Перельман отметил, что вся биосфера по вертикали отчетливо разделяется на две зоны: верхнюю, в которой происходит фотосинтез, и нижнюю, где фотосинтетические реакции невозможны. Он предложил верхнюю зону называть «фитосферой», а нижнюю — «редусферой». Н. Б. Вассоевич, критикуя эти названия, предлагал соответственно для зон название «фотобиосфера» и «мелабиосфера» (от древнегреческого корня «мела(н)», темный). Однако термин «мелабиосфера» также не является вполне удачным, поскольку он лишь одной буквой отличается от других терминов Н. Б. Вассоевича — «метабиосфера» и «мегабиосфера». Представляется поэтому, что нижнюю зону биосферы лучше называть не мелабиосферой, как предлагает Н. Б. Вассоевич, а меланобиосферой.
Граница между фотобиосферой и меланобиосферой на суше почти совпадает с дневной поверхностью: свет проникает в глубь почвы лишь на несколько миллиметров. В водной среде положение границы определяется прозрачностью воды. Толщина зоны фотосинтеза изменяется от нескольких сантиметров в быстротекущих реках, несущих значительное количество ила, до первой сотни метров (максимально до 180 м) на удаленных от суши участках океана. В соответствии с этим мощность фотобиосферы колеблется от нескольких миллиметров до первой сотни метров (на суше — вверх от дневной поверхности: вековые леса, в океане — вниз от поверхности моря: зона фотосинтеза). Мощность меланобиосферы на 1—2 порядка больше: в океанах — это вся водная толща ниже зоны фотобиосферы и заселенный слой донных осадков, на континентах — слой биосферы от дневной поверхности до нижней границы распространения активной бактериальной жизни.
Коренное отличие фотобиосферы от меланобиосферы состоит в структуре их живого вещества: в первом случае оно представлено фотоавтотрофами и гетеротрофами, во втором — фотоавтотрофы отсутствуют (однако в некоторых случаях их заменяют хемоавтотрофы). Впрочем, и среди гетеротрофов в меланобиосфере живут лишь виды, приспособившиеся к отсутствию света. Что касается человека, то он, расселяясь в пещерах, начал осваивать меланобиосферу много тысячелетий назад. Затем, переселившись в более уютную фотобиосферу, он начал углубляться в меланобиосферу своими рудниками. А сейчас многие из нас, пользуясь городским метрополитеном, ежедневно совершают «суточные миграции» в меланобиосферу.
Фотобиосферу и меланобиосферу можно разбить по вертикали и на более дробные зоны. Так, советский исследователь Юрий Петрович Бяллович ввел понятие биогеоценотического горизонта, или биогеогоризонта, определив его следующим образом: «Биогеоценотический горизонт есть вертикально обособленная и по вертикали далее нерасчленимая структурная часть биогеоценоза. Сверху донизу биогеоценотический горизонт однороден по составу биогеоценотических компонентов, по взаимосвязям их, по происходящим в нем превращениям вещества и энергии, и в этих же отношениях он отличается от соседних биогеоценотических горизонтов, служащих ему кровлей и постелью». Первопричиной деления биосферы на биогеогоризонты, по Бялловичу, является радиальное направление гравитации, солнечной радиации и земного излучения. В экосистемах всех рангов можно проследить не только эти элементарные, далее нерасчленимые, биогеогоризонты, но и слои более высоких рангов, которые целесообразно называть экогоризонтами[41]. Экогоризонтами высшего — глобального — ранга и являются фотобиосфера и меланобиосфера. Выделяемые В. И. Вернадским пленки жизни можно рассматривать как частный случай экогоризонтов.
Итак, по горизонтали биосфера делится на экосистемы, по вертикали — на экогоризонты. Действие закона всемирного тяготения приводит к тому, что взаимозависимость между двумя соседними экогоризонтами обычно больше, чем между соседними экосистемами.
Все экосистемы биосферы Земли по ландшафтному принципу можно разделить на три основные группы: а) морские экосистемы; б) экосистемы суши; в) экосистемы континентальных водоемов. Только морские экосистемы объединены в единую грандиозную экосистему — Мировой океан. Другие типы экосистем имеют дисперсное распространение: экосистемы наземных водоемов окружены сушей, а суша, в свою очередь, океаном. В современную эпоху они занимают следующие площади: Мировой океан — 361,2 млн. км², суша — 145,7 млн. км², континентальные водоемы — лишь 3,2 млн. км². Рассмотрим, как распределено живое вещество в этих основных типах экосистем биосферы и какие следы оставляют они в геологических отложениях.
«Биогенные соли в глубине и наличие света у поверхности» — так в афористической форме выразил советский океанолог Ю. Ю. Марти основную проблему морских экосистем. Мировой океан включает в себя водную толщу (океанологи ее называют пелагиалью) и дно (бенталь). Пелагиаль в пределах фотобиосферы в океанологии называют эвфотической зоной; нижняя часть пелагиали именуется афотической зоной. По существу, это три самостоятельных экогоризонта океана (сверху вниз: эвфотическая зона, афотическая зона и бенталь), каждый из которых характеризуется своим специфическим живым веществом и условиями среды. В некоторых полузамкнутых бассейнах с затрудненной циркуляцией вод (типа Черного моря) обнаруживается другой своеобразный слабо заселенный экогоризонт — зона сероводородного заражения, где прозябают только несколько видов анаэробных бактерий.
В. И. Вернадский выделил в океане две жизненные пленки (планктонную и донную). Обе они приурочены к границам раздела фаз: планктонная — газообразной и жидкой, донная — жидкой и твердой (рис. 2).
Из жизненного опыта нам хорошо известно, что жизнь размещена в биосфере очень неравномерно, и площади, густо населенные живыми организмами, чередуются с пустынными пространствами. Такая неравномерность распределения живого вещества наблюдается в биосфере повсеместно — на суше и на море, на земле и под землей.
В. И. Вернадский выделял две формы концентраций жизни: жизненные пленки, прослеживаемые на огромных площадях (например, планктонная пленка жизни, покрывающая всю верхнюю часть водной толщи океана), и сгущения жизни, имеющие более локальное распространение (например, сгущения стоячих водоемов). Мощность концентраций жизни обычно измеряется единицами или десятками, значительно реже — одной-двумя сотнями метров, т. е. по отношению к биосфере в целом — ничтожными величинами. Остальная часть биосферы представляет собой зону разрежения живого вещества.
Пленки и сгущения жизни являются областями наибольшей биогенной миграции атомов и трансформации энергии в биосфере. Развивая идеи Вернадского, А. И. Перельман отметил, что вся биосфера по вертикали отчетливо разделяется на две зоны: верхнюю, в которой происходит фотосинтез, и нижнюю, где фотосинтетические реакции невозможны. Он предложил верхнюю зону называть «фитосферой», а нижнюю — «редусферой». Н. Б. Вассоевич, критикуя эти названия, предлагал соответственно для зон название «фотобиосфера» и «мелабиосфера» (от древнегреческого корня «мела(н)», темный). Однако термин «мелабиосфера» также не является вполне удачным, поскольку он лишь одной буквой отличается от других терминов Н. Б. Вассоевича — «метабиосфера» и «мегабиосфера». Представляется поэтому, что нижнюю зону биосферы лучше называть не мелабиосферой, как предлагает Н. Б. Вассоевич, а меланобиосферой.
Граница между фотобиосферой и меланобиосферой на суше почти совпадает с дневной поверхностью: свет проникает в глубь почвы лишь на несколько миллиметров. В водной среде положение границы определяется прозрачностью воды. Толщина зоны фотосинтеза изменяется от нескольких сантиметров в быстротекущих реках, несущих значительное количество ила, до первой сотни метров (максимально до 180 м) на удаленных от суши участках океана. В соответствии с этим мощность фотобиосферы колеблется от нескольких миллиметров до первой сотни метров (на суше — вверх от дневной поверхности: вековые леса, в океане — вниз от поверхности моря: зона фотосинтеза). Мощность меланобиосферы на 1—2 порядка больше: в океанах — это вся водная толща ниже зоны фотобиосферы и заселенный слой донных осадков, на континентах — слой биосферы от дневной поверхности до нижней границы распространения активной бактериальной жизни.
Коренное отличие фотобиосферы от меланобиосферы состоит в структуре их живого вещества: в первом случае оно представлено фотоавтотрофами и гетеротрофами, во втором — фотоавтотрофы отсутствуют (однако в некоторых случаях их заменяют хемоавтотрофы). Впрочем, и среди гетеротрофов в меланобиосфере живут лишь виды, приспособившиеся к отсутствию света. Что касается человека, то он, расселяясь в пещерах, начал осваивать меланобиосферу много тысячелетий назад. Затем, переселившись в более уютную фотобиосферу, он начал углубляться в меланобиосферу своими рудниками. А сейчас многие из нас, пользуясь городским метрополитеном, ежедневно совершают «суточные миграции» в меланобиосферу.
Фотобиосферу и меланобиосферу можно разбить по вертикали и на более дробные зоны. Так, советский исследователь Юрий Петрович Бяллович ввел понятие биогеоценотического горизонта, или биогеогоризонта, определив его следующим образом: «Биогеоценотический горизонт есть вертикально обособленная и по вертикали далее нерасчленимая структурная часть биогеоценоза. Сверху донизу биогеоценотический горизонт однороден по составу биогеоценотических компонентов, по взаимосвязям их, по происходящим в нем превращениям вещества и энергии, и в этих же отношениях он отличается от соседних биогеоценотических горизонтов, служащих ему кровлей и постелью». Первопричиной деления биосферы на биогеогоризонты, по Бялловичу, является радиальное направление гравитации, солнечной радиации и земного излучения. В экосистемах всех рангов можно проследить не только эти элементарные, далее нерасчленимые, биогеогоризонты, но и слои более высоких рангов, которые целесообразно называть экогоризонтами[41]. Экогоризонтами высшего — глобального — ранга и являются фотобиосфера и меланобиосфера. Выделяемые В. И. Вернадским пленки жизни можно рассматривать как частный случай экогоризонтов.
Итак, по горизонтали биосфера делится на экосистемы, по вертикали — на экогоризонты. Действие закона всемирного тяготения приводит к тому, что взаимозависимость между двумя соседними экогоризонтами обычно больше, чем между соседними экосистемами.
Все экосистемы биосферы Земли по ландшафтному принципу можно разделить на три основные группы: а) морские экосистемы; б) экосистемы суши; в) экосистемы континентальных водоемов. Только морские экосистемы объединены в единую грандиозную экосистему — Мировой океан. Другие типы экосистем имеют дисперсное распространение: экосистемы наземных водоемов окружены сушей, а суша, в свою очередь, океаном. В современную эпоху они занимают следующие площади: Мировой океан — 361,2 млн. км², суша — 145,7 млн. км², континентальные водоемы — лишь 3,2 млн. км². Рассмотрим, как распределено живое вещество в этих основных типах экосистем биосферы и какие следы оставляют они в геологических отложениях.
«Биогенные соли в глубине и наличие света у поверхности» — так в афористической форме выразил советский океанолог Ю. Ю. Марти основную проблему морских экосистем. Мировой океан включает в себя водную толщу (океанологи ее называют пелагиалью) и дно (бенталь). Пелагиаль в пределах фотобиосферы в океанологии называют эвфотической зоной; нижняя часть пелагиали именуется афотической зоной. По существу, это три самостоятельных экогоризонта океана (сверху вниз: эвфотическая зона, афотическая зона и бенталь), каждый из которых характеризуется своим специфическим живым веществом и условиями среды. В некоторых полузамкнутых бассейнах с затрудненной циркуляцией вод (типа Черного моря) обнаруживается другой своеобразный слабо заселенный экогоризонт — зона сероводородного заражения, где прозябают только несколько видов анаэробных бактерий.
В. И. Вернадский выделил в океане две жизненные пленки (планктонную и донную). Обе они приурочены к границам раздела фаз: планктонная — газообразной и жидкой, донная — жидкой и твердой (рис. 2).
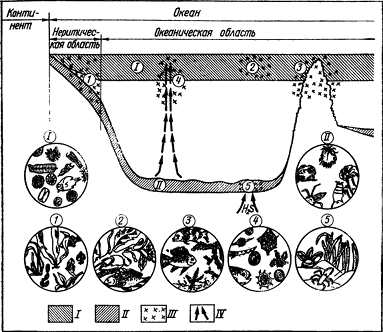 Рис. 2. Экогоризонты, концентрации и разрежения жизни Мирового океана: I — планктонная пленка жизни; II — донная пленка жизни; III — сгущения жизни; 1 — прибрежное; 2 — саргассовое; 3 — рифовое; 4 — апвеллинговое; 5 — абиссальное рифтовое; IV — подъем глубинных вод; А — разрежение жизни
Рис. 2. Экогоризонты, концентрации и разрежения жизни Мирового океана: I — планктонная пленка жизни; II — донная пленка жизни; III — сгущения жизни; 1 — прибрежное; 2 — саргассовое; 3 — рифовое; 4 — апвеллинговое; 5 — абиссальное рифтовое; IV — подъем глубинных вод; А — разрежение жизни
Планктонная пленка жизни В. И. Вернадского в основном соответствует эвфотической зоне океана. По составу живого вещества она резко отличается от наземных экосистем: доминируют здесь организмы, взвешенные в воде и неспособные противостоять течениям (сообщество этих организмов и есть планктон — от греческого корня «планктос» — парящий, блуждающий). Совокупность фотоавтотрофных планктонных организмов называют фитопланктоном, гетеротрофных — зоопланктоном. Специфичность планктона как особого сообщества водных организмов впервые была показана знаменитым немецким биологом Иоганном Петером Мюллером (1801—1858). До последнего времени считалось, что по первичной продукции лидером среди организмов планктона являются одноклеточные водоросли (главным образом диатомовые, измеряемые десятками и сотнями микронов), однако недавние исследования показали, что от 30 до 80% первичной продукции океана дают значительно более мелкие (0,4—1 мкм) фотосинтезирующие организмы, которые раньше ускользали от внимания исследователей из-за ничтожности своих размеров и несовершенства лабораторного оборудования. Эти организмы назвали пикопланктоном (от исп. «pico» — малая величина). Первыми идентифицированными представителями пикопланктона оказались цианобактерии. Результаты совместного советско-французского исследования, проведенного в 1983 г., позволили выявить в составе пикопланктона и эукариоты, но определить их пока не удалось. Оказалось, что максимум распределения пикопланктона приурочен к акваториям тропических и субтропических морей. А поскольку в его составе преобладают цианобактерии, не приходится удивляться, что пикопланктон весьма нетребователен к содержанию азота в воде — ведь цианобактерии могут поглощать его и из воздуха! И еще одна специфическая особенность: вклад пикопланктона в первичную продукцию возрастает с глубиной, поскольку организмы пикопланктона, по-видимому, способны осуществлять фотосинтез при очень низкой интенсивности солнечного света[42]. Состав зоопланктона очень разнообразен. Самыми распространенными и важными в экологическом отношении в составе зоопланктона являются копеподы (веслоногие рачки), обычный размер которых — всего 2—3 мм, а максимальная величина — до 10 мм. По биомассе им уступают эвфаузииды — несколько более крупные рачки размером до 5 см, очень похожие на креветок. Обычно они образуют огромные скопления в океане, которые рыбаки называют крилем. Криль — «дежурное» блюдо китов (индивидуальная суточная норма — 1,5 т). К зоопланктону относятся также медузы, сальпы, некоторые моллюски, простейшие, а также многие другие организмы (зачастую только в виде икры, личинок или молоди). Плотность населения в планктонной пленке такова, что девять десятых живых организмов, будь то растения или животные, поедаются раньше, чем наступает их естественная смерть: рачки-копеподы питаются диатомовыми водорослями, копепод пожирают более крупные рачки, и т. д. Количество живых организмов в планктонной пленке быстро убывает с глубиной. По данным одной из экспедиций 30‑х годов, содержание живых организмов в 1 л морской воды оказалось следующим: в поверхностном слое — 10 147 особей, на глубине 50 м — 9443, 100 м — 2749. Мощность планктонной пленки как своеобразной концентрации жизни Вернадский оценивал в 50—60 м. Эвфотическая зона — это огород океана. Именно здесь синтезируется большая часть автотрофного живого вещества океана (об исключениях поговорим позже). Оно кормит чуть ли не весь океан (а его накормить не так-то просто!), и накопленная им энергия является энергетическим источником большинства геохимических процессов, происходящих в океане. Огромна породообразующая роль планктона — он поставляет сырье для будущих горных пород. Планктонная пленка жизни продуцирует огромное количество необиогенного вещества, которое, однако, не может в ней накапливаться, а опускается под действием силы тяжести сквозь водную толщу вниз, пока не достигнет дна. Под планктонной пленкой располагается мощная водная афотическая зона «разрежения жизни», по Вернадскому. Афотическая зона водной толщи превышает эвфотическую по мощности в 40 раз (средняя глубина океана 3800 м). Плотность живого вещества здесь на несколько порядков ниже, чем в эвфотической зоне. Это — область вечного мрака, и собственного автотрофного живого вещества в афотической зоне нет. Гетеротрофные организмы питаются здесь детритом, поступающим из планктонной пленки жизни, или являются хищниками. Детрит представлен главным образом пеллетами, которые многократно реутилизируются живыми организмами в процессе погружения. Пищевая ценность детрита при этом неуклонно снижается. Вся водная толща океана представляет собой, по существу, транзитную зону. В твердом виде биогенное вещество здесь не накапливается, однако содержание элементов минерального питания в растворенном виде в этой зоне выше, чем в эвфотической. Водная толща Мирового океана подстилается донной пленкой жизни. Ее обитателей Э. Геккель назвал бентосом (от греч. «бентос» — глубина). К бентосу относится 157 тыс. из 160 тыс. видов морских животных. В его состав входят бактерии, простейшие и многоклеточные животные разных типов. К бентосу относятся и прикрепленные ко дну многоклеточные водоросли, однако они хуже, чем планктонные, переносят недостаток света и распространены до глубин менее 50 м. Размеры бентосных организмов варьируют в очень широких пределах и могут различаться на несколько порядков. Во времена Вернадского донная пленка была изучена только в пределах шельфа, и Владимир Иванович допустил невольную ошибку — данные по шельфу он экстраполировал на донную пленку в целом, считая ее вместилищем жизни более богатым, чем планктонная пленка. Однако сейчас установлена крайняя неравномерность заселения бентосной пленки живыми организмами. Удаленные от континентов участки акваторий, по площади составляющие ¾ территории Мирового океана, обладают лишь 1% суммарной биомассы бентоса. Интересный эксперимент провели недавно ученые Колумбийского университета (США) в восточно-экваториальной части Тихого океана. Здесь на глубине 4873 м была установлена фотокамера, автоматически проводившая фотосъемку океанского дна через каждые 4 ч в течение 202 дней. За это время в поле зрения камеры проползло или прошагало только 35 животных. Иначе говоря, животное проходило здесь только раз в шесть дней! Между распределением биомасс планктонной и донной пленок жизни существует тесная корреляционная зависимость, и акваториям с высокой биомассой планктона, как правило, соответствуют участки с повышенным содержанием живого вещества на бентали. В честь известного советского океанолога, открывшего эту зависимость, ее называют«принципом соответствия Л. А. Зенкевича». Если образование планктонной (поверхностной) пленки обусловлено проникновением солнечного света в верхние слои океана, то скопление жизни в донной пленке жизни определяется наличием… дна. Конечно, дно само по себе не кормит, но оно задерживает все то, что не успели съесть раньше. Кроме того, твердый субстрат дает возможность укрытия (а в океане укрытий не так-то много). Биосферная роль бентали — донной пленки жизни — не меньше, чем планктонной. Если планктонная пленка — огород океана, то бенталь — это склад его готовой продукции. Здесь на века замуровывается то, что, будучи создано живым веществом океана, ускользнуло из биотического круговорота благодаря специфической обстановке бентали. Планктонная пленка жизни поставляет сырье для осадков, а донная пленка — основной из экогоризонтов океана, где происходит их накопление. Основной, но, как показали недавние исследования, не единственный. Ювеналий Петрович Зайцев, член-корреспондент АН УССР, открыл интересное явление — «антидождь трупов». Оказалось, что после смерти всплывают тела не только крупных организмов (как это считалось раньше), но и всякой мелкой морской живности. В конце концов они, конечно, тонут, однако, находясь в поверхностном слое, успевают значительно обогатить его растворенным органическим веществом. Другой источник неживой органики в приповерхностном слое — органическое вещество, адсорбированное пузырьками газа, поднимающимися из морских глубин. Это явление дает органики в 10 раз больше, чем образуется ее здесь в процессе фотосинтеза. В результате у поверхности моря накапливается много органического вещества — главным образом в коллоидной форме. Во время штормов оно сбивается в белоснежную пену — ту самую, из которой, согласно древнегреческому мифу, родилась Венера, богиня любви и красоты… Население приповерхностного слоя водной толщи очень своеобразно. 5‑сантиметровый слой воды перехватывает 40% солнечного излучения, причем поглощается главным образом ультрафиолетовая часть спектра. Однако, как это ни парадоксально, фотосинтез тут подавлен. Основание трофической пирамиды составляют сапротрофные микроорганизмы, перерабатывающие неживое органическое вещество, благо оно здесь в избытке. Количество бактерий в приповерхностном слое в сотни и тысячи раз больше, чем в нижележащих горизонтах водной толщи. Вторую ступень трофической пирамиды составляют мельчайшие гетеротрофы — простейшие, личинки всевозможных моллюсков, червей, ракообразных, рыб и других животных. Очень много в приповерхностном слое молоди и зародышей организмов — икринок, которые держатся у самой поверхности воды. Наконец, последующие ступени трофической пирамиды составляют более крупные беспозвоночные, рыбы и морские птицы. «Инкубатором пелагиали» назвал Ю. П. Зайцев приповерхностный слой океана. Здесь концентрируются будущие поколения морских обитателей. В то же время именно поверхность моря сейчас больше всего подвергается загрязнению нефтепродуктами, и из-за этого гибнет бесчисленное количество нежной молоди. По выражению Ю. П. Зайцева (1974), «область максимума отрицательного воздействия на живое совпадает с областью максимума чувствительности населения». В опасности завтрашний день океана… Помимо жизненных пленок, в океане существуют сгущения и разрежения жизни. Пустыни на суше человечеству известны давно; в океане они открыты сравнительно недавно. Так, огромная водная пустыня обнаружена в Тихом океане в районе Гавайских островов. Концентрация живого вещества здесь составляет едва 3·10−6%. Иначе говоря, чтобы набрать литровую банку морских организмов, пришлось бы профильтровать ни много ни мало — 30 млн. л океанской воды. Сгущения жизни в океане В. И. Вернадский разделяет на три типа: а) прибрежные; б) саргассовые; 3) рифовые (см. рис. 2). Прибрежные сгущения жизни, по существу, возникают там, где встречаются обе пленки жизни — планктонная и донная — и сочетают в себе те жизненные блага, которые они несут: солнечный свет и твердый субстрат. К тому же прибрежные сгущения жизни характеризуются обильным притоком минеральных и органических веществ с континента и имеют еще одно благо — интенсивное перемешивание водной толщи (это делает возможным многократное использование элементов минерального питания). В результате сочетания всех этих благ в прибрежных сгущениях биомасса планктона в сотни, а бентоса — во многие тысячи раз больше, чем в глубоководных районах. «Море как часть биосферы начинается не постепенно, как, скажем, лес начинается с разреженной опушки, — пишет Ю. П. Зайцев. — Море начинается сразу со сгустка организмов, с многочисленных и уплотненных сообществ». Соответственно высокой оказывается и степень переработки осадочного материала. В прибрежных сгущениях жизни она такова, что экскременты морских организмов зачастую оказываются основным компонентом осадков. Отличительной чертой прибрежных сгущений жизни является преобладание многоклеточных, а не одноклеточных организмов. Характерно при этом сравнительное однообразие жизни в прибрежных сгущениях — от одного до 12 видов организмов здесь могут составлять до 95% биоценозов. Примером таких однородных скоплений являются мидиевые «банки». Во всех морях, кроме тропических, мидии образуют мощный прибрежный бордюр. Их биомасса иногда достигает десятков килограммов на квадратный метр дна. Иллюстрацией деятельности прибрежных скоплений жизни является исследование мидиевых «банок» Белого моря, выполненное К. А. Воскресенским в 40‑е годы под непосредственным влиянием идей Вернадского. К. А. Воскресенский выполнил удивительную работу — если не единственную в своем роде, то во всяком случае одну из немногих в то время и тем особенно ценную. Лабораторными экспериментами и натурными наблюдениями он доказал, что биофильтрационная деятельность мидиевых «банок» регулирует коллоидный состав прибрежных вод, осадконакопление в пределах отмелого побережья и — трудно в это поверить сразу! — даже циркуляцию вод в прибрежной зоне. Все эти функции осуществляются путем фильтрации мидиями прибрежных вод. Биогенная циркуляция создается за счет того, что вода, освобожденная от коллоидных частиц, становится легче и поднимается вверх. Другой тип концентрации жизни представляют собой саргассовые сгущения — участки моря, переполненные многоклеточными, не прикрепленными ко дну водорослями: саргассумом или филлофорой. Такой тип сгущения характеризуется очень высокой биомассой и чрезвычайно низкой продуктивностью живого вещества. Это чудо природы — «траву», плавающую в открытом море, там, где глубина достигает 4—6 км, первым описал Христофор Колумб. «С рассветом встретили столько травы, что море, казалось, покрылось ею, как льдом. Траву несло с запада» — такую запись сделал он в своем дневнике 21 октября 1492 года. А на следующий день записал: «В течение части дня травы не было, потом она пошла очень густо». Свое название бурая водоросль саргассум получила от португальского слова «саргасс» — гроздь винограда. Это желто-коричневые растения с сильно рассеченными «листьями», характерную особенность которых составляют шаровидные пузырьки, наполненные воздухом (они-то и напоминают виноград). Спутанная масса водорослей и обитающих здесь животных представляет собой саргассовое сгущение жизни, выделенное Вернадским. Классическим примером сгущения жизни такого рода является экосистема Саргассового моря, расположенного у восточных берегов Северной Америки. Его биомасса составляет 4—11 млн. т, т. е. примерно 1% всей биомассы живого вещества фотоавтотрофов Мирового океана. Низкая же годовая продуктивность объясняется своеобразным гидрологическим режимом моря: оно ограничено по контуру эллиптическим течением, и вертикальная перемешиваемость вод слабая. На территории нашей страны саргассовое сгущение жизни впервые было обнаружено в 1908 г. в северо-западной части Черного моря одним из основоположников отечественной гидробиологии академиком Сергеем Алексеевичем Зерновым (1871—1945). Здесь на площади около 11 000 км² на мелководье располагается скопление не прикрепленной ко дну багрянки филлофоры, по биомассе (5,5 млн. т) не уступающее сгущению Саргассового моря. Сейчас здесь, на «Филлофорном поле Зернова» (так его называют в честь первооткрывателя), организована добыча филлофоры для производства агароида, который находит применение в различных отраслях хозяйства. Существовали ли саргассовые сгущения жизни в геологическом прошлом? Известные советские палеонтологи Р. Ф. Геккер и Р. Л. Мерклин еще в 40‑е годы высказали хорошо подтвержденное фактами предположение о существовании саргассового сгущения 30 млн. лет назад в олигоценовом море, занимавшем территорию нынешнего Северного Кавказа. Польские ученые А. Ерзманьская и Я. Котларчук считают, что другое такое сгущение жизни одновременно возникло и на территории современных Карпат. Начиная с миоцена (15 млн. лет) под действием тектонических движений и морских течений оно стало перемещаться на запад. К настоящему времени, преодолев 8 тыс. км, оно достигло… Саргассового моря. Если это действительно так, сгущение жизни Саргассового моря — единственное в своем роде, совершившее столь длительное и далекое путешествие по морским просторам. Рифовое сгущение, коралловые рифы, — третий и последний тип из известных во времена Вернадского сгущений жизни. Коралловые рифы — это массовые мелководные поселения коралловых полипов и других морских организмов, имеющих твердый известковый скелет. Поскольку наиболее известными современными рифостроителями являются кораллы, все рифы иногда называют коралловыми, однако это не совсем верно. Помимо кораллов, рифы создаются деятельностью разнообразных живых существ: зеленых и красных водорослей, моллюсков, иглокожих и других организмов, причем пальма первенства принадлежит зеленым водорослям рода Halimeda и красным Lithotamnion. Ткани самих кораллов буквально набиты фотосинтезирующими симбиотическими водорослями зооксантеллами. В создании автотрофного живого вещества коралловых рифов большую роль играют также цианобактерии. Рифовые сгущения жизни — одна из самых высокопродуктивных экосистем биосферы. Растут рифы со скоростью 10 см/год, а иногда и до 25. Общая площадь всех рифов мира составляет 600 тыс. км² (что равняется территории Мадагаскара). Один лишь Большой Барьерный риф, протянувшийся вдоль восточного побережья Австралии, превосходит по объему все постройки человека. Состоят рифы из карбоната кальция. В наше время этим никого не удивишь, но попробуйте представить себе, какое впечатление произвела эта новость на плантатора прошлого века, живущего на коралловом острове и годами ввозящего известь из Англии! Об этом случае рассказал один путешественник того времени. Высокая продуктивность экосистемы коралловых рифов в значительной мере объясняется пористостью самого рифового сооружения: рифообразующие организмы могут «прогонять» через себя огромные объемы воды, отфильтровывая питательные вещества. Честь открытия этого интереснейшего явления принадлежит советскому ученому доктору биологических наук Юрию Ивановичу Сорокину. По его расчетам, вся толща океанских вод профильтровывается рифами за 40 тыс. лет. Благодаря биофильтрации значительных водных масс низкая концентрация биогенных элементов не лимитирует развитие коралловых рифов. Вековечную проблему океанской жизни — «биогенные соли в глубине» — эта экосистема разрешает вполне успешно. Область распространения современных рифов ограничена водами, имеющими на протяжении года температуру на поверхности не ниже 18°. В силу этого ограничения рифовые сгущения жизни сосредоточены в полосе между 37° с. ш. и 37° ю. ш. Есть у рифообразователей и другие «капризы» — морская вода должна иметь определенную соленость (от 2,7 до 4,0%), быть незамутненной, содержать много кислорода. Наконец, кораллам (точнее, живущим в симбиозе с ними водорослям) нужен свет, и поэтому они живут на глубине не более 40—50 м. А мощность коралловых рифов иногда достигает 1200—1400 м. Каким же образом остатки мелководных организмов оказались на такой значительной глубине? Первым правильно сумел ответить на этот вопрос Чарлз Дарвин в классическом труде «Коралловые рифы» (1842). Он показал, что формирование коралловых рифов происходит при непрерывном погружении морского дна. Своим ростом кораллы (и другие организмы) компенсируют это погружение, и глубина моря остается примерно одинаковой. А поскольку для длительного роста рифов требуется сохранение скорости прогибания на одном и том же месте, цепочки рифов обычно бывают приурочены к крупным долго живущим разломам. Это обстоятельство позволяет использовать древние рифы для реконструкции тектонического режима геологического прошлого. Ископаемые рифы имеют широкое распространение. По мнению академика Д. В. Наливкина, в кайнозое и мезозое рифовые известняки составляют главную массу всех известняков. В Ленинграде рифовые известняки можно увидеть не только в Геологоразведочном музее: они использовались для отделки подземного вестибюля станции метро «Площадь Восстания» (а в Москве — станция «Комсомольская»). Присмотревшись, в них можно найти органические остатки — обломки морских лилий (название может ввести в заблуждение: морские лилии — не растения, а животные, ныне вымирающие). Мы рассмотрели сгущения жизни в океане, выделенные В. И. Вернадским. Современной наукой установлены еще два типа характерных сгущений: апвеллинговое и абиссальное рифтовое. Апвеллингом (от англ. up — наверх и to well — хлынуть) называется явление подъема глубинных океанских вод к поверхности. А поскольку на большей части Мирового океана развитие жизни лимитируется главным образом недостатком неорганического азота и фосфора, то любой подъем на поверхность донных вод, обогащенных минеральными элементами и углекислотой, оказывает благотворное влияние на развитие жизни. Особенно ярко эта закономерность проявляется в тропических и субтропических районах. Поэтому именно здесь сосредоточены все крупнейшие апвеллинги планетарного масштаба: Канарский, Бенгельский, Сомалийский, Калифорнийский, Перуанский и Экваториальный. Вот как описывает современный путешественник апвеллинговое сгущение жизни у берегов Аравийского полуострова: «Казалось, что даже в самой атмосфере района апвеллинга ощущалось богатство морской жизни. По сравнению с кондиционированным воздухом в помещениях на судне холодный влажный воздух моря имел вполне определенный стойкий запах рыбы… Стаи дельфинов, порой столь многочисленные, что, казалось, они простираются до самого горизонта, окружали судно, отставая лишь на мгновение, чтобы порезвиться в волнах. Все море, казалось, кипело жизнью». Воды, обогащенные элементами минерального питания и углекислотой, обычно поднимаются с небольших глубин: приблизительно 100 м и очень редко до 300 м. Скорость подъема вод очень небольшая — от 1 до 5 м в день (представьте лифт, который поднимается с такой скоростью!). Самая мощная продуцирующая система Мирового океана находится в зоне Перуанского апвеллинга. С его площади, занимающей всего 0,02% всей акватории океана, получают до 15—20% мирового улова рыбы. О безнадежных затеях иногда говорят: «Ищи ветра в поле». Еще труднее, пожалуй, отыскать следы течения в море, существовавшем миллионы лет назад. Выручают некоторые характерные виды радиолярий и своеобразные сочетания различных типов осадков, по которым сейчас успешно выявляют древние апвеллинговые сгущения жизни. Так, львовский ученый, доктор геолого-минералогических наук Юрий Николаевич Сеньковский на основании исследования кремнистых пород недавно оконтурил апвеллинг в море, существовавшем на территории Карпат в меловое время. Выполнены и более древние реконструкции. Показано, например, что в палеозое апвеллинговые сгущения жизни были приурочены к окраинам Сибири, Австралии и Северной Америки. Свидетельством этого являются залежи морских фосфоритов, горючих сланцев и кремнистых толщ в этих регионах. Наконец, последним из ныне известных сгущений жизни в океане (рис. 3) являются абиссальные рифтовые (не путать с рифовыми!). Эти сгущения жизни — бесспорно, самые удивительные! — впервые предстали перед взором человека 15 февраля 1977 г., когда американский подводный аппарат «Алвин» в 280 км к северо-востоку от Галапагосских островов достиг дна Тихого океана на глубине 2540 м. Американские ученые под руководством Р. Бэлларда производили в то время исследования гидротермальных источников рифтов — зон раздвижения земной коры. Один из наблюдателей, профессор Массачусетского технологического института Дж. М. Эдмонд, так описывает это историческое погружение: «Типичный базальтовый ландшафт выглядел довольно уныло: монотонные поля бурых «подушек»[43] разбиты многочисленными трещинами; на площади несколько квадратных метров не всегда можно было увидеть живое существо… Но здесь мы оказались в оазисе. Рифы из мидий и целые поля гигантских двустворок, крабы, актинии и крупные рыбы, казалось, купались в мерцающей воде… Мы наткнулись на поле горячих источников. Внутри круга диаметром около 100 м теплая вода струилась из каждой расселины, каждого отверстия в морском дне». Слово «оазисы» сразу было подхвачено мировой прессой, их цветные фотографии появились во многих иллюстрированных журналах. Однако поначалу было совершенно непонятно, откуда в абиссали, в царстве вечного мрака, такой расцвет жизни? Во всех руководствах по экологии, вплоть до новейших, неизменно повторяется одна и та же мысль: «Без растений экосистемы не могут существовать, так как у них не будет источника органического вещества, а это означает то же самое, что отсутствие пищи». Чем же при полном отсутствии растений (откуда им взяться на глубине 2,5 км?) питаются эти странные гиганты? Ведь нежиться даже в самой теплой воде можно только при наличии надежной кормовой базы!
 Рис. 3. Абиссальное рифтовое сгущение жизни. На переднем плане — рифтии (слева и справа), двустворчатые моллюски (среди «подушечной лавы»), крабы, рыбы. На среднем и заднем плане — «черные курильщики» (в виде труб) и «белые курильщики», покрытые «зарослями» полихет. На заднем плане справа — прекративший свое существование «черный курильщик» с распадающимся сообществом организмов
Рис. 3. Абиссальное рифтовое сгущение жизни. На переднем плане — рифтии (слева и справа), двустворчатые моллюски (среди «подушечной лавы»), крабы, рыбы. На среднем и заднем плане — «черные курильщики» (в виде труб) и «белые курильщики», покрытые «зарослями» полихет. На заднем плане справа — прекративший свое существование «черный курильщик» с распадающимся сообществом организмов
«Есть многое на свете, друг Горацио, что и не снилось нашим мудрецам», — обронил как-то шекспировский Гамлет. Действительно, кто мог бы себе представить, что в рифтовых сгущениях основой жизни окажутся не фотоавтотрофы, как везде на Земле, а хемоавтотрофные микроорганизмы? Между тем исследования показали, что дело обстоит именно так. Гидротермальные источники, действующие на дне океана, несут эндогенный сероводород, который используют большинство хемоавтотрофов (некоторые микроорганизмы утилизируют водород, аммиак, NO2, Fe2+, Mn2+). Следующее трофическое звено — макрофауна — питается хемоавтотрофными микроорганизмами (архебактериями и бактериями), и, таким образом, эти абиссальные сообщества (единственные в своем роде!) являются приемниками эндогенной, а не солнечной энергии. В этом — первая и самая главная отличительная особенность абиссальных рифтовых сгущений жизни. Вторая особенность — широкое распространение симбиотрофных организмов. Хемоавтотрофные бактерии зачастую (но не всегда!) функционируют прямо в теле многоклеточных организмов и даже образуют там кристаллики самородной серы. В особом органе погонофор, где сосредоточены бактерии, их численность достигает 3,7 млрд. клеток на 1 г веса. Пищеварительная система у такого рода симбиотрофных организмов полностью атрофирована: у них нет ни рта, ни желудка, ни кишечника (это, кстати, причиняет большие неприятности ученым — из-за отсутствия «брюха» они не могут решить, где у этих организмов брюшная, а где — спинная сторона тела). Симбиоз многоклеточных животных с фотоавтотрофами был известен и ранее, однако их сосуществование с хемоавтотрофами впервые было установлено в абиссальных рифтовых сгущениях жизни (впоследствии такого рода симбионты были найдены и в других экосистемах). Третьей отличительной особенностью рифтовых сгущений жизни является удивительное своеобразие их органического мира. Он настолько экзотичен, что каждый вновь открытый оазис получал собственное название: «Розовый сад» (он был первым), «Райский сад», «Мидиевая банка», «Одуванчики», «Лужайка для пикников»… В абиссальных рифтовых сгущениях описано больше десятка новых семейств и подсемейств кольчатых червей, погонофор, гастропод, ракообразных, много новых родов разных типов. Только в последнем по времени открытия оазисе на подводном хребте Хуан-де-Фука зафиксировано 14 новых видов многоклеточных животных. Многие из обитателей абиссальных сгущений пока не попали в механические «руки» (манипуляторы) подводных аппаратов и известны лишь по фотографиям. Самым характерным обитателем рифтовых сгущений жизни являются рифтии — представители новооткрытого рода погонофор, получившего свое родовое название по обитанию в районе рифтов. По описаниям очевидцев, их перепутанные, срастающиеся концами белые трубки и красные плюмажи сразу бросаются в глаза при приближении подводного аппарата к сгущениям жизни. Рифтии — крупные червеобразные животные длиной до 1,5 м при диаметре 3,5—4 см, живущие в гибких цилиндрических трубках из белка и хитина. Это типичные симбиотрофные животные, питающиеся за счет «вмонтированных» в них хемоавтотрофных бактерий. Другими характерными обитателями абиссальных рифтовых сгущений являются полихеты, подобно рифтиям, живущие в трубках (но не хитиновых, а известковых). Один из новооткрытых представителей полихет получил название помпейского червя за свое близкое поселение к гидротермальным источникам, извергающим дымы (их называют белыми и черными курильщиками). Слепые крабы и рыбы в рифтовых сгущениях жизни пасутся на «зарослях» рифтий и полихет, как коровы на заливных лугах. А вдоль трещин, по которым изливаются гидротермы, селятся гигантские (до 30 см) двустворчатые моллюски. Их плоть, как и у рифтий, инкрустирована тионовыми бактериями. Гигантизм обитателей — еще одна отличительная особенность рифтовых сгущений жизни. Длина «обычных» (не рифтовых) погонофор, как правило, не превышает 10 см. Рифтии же, как упоминалось, достигают 1,5 м. Под стать им и гигантские двустворчатые моллюски размером с суповую тарелку или даже блюдо (до 25—30 см в поперечнике) и с толщиной раковины до 8 мм. Растут они в 500 раз быстрее, чем их ближайшие родственники, живущие вне зоны рифта и имеющие на порядок меньшие размеры (2—3 см в поперечнике). Интересно, что «гигантомания» в рифтовых сгущениях затронула даже бактерии, которые достигают здесь 0,11 мм — величины, для этого подцарства органического мира вообще неслыханной! Плотность жизни в рифтовых сгущениях необычайно высока. В районе Галапагосских островов на глубине 2500 м биомасса одних только рифтий составляет от 10 до 15 кг/м², тогда как обычно на такой глубине плотность живого вещества донной пленки лишь 0,1—10 г/м². Поток эндогенного вещества повышает биомассу на 3—5 порядков! Эти расчеты показывают независимость рифтовых сгущений жизни от солнечной энергии — при полнейшей подчиненности гидротермам, длительность действия которых исчисляется, по-видимому, первыми десятками лет. Ученые французской экспедиции «Биоспариз», посетившие в 1984 г. открытые ими ранее оазисы абиссальной жизни, зафиксировали серьезные изменения в структуре сообществ, произошедшие всего за два года. Там, где гидротермальная деятельность за это время прекратилась, распались и сообщества организмов. Лишь скопления раковин гигантских моллюсков порой свидетельствовали о недавнем расцвете жизни. Размеры оазисов небольшие: десятки метров в поперечнике — как лужайки в лесу или залы музеев. Все известные до последнего времени оазисы расположены в восточной части Тихого океана на глубинах 1500—3000 м, причем большая их часть — на оси срединно-океанического Восточно-Тихоокеанского поднятия: на 21° с. ш., 10—13° с. ш., у экватора (Галапагосский рифт) и у острова Пасхи (27° ю. ш.). Самое северное сгущение жизни находится за пределами Восточно-Тихоокеанского поднятия, в проливе Хуан-де-Фука у южной оконечности острова Ванкувер. В пределах каждого из этих участков, как правило, имеется несколько изолированных эфемерных оазисов, расположенных друг от друга на расстоянии нескольких сотен метров. Общая протяженность меридианального пояса рифтовых сгущений жизни составляет в Тихом океане около 8 тыс. км. В марте 1984 г. учеными Скриппсовского океанографического института в США было сделано неожиданное открытие: «рифтовое» сгущение жизни было встречено вне зоны рифта — в Атлантическом океане, недалеко от побережья Флориды, на глубине 3266 м. Первооткрыватели не зафиксировали в нем каких-либо специфических особенностей, отличающих его от ранее известных, «подлинно рифтовых» сгущений жизни. Источником питания хемоавтотрофных микроорганизмов здесь также служил сероводород, однако последний поступал не из рифта, а из зоны шельфа. На этом основании ученые сделали далеко идущий вывод, что подобные сообщества могут существовать не только в рифтовых зонах, как ранее предполагалось, и не требуют повышенных температур: им необходимы лишь восстановленные неорганические соединения, используемые в качестве пищи. Будем ждать новых открытий. Пока трудно судить, насколько значительной в общем балансе вещества и энергии в океане может быть роль абиссальных рифтовых сгущений жизни (до выяснения обстоятельств это название сохраняем). Не исключено, что в самих рифтовых сообществах используется лишь небольшая часть первичной продукции, создаваемой хемоавтотрофными бактериями. Как предполагает французский ученый Мишель Ру, остальная ее часть рассеивается на значительной площади акватории Мирового океана. Наряду с планктонной пленкой жизни абиссальные рифтовые сообщества могут оказаться еще одним «огородом» океана — природа, как известно, любит дублировать сконструированные ею системы жизнеобеспечения. Когда появились в биосфере абиссальные рифтовые сгущения жизни? По крайней мере 570 млн. лет назад — именно такой возраст имеет древнейшая достоверная находка их ископаемых остатков, сделанная на территории Ирландии. Впрочем, в отложениях венда (670—590 млн. лет) академиком Борисом Сергеевичем Соколовым уже давно были описаны остатки трубчатых организмов Sabelliditida, которые, как полагают теперь французские ученые супруги Термье, могут являться предками рифтий. Если это действительно так, рифтовые сгущения жизни — одни из древнейших экосистем, существующих на нашей планете. Не нуждаясь в солнечном свете, они могли возникнуть даже до появления на Земле первых фотоавтотрофов. Так это или нет, покажут дальнейшие исследования — геологических свидетельств пока явно недостаточно. Открытие абиссальных рифтовых сгущений жизни справедливо расценивается как одно из крупнейших биологических открытий, сделанных в 70‑е годы. Их исследования интенсивно продолжаются. И можно согласиться с американским ученым Дж. Ф. Грэсслом, который сказал, что после открытия абиссальных рифтовых сгущений «изменилось наше представление о жизни в глубинах морей, и еще много сюрпризов ждет нас в ближайшем будущем». Можно сказать и больше — коренным образом изменилось и наше прежнее представление о биосфере в целом: теперь уже о ней невозможно говорить как о системе, ход жизни в которой поддерживается исключительно первичной продукцией фотоавтотрофов. Таковы современные данные о сгущениях жизни в океане. Экосистемы суши значительно отличаются от морских, хотя имеются и некоторые черты сходства. Как и в океане, на суше имеются две пленки жизни, причем верхняя также расположена в фотобиосфере, а нижняя — в меланобиосфере. Но на этом сходство, по существу, и кончается. Верхняя пленка жизни на суше — наземная; она идет вверх от поверхности почвы до верхней границы биосферы. Это — привычные нам ландшафты, которые описывать нет необходимости. Ниже располагается весьма специфическая почвенная пленка жизни, представляющая собой густо заселенную сложную трехфазную систему. Как показал М. С. Гиляров[44], трехфазность, пористость и агрегатный характер почвы дают возможность существования самых разнообразных экологических групп организмов. Так, для микроорганизмов даже отдельная частица почвы обеспечивает одновременную активность как аэробных (на поверхности частицы), так и анаэробных (внутри нее) организмов. Простейшие, коловратки, мелкие нематоды живут в водной среде: в пленке воды, обволакивающей твердые частицы почвы. А для несколько более крупных обитателей, например мелких клещей, почва представляет как бы разветвленную систему пещер. И лишь для крупных обитателей (дождевых червей, личинок насекомых и т. д.) средой обитания является почва в целом. Концентрация жизни в почвенной пленке выше, чем в наземной. Так, в 1 см³ лесной почвы насчитывается в среднем 10 млн. бактерий, 200 тыс. микроскопических водорослей и 20 тыс. простейших, а длина содержащихся в нем гифов грибов достигает фантастической величины — 2 км! Деятельности живого вещества в почве В. И. Вернадский посвятил специальную работу, рукопись которой недавно была обнаружена в Киеве[45]. На суше обе пленки жизни имеют непосредственный контакт, а не разделяются по вертикали несколькими километрами слабо заселенного жизнью пространства, как в океане. Здесь нет проблемы, ограничивающей продуктивность океана («биогенные соли на глубине»), и потребные им элементы фотоавтотрофные организмы добывают из почвы. Туда же после отмирания попадают их остатки — необиогенное органическое вещество. Такое тесное взаимодействие пленок жизни приводит к тому, что их часто объединяют в общее понятие биогеоценотического покрова. Ниже под тонким слоем почвы (ее мощность составляет несколько дециметров, реже 1—1,5 м) располагается область подземного разрежения жизни, которую болгарский ученый, профессор Любомир Цветков недавно предложил называть стигобиосферой (от названия мифической реки Стикс, согласно верованиям древних греков, протекающей через подземное царство мертвых Аид). Стигобиосфера — мир, слабо затронутый научным исследованием. За редким исключением, каковое представляют пещеры, здесь везде — от нижней границы почвы до нижнего предела биосферы — возможна главным образом бактериальная, микроскопическая, жизнь. Сферой своего обитания подземные узники выбирают влагу — они живут в подземных водах и на их контакте с горными породами. На материале изучения ряда регионов Советского Союза крупный ленинградский гидрогеолог и геохимик Марк Савельевич Гуревич (1910—1975) установил в 1967 г. наличие сопряженной микробиологической и биогидрогеохимической зональности подземных вод. В дальнейшем эти работы были продолжены микробиологом Людмилой Евстафьевной Крамаренко (табл. 3). По существу, зоны, выделяемые М. С. Гуревичем и Л. Е. Крамаренко, являются определенными экогоризонтами подпочвенной части меланобиосферы в ее континентальной части. Таблица 3 Схема гидробиохимической зональности земной коры, по Л. Е. Крамаренко (1975, 1983)
| Зоны | Eh, мВ | Микроорганизмы (примеры) | К[46] | Продукты жизнедеятельности бактерий (биогенное вещество) |
|---|---|---|---|---|
| Аэробная (I) | +800÷+100 | Тионовые, нитрифицирующие, метаноокисляющие, водородокисляющие бактерии | До ∞ | SO42−, NO3−, NO2−, CO2, окисные формы металлов |
| Смешанная (II) | +100÷−200 | Тионовые, нитрифицирующие, водородокисляющие, денитрифицирующие, сульфатвосстанавливающие, метанпродуцирующие, водородпродуцирующие бактерии | Около 1 | SO42−, NO3−, NO2−, CO2, окисные формы металлов H2S, N2, H2, CH4, восстановленные формы металлов |
| Анаэробная (III) | −200÷−400 и ниже | Сульфатвосстанавливающие, денитрифицирующие, метанпродуцирующие, водородпродуцирующие бактерии | До 0 | H2S, N2, H2, CH4, восстановленные формы металлов |
| Отсутствия бактерий (IV) | Не обнаруживаются | CH4, N2, H2S, восстановленные формы металлов |
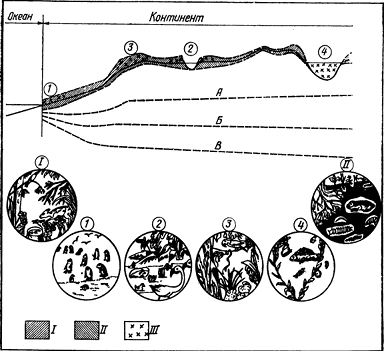 Рис. 4. Экогоризонты, концентрации и разрежения жизни на суше: I — наземная пленка жизни; II — почвенная пленка жизни; III — сгущения жизни; 1 — береговое; 2 — пойменное; 3 — влажных дождевых лесов тропиков и, отчасти субтропиков; 4 — сгущения стоячих водоемов; А—В — разрежения жизни: А — аэробный подземный экогоризонт; Б — аэробно-анаэробный подземный экогоризонт; В — анаэробный подземный экогоризонт
Рис. 4. Экогоризонты, концентрации и разрежения жизни на суше: I — наземная пленка жизни; II — почвенная пленка жизни; III — сгущения жизни; 1 — береговое; 2 — пойменное; 3 — влажных дождевых лесов тропиков и, отчасти субтропиков; 4 — сгущения стоячих водоемов; А—В — разрежения жизни: А — аэробный подземный экогоризонт; Б — аэробно-анаэробный подземный экогоризонт; В — анаэробный подземный экогоризонт
Живое вещество распределяется неравномерно не только в вертикальном сечении биосферы, но и по площади. Владимир Иванович выделял на суше: а) береговые и б) пойменные сгущения жизни. Учитывая современные данные по определению биомассы различных экосистем, выполненные Н. И. Базилевич, Л. Е. Родиным и И. Н. Розовым, можно выделить третий тип сгущений такого рода — влажных дождевых лесов тропиков и отчасти субтропиков. Береговые сгущения жизни, по Вернадскому, представляют собой морские побережья и прибрежные территории суши. Высокая концентрация живого вещества обусловлена здесь благоприятной обстановкой, создаваемой контактом двух основных сред обитания живых организмов — морской и наземной. Два соседних сообщества — наземное, береговое, и морское, прибрежное, — непосредственно примыкают друг к другу и находятся в тесном взаимодействии. Однако везде ли на Земле морские побережья являются сгущениями жизни? Посмотрим на физическую карту мира. Значительную часть прибрежных территорий Америки, Африки, Азии и Австралии занимают пустыни. Арктические и антарктические побережья по абсолютным цифрам плотности живого вещества также нельзя отнести к сгущениям жизни. Очевидно, что на концентрацию жизни большое влияние оказывает и климатический фактор. Однако если мы сравним плотность жизни в центре Антарктического континента или Гренландии и на их побережьях, то убедимся, что для данных климатических условий побережье, действительно, сгущением жизни является. Лишь при благоприятном климате, например на Средиземноморье, морские побережья служат наглядным примером сгущений жизни в абсолютном выражении. Второй тип сгущения жизни на материках — пойменный — Вернадский охарактеризовал в «Очерках геохимии» как «скопление жизни в бассейнах великих рек», включая сюда не только плодородные долины рек, но и их дельты. По современным данным, эти экосистемы характеризуются наивысшей на суше продуктивностью: при ничтожной площади (меньше 1%) они производят 10% живого вещества. Увеличение продукции более чем на порядок против среднего стандарта наземных экосистем объясняется обильным снабжением пойм и особенно дельт элементами минерального питания. Вернадский в качестве примеров пойменных сгущений жизни приводил Амазонку, Ориноко, Замбези, Обь и Иртыш. Однако и здесь климатический фактор вносит свои коррективы — наиболее значительной фитомассой обладают экосистемы поймы только в пределах субтропического и тропического пояса. Поэтому долины наших великих рек Оби и Иртыша примерами сгущений живого вещества служить не могут. «По-разному соткан ковер, накинутый богатой цветами флорой на обнаженное тело земли; он гуще там, где солнце поднимается выше на всегда безоблачном небе, — и реже у полюсов, где постоянно возвращающиеся морозы убивают то распускающуюся почку, то созревающий плод», — писал Александр Гумбольдт. Там, где «солнце поднимается выше», и располагается последний тип сгущений живого вещества на континентах — влажных дождевых лесов тропиков и отчасти субтропиков. Их фитомасса достигает рекордной величины: 650 т/га (в нашей тайге 200—250 т/га), а суммарная фитомасса тропических и субтропических лесов составляет около 60% всего живого вещества нашей планеты. Тропические леса, по образному сравнению Д. В. Панфилова, напоминают застывший в падении зеленый водопад. Гетеротрофы по биомассе значительно уступают фотоавтотрофам и отличаются поразительным разнообразием видового состава. Так, во влажных тропических лесах на площади всего в несколько квадратных километров встречается в среднем 97 видов млекопитающих, 464 вида птиц, 93 вида пресмыкающихся, 54 вида земноводных. При этом биомасса почвенных животных в 4 раза выше, чем биомасса наземных обитателей. Основу почвенной зоомассы составляют дождевые черви. Казалось бы, при огромной биомассе живого вещества во влажных дождевых лесах тропиков и субтропиков должно происходить интенсивное накопление необиогенного вещества. Однако поскольку деревья в этих лесах являются вечнозелеными, сезонного поступления отмершей органики в опад не происходит, и толщина лесной подстилки здесь не превышает нескольких сантиметров. Насекомые, главным образом термиты, очень быстро уничтожают отмершие части растений (опавшие листья, ветви, упавшие и даже стоящие на корню стволы отмерших деревьев). Толстые ветви деревьев уничтожаются насекомыми почти полностью за 3—5 лет, а лежащие на земле сухие листья и мелкие веточки исчезают по прошествии немногих месяцев. Завершают процесс разложения органики грибы и бактерии. В обилии почвенных сапротрофов в тропических лесах легко убедиться: достаточно провести ногой борозду по почве — и взору откроется густое переплетение беловатых нитей мицелиев грибов, живущих в симбиозе с древесными растениями. В результате столь активной деятельности сапротрофов необиогенное вещество во влажных дождевых лесах тропиков и субтропиков практически не накапливается. Отличительной особенностью живого вещества суши является его состав: абсолютным преобладанием по биомассе и продуктивности характеризуются высшие растения. Все остальные восемь подцарств органического мира находятся в резко подчиненном положении. Но и среди высших растений также существуют свои лидеры — ими являются древесные растения. Биомасса лесов еще совсем недавно составляла 84% от общей биомассы живого вещества суши (из них тропических и субтропических лесов, как мы помним, — 60%). Однако в последние годы биомасса лесов стремительно сокращается. Французский поэт Жак Превер пишет по этому поводу:
Все меньше и меньше остается лесов:
Их истребляют,
Их убивают,
Их сортируют
И в дело пускают.
Их превращают
В бумажную массу,
Из которой получают миллиарды газетных листов,
Настойчиво обращающих внимание публики
На крайнюю опасность истребления лесов.
Глава четвертая. Самая мощная геологическая сила
Невозможно понять историю осадочных пород, если не принимать во внимание мощного влияния организмов на миграцию веществ в земной коре и на образование осадков.Л. С. Берг. 1944
 Работа живого вещества в биосфере, по Вернадскому, может проявляться в двух основных формах: а) химической (биохимической) — I род геологической деятельности; б) механической — II род такой деятельности.
Геологическая деятельность I рода — построение тела организмов и переваривание пищи — конечно, является более значимой. Классическим стало функциональное определение жизни, данное Фридрихом Энгельсом: «Жизнь есть способ существования белковых тел, существенным моментом которого является постоянный обмен веществ с окружающей их внешней природой, причем с прекращением этого обмена веществ прекращается и жизнь»[49] (курсив Энгельса. — А. Л.).
Собственно говоря, постоянный обмен веществ между живым организмом и внешней средой и обусловливает проявление большинства функций живого вещества в биосфере, которые мы рассмотрим в этой главе книги. По подсчетам специалистов, в течение жизни человека через его тело проходит 75 т воды, 17 т углеводов, 2,5 т белков, 1,3 т жиров. Между тем по геохимическому эффекту своей геологической деятельности I рода человек отнюдь не самый важный вид разнородного живого вещества биосферы. Наиболее эффективную деятельность в этом отношении осуществляют на суше — грунтоеды (главным образом дождевые черви), а в океане — илоеды и фильтраторы.
Первым обратил внимание на геологическую деятельность дождевых червей Чарлз Дарвин. За год до своей смерти, в 1881 г., он выпустил фундаментальный труд «Образование растительного слоя деятельностью дождевых червей» (столетие со дня его выхода в свет было отмечено проведением Международного симпозиума по экологии дождевых червей). Дарвин убедительно доказал, что весь слой почвы непрерывно перерабатывается дождевыми червями. По его подсчетам, слой экскрементов, выделяемых дождевыми червями на плодородных почвах Англии, составляет около 5 мм в год. Отсюда следует, что почвенный пласт мощностью в 1 м дождевые черви полностью пропускают через свой кишечник за 200 лет!
Кажется, что дождевые черви созданы самой природой специально для пищеварения: они способны переваривать почти любые органические вещества, включая и новинки человеческой технологии. При этом дождевые черви заглатывают многие элементы в количествах, значительно превышающих их потребности, а избытки выделяют в окружающую среду. В результате почва, в которой находится достаточное количество червей, содержит вдвое больше магния, впятеро — азота, в 11 раз — калия, чем лишенная червей. Можно себе представить, каких размеров достигает суммарный геохимический эффект деятельности дождевых червей, если при этом учесть, что по произведенным в США расчетам биомасса червей там в 10 раз превышает массу человеческого населения!
В донных отложениях современных водоемов фекальные комочки беспозвоночных распространены также очень широко и нередко являются основной частью осадка. В Южной Атлантике, например, современные осадки илов почти нацело слагаются фекалиями фильтрующих планктонных ракообразных, а по берегам Северного моря донные осадки, образованные фекалиями мидий (также — фильтраторов), имеют мощность до 8 м. По своей «пропускной способности» с дождевыми червями в океане могут конкурировать их близкие родственники, представители того же типа кольчатых червей — полихеты. Как и в случае дождевых червей, химический состав субстрата при этом существенно изменяется: по сравнению с исходными илами он обогащается кальцием, железом, магнием, калием и фосфором.
Таким образом, пищеварительная деятельность грунтоедов на суше и илоедов и фильтраторов в океане является важным геохимическим фактором биосферы. По мнению ученых Ростовского университета, настало время говорить о рождении новой отрасли геохимии — трофической геохимии. Ее основной проблемой является исследование процессов химического преобразования субстрата при его прохождении через пищеварительный тракт массовых видов грунтоедов, илоедов и фильтраторов.
Ископаемые остатки фекалий — их называют копролитами — известны в геологических отложениях начиная с ордовика. Бесспорно, однако, что большая часть копролитов при геологических описаниях не учитывается. Происходит это из-за слабой изученности вопроса и из-за отсутствия четких диагностических признаков для определения копролитов. Чаще фиксируются в древних осадках следы проедания илоедов — бесспорные свидетельства существования в геологическом прошлом I рода геологической деятельности живого вещества.
Механическая работа живого вещества — II род геологической деятельности — проявляется как в наземных, так и в водных экосистемах.
Интересно, что первая научная работа В. И. Вернадского, написанная еще в студенческие годы, была посвящена именно механической деятельности живого вещества. Она была составлена В. И. Вернадским по материалам почвенной экспедиции 1884 г. в Екатеринославскую губернию и опубликована В. В. Докучаевым без ведома автора в 1889 г.[50] (Вернадский в это время находился в заграничной научной командировке). В этой работе В. И. Вернадский, едва перешагнувший за рубеж своего двадцатилетия, впервые в мировой литературе попытался количественно оценить влияние роющей деятельности сурка. Для самого Вернадского эта работа, видимо, послужила той своеобразной затравкой, из которой через 40 с лишним лет окончательно выкристаллизовалась стройная концепция биосферы.
Роющая деятельность в наземных экосистемах осуществляется главным образом млекопитающими и распространена повсеместно: в лесах, на лугах, в тундрах, степях, полупустынях и пустынях. При этом наибольшее развитие роющая деятельность животных получает в открытых ландшафтах аридной зоны из-за отсутствия там естественных убежищ. Мелкие млекопитающие этой зоны — сурки, суслики и слепыши — буквально перелопачивают почвы и подстилающие их отложения. Благодаря их выбросам в верхние слои почвы попадают первичные невыветрившиеся минералы, которые, разлагаясь, вовлекаются в биогеохимический круговорот. Попутно эти животные иногда оказывают ценные услуги геологам. Вспоминается следующий факт, описанный в научной литературе.
В годы Великой Отечественной войны возникла острая нехватка писчего мела — все его месторождения, расположенные на Украине и в Белгородской области, были захвачены врагом. Новое месторождение, пригодное для эксплуатации, в кратчайший срок было найдено в Казахстане по выбросам из нор сурков. Может быть, заслуга сурков и меньше, чем гусей (которые, как известно, «Рим спасли»), но добрым словом их помянуть стоит.
Как показали исследования Бориса Даниловича Абатурова, в экосистемах полупустыни роющая деятельность животных представляет собой одну из важнейших форм механической работы, ведущей к глубокому преобразованию почв и даже рельефа страны. Норы млекопитающих обеспечивают более интенсивное увлажнение почв атмосферными осадками и поступление воды в более глубокие ее горизонты. Роющая деятельность, продолжаясь в течение тысячелетий, стала важнейшим фактором формирования характерного для полупустынь микрорельефа «сусликогенного» облика, состоящего из микроповышений и западин.
В пустынях в результате совокупной деятельности ветра, растений (шишконосной эфедры), паукообразных, насекомых и мелких млекопитающих (полуденной песчанки) возникают целые «крепостные» сооружения высотой до 5 м. В увлекательной книге А. В. Смирнова «Мир растений» (1982) так описывается процесс их возникновения на примере Каракумов.
Все начинается с того, что вокруг кустиков эфедры ветер надувает холмики песка. Опавшие ветви эфедры цементируются песком, в котором много гипса. Образуется песчаный бугор с прочным куполом, в котором сохраняется постоянная температура. В нем поселяются песчанки, паукообразные, насекомые — обычные обитатели пустыни. Песчанки проделывают в бугре множество ходов, зачастую перегрызая корневища эфедры. Куст местами усыхает, но разрастается дальше своими корневищами. Каждый новый куст собирает вокруг себя песок, и история повторяется сначала. Так в пустыне появляются песчаные города с крепостными стенами (остатками первичного бугра) и башнями (вторичными дочерними буграми).
В тропиках подобными своеобразными памятниками механической деятельности живого вещества являются термитники. Их высота достигает 15 м! Если бы люди умели строить сооружения в таком же соотношении к размерам своего тела, как термиты, то Землю бы украшали небоскребы высотой до 3400 м. Исходный материал для своего строительства термиты, к вящей радости геологов, извлекают с глубины до 12 м (суркам до них далеко!). При этом, располагая термитники на линиях тектонических нарушений и в зонах повышенной трещиноватости пород, они оказывают геологам дополнительные услуги, помогая тем самым в составлении крупномасштабных структурногеоморфологических карт.
В лесных экосистемах важную механическую деятельность в виде строительства плотин осуществляют бобры. Длина некоторых бобровых плотин достигает 100 м, а рекорд, зафиксированный на Джефферсон-Ривер (штат Монтана, США), составляет 700 м! Течение рек из-за плотин замедляется; иногда образуется даже целая цепь прудов со слабо проточной водой и широкие выровненные поймы. В запрудах постепенно оседает ил. Если это происходит достаточно долго, то его отложения достигают большой мощности. При перемещении бобровых запруд или их исчезновении плодородные илы обнажаются, покрываясь густой растительностью. Таким путем в Северной Америке образовались богатые пастбища и луга, которые называют «бобровыми». Подобно человеку, но без использования техники, бобры преобразовали облик целой страны. Не зря индейцы считали бобра своим младшим братом, а у американцев существует пословица: «трудиться, как бобр».
Термитники в тропиках и «крепостные» сооружения в пустынях, сусликогенный рельеф полупустынь и бобровые луга лесной зоны — примеры грандиозной механической деятельности живого вещества в наземных экосистемах. С неменьшим успехом эта деятельность осуществляется живым веществом и в морских экосистемах.
На дне моря, как и на суше, организмы роют себе укрытия, причем не только в мягком, но и в скальном грунте. Олигохеты и полихеты углубляются в грунт на 40 см и более. Двустворчатые моллюски зарываются обычно неглубоко, но некоторые из них, например миа, роют норы, которым позавидует и сурок: они достигают глубины нескольких метров. В зоне прибоя и на перемываемом волнами песке — вот беда! — норы не выроешь и гнездо не совьешь. Приходится сверлить скальные породы. И сверлят. Сверлят водоросли и губки, бактерии и моллюски, цианобактерии, полихеты, морские ежи, рачки, мшанки… При этом, если сверление производится механически, образуются мельчайшие «опилки» диаметром в десятки микронов. Только сверлящие губки, живущие на 1 м² морского дна, выделяют за год 250 г таких «опилок».
Сверлильщики появились в далеком геологическом прошлом. Источенные ими породы находят даже в докембрийских отложениях, и поныне они продолжают свою разрушительную работу. Сверлящая деятельность моллюсков фолад вызывает иногда катастрофические последствия. Когда недавно в районе Сочи в результате непродуманного строительства берег лишился гальки, он начал отступать со скоростью до 4 м в год. Главным виновником разрушения были фолады, которые заселили каждый метр скального берега, сложенного сланцами, и принялись дружно сверлить себе подводные норки. К счастью, был найден выход: берег стали укреплять поперечными стенками, а между ними засыпать гальку. В результате сверлильщики были уничтожены: движущаяся под ударами волн галька перемолола их. А в Западной Европе не менее опасную деятельность производит случайно завезенный из Китая мохнаторукий краб — он проник во многие реки и, строя свои норы, подрывает берега и разрушает плотины.
Механическая деятельность живого вещества в значительной мере определяет микрорельеф морского дна. Положительные формы рельефа создаются активными фильтраторами и морской растительностью, скрепляющей отложения, а отрицательные — теми бентосными организмами, которые активно взмучивают ил: бентосными рыбами, декаподами, морскими ежами. Амплитуда микрорельефа обычно не превышает 30—50 см, однако угри, например, способны создавать и более глубокие воронки — до 1 м при диаметре до 3 м. В тропических и субтропических морях дно в прибрежной зоне обычно сплошь перерыто креветкой, выбросы которой имеют форму конусов высотой от 6 до 50 см. А в Баренцевом море в мелководных заливах с илисто-песчаным грунтом микрорельеф дна определяется конусами выбросов морских червей с высотой 15 см и диаметром основания 20 см. Невольно возникает аналогия с сусликогенным рельефом полупустынь (правда, поверхность морского дна более сглажена воздействием волн). Характер геологической деятельности II рода, осуществляемой живым веществом, оказывается принципиально сходным в море и на суше.
В. И. Вернадский, как мы упоминали в начале главы, подразделял геологическую деятельность живого вещества по характеру протекающих процессов. Несколько иначе подошел к этому вопросу современник Владимира Ивановича Н. И. Андрусов. Он писал, что химическая деятельность организмов вообще, имеющая геологическое значение, может быть сведена к двум категориям: во-первых, к образованию на наружной поверхности или внутри их твердых выделений, способных сохраняться; во-вторых, к образованию жидких и газообразных выделений, способных вступать в различные химические реакции с окружающим неорганическим миром. По существу, эту же мысль развивала на современном материале ленинградский микробиолог, доктор биологических наук Т. В. Аристовская. Она указала, что миграция атомов химических элементов может быть как прямым, так и косвенным результатом жизнедеятельности организмов (в первую очередь бактерий). В табл. 4 совмещены классификационные подходы Вернадского (горизонтальные ряды) и Андрусова — Аристовской (вертикальные столбцы).
Таблица 4
Характер и локализация процессов, осуществляемых живым веществом
Работа живого вещества в биосфере, по Вернадскому, может проявляться в двух основных формах: а) химической (биохимической) — I род геологической деятельности; б) механической — II род такой деятельности.
Геологическая деятельность I рода — построение тела организмов и переваривание пищи — конечно, является более значимой. Классическим стало функциональное определение жизни, данное Фридрихом Энгельсом: «Жизнь есть способ существования белковых тел, существенным моментом которого является постоянный обмен веществ с окружающей их внешней природой, причем с прекращением этого обмена веществ прекращается и жизнь»[49] (курсив Энгельса. — А. Л.).
Собственно говоря, постоянный обмен веществ между живым организмом и внешней средой и обусловливает проявление большинства функций живого вещества в биосфере, которые мы рассмотрим в этой главе книги. По подсчетам специалистов, в течение жизни человека через его тело проходит 75 т воды, 17 т углеводов, 2,5 т белков, 1,3 т жиров. Между тем по геохимическому эффекту своей геологической деятельности I рода человек отнюдь не самый важный вид разнородного живого вещества биосферы. Наиболее эффективную деятельность в этом отношении осуществляют на суше — грунтоеды (главным образом дождевые черви), а в океане — илоеды и фильтраторы.
Первым обратил внимание на геологическую деятельность дождевых червей Чарлз Дарвин. За год до своей смерти, в 1881 г., он выпустил фундаментальный труд «Образование растительного слоя деятельностью дождевых червей» (столетие со дня его выхода в свет было отмечено проведением Международного симпозиума по экологии дождевых червей). Дарвин убедительно доказал, что весь слой почвы непрерывно перерабатывается дождевыми червями. По его подсчетам, слой экскрементов, выделяемых дождевыми червями на плодородных почвах Англии, составляет около 5 мм в год. Отсюда следует, что почвенный пласт мощностью в 1 м дождевые черви полностью пропускают через свой кишечник за 200 лет!
Кажется, что дождевые черви созданы самой природой специально для пищеварения: они способны переваривать почти любые органические вещества, включая и новинки человеческой технологии. При этом дождевые черви заглатывают многие элементы в количествах, значительно превышающих их потребности, а избытки выделяют в окружающую среду. В результате почва, в которой находится достаточное количество червей, содержит вдвое больше магния, впятеро — азота, в 11 раз — калия, чем лишенная червей. Можно себе представить, каких размеров достигает суммарный геохимический эффект деятельности дождевых червей, если при этом учесть, что по произведенным в США расчетам биомасса червей там в 10 раз превышает массу человеческого населения!
В донных отложениях современных водоемов фекальные комочки беспозвоночных распространены также очень широко и нередко являются основной частью осадка. В Южной Атлантике, например, современные осадки илов почти нацело слагаются фекалиями фильтрующих планктонных ракообразных, а по берегам Северного моря донные осадки, образованные фекалиями мидий (также — фильтраторов), имеют мощность до 8 м. По своей «пропускной способности» с дождевыми червями в океане могут конкурировать их близкие родственники, представители того же типа кольчатых червей — полихеты. Как и в случае дождевых червей, химический состав субстрата при этом существенно изменяется: по сравнению с исходными илами он обогащается кальцием, железом, магнием, калием и фосфором.
Таким образом, пищеварительная деятельность грунтоедов на суше и илоедов и фильтраторов в океане является важным геохимическим фактором биосферы. По мнению ученых Ростовского университета, настало время говорить о рождении новой отрасли геохимии — трофической геохимии. Ее основной проблемой является исследование процессов химического преобразования субстрата при его прохождении через пищеварительный тракт массовых видов грунтоедов, илоедов и фильтраторов.
Ископаемые остатки фекалий — их называют копролитами — известны в геологических отложениях начиная с ордовика. Бесспорно, однако, что большая часть копролитов при геологических описаниях не учитывается. Происходит это из-за слабой изученности вопроса и из-за отсутствия четких диагностических признаков для определения копролитов. Чаще фиксируются в древних осадках следы проедания илоедов — бесспорные свидетельства существования в геологическом прошлом I рода геологической деятельности живого вещества.
Механическая работа живого вещества — II род геологической деятельности — проявляется как в наземных, так и в водных экосистемах.
Интересно, что первая научная работа В. И. Вернадского, написанная еще в студенческие годы, была посвящена именно механической деятельности живого вещества. Она была составлена В. И. Вернадским по материалам почвенной экспедиции 1884 г. в Екатеринославскую губернию и опубликована В. В. Докучаевым без ведома автора в 1889 г.[50] (Вернадский в это время находился в заграничной научной командировке). В этой работе В. И. Вернадский, едва перешагнувший за рубеж своего двадцатилетия, впервые в мировой литературе попытался количественно оценить влияние роющей деятельности сурка. Для самого Вернадского эта работа, видимо, послужила той своеобразной затравкой, из которой через 40 с лишним лет окончательно выкристаллизовалась стройная концепция биосферы.
Роющая деятельность в наземных экосистемах осуществляется главным образом млекопитающими и распространена повсеместно: в лесах, на лугах, в тундрах, степях, полупустынях и пустынях. При этом наибольшее развитие роющая деятельность животных получает в открытых ландшафтах аридной зоны из-за отсутствия там естественных убежищ. Мелкие млекопитающие этой зоны — сурки, суслики и слепыши — буквально перелопачивают почвы и подстилающие их отложения. Благодаря их выбросам в верхние слои почвы попадают первичные невыветрившиеся минералы, которые, разлагаясь, вовлекаются в биогеохимический круговорот. Попутно эти животные иногда оказывают ценные услуги геологам. Вспоминается следующий факт, описанный в научной литературе.
В годы Великой Отечественной войны возникла острая нехватка писчего мела — все его месторождения, расположенные на Украине и в Белгородской области, были захвачены врагом. Новое месторождение, пригодное для эксплуатации, в кратчайший срок было найдено в Казахстане по выбросам из нор сурков. Может быть, заслуга сурков и меньше, чем гусей (которые, как известно, «Рим спасли»), но добрым словом их помянуть стоит.
Как показали исследования Бориса Даниловича Абатурова, в экосистемах полупустыни роющая деятельность животных представляет собой одну из важнейших форм механической работы, ведущей к глубокому преобразованию почв и даже рельефа страны. Норы млекопитающих обеспечивают более интенсивное увлажнение почв атмосферными осадками и поступление воды в более глубокие ее горизонты. Роющая деятельность, продолжаясь в течение тысячелетий, стала важнейшим фактором формирования характерного для полупустынь микрорельефа «сусликогенного» облика, состоящего из микроповышений и западин.
В пустынях в результате совокупной деятельности ветра, растений (шишконосной эфедры), паукообразных, насекомых и мелких млекопитающих (полуденной песчанки) возникают целые «крепостные» сооружения высотой до 5 м. В увлекательной книге А. В. Смирнова «Мир растений» (1982) так описывается процесс их возникновения на примере Каракумов.
Все начинается с того, что вокруг кустиков эфедры ветер надувает холмики песка. Опавшие ветви эфедры цементируются песком, в котором много гипса. Образуется песчаный бугор с прочным куполом, в котором сохраняется постоянная температура. В нем поселяются песчанки, паукообразные, насекомые — обычные обитатели пустыни. Песчанки проделывают в бугре множество ходов, зачастую перегрызая корневища эфедры. Куст местами усыхает, но разрастается дальше своими корневищами. Каждый новый куст собирает вокруг себя песок, и история повторяется сначала. Так в пустыне появляются песчаные города с крепостными стенами (остатками первичного бугра) и башнями (вторичными дочерними буграми).
В тропиках подобными своеобразными памятниками механической деятельности живого вещества являются термитники. Их высота достигает 15 м! Если бы люди умели строить сооружения в таком же соотношении к размерам своего тела, как термиты, то Землю бы украшали небоскребы высотой до 3400 м. Исходный материал для своего строительства термиты, к вящей радости геологов, извлекают с глубины до 12 м (суркам до них далеко!). При этом, располагая термитники на линиях тектонических нарушений и в зонах повышенной трещиноватости пород, они оказывают геологам дополнительные услуги, помогая тем самым в составлении крупномасштабных структурногеоморфологических карт.
В лесных экосистемах важную механическую деятельность в виде строительства плотин осуществляют бобры. Длина некоторых бобровых плотин достигает 100 м, а рекорд, зафиксированный на Джефферсон-Ривер (штат Монтана, США), составляет 700 м! Течение рек из-за плотин замедляется; иногда образуется даже целая цепь прудов со слабо проточной водой и широкие выровненные поймы. В запрудах постепенно оседает ил. Если это происходит достаточно долго, то его отложения достигают большой мощности. При перемещении бобровых запруд или их исчезновении плодородные илы обнажаются, покрываясь густой растительностью. Таким путем в Северной Америке образовались богатые пастбища и луга, которые называют «бобровыми». Подобно человеку, но без использования техники, бобры преобразовали облик целой страны. Не зря индейцы считали бобра своим младшим братом, а у американцев существует пословица: «трудиться, как бобр».
Термитники в тропиках и «крепостные» сооружения в пустынях, сусликогенный рельеф полупустынь и бобровые луга лесной зоны — примеры грандиозной механической деятельности живого вещества в наземных экосистемах. С неменьшим успехом эта деятельность осуществляется живым веществом и в морских экосистемах.
На дне моря, как и на суше, организмы роют себе укрытия, причем не только в мягком, но и в скальном грунте. Олигохеты и полихеты углубляются в грунт на 40 см и более. Двустворчатые моллюски зарываются обычно неглубоко, но некоторые из них, например миа, роют норы, которым позавидует и сурок: они достигают глубины нескольких метров. В зоне прибоя и на перемываемом волнами песке — вот беда! — норы не выроешь и гнездо не совьешь. Приходится сверлить скальные породы. И сверлят. Сверлят водоросли и губки, бактерии и моллюски, цианобактерии, полихеты, морские ежи, рачки, мшанки… При этом, если сверление производится механически, образуются мельчайшие «опилки» диаметром в десятки микронов. Только сверлящие губки, живущие на 1 м² морского дна, выделяют за год 250 г таких «опилок».
Сверлильщики появились в далеком геологическом прошлом. Источенные ими породы находят даже в докембрийских отложениях, и поныне они продолжают свою разрушительную работу. Сверлящая деятельность моллюсков фолад вызывает иногда катастрофические последствия. Когда недавно в районе Сочи в результате непродуманного строительства берег лишился гальки, он начал отступать со скоростью до 4 м в год. Главным виновником разрушения были фолады, которые заселили каждый метр скального берега, сложенного сланцами, и принялись дружно сверлить себе подводные норки. К счастью, был найден выход: берег стали укреплять поперечными стенками, а между ними засыпать гальку. В результате сверлильщики были уничтожены: движущаяся под ударами волн галька перемолола их. А в Западной Европе не менее опасную деятельность производит случайно завезенный из Китая мохнаторукий краб — он проник во многие реки и, строя свои норы, подрывает берега и разрушает плотины.
Механическая деятельность живого вещества в значительной мере определяет микрорельеф морского дна. Положительные формы рельефа создаются активными фильтраторами и морской растительностью, скрепляющей отложения, а отрицательные — теми бентосными организмами, которые активно взмучивают ил: бентосными рыбами, декаподами, морскими ежами. Амплитуда микрорельефа обычно не превышает 30—50 см, однако угри, например, способны создавать и более глубокие воронки — до 1 м при диаметре до 3 м. В тропических и субтропических морях дно в прибрежной зоне обычно сплошь перерыто креветкой, выбросы которой имеют форму конусов высотой от 6 до 50 см. А в Баренцевом море в мелководных заливах с илисто-песчаным грунтом микрорельеф дна определяется конусами выбросов морских червей с высотой 15 см и диаметром основания 20 см. Невольно возникает аналогия с сусликогенным рельефом полупустынь (правда, поверхность морского дна более сглажена воздействием волн). Характер геологической деятельности II рода, осуществляемой живым веществом, оказывается принципиально сходным в море и на суше.
В. И. Вернадский, как мы упоминали в начале главы, подразделял геологическую деятельность живого вещества по характеру протекающих процессов. Несколько иначе подошел к этому вопросу современник Владимира Ивановича Н. И. Андрусов. Он писал, что химическая деятельность организмов вообще, имеющая геологическое значение, может быть сведена к двум категориям: во-первых, к образованию на наружной поверхности или внутри их твердых выделений, способных сохраняться; во-вторых, к образованию жидких и газообразных выделений, способных вступать в различные химические реакции с окружающим неорганическим миром. По существу, эту же мысль развивала на современном материале ленинградский микробиолог, доктор биологических наук Т. В. Аристовская. Она указала, что миграция атомов химических элементов может быть как прямым, так и косвенным результатом жизнедеятельности организмов (в первую очередь бактерий). В табл. 4 совмещены классификационные подходы Вернадского (горизонтальные ряды) и Андрусова — Аристовской (вертикальные столбцы).
Таблица 4
Характер и локализация процессов, осуществляемых живым веществом
| Род геологической деятельности | Характер процесса \ Где протекает процесс | Внутри организма | Вне организма |
|---|---|---|---|
| I | Химический (биохимический) | Переваривание пищи, построение тела организма | Выделение во внешнюю среду продуктов метаболизма и экскретов; внеклеточное пищеварение |
| II | Механический | Пропускание через желудочный тракт грунтоедов и илоедов неорганических компонентов пищи, не подверженных перевариванию | Перемещение неживого вещества организмами в ходе жизнедеятельности |
| № п/п | Функции | Краткая характеристика происходящих процессов |
|---|---|---|
| 1 | Энергетическая | Поглощение солнечной энергии при фотосинтезе, а химической энергии — путем разложения энергонасыщенных веществ; передача энергии по пищевой цепи разнородного живого вещества |
| 2 | Концентрационная | Избирательное накопление в ходе жизнедеятельности определенных видов вещества: а) используемых для построения тела организма; б) удаляемых из него при метаболизме |
| 3 | Деструктивная | 1) Минерализация необиогенного органического вещества; 2) разложение неживого неорганического вещества; 3) вовлечение образовавшихся веществ в биотический круговорот |
| 4 | Средообразующая | Преобразование физико-химических параметров среды (главным образом за счет необиогенного вещества) |
| 5 | Транспортная | Перенос вещества против силы тяжести и в горизонтальном направлении |
 Рис. 5. Минералы, образуемые живыми организмами (по Ловенштаму, 1984). Наверху — названия царств органического мира (по Маргелис, 1983), внизу — названия типов
Рис. 5. Минералы, образуемые живыми организмами (по Ловенштаму, 1984). Наверху — названия царств органического мира (по Маргелис, 1983), внизу — названия типов
По степени концентрации химических элементов Вернадский разбил живые организмы на 4 группы. В первую группу — «организмы какого-либо элемента» — были включены организмы, концентрирующие данный элемент в количестве 10% и выше. Существуют, например, кремниевые организмы (диатомовые водоросли, радиолярии, кремниевые губки), кальциевые (бактерии, водоросли, простейшие, моллюски, брахиоподы, иглокожие, мшанки и кораллы), железные (железобактерии) и т. д. Во вторую группу — «богатые каким-либо элементом» — относились организмы, содержащие данный элемент в количестве около 1% и выше (до 10%). При этом содержание элементов в первых двух группах должно быть выше, чем кларк данного элемента. Третью группу составляют «обычные организмы», четвертую — «бедные данным элементом». Развивая эти представления с несколько иных позиций, югославский геохимик В. Омальев недавно ввел понятие биогеохимического фона, которое он предложил обозначать термином «вернадский» (по аналогии с кларком). Биогеохимический фон, или вернадский, — это среднее содержание какого-либо элемента в живом веществе — как в разнородном живом веществе биосферы в целом, так и в живом веществе отдельных типов, классов, родов или видов живых организмов. Один из основоположников геохимии, известный норвежский ученый Виктор Мориц Гольдшмидт (1888—1947) в разработанной им геохимической классификации элементов выделил особую группу биофильных элементов, включив туда углерод, водород, кислород, азот, фосфор, серу, хлор и йод. Следуя по этому пути, академик Б. Б. Полынов в 1948 г. предложил выделять группу элементов-органогенов, подразделяя ее на: а) абсолютные органогены, без которых совершенно невозможно существование организмов (водород, углерод, кислород, азот, фосфор, сера, калий, магний) и б) специальные органогены, необходимые для многих организмов, но необязательные для всех. Через 8 лет В. А. Ковда добавил в число абсолютных органогенов еще 6 элементов: йод, бор, кальций, железо, медь и кобальт. В дальнейшем число органогенов неудержимо росло, и в настоящее время установлено, что, если учитывать и те элементы, которые содержатся в небольших количествах, в состав живого вещества входят все элементы таблицы Менделеева. При этом, как установили Г. Н. Саенко, М. Д. Корякова, В. Ф. Макиенко и И. Г. Добромыслова, организмы концентрируют из среды не один какой-либо элемент, а целую группу их, обычно состоящую из 4—7 поливалентных элементов. Это явление получило название специфического группового концентрирования. Интенсивность вовлечения химического элемента в биотический круговорот академик Б. Б. Полынов предложил измерять частным от деления числа, показывающего количество элемента в золе организма, на число, характеризующее его содержание в исходной породе. Позднее ученик Б. Б. Полынова профессор А. И. Перельман стал называть эту величину «коэффициентом биологического поглощения». В целом для биосферы говорят о биофильности элементов: отношении их среднего содержания в живом веществе к кларку данного элемента в литосфере. Наибольшей биофильностью характеризуется углерод, менее биофильны азот и водород. Таблица 6 Сопоставление данных по разведанным запасам некоторых химических элементов и их ежегодному накоплению фотоавтотрофами (по Бойченко и др., 1968)
| Элемент | Концентрируется ежегодно при фотосинтезе, т | Мировые запасы сырья, т |
|---|---|---|
| Углерод | 1011 | 1012 |
| Фосфор | 109 | 1010 |
| Хром | 105 | 108 |
| Марганец | 107 | 108 |
| Железо | 108 | 1011 |
| Кобальт | 105 | 106 |
| Никель | 106 | 107 |
| Медь | 107 | 108 |
| Цинк | 107 | 107 |
| Молибден | 105 | 106 |
Глава пятая. Три фактора: био, эко и тафо
Окончательное количество и качество естественных продуктов зависит не только от условий образования, но и в большой степени от условий сохраненияИоганнес Вальтер, 1895
 Формирование палеобиогенного вещества в земной коре регулируется тремя факторами. Эти факторы следующие:
первый — продуктивность живого вещества, которое служит исходным материалом при формировании палеобиогенного вещества: биотический (биофактор);
второй — условия, благоприятные для концентрации необиогенного вещества: экологический (экофактор);
третий — обстановка, обеспечивающая переход биогенного вещества в ископаемое состояние: тафономический (тафофактор).
В третьей главе мы рассмотрели, как распределено живое вещество в биосфере. Однако всегда ли высокая концентрация живого вещества и значительная его продуктивность — биофактор — приводят к увеличению количества биогенного вещества в осадках? В некоторых случаях такая связь фиксируется совершенно четко: например, в океане, по свидетельству А. П. Лисицына и М. Е. Виноградова (1982), «максимальное количество биогенного материала накапливается в донных осадках там, где выше первичная продукция». А в озерах, наоборот, установлено, что озера с низким уровнем биологической продуктивности образуют илы с высоким содержанием необиогенного органического вещества. Видимо, существуют и какие-то другие факторы, помимо «био», которые регулируют накопление в осадках биогенного вещества.
С действием этих факторов ученые впервые столкнулись в середине прошлого века, когда протягивали через Атлантику «телеграфический канат» (слова «кабель» еще не существовало). Начиналось целенаправленное исследование океанов, рождалась новая наука — океанология. Впервые были изучены осадки океанских глубин. Их состав оказался неожиданным.
«Отчего же не найдено рыбьих костей, зубов, чешуй, раковин, морских звезд, кораллов и других частей животных, не подверженных скорому тлению?» — писал научный обозреватель того времени. Объяснение было найдено правильное: «Когда эти жители вод, близких к поверхности, умирают, трупы их делаются добычею животных низшей организации, которые, в свою очередь, доходят тем же путем до простых организмов… Еще задолго до того времени, когда остатки животных, обитающих на поверхности моря, дойдут до дна и ассимилируются с его массою, они, может быть, проходят по разным инстанциям и оканчивают это странствование у микроскопических животных, населяющих самые глубокие части моря».
Попытаемся представить себе на минуту, что на Земле существует исключительно автотрофное живое вещество. Тогда накопление необиогенного вещества регулировалось бы только его продуктивностью (другое дело, что жизнь исчерпала бы себя за несколько сотен или тысяч лет). Но на реальной Земле, как известно, существует и гетеротрофное живое вещество. В большинстве случаев сапротрофы при мощной поддержке абиотических факторов с честью справляются со своей трудной задачей — разложить необиогенное вещество на простые составные части, пригодные для повторного использования живым веществом: углекислый газ, воду, сероводород, аммиак и т. д.
Иногда, однако, полного разложения необиогенного вещества все же не происходит. Деятельность сапротрофов может быть подавлена как сухостью климата (в пустыне), так и чрезмерной увлажненностью обстановки (в болоте), низкой температурой (в тундре) или токсичностью среды. В воде сфагновых болот, например, имеются фенолы, обусловливающие ее стерильность. И не зря в средние века моряки, отправляясь в дальние плавания, запасались именно такой водой. Неоценимую помощь оказывает она и археологам, консервируя на века деревянные остатки человеческой культуры: остатки свай, настилы мостовых, лодки, весла, мелкие поделки.
Полнота разложения необиогенного вещества в значительной мере зависит и от длительности протекания процессов деструкции. Так, в озерах и неглубоких частях океана, где планктонную пленку от донной отделяют лишь немногие метры водной толщи, количество попавшего в осадок детрита будет, конечно, значительно больше, чем в глубоководных морских бассейнах. Сейчас подсчитано, что из всей необиогенной органики в глубоководной части океана накапливается только 2,7%, на шельфах и континентальном склоне — 26,3%, а на заболоченных участках суши и во внутриконтинентальных мелководных водоемах — 71%.
Московский ученый Дмитрий Викторович Панфилов, труды которого мы уже неоднократно цитировали, выступая на 23‑м Международном географическом конгрессе в Москве (1976), впервые предложил выделить три основных типа экосистем: транзитный, автономный и аккумулятивный. Транзитные экосистемы характеризуются постоянным привносом и выносом элементов минерального питания растений, автономные — слабым привносом и выносом, а аккумулятивные — значительным привносом и слабым выносом вещества (рис. 6). В качестве примеров аккумулятивных систем Д. В. Панфилов привел океанические абиссали, мангры, болота и др. Накапливается же здесь не что иное, как необиогенное вещество — органическое (в манграх и болотах) или неорганическое (океанические абиссали). Дальнейшее развитие и уточнение этой концепции было произведено Д. В. Панфиловым в монографии «Классификация, география и антропогенная трансформация экосистем» (1980), написанной совместно с Ю. А. Исаковым и Н. С. Казанской.
Формирование палеобиогенного вещества в земной коре регулируется тремя факторами. Эти факторы следующие:
первый — продуктивность живого вещества, которое служит исходным материалом при формировании палеобиогенного вещества: биотический (биофактор);
второй — условия, благоприятные для концентрации необиогенного вещества: экологический (экофактор);
третий — обстановка, обеспечивающая переход биогенного вещества в ископаемое состояние: тафономический (тафофактор).
В третьей главе мы рассмотрели, как распределено живое вещество в биосфере. Однако всегда ли высокая концентрация живого вещества и значительная его продуктивность — биофактор — приводят к увеличению количества биогенного вещества в осадках? В некоторых случаях такая связь фиксируется совершенно четко: например, в океане, по свидетельству А. П. Лисицына и М. Е. Виноградова (1982), «максимальное количество биогенного материала накапливается в донных осадках там, где выше первичная продукция». А в озерах, наоборот, установлено, что озера с низким уровнем биологической продуктивности образуют илы с высоким содержанием необиогенного органического вещества. Видимо, существуют и какие-то другие факторы, помимо «био», которые регулируют накопление в осадках биогенного вещества.
С действием этих факторов ученые впервые столкнулись в середине прошлого века, когда протягивали через Атлантику «телеграфический канат» (слова «кабель» еще не существовало). Начиналось целенаправленное исследование океанов, рождалась новая наука — океанология. Впервые были изучены осадки океанских глубин. Их состав оказался неожиданным.
«Отчего же не найдено рыбьих костей, зубов, чешуй, раковин, морских звезд, кораллов и других частей животных, не подверженных скорому тлению?» — писал научный обозреватель того времени. Объяснение было найдено правильное: «Когда эти жители вод, близких к поверхности, умирают, трупы их делаются добычею животных низшей организации, которые, в свою очередь, доходят тем же путем до простых организмов… Еще задолго до того времени, когда остатки животных, обитающих на поверхности моря, дойдут до дна и ассимилируются с его массою, они, может быть, проходят по разным инстанциям и оканчивают это странствование у микроскопических животных, населяющих самые глубокие части моря».
Попытаемся представить себе на минуту, что на Земле существует исключительно автотрофное живое вещество. Тогда накопление необиогенного вещества регулировалось бы только его продуктивностью (другое дело, что жизнь исчерпала бы себя за несколько сотен или тысяч лет). Но на реальной Земле, как известно, существует и гетеротрофное живое вещество. В большинстве случаев сапротрофы при мощной поддержке абиотических факторов с честью справляются со своей трудной задачей — разложить необиогенное вещество на простые составные части, пригодные для повторного использования живым веществом: углекислый газ, воду, сероводород, аммиак и т. д.
Иногда, однако, полного разложения необиогенного вещества все же не происходит. Деятельность сапротрофов может быть подавлена как сухостью климата (в пустыне), так и чрезмерной увлажненностью обстановки (в болоте), низкой температурой (в тундре) или токсичностью среды. В воде сфагновых болот, например, имеются фенолы, обусловливающие ее стерильность. И не зря в средние века моряки, отправляясь в дальние плавания, запасались именно такой водой. Неоценимую помощь оказывает она и археологам, консервируя на века деревянные остатки человеческой культуры: остатки свай, настилы мостовых, лодки, весла, мелкие поделки.
Полнота разложения необиогенного вещества в значительной мере зависит и от длительности протекания процессов деструкции. Так, в озерах и неглубоких частях океана, где планктонную пленку от донной отделяют лишь немногие метры водной толщи, количество попавшего в осадок детрита будет, конечно, значительно больше, чем в глубоководных морских бассейнах. Сейчас подсчитано, что из всей необиогенной органики в глубоководной части океана накапливается только 2,7%, на шельфах и континентальном склоне — 26,3%, а на заболоченных участках суши и во внутриконтинентальных мелководных водоемах — 71%.
Московский ученый Дмитрий Викторович Панфилов, труды которого мы уже неоднократно цитировали, выступая на 23‑м Международном географическом конгрессе в Москве (1976), впервые предложил выделить три основных типа экосистем: транзитный, автономный и аккумулятивный. Транзитные экосистемы характеризуются постоянным привносом и выносом элементов минерального питания растений, автономные — слабым привносом и выносом, а аккумулятивные — значительным привносом и слабым выносом вещества (рис. 6). В качестве примеров аккумулятивных систем Д. В. Панфилов привел океанические абиссали, мангры, болота и др. Накапливается же здесь не что иное, как необиогенное вещество — органическое (в манграх и болотах) или неорганическое (океанические абиссали). Дальнейшее развитие и уточнение этой концепции было произведено Д. В. Панфиловым в монографии «Классификация, география и антропогенная трансформация экосистем» (1980), написанной совместно с Ю. А. Исаковым и Н. С. Казанской.
 Рис. 6. Типы экосистем (по Панфилову, 1976): 1 — транзитный тип; 2 — автономный; 3 — аккумулятивный. Стрелками обозначен поток питательных веществ
Рис. 6. Типы экосистем (по Панфилову, 1976): 1 — транзитный тип; 2 — автономный; 3 — аккумулятивный. Стрелками обозначен поток питательных веществ
Фактор, обеспечивающий концентрацию необиогенного вещества в экосистемах, недавно было предложено называть экофактором[57]. Первым, кто подчеркнул важности этого фактора при накоплении каустобиолитов, был известный советский микробиолог Владимир Оттонович Таусон (1894—1946), автор блестяще написанных научно-популярных книг «Наследство микробов» (1947) и «Великие дела маленьких существ» (1948). К компетенции экофактора можно отнести и условия, исключающие привнос в экосистему значительных количеств терригенного материала, разбавляющего необиогенное вещество: выровненный рельеф окружающей территории и слабые колебательные движения земной коры. Те участки биосферы, в которых происходит формирование биогенных или хемогенных осадков, не нарушаемое привносом терригенного материала, называют бассейнами концентрации. К ним, по представлениям академика Дмитрия Васильевича Наливкина (1889—1982), относятся бассейны, где накапливаются фосфатные, марганцевые, железистые и другие осадки; бассейнами концентрации являются и торфяные болота. Все приведенные выше рассуждения относятся к той начальной стадии формирования осадочных пород, которая протекает в биосфере. В масштабе геологического времени консервация необиогенного вещества в аккумулятивных экосистемах — явление сугубо временное, нечто вроде хранения багажа в автоматической камере на вокзале. Нормально жизнь забирает обратно свой багаж (то бишь необиогенное вещество). Нужны какие-то исключительные обстоятельства, чтобы ячейка камеры хранения осталась невскрытой, а багаж — замурованным в ней. При аккумуляции осадков таким чрезвычайным обстоятельством является процесс захоронения. «Основным содержанием процесса образования любого местонахождения (органических остатков. — А. Л.) нужно считать превращение органических остатков из компонентов биосферы в компоненты литосферы, — писал И. А. Ефремов[58]. — Продукты биосферы, переходя в литосферу, становятся членами минеральных ассоциаций осадочных пород и в качестве таковых подчиняются всем закономерностям и дальнейшим судьбам процессов литосферы. Самый переход остатков биосферы в литосферу мы и называем захоронением». Только так. Только будучи удаленным из биосферы, т. е. став изолированным от процессов активного разложения, необиогенное вещество получает шанс на сохранение в течение миллионов лет геологической истории. Только перестав быть осадком и превратившись в горную породу. Классик геологии немецкий ученый Иоганнес Вальтер, слова которого мы цитировали в эпиграфе к этой главе, лаконично сформулировал следующий тезис: «Отложениями мы называем все образующиеся скопления вещества — породами делаются лишь те, которые сохранились». Переход биогенного вещества в ископаемое состояние, его трансформацию из необиогенного вещества в палеобиогенное можно назвать тафофактором формирования осадочных пород (от греческого слова «тафос» — могила). Применительно к палеонтологии учение о захоронении органических остатков в земной коре (тафономию) создал Иван Антонович Ефремов (1907—1972) — выдающийся палеонтолог и писатель, известный миллионам читателей как автор романов «Туманность Андромеды», «Лезвие бритвы» и других произведений. Наиболее общим случаем захоронения необиогенного вещества на суше является погружение бассейна его аккумуляции. Однако захоронение может быть и внезапным. На памяти человечества происходили случаи катастрофических захоронений целых городов. Общеизвестен случай с Помпеей, засыпанной слоем вулканического пепла толщиной 4 м. А вот другой пример, менее известный: 31 мая 1970 г. город Юнгай в горах Перу в течение нескольких секунд был затоплен десятиметровым (!) слоем грязи. Такого же рода внезапные захоронения могли происходить и в геологическом прошлом. Не столь уж редки, например, находки «ископаемых лесов» — скоплений минерализованных древесных стволов, захороненных на месте своего произрастания. А ведь если дерево не успело сгнить до того, как его перекрыло осадками, значит, процесс его захоронения происходил очень быстро. В океанах захоронение малоустойчивого биогенного вещества также может происходить в результате очень интенсивного осадконакопления и изоляции тем самым очередных порций осадка от агрессивного воздействия морских вод и деятельности донной фауны. Общая обстановка на суше не способствует биогенному осадкообразованию. И. А. Ефремов предложил называть ультрафациями те обстановки, которые благоприятствуют захоронению биогенного вещества. От материка же, как отмечает Ефремов, в геологической летописи остается пустота, окруженная поясом ультрафаций. Полную противоположность континенту в этом отношении представляет океан, точнее, его донная пленка. Уже упоминалось, что донная пленка жизни имеет исключительное значение в формировании осадочных пород, поскольку здесь существуют условия для сохранения биогенного вещества в течение длительного геологического времени. «По мере падения сверху остатков жизни и взмученных частей косной материи — нижние слои морской грязи становятся безжизненными, и образованные жизнью химические тела не успевают перейти в газообразные продукты или войти в новые живые вещества. Живой слой грязи никогда не превышает немногих метров, между тем как он непрерывно растет с поверхности. Снизу он неустанно замирает»[59]. Именно здесь звуча последние аккорды великой симфонии жизни, а необиогенное вещество, выходя из биосферы, превращается с ходом геологического времени в палеобиогенное. Количество биогенного вещества, переходящего в ископаемое состояние, очень невелико по отношению к годичной продукции живого вещества. У углерода, например, оно составляет для биосферы в целом 0,05%, для Мирового океана — 0,4% (0,7% для подводных окраин и 0,1% для пелагиали). Такого рода ничтожные доли вещества, ускользающие из биотического круговорота благодаря переходу в ископаемое состояние, слагают биогенное вещество метабиосферы. Формирование осадочных пород Владимир Иванович изобразил в виде схемы (рис. 7). Из нее следует, что в 20—30‑е годы Вернадский представлял себе исключительно подводное формирование осадочных пород. В 40‑е годы, однако, Л. С. Берг и Б. Л. Личков обосновали мнение, что осадочные породы могут формироваться и на поверхности материков, в ходе процессов почвообразования (рис. 8). Владимир Иванович эту идею поддержал: статью Б. Л. Личкова на эту тему он представил для опубликования в «Известия АН СССР» и сочувственно упомянул о его работах в «Химическом строении биосферы Земли и ее окружения» (1965, с. 128).
 Рис. 7. Принципиальная схема формирования осадочных пород (по Вернадскому, 1936)
Рис. 7. Принципиальная схема формирования осадочных пород (по Вернадскому, 1936)
 Рис. 8. Принципиальная схема формирования осадочных пород (по Личкову, 1946)
Рис. 8. Принципиальная схема формирования осадочных пород (по Личкову, 1946)
Накопление осадков и их дальнейшие превращения происходят в динамических системах особого рода, которые В. И. Вернадский назвал биокосными естественными телами. Это понятие В. И. Вернадский ввел в 1938 г.[60], выделив в биосфере, помимо живых и косных тел, системы, где осуществляется их взаимодействие. В качестве примеров биокосных естественных тел Владимир Иванович приводил почвы, леса, кору выветривания, илы, поверхностные воды, океан, биосферу в целом. «Организованные биокосные тела занимают значительную часть повесу и по объему биосферы, — пишет В. И. Вернадский[61]. — Их остатки, после гибели организмов, их составляющих, образуют биогенные породы, которые образуют огромную часть стратисферы». Формируются ли осадочные породы в результате преобразования донных осадков водоемов или почв — в обоих случаях их образование происходит в биокосных естественных телах (или по А. И. Перельману, — биокосных системах). Какую же роль выполняет при этом живое вещество и какие процессы происходят в биокосных системах? Обратимся к рис. 9 и 10. Отличительная особенность биокосных систем — присутствие в них живого вещества. Если входящие в состав системы живые организмы погибнут, то система потеряет право считаться биокосной. Содержание живого вещества во всех биокосных системах является небольшим (по отношению к общей массе), но именно живому веществу принадлежит ведущая роль в их функционировании.
 Рис. 9. Схема формирования осадочных пород в биокосных системах с последующим захоронением
Рис. 9. Схема формирования осадочных пород в биокосных системах с последующим захоронением
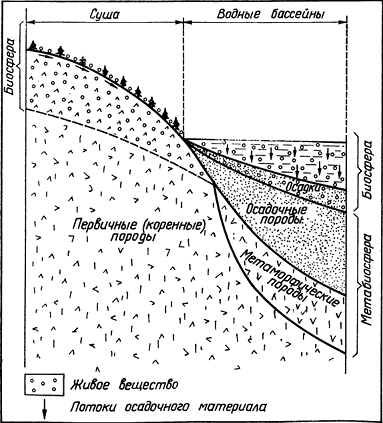 Рис. 10. Схема формирования горных пород метабиосферы
Рис. 10. Схема формирования горных пород метабиосферы
Раз в системе есть живое вещество — значит, обязательно есть и продукты его жизнедеятельности, и отмершая органика, иначе говоря, необиогенное вещество. Обязательно есть в биокосных системах и абиогенное вещество (оно может поступать дополнительно в виде терригенной примеси, продуктов вулканизма или внеземного материала — земное и внеземное вещество мы на этой схеме объединяем). Может присутствовать (но не обязательно) и палеобиогенное вещество, например, известняки, фосфориты и т. д. Живое вещество в ходе процессов жизнедеятельности непрерывно потребляет и вовлекает в биотический круговорот все новые порции абиогенного вещества (мы помним, что в этом заключается одна из основных функций живого вещества в биосфере). Усваивается большей частью абиогенное вещество земного происхождения, но видимо, используется и часть абиогенного вещества внеземного происхождения. Наряду с абиогенным живое вещество потребляет и необиогенное. Утилизируется им и палеобиогенное вещество (так, сверлильщики используют для построения своих раковин биогенные известняки). Высшие растения осваивают биогенные минералы фосфора, а некоторые виды бактерий — органическое вещество углей и нефтей; тионовые бактерии окисляют биогенные сульфиды, которые созданы трудами их предшественников — сульфатредудирующих бактерий, и т. д. Таким образом, как 40 рек впадают в Байкал, а вытекает только одна, так и живое вещество вовлекает в свой состав все типы неживого вещества, а продуцирует только один тип — необиогенное вещество. Биокосные системы творят осадочные породы Земли. После захоронения жизнь в них замирает, и биокосное естественное тело превращается в косное. В результате из четырех типов веществ, входящих в состав биокосных систем, сохраняются два типа неживого вещества косных систем: абиогенное и биогенное. Процесс формирования осадочных пород — литогенез — геологи делят на четыре стадии. Первая из них — стадия гипергенеза — является как бы подготовительной: в это время образуются исходные продукты, из которых в дальнейшем формируются осадочные породы. Во второй стадии — седиментогенеза — происходит перенос вещества и образование рыхлых несцементированных осадков: песков, глин, ракушняка, илов, торфа и т. д. Биогенная компонента представлена здесь необиогенным веществом. Третья стадия носит название диагенеза. В этот период рыхлые, зачастую сыпучие осадки преобразуются в плотные прочно сцементированные породы: песчаники, аргиллиты, известняки, ископаемые угли. Необиогенное вещество после захоронения переходит в палеобиогенное. Наконец, в четвертой стадии — ее называют катагенезом — происходит дальнейшее уплотнение и преобразование осадочных пород. Как мы увидим дальше, живое вещество играет важнейшую, а большей частью — определяющую роль на трех первых стадиях литогенеза, и лишь на стадии катагенеза его влияние сходит на нет. Не участвует живое вещество и в процессах метаморфизма, в результате которых осадочные породы переходят в метаморфические. Вместо термина «гипергенез» часто употребляется более близкий по смыслу и простой — выветривание. Появился он тогда, когда думали, что основным агентом разрушения горных пород являются чисто физические факторы, и в их числе — ветер. Такие представления господствовали в науке примерно до 40‑х годов нашего века. Вернадский писал в 1938 г.: «Процесс выветривания горных пород есть биокосный процесс, что обычно не учитывается. Мне кажется, это объясняет отставание этой области химической геологии (коры выветривания) от современного уровня знания. К нему подходят только как к процессу физико-химическому. Биогеохимический подход к решению этой проблемы должен дать много»[62]. Подход, о котором писал В. И. Вернадский, действительно, много дал для познания процессов гипергенеза. В нашей стране в 40‑е годы инициатором и организатором исследований в этом направлении явился выдающийся почвовед, геохимик и географ академик Борис Борисович Полынов (1877—1952). С геологических позиций было важно выяснить, как происходит выветривание в горных районах: именно там сильнее всего происходит денудация и именно оттуда выветрившийся материал поступает в бассейны аккумуляции. Поэтому исследования Б. Б. Полынова и его соратников проводились на скальных породах Урала, Кавказа и Тянь-Шаня. Этими работами было показано, что даже самые первые стадии выветривания скальных пород — образование мелкозема — происходят при активном воздействии живых организмов. Б. Б. Полынов сделал вывод: «Представление о том, что происходит какое-то стерильное — абиотическое — выветривание, несомненно, является совершенно нереальным, придуманным и не подтвержденным фактами. Мы, понятно, не отрицаем возможности термического дробления массивной породы, как не отрицаем возможности реакций окисления и карбонатизации, но мы категорически отрицаем возможность изолированного, стерильного проявления этих процессов, проявления их как чисто абиотических, подготовительных к захвату организмами минеральных элементов». Время подтвердило справедливость этой позиции. Сейчас показано, что наиболее активное биогенное выветривание горных пород происходит в пределах довольно тонкой поверхностной пленки, толщина которой на скальных породах обычно колеблется от долей миллиметра до 5 мм и лишь изредка превышает 1 см. Она развита во всех климатических поясах земного шара. Там, где условия существования наиболее суровые, живые организмы селятся не на поверхности, а внутри камня. Такая форма жизни получила название эндолитной: от греческих корней «эндо» (внутри) и «литос» (камень). Уход в камень позволяет живым организмам спастись от палящих солнечных лучей и несколько сгладить суточные температурные скачки, характерные для самых экстремальных местообитаний Земли — пустынь полярных и жарких стран. В Антарктиде, казалось бы, можно не опасаться солнечного удара, но знаете ли вы, что именно Антарктида — самый солнечный материк на нашей планете? Избыток солнца и очень неприятные для живых организмов колебания температуры около 0° (в летний сезон) заставляют гегемонов антарктической биоты — лишайники — прятаться под камнями. Селятся они и в трещинах, порой почти незаметных. Биомасса лишайников в Антарктиде достигает довольно внушительной величины — 175 г/м². Функционируют они, естественно, только летом, а зимой впадают в состояние анабиоза. В противоположной ситуации — в пустынях приэкваториальных стран такую же скрытную жизнь внутри камня ведут прокариоты — цианобактерии и бактерии. Они активно «всверливаются» в скалы, причем глубина их нор достигает порой 3 см. Из скальных пород пустынь Америки, Азии и Африки выделено около 150 культур цианобактерий. Несмотря на незначительную мощность поверхностной пленки биогеохимического выветривания, действие ее в зоне гипергенеза играет решающую роль: именно здесь происходят первичная дезинтеграция и минеральные преобразования скальных пород. Основными «действующими лицами» являются бактерии, цианобактерии, аэрофильные водоросли и грибы, подготавливающие субстрат для заселения лишайников и высших растений (последние воздействуют на скалы механически, разламывая их своими корнями). Среди автотрофных организмов наиболее активными являются нитрифицирующие бактерии, а среди гетеротрофов — микроскопические грибы. Из поверхностных слоев зоны гипергенеза изверженных пород канадские микробиологи выделили 31 вид микроскопических грибов. Так, при определяющем влиянии живого вещества протекают процессы гипергенеза в наземных экосистемах. В водных экосистемах гипергенез также регулируется деятельностью живого вещества, в данном случае — бентосных сверлящих организмов. «Действующие лица» здесь те же: бактерии, цианобактерии, водоросли и грибы. Правда, в море нет лишайников, да и высшие растения как будто в процессах гипергенеза не участвуют, но их с успехом заменяют многоклеточные животные — сверлильщики, осуществляющие деструктивную функцию. Седиментогенез — следующая стадия процесса литогенеза. Какую же роль играет здесь живое вещество? Еще несколько десятилетий назад, не мудрствуя лукаво, полагали, что роль живых организмов сводится к поставке в осадок своих бренных остатков, каковыми и сложены некоторые известняки, кремнистые породы, ископаемыеугли. Лишь в 70‑е годы выяснилось, что в водных экосистемах живое вещество, помимо этого, регулирует весь механизм очистки вод как от терригенной, вулканогенной и другой неорганической взвеси, так и от биогенного детрита. Процесс этот получил название биофильтрации. Мощность фильтра живого вещества огромна. Так, суммарная суточная потребность в пище фильтраторов Мирового океана составляет 10 млрд. т, а взвешенный сток всех рек мира за год — не за сутки! — равняется 18 млрд. т. В предыдущих главах мы уже упоминали об исследованиях мидиевых банок Белого моря, проведенных Кириллом Александровичем Воскресенским в 40‑е годы. Продемонстрировав поразительную способность мидий быстро и эффективно осаждать из морской воды взвешенные в ней частицы, К. А. Воскресенский сделал знаменательный вывод: «Факт вовлечения в придонную циркуляцию значительного по толщине слоя воды в соединении с активным отнятием из него взвесей населенным дном заставляет пересмотреть процессы осадочной дифференциации. Закон Стокса и его модификации, учитывающие лишь закономерности механики, физики и химии, становятся недостаточными в динамическом поле, где проявляют себя биомассы фильтраторов. Вблизи населенного дна относительно простые законы снимаются более высокими биогидрологическими». Процессы осадочной дифференциации действительно пришлось пересматривать, когда в океанологии стали применяться седиментационные ловушки — специальные приспособления, служащие для накопления вещества, поступающего в осадок. Уже первые исследования такого рода показали, что осаждение материала в море происходит не «частица за частицей» (как предполагалось умозрительно), а главным образом в виде фекальных пеллет размером от десятков микронов до 1—4 мм. Содержание таких пеллет в осадках, как правило, превышает 60%, а в некоторых случаях осадки сложены пеллетами полностью! Исследования показали, что биофильтрация и соответственно биоседиментация (от англ. sedimentation — осаждение) осуществляется в океане главным образом многоклеточными животными двух пленок жизни — планктонной и бентосной. Некоторый вклад в биоседиментацию вносит и нектон. Наглядный пример нектонных фильтраторов — киты, процеживающие морскую воду для добывания криля. Процесс биофильтрации зоопланктоном ограничен главным образом верхним пятисотметровым слоем водной толщи. Наиболее активными фильтраторами здесь являются мелкие ракообразные: веслоногие и ветвистоусые рачки, остракоды, эвфаузииды (криль), а также коловратки, простейшие и моллюски. Большинство организмов зоопланктона производит безвыборочное удаление из морской воды взвешенных частиц с минимальным размером 1—2 мкм, а некоторые организмы — даже более мелких. Благодаря механизму биофильтрации все мелкие взвешенные частицы без различия их генезиса объединяются в пеллеты, покрытые защитной оболочкой из органического вещества, и выделяются в окружающую среду. Скорость их осаждения в морской воде составляет в среднем около 100 м/сут, что в сотни раз больше, чем скорость осаждения исходных частиц взвеси. При прохождении через водную толщу пеллеты повторно многократно используются в качестве пищи более глубоководными организмами (в том числе и нектонными) и, таким образом, переупаковываются для дальнейшей транспортировки к морскому дну. Роль нектонных организмов в этих превращениях особенно велика в апвеллинговых сгущениях жизни. В огромных масштабах биофильтрацию осуществляют и организмы другой пленки жизни — бентосной. На шельфе фильтрационная система бентоса является наиболее развитой; здесь она по своей производительности местами даже превосходит фильтрационную систему зоопланктона — в целом по океану более мощную. Основными «исполнителями» здесь являются массовые виды моллюсков, а также так называемые морские желуди — усоногие рачки балянусы. Бентосные фильтраторы, как и планктонные, фильтрование производят безвыборочно, однако между ними существует и различие. Заключается оно в том, что бентосные организмы отфильтровывают более крупные частицы (вплоть до песка), которые не улавливаются зоопланктоном. При этом тонкость очистки и эффективность фильтрации остаются очень высокими (не зря моллюски уже давно используются в системах биологической очистки водной среды). Так, каждая мидия за свою жизнь прокачивает через себя 200 тыс. л воды, а все мидии Черного моря профильтровывают весь его объем за 1 год 5 месяцев. Цифра сама по себе впечатляющая; однако если мы вспомним, что весь Мировой океан профильтровывается зоопланктоном всего за полгода, то убедимся, что бентосный биофильтр по своей мощности планктонному уступает. Что касается балянусов, то они в пределах своих колоний ежегодно образуют 8—16 мм осадков, из которых на пеллеты приходится 5—13 мм, а на раковинный материал — 3 мм. Таким образом, процесс седиментогенеза в водных бассейнах (а ведь именно там происходит формирование большей части осадочных пород!) регулируется живым веществом. Живые организмы образуют своеобразный конвейер, по которому в форме пеллет передаются частицы взвеси в ходе седиментогенеза. Достигнув морского дна, пеллеты перерабатываются бентосными организмами и распадаются. Роль пеллетного транспорта, таким образом, затушевывается, и при чисто механическом подходе остается неясным, как при данном гидродинамическом режиме могли попасть в осадки столь мелкие частицы. Подтверждается давний вывод К. А. Воскресенского: «Закон Стокса и его модификации… становятся недостаточными в динамическом поле, где проявляют себя биомассы фильтраторов». Биофильтрацию осуществляют, как известно, многоклеточные животные. Роль растений в седиментогенезе значительно скромнее: они лишь поставляют исходный материал для некоторых типов горных пород (ископаемые угли) да служат механическим барьером, тормозящим потоки терригенных частиц и обусловливающим их седиментацию. В водных экосистемах роль такого барьера играют заросли водорослей и водных цветковых растений, на суше — разнообразные высшие растения. Хорошо развитый травяной покров в саваннах и прериях задерживает эоловые частицы. В результате корни растений перекрываются пылью, а верхняя часть растений отмирает. Затем появляется новая растительность, и процесс повторяется. По некоторым данным, именно таким образом накапливаются на суше мощные лёссовые толщи. Значительную роль в седиментогенезе на континентах играют и некоторые почвенные насекомые (термиты и муравьи), поставляющие на поверхность осадочный материал буквально из-под земли. Осадконакопление в этом случае — очередной парадокс! — происходит не сверху вниз, как обычно, а снизу вверх. Предполагают, что в тропической Африке термиты таким образом сформировали слой глинистых песков мощностью 4 м. Многообразна роль живого вещества в седиментогенезе, но главным, конечно, является процесс биофильтрации. Резюмируя достижения биоседиментологии, член-корреспондент АН СССР Александр Петрович Лисицын сказал: «В «живом океане» биос и связанный с ним поток пеллетов определяют процессы седиментогенеза, подготовки биогенного осадочного материала, его транспортировки в пеллетах и отложение, а также дальнейшие преобразования пеллетного материала с использованием заключенной в пеллетах органики в ходе диагенеза и катагенеза». Действительно, и дальнейшие преобразования осадков в значительной мере определяются деятельностью живого вещества. Даже после попадания в осадок странствия его частиц по кишечным трактам живых организмов не прекращаются: верхний слой осадков мощностью до 1 м интенсивно перерабатывается илоедами. В современных морских экосистемах наиболее активными илоедами являются кольчатые черви (аннелиды), отдельные виды которых способны пропустить за год до 1,5 м осадков «через кишечную тюрьму». (Заболоцкий).
 Рис. 11. Переработка донных осадков современным двустворчатым моллюском Yoldia hyperborea (по Йохансону, 1985)
Рис. 11. Переработка донных осадков современным двустворчатым моллюском Yoldia hyperborea (по Йохансону, 1985)
Процессы перекапывания и перемешивания осадков живыми организмами называют биотурбацией (рис. 11, 12). Они протекают как в морских, так и в пресноводных экосистемах. В Байкале на глубине 380 м через иллюминатор спускаемого аппарата «Пайсис» старший научный сотрудник Байкальской биологической станции В. Н. Максимов наблюдал следующую картину: «У самого дна, не выше 2—3 м от него, вверх или вниз головой парят голомянки. Время от времени они приближаются к скоплениям ила, стремительно врезаясь, взмучивают их и взмывают затем вверх в облаке поднятой ими мути. Невольно приходит на ум сравнение: голомянки — пахари дна. Фонтанчики мути периодически появляются во всем поле зрения. И это происходит веками!»
 Рис. 12. Нарушение слоистости донных осадков пескожилом. Штриховкой и другими условными обозначениями показаны осадки разного состава (по Алексееву и Найдину, 1973)
Рис. 12. Нарушение слоистости донных осадков пескожилом. Штриховкой и другими условными обозначениями показаны осадки разного состава (по Алексееву и Найдину, 1973)
В морских экосистемах «пахарями дна» служат зарывающиеся в ил черви, голотурии, морские ежи, морские звезды, моллюски, офиуры, высшие раки и некоторые рыбы (камбала), в пресноводных — насекомые, олигохеты и моллюски. Каждый из них воздействует на осадки по-своему, а их совокупная деятельность приводит к повышению аэрации отложений, увеличению их рыхлости и полному изменению текстуры. На Всесоюзном совещании «Биоседиментация в морях и океанах» в Теберде в 1983 г. настоящую сенсацию вызвал доклад сотрудника Института океанологии им. П. П. Ширшова АН СССР В. М. Купцова «Постседиментационное перемешивание донных осадков бентическими организмами». Докладчик рассказал о необычном исследовании: по радиоактивному распаду производилось определение возраста различных интервалов осадков в их верхнем 20‑сантиметровом слое. Результаты оказались неожиданными: предполагаемого увеличения возраста по мере погружения в осадок не наблюдалось! Цифры абсолютного возраста имели широкий размах варьирования со средним значением около 7 тыс. лет. В. М. Купцов сделал единственно правильный вывод: верхний слой осадка настолько интенсивно перемешан живыми организмами, что его возраст может быть определен только в целом, без подразделения на отдельные интервалы. Воздействию макрофауны на осадки в последние годы уделяется все большее внимание. Этому вопросу посвящаются специальные симпозиумы. Как пишут советские океанологи Н. А. Айбулатов и В. А. Друшиц, «построение различных моделей, выведение каких-либо расчетных зависимостей без учета влияния биологической активности возможно лишь для анаэробных областей и для осадков, лежащих ниже зоны биотурбации». Большое значение при диагенетическом преобразовании осадков имеет деятельность сапротрофных микроорганизмов. «Биологический реактор» диагенеза работает главным образом на органическом веществе, и большинство химических реакций осуществляется бактериями. Фекалии макрофауны создают для их развития особенно благоприятную среду. Раньше думали, что глубина проникновения сапротрофных микроорганизмов в толщу осадков очень невелика и ограничивается первыми сантиметрами. Сейчас показано, что бактерии распространены повсеместно по всей изученной микробиологами толще морских осадков, т. е. не менее чем на 3—6 м от поверхности дна. Существует предположение, что процессы активной жизнедеятельности бактерий могут продолжаться до глубины 200—250 м от поверхности осадка. Лимитирующими факторами для их развития служат только исчерпание запасов органического детрита и температуры, значительно превышающие 100°. Если содержание органического детрита в осадках достаточно велико, аэробные процессы в толще осадков сменяются анаэробными, если же органики недостаточно (например, в глубоководных участках океана, удаленных от континентов), зона анаэробного диагенеза в осадках не развивается. Микробиологические процессы, происходящие в донных илах, приводят к преобразованию их органического вещества, изменению физико-химических параметров среды и к перестройке минерального состава отложений. Ведущая роль в процессах анаэробного диагенеза принадлежит сульфатредуцирующим бактериям. Сульфатредуцирующие бактерии, как известно, являются гетеротрофными организмами. Энергетическим источником их существования служат органические соединения, полностью минерализуемые бактериями с образованием углекислоты. Исследования, проведенные доктором геолого-минералогических наук Аллой Юльевной Леин, показали, что эта углекислота участвует в формировании диагенетических карбонатов, дискуссия об образовании которых продолжалась долгие годы. Ключом к решению проблемы оказалось изучение изотопного состава углерода загадочных карбонатов. По этому показателю было достоверно установлено, что никакой другой источник углекислоты, кроме анаэробной деструкции органического детрита, не мог обеспечить формирование диагенетических карбонатов. Другими характерными минералами анаэробной зоны диагенеза морских осадков являются пирит и другие сульфиды железа. Подобно карбонатам, они образуются в результате взаимодействия биогенного вещества (в данном случае — сероводорода) с абиогенным. «Память» о биогенном сероводороде запечатлевается в изотопном составе серы этих минералов. Живое вещество в этом случае является лишь поставщиком исходного материала для сульфидов, но не участвует непосредственно в их формировании. Большое значение на стадии диагенеза имеет и средообразующая деятельность сульфатредуцирующих бактерий, а именно, изменение ими pH среды. Таковы события, происходящие «под занавес» начальных стадий литогенеза. Для правильного понимания процессов формирования осадочных пород в условиях биосферы при определяющем влиянии живого вещества очень важными являются три основных положения, которые В. И. Вернадский называл «биогеохимическими принципами». В его формулировке[63] они звучат следующим образом: I принцип: «Биогенная миграция атомов химических элементов в биосфере всегда стремится к максимальному своему проявлению». II принцип: «Эволюция видов в ходе геологического времени, приводящая к созданию форм жизни, устойчивых в биосфере, идет в направлении, увеличивающем биогенную миграцию атомов биосферы» (или в другой формулировке: «При эволюции видов выживают те организмы, которые своею жизнью увеличивают биогенную геохимическую энергию»). III принцип: «В течение всего геологического времени, с криптозоя, заселение планеты должно было быть максимально возможное для всего живого вещества, которое тогда существовало». Для Вернадского I биогеохимический принцип был тесно связан со способностью живого вещества неограниченно размножаться в оптимальных условиях. «Вихрь атомов», который представляет собой жизнь, стремится к безграничной экспансии. Следствием этого и является максимальное проявление биогенной миграции атомов в биосфере. II биогеохимический принцип, по существу, затрагивает кардинальную проблему современной биологической теории — вопрос о направленности эволюции организмов. По мысли Вернадского, преимущества в ходе эволюции получают те организмы, которые приобрели способность усваивать новые формы энергии или «научились» полнее использовать химическую энергию, запасенную в других организмах. В ходе биологической эволюции, таким образом, увеличивается КПД биосферы в целом. Чисто математически это показал недавно Вячеслав Викторович Алексеев, который на основе расчетов пришел к следующим выводам: «Эволюция должна идти в направлении увеличения скорости обмена веществом в системе». И далее: «Становится понятным, почему образовались ферменты, роль которых заключается в резком увеличении скоростей реакций, идущих при обычных условиях исключительно медленно». II биогеохимический принцип Вернадского получает подтверждения на самом разнообразном эмпирическом материале. Так, в 1956 г. почвовед Виктор Абрамович Ковда, ныне член-корреспондент АН СССР, обобщил результаты химического исследования более 1300 образцов золы современных высших растений. На этом обширнейшем фактическом материале автор пришел к выводу, что (за несколькими исключениями) зольность растений возрастает от представителей древних таксонов к более молодым. Эта закономерность свидетельствует о все более активном вовлечении растениями минеральных веществ в биогеохимический круговорот и является одним из частных проявлений II биогеохимического принципа. Вообще его проявления в биосфере очень многообразны и довольно неожиданны. Возьмем другой пример из области ботаники. Магаданский ботаник доктор биологических наук Андрей Павлович Хохряков недавно установил своеобразную направленность эволюции высших растений — интенсификацию смен органов в ходе индивидуального развития организма. Так, по мнению Хохрякова, у древних древовидных плаунов — лепидодендронов — смене была подвержена только часть листьев. У более продвинутых в эволюционном отношении растений — папоротникообразных — опадают также только листья, но у них в единицу времени по отношению к массе всего тела сменяется большая их часть, чем у лепидодендронов. У наиболее примитивных голосеменных — саговников — сменам также подвержены только листья, да и то за исключением оснований. У хвойных периодически сменяются ветви и кора. Наконец, на примере цветковых растений мы наиболее четко видим переход от многолетних форм (деревья и кустарники) к однолетним (травы). Этот же переход наблюдается и у других таксонов высших растений: среди древних хвощей и плаунов господствовали древовидные формы, а современные нам хвощи и плауны — травы; среди папоротников в геологическом прошлом было много древовидных, а сейчас древовидные папоротники вымирают. Такая интенсификация смен, естественно, приводит к усилению биогенной миграции атомов в биосфере. И здесь «работает» II принцип… Правда, хвойные почему-то не хотят становиться травами, а мхи, наоборот, никогда не были деревьями. А. П. Хохряков, будучи ботаником, рассматривает только растения; в более широком плане подошел к вопросу о направленности эволюции крупный советский геохимик, профессор Александр Ильич Перельман. Он вычислил, что по отношению логарифмов ежегодной продукции к «моментальной биомассе» живого вещества (коэффициент К) современные экосистемы образуют следующий ряд: I. Таежные ландшафты (0,54—0,55). II. Ландшафты влажных лиственных лесов: а) умеренного пояса (0,59—0,62); б) субтропического пояса (0,66); в) тропического пояса (0,68). III. Травяные ландшафты (0,83—0,95). Можно предполагать, что это — своеобразный «эволюционный ряд» ландшафтов и что несуществующие ныне ландшафты имели значение К меньше 0,5. Наконец, III биогеохимический принцип также связан со «всюдностью» или «давлением» жизни. Этот фактор обеспечивает безостановочный захват живым веществом любой территории, где возможно нормальное функционирование живых организмов. В связи с этим рассмотрим, как происходило освоение жизнью поверхностной оболочки Земли. В первые десятилетия нашего века ни у кого не было сомнений, что жизнь на Земле появилась лишь в кембрии, т. е., по современным датировкам, около 600 млн. лет назад. Честь открытия докембрийских микробиот принадлежит знаменитому американскому геологу Чарлзу Д. Уолкотту (1850—1927), высказавшему идею о бактериогенном происхождении докембрийских известняков и опубликовавшему в 1915 г. первую заметку с описанием остатков микроорганизмов из этих отложений. Идея Уолкотта показалась современникам абсурдной («этого не может быть, потому что не может быть никогда», используя крылатое выражение А. П. Чехова), а его описание докембрийских микроорганизмов не было принято всерьез. Сейчас Ч. Д. Уолкотта по праву называют пионером палеонтологии докембрия; учреждена специальная медаль «За изучение организмов докембрия», которая носит его имя. Другим первооткрывателем докембрийских микробиот является В. Грюнер, в 1922 г. описавший органические остатки из протерозойских железорудных формаций штата Миннесота (США). Собственно, исследованиями Ч. Д. Уолкотта и В. Грюнера и исчерпывались сведения о докембрийской жизни к тому моменту, когда Вернадский сформулировал свой знаменитый тезис об отсутствии в истории Земли безжизненных геологических эпох. Новый этап исследований начался в 40‑е годы. В 1939 г. изучением органических остатков докембрия начал заниматься А. Г. Вологдин (впоследствии — лауреат медали имени Уолкотта); в 1943 г. он организовал в Палеонтологическом институте АН СССР лабораторию по изучению древнейших организмов. В 1947 г. австралийский геолог Р. Спригг опубликовал первое описание фауны, впоследствии ставшей известной всему миру под названием «эдиакарской». Несколько лет спустя американские ученые С. А. Тайлер и Э. С. Бархгорн при микроскопическом исследовании протерозойских пород Канадского щита открыли хорошо сохранившиеся органические остатки. Крупнейшим событием в развитии науки о древнейшей жизни явился Всесоюзный симпозиум по палеонтологии докембрия и раннего кембрия, проходивший в Новосибирске осенью 1965 г. После этого симпозиума словосочетание «палеонтология докембрия» перестало шокировать даже самых отчаянных скептиков. Публикации по новой отрасли знаний пошли широким потоком. «С удивительной быстротой мы стали проникать в глубины времени и теперь твердо знаем, что менее чем шестистам миллионам лет фанерозойской палеонтологической истории предшествовали по крайней мере еще три миллиарда лет жизни», — сказал академик Б. С. Соколов. В настоящее время данные о древнейшей жизни основываются главным образом на изучении следующих объектов: а) так называемых акритарх — микроископаемых неустановленной природы, скорее всего относящихся к фитопланктону; б) строматолитов — слоистых карбонатных образований, возникших в результате жизнедеятельности цианобактерий и бактерий; в) минерализованных остатков прокариот, «запечатанных» в кремнях. Помимо этого, производятся изотопные исследования докембрийских пород, поскольку известно, что у углерода и серы соотношение изотопов пригодно для распознавания абиогенных и биогенных объектов. В нашей стране наибольшее развитие получили исследования строматолитов. Знаменательна фраза известного американского ученого профессора Престона Клауда в докладе на 27‑м Международном геологическом конгрессе: «Говорить о строматолитах на Московской сессии Международного конгресса все равно, что ехать в Тулу со своим самоваром». В последние годы в СССР успешно развивается изучение минерализованных остатков докембрийских прокариот, осуществляемое во Всесоюзном геологическом институте имени А. П. Карпинского В. К. Головенком и М. Ю. Беловой. В результате исследований, проведенных за последние четверть века, вырисовывается следующая картина развития биосферы на ранних этапах ее эволюции (табл. 7). Таблица 7 Геохронологическая шкала и основные события истории биосферы
 Дата рождения Земли — 4,6—4,7 млрд. лет назад — установлена по космохимическим и астрофизическим данным. Земля в это время подвергалась интенсивной метеоритной бомбардировке. Каковы были физико-химические условия ка поверхности, сказать трудно, однако ясно, что они были иными, чем в течение всей последующей геологической истории. Существование жизни в это время на Земле маловероятно, а геологические образования этого возраста неизвестны.
Возраст древнейших метаосадочных пород Земли — 3,8 млрд. лет (формация Исуа в Юго-Западной Гренландии). Остатков живых организмов в них не найдено, однако изотопный состав углерода однозначно свидетельствует об активных процессах жизнедеятельности, происходивших в то время. Таким образом, «начало геологической и биологической истории совпадает с точностью до сотен миллионов лет»[64].
Первые микроскопически распознаваемые органические остатки появляются в породах с возрастом 3,5 млрд. лет. Представлены они только прокариотами: цианобактериями и архебактериями. В течение первых 2 млрд. лет — половина геологической истории! — жизнь на Земле была представлена исключительно экосистемами прокариот. Некоторые из них отличаются поразительной устойчивостью. Так, экосистемы строматолитов почти не изменились за 2,7 млрд. лет своего существования.
Согласно представлениям А. Г. Пономаренко, описавшего недавно эволюцию экосистем на всем протяжении геологической истории, архейские экосистемы характеризовались значительными колебаниями биомассы и продуктивности. Жизнь существовала локально, занимая, по-видимому, лишь зону шельфа (хотя, по мнению Г. А. Заварзина, она могла распространяться частично и на континенты). Сильная эрозия на слабо заселенных или нацело лишенных жизни континентах препятствовала возникновению стабильной озерно-речной сети. Сток с континентов был преимущественно плащевым, и в море попадало много пелитового материала. Это, в свою очередь, сужало эвфотическую зону на шельфах и лимитировало развитие фотосинтезирующих прокариотов. Из-за слабого развития жизни на материках мощность континентальной коры выветривания, по данным Б. М. Михайлова, нигде в докембрии не превышала первых метров (что на 1—2 порядка меньше, чем в фанерозое).
К интервалу 1,9—2,0 млрд. лет приурочены древнейшие остатки фотоавтотрофных эукариот — микроскопических зеленых водорослей. В этом же интервале известны и первые проблематичные пеллеты. Фекальное их происхождение пока не доказано. Если оно подтвердится, то можно будет говорить о появлении в среднем протерозое первых многоклеточных животных (скорее всего, представителей зоопланктона). Другим косвенным свидетельством существования многоклеточных животных (в этом случае — бентосных) являются следы ползания в отложениях верхнего протерозоя с возрастом 1,35 млрд. лет. А первые достоверные телесные остатки многоклеточных животных относятся к отложениям венда (650—570 млн. лет). Именно эта фауна, открытая Р. Сприггом и детально описанная М. Глесснером, по месту первоначальной находки (Эдиакара в Южной Австралии) получила название эдиакарской. Сейчас она установлена в 17 крупных регионах, расположенных на пяти континентах. В нашей стране наиболее крупные местонахождения находятся в Беломорье, Приднестровье и на Оленекском поднятии в Якутии.
Эта вендская фауна поистине изумительна. Она представлена формами, не встречающимися больше ни в каких других отложениях. Поражает прежде всего размер отпечатков (остатки медуз достигают почти метра в поперечнике) и полное отсутствие скелетов даже у тех организмов, которые без них, казалось бы, немыслимы. Так, встречены отпечатки, напоминающие обнаженных моллюсков и членистоногих, и трехлучевой диск, похожий на представителя иглокожих. Помимо бентосной макрофауны (среди которой преобладают кишечнополостные), имеются остатки микрофитопланктона, бентосных водорослей и даже… грибов. Установлено, что бентосные организмы неглубоко зарывались в осадок, и степень его биогенной переработки невелика. Удивительная сохранность остатков мягкотелых организмов свидетельствует о слабом развитии сапротрофов в экосистемах докембрия.
Вендская биота является венцом органического мира докембрия. К началу фанерозоя, таким образом, представлены уже все четыре царства живых организмов (дробянки, растения, грибы и животные), а из девяти подцарств на арену жизни не вышли лишь высшие растения да, возможно, миксомицеты (они вообще неизвестны в ископаемом состоянии). При этом поверхностная оболочка планеты была еще слабозаселенной — освоены были лишь главным образом мелководья, хотя живое вещество уже принимало деятельное участие в различных геологических процессах, в частности в рудообразовании.
Рубеж докембрия и фанерозоя В. В. Меннер и Н. А. Штрейс назвали «величайшей биостратиграфической границей». Он знаменателен внезапным появлением наружных скелетов у представителей самых разнообразных типов многоклеточных животных: губок, брахиопод, членистоногих, иглокожих, моллюсков… О причинах такой внезапной «скелетизации» много спорят; единого мнения пока нет. Возможно, что это связано с прогрессирующим увеличением доли кислорода в атмосфере (минерализация скелетов у эукариот осуществляется коллагеном, а для его образования, в свою очередь, требуется достаточно высокое парциальное давление кислорода).
Процесс освоения жизненного пространства продолжался в палеозое. Предполагают, что из мелководья жизнь стала распространяться как в глубь океанов, так и внутрь материков. Освоение материков первыми высшими растениями началось в ордовике, но до середины девона оно происходило довольно медленно. Появление первых зарослей растений по берегам морей сильно изменило характер стока в океан. Возникают прибрежные лагуны — отстойники пелитового материала, дальше которых поступление терригенного вещества в океан сокращается. Вода в морях становится прозрачнее, мощность эвфотической зоны увеличивается, продуктивность фитопланктона растет.
В позднем девоне, как показала ленинградский палеоботаник Н. С. Снигиревская, на поверхности нашей планеты впервые появляются леса. Это обстоятельство значительно уменьшило поверхностный сток с континентов и ослабило эрозию. На поверхности материков впервые возникают и постоянные пресноводные водоемы — озера и реки.
Однако палеозойские и мезозойские флоры по сравнению с современными обладали еще довольно слабыми противоэрозионными и почвообразовательными свойствами. Наклонные рыхлые субстраты не зарастали и быстро размывались. Интенсивно развивалась растительность лишь в понижениях рельефа, куда сносились питательные элементы.
Привычные нам ландшафты и знакомый «ансамбль» живого вещества сформировались на Земле лишь в меловое время, после появления цветковых растений. Возникновение нового типа растительности обусловило бо́льшую устойчивость поверхности материков к эрозии; возросла скорость зарастания оголенных грунтов, ускорился процесс почвообразования. При этом за счет более активного воздействия растительности и продуктов ее опада на грунты увеличилась мощность зоны гипергенеза (особенно в тропической зоне).
Остановится ли на этом эволюция? «По-видимому, масса живого вещества растет в ходе геологического времени, и процесс захвата земной коры живым веществом еще не закончен» — так отвечает на этот вопрос В. И. Вернадский[65]. Вернадский ставит и другую проблему, для решения которой у него не хватало фактического материала: «Жизнь проникает всюду, где ее не было, но мы не можем утверждать, что это действительно были всегда свободные от жизни области планеты, никогда в другие геологические времена ею не занятые. Представляется возможным, что эти свободные от жизни области образовались в ближайшие геологические эпохи, и мы наблюдаем только освоение новыми формами жизни областей, в которых старое живое вещество почему бы то ни было исчезло… Но возможно и допущение, что мы здесь видим и реальное расширение области жизни, причем шла длительная эволюция организмов, приспособляющихся к новым условиям. Мне кажется, иначе трудно уверенно объяснять приспособления глубоководных, живущих глубже 6 км, организмов, но доказанным это считаться не может[66].
Данные современной науки подтверждают, что «реальное расширение жизни» в геологической истории действительно происходило.
Дата рождения Земли — 4,6—4,7 млрд. лет назад — установлена по космохимическим и астрофизическим данным. Земля в это время подвергалась интенсивной метеоритной бомбардировке. Каковы были физико-химические условия ка поверхности, сказать трудно, однако ясно, что они были иными, чем в течение всей последующей геологической истории. Существование жизни в это время на Земле маловероятно, а геологические образования этого возраста неизвестны.
Возраст древнейших метаосадочных пород Земли — 3,8 млрд. лет (формация Исуа в Юго-Западной Гренландии). Остатков живых организмов в них не найдено, однако изотопный состав углерода однозначно свидетельствует об активных процессах жизнедеятельности, происходивших в то время. Таким образом, «начало геологической и биологической истории совпадает с точностью до сотен миллионов лет»[64].
Первые микроскопически распознаваемые органические остатки появляются в породах с возрастом 3,5 млрд. лет. Представлены они только прокариотами: цианобактериями и архебактериями. В течение первых 2 млрд. лет — половина геологической истории! — жизнь на Земле была представлена исключительно экосистемами прокариот. Некоторые из них отличаются поразительной устойчивостью. Так, экосистемы строматолитов почти не изменились за 2,7 млрд. лет своего существования.
Согласно представлениям А. Г. Пономаренко, описавшего недавно эволюцию экосистем на всем протяжении геологической истории, архейские экосистемы характеризовались значительными колебаниями биомассы и продуктивности. Жизнь существовала локально, занимая, по-видимому, лишь зону шельфа (хотя, по мнению Г. А. Заварзина, она могла распространяться частично и на континенты). Сильная эрозия на слабо заселенных или нацело лишенных жизни континентах препятствовала возникновению стабильной озерно-речной сети. Сток с континентов был преимущественно плащевым, и в море попадало много пелитового материала. Это, в свою очередь, сужало эвфотическую зону на шельфах и лимитировало развитие фотосинтезирующих прокариотов. Из-за слабого развития жизни на материках мощность континентальной коры выветривания, по данным Б. М. Михайлова, нигде в докембрии не превышала первых метров (что на 1—2 порядка меньше, чем в фанерозое).
К интервалу 1,9—2,0 млрд. лет приурочены древнейшие остатки фотоавтотрофных эукариот — микроскопических зеленых водорослей. В этом же интервале известны и первые проблематичные пеллеты. Фекальное их происхождение пока не доказано. Если оно подтвердится, то можно будет говорить о появлении в среднем протерозое первых многоклеточных животных (скорее всего, представителей зоопланктона). Другим косвенным свидетельством существования многоклеточных животных (в этом случае — бентосных) являются следы ползания в отложениях верхнего протерозоя с возрастом 1,35 млрд. лет. А первые достоверные телесные остатки многоклеточных животных относятся к отложениям венда (650—570 млн. лет). Именно эта фауна, открытая Р. Сприггом и детально описанная М. Глесснером, по месту первоначальной находки (Эдиакара в Южной Австралии) получила название эдиакарской. Сейчас она установлена в 17 крупных регионах, расположенных на пяти континентах. В нашей стране наиболее крупные местонахождения находятся в Беломорье, Приднестровье и на Оленекском поднятии в Якутии.
Эта вендская фауна поистине изумительна. Она представлена формами, не встречающимися больше ни в каких других отложениях. Поражает прежде всего размер отпечатков (остатки медуз достигают почти метра в поперечнике) и полное отсутствие скелетов даже у тех организмов, которые без них, казалось бы, немыслимы. Так, встречены отпечатки, напоминающие обнаженных моллюсков и членистоногих, и трехлучевой диск, похожий на представителя иглокожих. Помимо бентосной макрофауны (среди которой преобладают кишечнополостные), имеются остатки микрофитопланктона, бентосных водорослей и даже… грибов. Установлено, что бентосные организмы неглубоко зарывались в осадок, и степень его биогенной переработки невелика. Удивительная сохранность остатков мягкотелых организмов свидетельствует о слабом развитии сапротрофов в экосистемах докембрия.
Вендская биота является венцом органического мира докембрия. К началу фанерозоя, таким образом, представлены уже все четыре царства живых организмов (дробянки, растения, грибы и животные), а из девяти подцарств на арену жизни не вышли лишь высшие растения да, возможно, миксомицеты (они вообще неизвестны в ископаемом состоянии). При этом поверхностная оболочка планеты была еще слабозаселенной — освоены были лишь главным образом мелководья, хотя живое вещество уже принимало деятельное участие в различных геологических процессах, в частности в рудообразовании.
Рубеж докембрия и фанерозоя В. В. Меннер и Н. А. Штрейс назвали «величайшей биостратиграфической границей». Он знаменателен внезапным появлением наружных скелетов у представителей самых разнообразных типов многоклеточных животных: губок, брахиопод, членистоногих, иглокожих, моллюсков… О причинах такой внезапной «скелетизации» много спорят; единого мнения пока нет. Возможно, что это связано с прогрессирующим увеличением доли кислорода в атмосфере (минерализация скелетов у эукариот осуществляется коллагеном, а для его образования, в свою очередь, требуется достаточно высокое парциальное давление кислорода).
Процесс освоения жизненного пространства продолжался в палеозое. Предполагают, что из мелководья жизнь стала распространяться как в глубь океанов, так и внутрь материков. Освоение материков первыми высшими растениями началось в ордовике, но до середины девона оно происходило довольно медленно. Появление первых зарослей растений по берегам морей сильно изменило характер стока в океан. Возникают прибрежные лагуны — отстойники пелитового материала, дальше которых поступление терригенного вещества в океан сокращается. Вода в морях становится прозрачнее, мощность эвфотической зоны увеличивается, продуктивность фитопланктона растет.
В позднем девоне, как показала ленинградский палеоботаник Н. С. Снигиревская, на поверхности нашей планеты впервые появляются леса. Это обстоятельство значительно уменьшило поверхностный сток с континентов и ослабило эрозию. На поверхности материков впервые возникают и постоянные пресноводные водоемы — озера и реки.
Однако палеозойские и мезозойские флоры по сравнению с современными обладали еще довольно слабыми противоэрозионными и почвообразовательными свойствами. Наклонные рыхлые субстраты не зарастали и быстро размывались. Интенсивно развивалась растительность лишь в понижениях рельефа, куда сносились питательные элементы.
Привычные нам ландшафты и знакомый «ансамбль» живого вещества сформировались на Земле лишь в меловое время, после появления цветковых растений. Возникновение нового типа растительности обусловило бо́льшую устойчивость поверхности материков к эрозии; возросла скорость зарастания оголенных грунтов, ускорился процесс почвообразования. При этом за счет более активного воздействия растительности и продуктов ее опада на грунты увеличилась мощность зоны гипергенеза (особенно в тропической зоне).
Остановится ли на этом эволюция? «По-видимому, масса живого вещества растет в ходе геологического времени, и процесс захвата земной коры живым веществом еще не закончен» — так отвечает на этот вопрос В. И. Вернадский[65]. Вернадский ставит и другую проблему, для решения которой у него не хватало фактического материала: «Жизнь проникает всюду, где ее не было, но мы не можем утверждать, что это действительно были всегда свободные от жизни области планеты, никогда в другие геологические времена ею не занятые. Представляется возможным, что эти свободные от жизни области образовались в ближайшие геологические эпохи, и мы наблюдаем только освоение новыми формами жизни областей, в которых старое живое вещество почему бы то ни было исчезло… Но возможно и допущение, что мы здесь видим и реальное расширение области жизни, причем шла длительная эволюция организмов, приспособляющихся к новым условиям. Мне кажется, иначе трудно уверенно объяснять приспособления глубоководных, живущих глубже 6 км, организмов, но доказанным это считаться не может[66].
Данные современной науки подтверждают, что «реальное расширение жизни» в геологической истории действительно происходило.
Глава шестая. Метабиосфера
Стало очевидным единство стратисферы Земли, как результат развития былых биосфер планеты.Б. С. Соколов. 1975
 На Среднем проспекте в Ленинграде все пространство между 19‑й и 20‑й линиями занимает величественное здание, отделанное серым камнем. По праву его называют «Дворцом геологии». Когда-то здесь помещался Геологический комитет — по существу, единственное геологическое учреждение дореволюционной России, а сейчас размещены три головных учреждения Министерства геологии СССР: Всесоюзный геологический институт имени А. П. Карпинского (ВСЕГЕИ), Всесоюзная геологическая библиотека и Центральный геологоразведочный музей имени Ф. Н. Чернышева. Просторный, светлый вестибюль, широкая, идущая до самого верха лестница. Ее ступени выложены плитами так называемого ортоцератитового известняка с остатками ортоцерасов (вымерших головоногих моллюсков) и морских лилий. А у подножия лестницы — другие нерукотворные создания природы: глыба каменного угля из Донбасса, ископаемые стволы деревьев, шедевр мезозойской эры — аммонит. Следы былых биосфер…
В январе 1959 г. во «Дворце геологии» открылась V сессия Всесоюзного палеонтологического общества (ВПО). Такие сессии собираются каждый год; но тема этой была необычной: «Значение биосферы в геологических процессах».
Этот представительный форум — на нем присутствовало больше 500 ученых — был первым научным съездом, где так разнообразно и всесторонне была рассмотрена роль живого вещества в формировании земной коры. Ее инициатор, известный советский палеонтолог, впоследствии — лауреат Ленинской премии, Борис Павлович Марковский (1895—1966) сделал программный доклад «Жизнь как геологический фактор». А потом были заслушаны многочисленные более специальные сообщения, посвященные вопросам биогенного происхождения карбонатных пород, горючих ископаемых, железных и марганцевых руд, фосфоритов, бокситов…
V сессия ВПО подвела итог исследований, проведенных за несколько десятилетий, прошедших после опубликования работ Вернадского о биосфере. Но этим не исчерпывается ее значение — одновременно она открыла новый этап исследований роли жизни в геологических процессах. А ровно через 20 лет в том же зале собралась XXV, юбилейная, сессия ВПО, посвященная теме «Палеонтология и эволюция биосферы».
Идеи В. И. Вернадского по праву обрели всеобщее признание. В блестящую плеяду ученых, достойно развивающих эти идеи, входят многие современники и младшие современники Вернадского. Их усилиями достигнуты выдающиеся успехи в исследовании роли жизни в формировании земной коры. Этому вопросу и посвящена последняя глава нашего повествования.
Говорят, что настоящее — это ключ к познанию прошлого. Мы воспользуемся этим ключом, чтобы лучше понять, как формировались осадочные породы в геологическом прошлом. Однако пользоваться им будем осторожно, поскольку это все-таки ключ, а не отмычка, и не ко всем дверям (в нашем случае фациальным обстановкам) он подходит.
В самом упрощенном виде осадочная оболочка Земли — это стратиграфически наложенные друг на друга следы былых биосфер нашей планеты. Все вместе они слагают метабиосферу Земли: многокилометровую оболочку, облик которой в значительной мере определяется деятельностью живого вещества.
Осадочные породы согласно современным классификациям разделяются на следующие группы: 1) обломочные; 2) глинистые; 3) аллитные; 4) железистые; 5) марганцевые; 6) фосфатные; 7) карбонатные; 8) кремнистые; 9) соли; 10) каустобиолиты.
Все осадочные породы Земли (вплоть до самых древних) сформировались в условиях биосферы. Думается, что этот тезис в наиболее краткой форме выражает геологический аспект учения Вернадского. В той или иной форме живое вещество принимает деятельное участие в формировании всех осадочных пород, и все они включают в себя какое-то (пусть иногда и малое) количество палеобиогенного вещества. Соотношение абиогенного и палеобиогенного вещества в породах может быть различным: от ископаемых углей, малозольные разновидности которых почти нацело сложены палеобиогенным органическим веществом, и известняков, состоящих из палеобиогенного неорганического вещества, — до песчаников, где палеобиогенного материала может быть совсем немного. При этом палеобиогенное вещество в осадочных породах весьма разнообразно: растительный детрит, различные остатки организмов (миоспоры, крупномерные фрагменты растений, остатки планктона, спикулы губок, зубы акул, раковины, кости позвоночных и т. д.), янтарь, копролиты и такие микробиогенные минералы, как, например, сульфиды, карбонаты, гидроокислы железа и т. д. Выходит, что не так уж далек от истины был Омар Хайям, утверждая:
На Среднем проспекте в Ленинграде все пространство между 19‑й и 20‑й линиями занимает величественное здание, отделанное серым камнем. По праву его называют «Дворцом геологии». Когда-то здесь помещался Геологический комитет — по существу, единственное геологическое учреждение дореволюционной России, а сейчас размещены три головных учреждения Министерства геологии СССР: Всесоюзный геологический институт имени А. П. Карпинского (ВСЕГЕИ), Всесоюзная геологическая библиотека и Центральный геологоразведочный музей имени Ф. Н. Чернышева. Просторный, светлый вестибюль, широкая, идущая до самого верха лестница. Ее ступени выложены плитами так называемого ортоцератитового известняка с остатками ортоцерасов (вымерших головоногих моллюсков) и морских лилий. А у подножия лестницы — другие нерукотворные создания природы: глыба каменного угля из Донбасса, ископаемые стволы деревьев, шедевр мезозойской эры — аммонит. Следы былых биосфер…
В январе 1959 г. во «Дворце геологии» открылась V сессия Всесоюзного палеонтологического общества (ВПО). Такие сессии собираются каждый год; но тема этой была необычной: «Значение биосферы в геологических процессах».
Этот представительный форум — на нем присутствовало больше 500 ученых — был первым научным съездом, где так разнообразно и всесторонне была рассмотрена роль живого вещества в формировании земной коры. Ее инициатор, известный советский палеонтолог, впоследствии — лауреат Ленинской премии, Борис Павлович Марковский (1895—1966) сделал программный доклад «Жизнь как геологический фактор». А потом были заслушаны многочисленные более специальные сообщения, посвященные вопросам биогенного происхождения карбонатных пород, горючих ископаемых, железных и марганцевых руд, фосфоритов, бокситов…
V сессия ВПО подвела итог исследований, проведенных за несколько десятилетий, прошедших после опубликования работ Вернадского о биосфере. Но этим не исчерпывается ее значение — одновременно она открыла новый этап исследований роли жизни в геологических процессах. А ровно через 20 лет в том же зале собралась XXV, юбилейная, сессия ВПО, посвященная теме «Палеонтология и эволюция биосферы».
Идеи В. И. Вернадского по праву обрели всеобщее признание. В блестящую плеяду ученых, достойно развивающих эти идеи, входят многие современники и младшие современники Вернадского. Их усилиями достигнуты выдающиеся успехи в исследовании роли жизни в формировании земной коры. Этому вопросу и посвящена последняя глава нашего повествования.
Говорят, что настоящее — это ключ к познанию прошлого. Мы воспользуемся этим ключом, чтобы лучше понять, как формировались осадочные породы в геологическом прошлом. Однако пользоваться им будем осторожно, поскольку это все-таки ключ, а не отмычка, и не ко всем дверям (в нашем случае фациальным обстановкам) он подходит.
В самом упрощенном виде осадочная оболочка Земли — это стратиграфически наложенные друг на друга следы былых биосфер нашей планеты. Все вместе они слагают метабиосферу Земли: многокилометровую оболочку, облик которой в значительной мере определяется деятельностью живого вещества.
Осадочные породы согласно современным классификациям разделяются на следующие группы: 1) обломочные; 2) глинистые; 3) аллитные; 4) железистые; 5) марганцевые; 6) фосфатные; 7) карбонатные; 8) кремнистые; 9) соли; 10) каустобиолиты.
Все осадочные породы Земли (вплоть до самых древних) сформировались в условиях биосферы. Думается, что этот тезис в наиболее краткой форме выражает геологический аспект учения Вернадского. В той или иной форме живое вещество принимает деятельное участие в формировании всех осадочных пород, и все они включают в себя какое-то (пусть иногда и малое) количество палеобиогенного вещества. Соотношение абиогенного и палеобиогенного вещества в породах может быть различным: от ископаемых углей, малозольные разновидности которых почти нацело сложены палеобиогенным органическим веществом, и известняков, состоящих из палеобиогенного неорганического вещества, — до песчаников, где палеобиогенного материала может быть совсем немного. При этом палеобиогенное вещество в осадочных породах весьма разнообразно: растительный детрит, различные остатки организмов (миоспоры, крупномерные фрагменты растений, остатки планктона, спикулы губок, зубы акул, раковины, кости позвоночных и т. д.), янтарь, копролиты и такие микробиогенные минералы, как, например, сульфиды, карбонаты, гидроокислы железа и т. д. Выходит, что не так уж далек от истины был Омар Хайям, утверждая:
Эти мертвые камни у нас под ногами
Прежде были зрачками пленительных глаз…
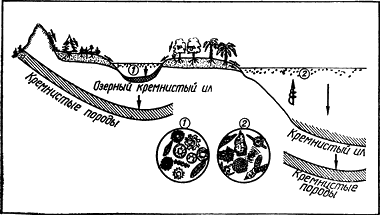 Рис. 13. Схема формирования кремнистых пород в биосфере: 1 — кремниевые организмы континентальных водоемов; 2 — кремниевые организмы морей
Рис. 13. Схема формирования кремнистых пород в биосфере: 1 — кремниевые организмы континентальных водоемов; 2 — кремниевые организмы морей
Если диатомиты — наиболее распространенные среди биогенных кремнистых пород, то описанные сравнительно недавно У. Г. Дистановым и З. И. Глезер силикофлагеллиты — самые редкие. Сложены они остатками другого представителя фитопланктона — кремниевых жгутиковых водорослей, или силикофлагеллат. В неогеновых толщах Юго-Восточной Европы силикофлагеллиты образуют самостоятельные пласты. Радиоляриты, как и предыдущие типы кремнистых пород, также сложены остатками планктона, однако радиолярии в отличие от диатомей и силикофлагеллат представляют собой не фито‑, а зоопланктон. Название «радиолярии» происходит от лат. radius — луч (радиолярии иначе называют лучевиками). Сложно построенный микроскопический скелет радиолярий представляет собой ажурный шар, от которого во все стороны, но строго закономерно, отходят длинные иглы. И кажется, что радиолярии созданы природой специально для изучения законов симметрии и совершенства. Кто-то назвал радиолярии самыми изящными и красивейшими образованиями, которые существуют в животном мире. Известны радиолярии с кембрия. По сравнению с диатомитами радиоляриты имеют меньшее распространение, однако мощность их пластов иногда достигает 38 м (палеоген о. Барбадос). Значительно более широкое развитие (по сравнению с «нормальными» радиоляритами) имеют радиоляриевые глины — породы, где панцири радиолярий обильно разбавлены терригенным материалом. Что касается спонголитов, то они когда-то считались очень редкими, а сейчас пласты морских спонголитов мощностью до 10—15 м известны в меловых и палеогеновых отложениях Украины, Кавказа и Центральной Европы, палеозое Приуралья, Северо-Востока СССР и других районов. Найдены и пресноводные спонголиты, мощность которых достигает 6—10 м. Современные спонголиты известны в Баренцевом море. Спонголиты представляют собой однородные мелкозернистые породы, состоящие из спикул донных животных, так называемых «кремниевых» губок. Спикулами называют элементы, из которых построен скелет губок. При их жизни скелет связан воедино органическим веществом, а после отмирания губок он распадается на отдельные спикулы. Длина спикул может достигать 2—3 мм, поперечное сечение измеряется сотыми долями миллиметра, а форма — очень разнообразна: от простой (игловидной) до изогнутой и звездчатой[69]. Нет сомнений, что все рассмотренные типы кремнистых пород, безусловно, сложены биогенным кремнеземом (опалом). Происхождение других кремнистых пород — трепелов, опок, яшм — вызывает дискуссии. Органические остатки в них довольно редки, и многие литологи считают эти породы хемогенными. Существует, однако, мнение, что опоки и трепелы образовались из диатомитов и спонголитов, а яшмы — из радиоляритов, утративших свою первичную структуру в ходе процессов диагенеза и катагенеза. Еще в конце прошлого века академик Феодосий Николаевич Чернышев, именем которого назван Геологоразведочный музей в Ленинграде, обнаружил в яшмах Южного Урала остатки обильной фауны радиолярий. «Наши яшмы представляют, несомненно, глубоководный шлам, — писал Чернышев в 1889 г., — причем кремнезем панцирей радиолярий и дал главнейший материал для скоплений кварца, рассеянных по всей массе породы». В первые десятилетия нашего века идею о преобладающем биогенном происхождении кремнистых пород высказывал Я. В. Самойлов. А недавнее исследование, выполненное в Институте литосферы АН СССР Валентиной Сергеевной Вишневской, показало, что породы, сложенные скелетными остатками кремневых организмов, составляют более 90% всех изученных ею кремнистых пород верхней юры — нижнего мела восточной части Малого Кавказа. На долю достоверно абиогенных пород, сформировавшихся вблизи подводных гидротерм из пересыщенных растворов, приходится менее 1% кремнистых толщ этого региона. Кремнистые осадки образуются в биосфере в результате концентрационной функции живого вещества. И в современную геологическую эпоху, и в геологическом прошлом (начиная с кембрия) накопление биогенного кремнезема происходило главным образом в морских экосистемах. При этом если первоначально оно осуществлялось донной пленкой жизни (кремневые губки) и зоопланктоном (радиолярии), то начиная с конца мезозоя основная роль в кремненакоплении переходит к фитопланктону (диатомеям). Озерное кремненакопление началось только в эоцене с появлением пресноводных форм диатомей и губок и имеет подчиненное значение. Рассказ о кремнистых породах закончим словами Вернадского: «Хотя в истории кремния роль организмов не так ярка, как в истории кальция, но и здесь без нее история данного химического элемента не может быть нами понята»[70]. Каустобиолиты — третья группа осадочных пород, характеризующаяся значительными концентрациями биогенного вещества. Термин «каустобиолиты» образован от трех греческих слов: «каустос» — горючий, «биос» — жизнь и «литос» — камень. Каустобиолитами являются торфы, сапропели, угли, горючие сланцы и нефть — современные осадки и горные породы, сложенные главным образом биогенным органическим веществом. «Органическое вещество, проникающее все вещество доступной изучению земной коры, все — биогенного происхождения… Медленными геологическими процессами эти органические вещества, остатки тел и метаболизма организмов из биосферы попадают в стратисферу, в метаморфическую оболочку», — писал Вернадский[71]. Наиболее интенсивное накопление органического вещества в современной биосфере происходит в болотах и некоторых озерах. Особняком стоит аккумуляция необиогенной органики в океанах. Обыденные понятия не всегда легко поддаются определению. Так, непросто определить, что такое болото. Существует и такое определение: «Болото можно охарактеризовать или как озеро, но со связанной водой, или как сушу, содержащую обычно 90% воды и лишь 10% сухого вещества». Действительно, одна часть сухого торфа способна удержать 15—25 частей воды! Оптимальными для торфонакопления условиями характеризуются громадные пространства территории нашей страны с умеренным гумидным климатом. Советскому Союзу принадлежит 60% мировых запасов торфа. Климатические условия являются для болотообразования «пусковым механизмом». В процессе развития торфяника роль климатических факторов уменьшается. Болото постепенно становится экосистемой, в значительной мере развивающейся по своим внутренним законам и сравнительно мало зависящей от внешней среды. Подчеркивая специфичность условий образования каустобиолитов, известный советский геолог К. Г. Войновский-Кригер (1894—1979) писал: «Накопление растительной массы, очевидно, подчинено не таким закономерностям, как накопление песка и алеврита: кроме геоморфологического фактора, здесь участвует и, может быть, играет основную роль биологический фактор». А наибольшие шансы перейти в ископаемое состояние имеют приморские и прибрежно-озерные торфяники в зонах прогибания земной коры. Другим типом экосистем, где происходит накопление необиогенного органического вещества, являются континентальные водоемы. Здесь в противоположность торфяникам накапливаются главным образом не остатки высших растений, а «сапропель» (по-гречески это значит «гнилой ил»): скопление остатков фито‑ и зоопланктона, донных и свободноплавающих организмов и экскрементов животных. Наконец, биогенное органическое вещество накапливается и в морских экосистемах, главным образом в мелководных лагунах. Основным фактором, который контролирует накопление необиогенного вещества в Мировом океане, является циркумконтинентальная зональность. По данным известного советского геохимика, лауреата премии им. В. И. Вернадского Евгения Александровича Романкевича, в периферических районах океана накапливается 87% всего органического вещества океана, в краевой части ложа — 10%, а в центральных областях — только 3%.
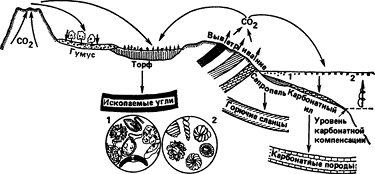 Рис. 14. Схема формирования каустобиолитов и карбонатных пород в биосфере: 1 — организмы — сапропелеобразователи; 2 — карбонатные организмы
Рис. 14. Схема формирования каустобиолитов и карбонатных пород в биосфере: 1 — организмы — сапропелеобразователи; 2 — карбонатные организмы
Так происходит накопление органического вещества в современной биосфере. Тем же путем происходило накопление биогенной органики и в геологическом прошлом (рис. 14). «Образование каменных углей имеет связь с болотами, с большими скоплениями растений, свойственными странам с сырым климатом, в устьях и дельтах больших рек, в равнинах их бассейнов, на берегах континентов и островов, в низинах областей приливов и отливов. Это все большие сгущения жизни, где масса органической материи, находящейся в состоянии медленного разложения, огромна. Возможно, что это самые большие сгущения жизни, нам вообще известные на суше», — писал Вернадский[72]. Ископаемые угли известны с девона — с того времени, когда в биосфере возникли леса. Черные, на первый взгляд невзрачные, ископаемые угли под микроскопом в шлифах чаруют гаммой оранжево-красных тонов. Сложены они большей частью углефицированными растительными тканями (их называют фитералами: суффикс тот же, что и в слове «минерал», а корень «фито» по-гречески — растение). В последнее время разработаны методы их диагностики: определяют исходный орган растения, его систематическую принадлежность и способ превращения. По этим данным можно составить представление о растительности, послужившей исходным материалом для формирования угля. Состав углей в ходе геологической истории существенно менялся. В карбоновых углях много спор; в позднекарбоновых углях впервые появляются массивные стволы древесных растений (раньше древесные растения были преимущественно трубчатыми, как сейчас бамбук, или же обладали рыхлой центральной частью — как камыш); в мезозое встречаются угли, спрессованные из листьев голосеменных или из иголочек смолы; для палеогена и неогена характерны лигниты — остатки хвойных с макроскопически различимой структурой древесины и т. д. Биогенное вещество, образованное гетеротрофами, в углях встречается довольно редко; главным образом это грибы, в палеогеновых и неогеновых углях наиболее распространенные. Найдены также остатки бактерий, членистоногих, позвоночных. Самые удивительные находки сделаны на двух буроугольных месторождениях Центральной Европы: Гейзельталь (палеоген) в ГДР и Турув (неоген) в Польше. Здесь найдены остатки богатой и разнообразной фауны позвоночных (вспомним цитированные выше слова Вернадского о «самых больших сгущениях жизни»): рыб, земноводных, пресмыкающихся (крокодилов, ящериц, змей), птиц, наконец, млекопитающих: тапиров, лошадей, сумчатых крыс, летучих мышей, полуобезьян. А в угле одного из небольших месторождений Италии известный западногерманский углепетрограф М. Тайхмюллер описала останки молодой особи «почти человека» — человекообразной обезьяны близкого нам рода. Малыш утонул в болоте… Если ископаемые угли соответствуют древним торфам, то горючие сланцы образовались из сапропелей, накапливающихся иногда в озерных, а чаще — в морских экосистемах. Классик литологии, академик Николай Михайлович Страхов (1900—1978) еще в 30‑е годы выделил следующие типы горючих сланцев: а) пелагические планктогенные горючие сланцы; б) пелагические бентогенные горючие сланцы; в) горючие сланцы заливного и предустьевого типов; г) горючие сланцы рифового типа. Легко заметить, что эти типы горючих сланцев сформированы различными концентрациями жизни, выделяемыми Вернадским: планктонной и донной пленками и прибрежным и рифовым сгущениями жизни. Биогенный материал горючих сланцев интенсивно перерабатывался донными организмами, и поэтому распознавание органических остатков в горючих сланцах иногда затруднительно. Тем не менее советский палеоботаник Михаил Дмитриевич Залесский (1877—1946), выполнивший классические исследования исходного материала горючих сланцев, установил, что их органическое вещество представлено главным образом остатками фитопланктона. Позднее было показано, что существуют и «полугорючие» сланцы, где в составе органического вещества преобладают остатки зообентоса (диктионемовые сланцы ордовика) и зоопланктона (доманиковые сланцы девона). В значительной мере биогенной является и неорганическая составляющая горючих и «полугорючих» сланцев: здесь обильно представлены панцири диатомовых водорослей, спикулы кремневых губок, остатки радиолярий и разнообразных организмов с карбонатным скелетом. Вопрос о генезисе твердых горючих ископаемых решается сравнительно просто, поскольку известны их современные аналоги. Сложнее дело обстоит с нефтью. Еще два-три десятилетия назад вопрос о ее генезисе вызывал ожесточенные дискуссии. Теперь, благодаря работам школы геологов-нефтяников, до недавнего времени возглавлявшейся членом-корреспондентом АН СССР Николаем Брониславовичем Вассоевичем (1902—1981), происхождение нефти из биогенного органического вещества мало у кого вызывает сомнения. Школой Н. Б. Вассоевича создана теория нефтеобразования, которая получила название «осадочно-миграционной». Нефть согласно этой теории — «детище литогенеза». Исходным материалом для нефти служат остатки организмов планктонной пленки жизни. «Рождение» нефти как жидкой фазы углеводородов, выделяющейся из рассеянного органического вещества, происходит в земной коре в пределах так называемой «главной зоны нефтеобразования», расположенной на глубине от 2 до 6 км ниже земной поверхности, где господствуют температуры от 60 до 160°С (именно в этом смысле нефть и является «детищем литогенеза» и именно поэтому неизвестны ее современные аналоги). Доказательством того, что нефть действительно образовалась из биогенного органического вещества, является изотопный состав входящего в нее углерода. Кроме того, в нефти найдены «биомолекулы» — соединения, аналоги которых имеются только в живом и биогенном веществе. Суммарные запасы каустобиолитов измеряются цифрой n. 1013 т. Однако, по подсчетам Н. Б. Вассоевича, это составляет только 0,36% всего органического углерода, содержащегося в осадочных породах. Биогенная органика наравне с кремнеземом, а возможно, и карбонатами представлена в метабиосфере главным образом в дисперсной форме. Итак, палеобиогенная органика метабиосферы образовалась на суше благодаря деятельности наземной пленки жизни, а в водных экосистемах — главным образом за счет планктонной пленки. Формирование каустобиолитов осуществляется при определяющем воздействии энергетической функции живого вещества. Именно это обстоятельство позволяет нам, сжигая каустобиолиты, использовать энергию солнечного луча миллионнолетней давности. Не зря каустобиолиты называют солнечными консервами. В качестве характерной особенности каустобиолитов Вернадский подчеркивал эволюцию их состава и свойств. «Они (каустобиолиты. — А. Л.) являют индивидуальность и неповторяемость в геологическом времени и представляют собой историческое явление, что выражается в их чрезвычайном химическом разнообразии и просто объясняется тем, что для каждого геологического момента организмы, тела которых они представляют, были неповторяемы, резко химически различны, так же, как различны те микробы, которые вызвали их образование и тела которых входят в их состав… В каустобиолитах мы видим ясное проявление эволюционного процесса»[73]. Не во всех осадочных породах биогенное вещество представлено такими высокими концентрациями, как в карбонатных, кремнистых породах и каустобиолитах. Беднее биогенным веществом фосфатные, железистые и марганцевые породы. К фосфатным обычно относят породы, содержащие не менее 10% окиси фосфора. Основными минералами, содержащими фосфор, являются тонкодисперсные минералы группы апатита. Полезными ископаемыми — фосфоритами — являются фосфатные породы с 12—40% окиси фосфора. Их выделяют несколько типов: пластовые, желваковые и ракушечные фосфориты, костяные брекчии и гуано. 95% всех запасов фосфоритов приурочено к морским отложениям. Мощность пластовых фосфоритов достигает 15—17 м. По внешнему виду они мало характерны. Иногда они белые, похожие на мел, чаще же темные, почти черные. Отсутствие у фосфоритов четких макроскопических признаков иногда вводит геологов в заблуждение. Так, в 30‑е годы в Казахстане (Каратау) фосфориты поначалу принимались за бокситы. А одно из сибирских месторождений было открыто не в поле, а на верхнем этаже «Дворца геологии», в Центральном геологоразведочном музее им. Ф. Н. Чернышева. Здесь в 1949 г. Наталья Александровна Красильникова, известный специалист по фосфоритам, просматривая музейные коллекции, обнаружила интересный образец. По этикетке, составленной четверть века назад, он числился песчаником, а оказался — высококачественным фосфоритом! Наверно, такие ошибки бывали и раньше — ведь название «апатит» образовано от греческого слова «апатао», что значит «обманываю»… Желваковые фосфориты представляют собой скопление конкреций. Насыщенность ими породы различна; встречаются также куски костей и фосфатизированных органических остатков: древесин, раковин моллюсков и т. д. Иногда присутствуют и фосфатизированные копролиты. Ракушечные фосфориты встречаются главным образом в отложениях ордовика. Это песчаники или конгломераты, переполненные раковинами мелких беззамковых брахиопод. Характерным примером являются так называемые оболовые песчаники Эстонии и Ленинградской области мощностью до 11 м, включающие в себя остатки Obolus и Schmidtia. Довольно редким типом фосфоритов являются костяные брекчии — пористые породы желто-бурых тонов, состоящие из обломков скелетов рыб или пещерных позвоночных. Наконец, завершает список типов фосфоритов гуано — огромные скопления (мощностью до 35 м) экскрементов морских птиц (на островах и океанских побережьях) или летучих мышей (в пещерах). На островах Чинча, расположенных у берегов Латинской Америки, ежегодно накапливается слой гуано мощностью 8 см. Таковы наиболее характерные типы фосфоритов. Происхождение их обсуждается уже много лет. Трудность проблемы заключается в том, что в фосфоритах редко сохраняются остатки сформировавших их организмов (костяные брекчии и оболовые песчаники представляют собой редкое исключение). Вместе с тем имеются убедительные доказательства того, что в круговороте фосфора живое вещество играет важную роль (рис. 15).
 Рис. 15. Схема биогеохимического цикла фосфора (по Дювиньо, 1974): 1 — накопление фосфора высшими растениями; 2 — разложение необиогенного органического вещества; 3 — экскременты; 4 — эрозия; 5 — поступление в океан; 6 — вулканический апатит; 7 — осаждение на небольшой глубине; 8 — осаждение на значительных глубинах; 9 — переход в ископаемое состояние; 10 — вовлечение в биотический круговорот диатомовыми водорослями
Рис. 15. Схема биогеохимического цикла фосфора (по Дювиньо, 1974): 1 — накопление фосфора высшими растениями; 2 — разложение необиогенного органического вещества; 3 — экскременты; 4 — эрозия; 5 — поступление в океан; 6 — вулканический апатит; 7 — осаждение на небольшой глубине; 8 — осаждение на значительных глубинах; 9 — переход в ископаемое состояние; 10 — вовлечение в биотический круговорот диатомовыми водорослями
Американский ученый Д. Мак-Коннел насчитал 76 фосфорсодержащих минералов, образующихся, скорее всего, в результате деятельности живого вещества. Кости позвоночных содержат до 60% фосфата кальция. Еще больше его в раковинах беззамковых брахиопод (помимо Obolus и Schmidtia, к ним относится и ныне живущая Lingula). В планктоне в целом двуокиси фосфора содержится в 1000 раз больше, чем в морской воде. Богаты фосфором также чешуя и скелеты рыб. До 50% фосфата кальция содержат панцири ракообразных. Концентрируется окись фосфора и в экскрементах. Так, в экскрементах морских животных содержание окиси фосфора в 3 раза выше, чем во вмещающих донных осадках. Важной «уликой» участия живого вещества в формировании фосфоритов является и высокий процент органики (до 36%) во многих фосфоритах. Залежи оболовых фосфоритов из-за высокого содержания органики имеют настолько темный цвет, что похожи на угольные пласты. В конце XIX — начале XX в. была выдвинута гипотеза об образовании фосфоритов в результате массовой гибели организмов (Дж. Меррей, Р. Ренар и Л. Кайе, А. Д. Архангельский). Весомым аргументом в пользу этой гипотезы являются костяные брекчии. В современных морях на некоторых участках морское дно покрыто слоем мертвых рыб и других животных по 2 м толщиной. Гипотеза Дж. Меррея долгое время подвергалась критике, однако сейчас показано, что в зонах апвеллинга массовые заморы рыбы происходят систематически с интервалом в несколько лет. Они-то и могли привести к накоплению костяных брекчий. Активным сторонником и пропагандистом идеи биогенного происхождения фосфоритов был Я. В. Самойлов. «Все месторождения фосфоритов, за самым небольшим исключением, органического происхождения, заключенный в них фосфор прошел через тело животного», — писал он. Действительно, копролиты часто встречаются в фосфоритах. Наиболее распространены мелкие копролиты, имеющие 0,2—1,0 мм в диаметре, но изредка попадаются и фосфатизированные копролиты крупных животных, например ихтиозавров. Много нового в познание генезиса фосфоритов внесло исследование известным геохимиком, доктором геолого-минералогических наук Глебом Николаевичем Батуриным современных фосфоритовых конкреций, обнаруженных в конце 60‑х — начале 70‑х годов на шельфах Юго-Западной Африки, Чили и Перу. Оказалось, что современное фосфоритообразование в океанах локализовано главным образом в зонах апвеллингов (впервые догадку об этом высказал в 30‑е годы ученик Вернадского геохимик А. В. Казаков). Довольно четко при этом прослеживается зависимость интенсивности фосфатонакопления от биологической продуктивности. Основным первичным концентратором фосфора из морской воды является фитопланктон, а в осадок фосфор попадает главным образом в виде фекальных пеллет зоопланктона. Осадки в ходе диагенеза дегидратируются и постепенно преобразуются в плотные фосфатные зерна и желваки. Это явление, впервые описанное Г. Н. Батуриным, в 1984 г. было признано открытием и получило название «эффект Батурина»[74]. Подобный путь фосфоритообразования, видимо, не является специфическим только для современной эпохи. Однако среди древних фосфоритов распространены и другие типы, несопоставимые с современными. Важнейшей эпохой древнего фосфоритонакопления была позднепротерозойско-раннекембрийская. По мнению советских геологов А. Л. Яншина, А. В. Ильина, Э. А. Еганова и австралийских геологов П. Дж. Кука и Дж. Г. Шерголда, обстановки и условия фосфатонакопления того времени значительно отличались от современных. Содержание фосфора в морской воде было значительно выше, чем ныне. Среди живых организмов, обладающих скелетом, процент фосфатных организмов тогда был выше, чем когда-либо. Фосфатонакопление происходило на шельфах в пределах эвфотической зоны (об этом свидетельствуют остатки водорослей в фосфоритоносной толще) и, видимо, не было связано с апвеллингом. При формировании фосфатных пород (в частности, фосфоритов) наиболее ярко проявляется концентрационная функция живого вещества. И если при сжигании горючих ископаемых человечество использует энергию, накопленную живым веществом миллионы лет назад, то, удобряя фосфоритами поля, оно возвращает в биотический круговорот фосфор обитателей древних морей. Транспортная функция живого вещества — перемещение необиогенного вещества из моря на континент — проявляется при накоплении гуано. Следующими группами осадочных пород являются железистые и марганцевые. Поскольку при их формировании живое вещество выполняет сходные функции, мы рассмотрим их совместно. Железистые осадочные породы залегают в виде пластов, линз или гнезд. В докембрийской части метабиосферы широко распространены своеобразные метаморфические породы — железистые кварциты, или джеспилиты. Они представляют собой яснослоистые породы, в которых чередуются тонкие прослойки, в различной степени обогащенные железом и кремнеземом. Возраст джеспилитов около 2 млрд. лет. Образование осадочных железистых и марганцевых руд фанерозоя происходило в морской или озерно-болотной обстановке. «Вероятно, нет ни одного металла, в образовании больших скоплений которого жизнь играла бы такую роль, как для марганца, — сказал Вернадский в своем выступлении на Конференции по вопросам генезиса руд железа, марганца и алюминия в апреле 1935 г.[75] — Для кальция и железа точно так же биогенные образования отвечают наибольшим их природным массам, но для них наряду с концентрациями такого происхождения известны не меньшие скопления, связанные с магматическими процессами, совершенно независимые от биосферы. Ничего подобного неизвестно для марганца». Марганцевыми обычно называют породы с содержанием окиси марганца больше 10%. Как и железистые породы, они залегают в форме пластов мощностью до 20 м и пластообразных или линзовидных залежей. Основные минералы марганцевых пород — окислы и гидроокислы марганца. Один из этих минералов назван в честь Владимира Ивановича вернадитом. Вопрос о роли жизни в формировании железистых и марганцевых пород волнует геологов уже больше 100 лет. И может быть, ни в каком другом вопросе науки об осадочных породах борьба мнений не принимала такие острые, подчас драматические формы. К. Г. Эренберг (это имя мы уже упоминали) еще в 1836 г. выявил в так называемых болотных (или озерных) железных рудах микробиогенные структуры. Бактерии, концентрирующие железо, были открыты в конце прошлого века. Великий русский микробиолог С. Н. Виноградский предложил для них название «железобактерии». В январе 1947 г. на сессии Отделения геолого-географических наук АН СССР выступил с докладом член-корреспондент АН СССР Александр Григорьевич Вологдин (1896—1971). Вологдин рассказал, что, следуя совету академика В. Л. Комарова поискать в горных породах остатки древних бактерий или следы их деятельности, оп предпринял микроскопическое исследование широкого круга геологических объектов. В противоположность ранее предпринимавшимся попыткам Г. Молиша, Я. В. Самойлова и В. С. Буткевича Вологдину посчастливилось найти оруднелые остатки железобактерий в железистых и марганцевых породах. Доклад А. Г. Вологдина вызвал суровую критику собравшихся. Докладчика упрекали в бездоказательности основных положений, в излишнем увлечении микроскопированием, в неубедительности фотографий микроорганизмов… И лишь четверть века спустя эта работа получила иную оценку. «Гипотеза Вологдина была отвергнута геологами, как совершенно необоснованная, что отчасти можно объяснить неподготовленностью геологической аудитории к восприятию высказанных идей. Главной же причиной была, как нам кажется, нечеткость представленных автором микрофотографий и недостаточная изученность в то время флоры железобактерий. Многие из известных теперь представителей этой группы в 40‑х годах еще не были описаны, и поэтому обнаруженные автором микроподобные структуры не имели достаточно сходных в морфологическом отношении живых аналогов». Эти слова, сказанные в 1973 г., принадлежат известному микробиологу, доктору биологических наук Т. В. Аристовской. Достигнуть значительного прогресса в области изучения железобактерий микробиологам удалось при исследованиях современного железомарганцевого рудообразования в лабораториях природы. Одной из таких природных лабораторий явились озера северо-запада нашей страны. Озерные руды представляют собой конкреции, содержащие до 80% окислов железа и марганца. По размеру конкреций выделяют различные виды руд: пороховую, градовидную, гороховидную, монетную, тарелочную, лепешечную, щитовидную и др. В 1926 г. при исследовании озерных руд Карелии было сделано выдающееся открытие. Оно принадлежит Борису Васильевичу Перфильеву (1891—1969), впоследствии профессору, лауреату Ленинской и Государственной премий. В озерных рудах Б. В. Перфильеву удалось обнаружить несомненные бактериальные структуры, а в донных осадках — микрозональность, обусловленную различной интенсивностью деятельности железобактерий. Борис Васильевич высказал предположение, что и в прошлые геологические эпохи механизм накопления железа также был бактериальным. Открытие Б. В. Перфильева послужило толчком для развития исследований по бактериогенному рудообразованию. Были описаны новые роды и виды железобактерий, принимающие участие в этом процессе. В 1936 г. Б. В. Перфильев описал новый род бактерий, который, по современным данным, играет основную роль в концентрации марганца и железа в донных осадках. Борис Васильевич именовал его «Metallogenium» — «рождающий металл». А другой исследователь, В. О. Калиненко в 1949 г. назвал один из новых видов железобактерий Leptothrix Wernadskyi. В результате кропотливых работ микробиологов было установлено, что образование железомарганцевых конкреций в озерах и болотах происходит в два этапа. Сначала окисные соединения этих металлов восстанавливаются в донных осадках сульфатредуцирующими и другими бактериями, и восстановленные соединения железа и марганца поступают из ила в воду. На втором этапе происходит обратный процесс — железобактерии окисляют растворенные в придонной воде закисные соединения железа и марганца с образованием ферригидрита, вернадита и других минералов. Установлено, что образование вернадита возможно лишь при очень быстром окислении двухвалентного марганца до четырехвалентного — настолько быстрого, что осуществляться оно может только с участием живого вещества. При этом бактериогенное окисление железа и марганца происходит при столь низких их концентрациях в растворах, что хемогенное осаждение элементов исключается. Другим естественным полигоном, где проводили исследования микробиологи, были области железомарганцевого конкрециообразования Мирового океана. На океанском дне на глубине от 4 до 6,5 км эти конкреции иногда образуют сплошной покров, напоминающий булыжную мостовую. Площадь, ими покрытая, составляет около 10% дна Мирового океана. Марганца в конкрециях около 25%, железа — около 15%; остальные 60% приходятся на долю кислорода, никеля, кобальта, меди и других 35 элементов таблицы Менделеева. Наравне с торфом, конкреционными фосфоритами и озерными железомарганцевыми рудами и в противоположность всем древним ископаемым это — возобновляемый источник сырья. Ежегодный прирост запасов железомарганцевых конкреций составляет миллионы тонн. Океанские конкреции иногда считают абиогенными, однако исследования показали, что живое вещество играет важную роль в их формировании. Железо в водной толще интенсивно потребляется фитопланктоном. Осаждение железа и марганца в значительной мере осуществляется фильтрующими организмами планктонной и донной пленок жизни. Дальнейшее преобразование соединений этих элементов на дне морей и океанов производится бактериями, многочисленные чехлы которых были найдены в конкрециях. На основании опыта этих исследований, накопленного за последние десятилетия, Т. В. Аристовская произвела ревизию выводов А. Г. Вологдина. Для изучения ею были выбраны образцы нескольких железистых минералов: лимонита, гетита и магнетита. Исследование производилось микроскопически в проходящем свете после специальной подготовки препаратов. Что же увидела Татьяна Вячеславовна? Гетит и лимонит оказались полностью состоящими из железистых отложений, повторяющих очертания клеток замурованных в них железобактерий. Сравнивая микроскопическую структуру минеральных масс с формами роста железобактерий, можно было обнаружить черты сходства между остатками микроорганизмов, слагающих минералы, и фрагментами колоний некоторых современных бактерий. Магнетит же в противоположность гетиту и лимониту показал полное отсутствие бактериальной структуры. Т. В. Аристовская изучала только железистые породы. Марганцевые руды палеогена месторождений Чиатуры и Тетрицкаро были исследованы ранее московским геологом, доктором геолого-минералогических наук Лазарем Ефимовичем Штеренбергом. В препаратах из руд, обработанных щавелевой кислотой, Л. Е. Штеренберг также описал биогенные структуры, сходные с колониями уже известного нам рода Metallogenium. Интенсивное исследование докембрийских железных руд различных районов СССР в настоящее время производится в Институте геологических наук АН УССР Владимиром Ивановичем Лазуренко. По его данным, количество найденных чехлов железобактерий в докембрийских рудах исчисляется тысячами. Рудные прослои в полосчатых рудах иногда оказываются нацело сложенными остатками железобактерий. После этих работ А. Г. Вологдина, Т. В. Аристовской, Л. Е. Штеренберга и В. И. Лазуренко можно считать доказанным биогенное происхождение не только современных железомарганцевых руд, но и их древних аналогов, включая докембрийские джеспилиты. При этом показано, что образование биогенного вещества железистых и марганцевых руд происходит несколько иначе, чем карбонатных, кремнистых и фосфатных пород: решающую роль здесь играет не планктонная, а донная пленка жизни. Однако по условиям формирования они сходны: во всех случаях накопление биогенного вещества происходит в водных экосистемах — в континентальных водоемах или в море; определяющую роль на стадии седиментогенеза и диагенеза играет концентрационная функция живого вещества. Вопрос о роли жизни в формировании аллитов и солей остается более дискуссионным. Аллитами, или глиноземистыми породами, называют горные породы, богатые свободным окислом алюминия(глиноземом). К их числу относится алюминиевое сырье — бокситы, горная порода, состоящая в основном из минералов гидроокиси алюминия. Как и фосфориты, бокситы очень разнообразны по своему внешнему виду. Известны бокситы черного, белого, желтого, красного и вишневого цветов, пористые и плотные, глинистые и песчаные. На первом курсе вуза на практических занятиях по общей геологии студентам в составе учебной коллекции иногда подсовывают битый кирпич — и неофиты от геологии дружно определяют его как боксит. Когда-то такой грех случился и со мной… Название «бокситы» связано отнюдь не с популярным видом спорта, а с местечком Бо на юге Франции, где бокситы впервые были найдены в 1821 г. Согласно господствующей точке зрения бокситы представляют собой ископаемую кору выветривания пород алюмосиликатного состава или продукты ее переотложения. Первые сведения о том, что расщепление алюмосиликатов может осуществляться биогенно за счет деятельности диатомовых водорослей, были получены экспериментально английскими учеными Дж. Мерреем и Р. Ирвином еще в конце прошлого века. Впоследствии В. И. Вернадский[76] и его ученики А. П. Виноградов и Е. А. Бойченко повторили эти эксперименты. Их результаты не оставляли места для сомнений: диатомовые водоросли действительно расщепляют алюмосиликаты и используют кремнезем для построения своих панцирей; глинозем же остается в водной среде в виде коллоидного раствора. Еще в самом начале нашего века В. И. Вернадский сделал вывод о большом, порообразующем значении этого процесса[77]. Бокситы, как известно, содержат значительное количество окиси железа. Поэтому в 1903 г. Т. Холландом было высказано предположение, что процесс формирования бокситов может быть связан с деятельностью железобактерий. Однако это было только предположением, не подтвержденным фактическим материалом — до тех пор, пока А. Г. Вологдин не обратил внимание на то, что остатки железобактерий, помимо железистых пород, имеют широкое распространение и в аллитах, где они составляют до 40—45% объема породы. Прошло еще несколько десятилетий, и во Всесоюзном геологическом институте им. А. П. Карпинского стали проводиться эксперименты по биогенному разложению вулканических пород бактериями. На основании этих опытов Л. Е. Крамаренко и О. Ф. Сафонова сделали вывод: в присутствии микроорганизмов полное превращение габбро-лабрадорита в глинозем в аэробной среде происходит за один год, а в анаэробной — за 7 лет. В анаэробной среде отложения глинозема без участия микроорганизмов вообще не происходит. Итак, один из возможных путей образования бокситов — разложение абиогенного вещества непосредственно микроорганизмами. Однако выветривание первичных пород живое вещество регулирует и опосредствованно — продуктами своего опада (необиогенным органическим веществом). Исследования почвоведов показали, что в современном выветривании в зоне муссонного климата органическое вещество гниющей подстилки тропического леса играет огромную роль. На этом основании известный советский геолог, вице-президент АН СССР, академик Александр Леонидович Яншин делает вывод, что «главная эпоха бокситообразования, эпоха разрушения горных пород, при котором кремнезем выносится, а глинозем остается в коре выветривания, образуя крупные месторождения алюминиевых руд — бокситов, могла наступить только после появления покрытосеменной растительности, примерно 100 млн. лет назад. С появлением покрытосеменных и образованием формации тропического леса почва в низких широтах стала получать кислоты, которых раньше в ней не было. Поэтому поиски бокситов в более древних корах выветривания, очевидно, будут малорезультативными». Так учение в роли жизни в геологических процессах помогает геологам искать рудные залежи в отложениях определенного возраста. Помимо деструктивной, при формировании бокситов проявляется и концентрационная функция живого вещества: известно, что некоторые растения накапливают алюминий (например, зола плаунов содержит его до 30%). Геологический аспект этой проблемы первым осознал академик Лев Семенович Берг (1876—1950), человек, чью специальность определить нелегко. Он был географом и ихтиологом, биологом-теоретиком и историком науки. Геологом Л. С. Берга обычно не считают, но среди его работ есть и такие: «О предполагаемой периодичности в образовании осадочных пород (1944), «Жизнь и почвообразование на докембрийских материках» (1944), «О происхождении уральских бокситов» (1945), «Почвы и водные осадочные породы» (1945), «О происхождении железных руд типа криворожских» (1947), «Солнечная активность в геологическом прошлом» (1947)… Этими работами Л. С. Берг вписал свое имя и в анналы геологической науки. Лев Семенович предполагал, что бокситы могли образоваться путем минерализации и переотложения остатков высших растений, содержащих значительное количество глинозема. Отложение бокситов, по мнению Берга, происходило на суше, в болотах или в мелких заболоченных пресноводных водоемах. Эти соображения у геологов долго не находили понимания. Неожиданная поддержка пришла со стороны микробиологов. Алюминий, как известно, образует в теле растений комплексные алюмоорганические соединения. Т. В. Аристовская и Л. В. Зыкина экспериментально изучили разложение соединений такого рода под действием железобактерии Metallogenium и установили, что в этом случае происходит осаждение гидроокиси алюминия. «Рождающая металл» бактерия оказалась способной производить руды не только железа и марганца, но и алюминия. С полным основанием авторы заключили: «Мы полагаем, что обнаруженные нами явления могут послужить ключом к разгадке бокситообразования». Для проверки этой гипотезы Т. В. Аристовская изучила образцы современных и древних бокситов. Из современных латеритных бокситов Гавайских островов была выделена культура Metallogenium, развивающаяся в симбиозе с пенициллиновым грибком. Изученные образцы древних бокситов жизнеспособных микробных клеток не содержали, однако в массовых количествах в них были обнаружены фоссилизированные клетки, морфологически сходные с бактериальными. Эти факты могут служить серьезными аргументами возможности биогенного образования бокситов. Ярлык «хемогенная порода» с бокситов, видимо, придется снимать. К чисто хемогенным образованиям обычно относят соли — осадочные породы, состоящие из легко растворимых в воде минералов. Бесспорно, что процесс выпаривания насыщенных растворов и кристаллизации солей — этот эффектный процесс, который мы помним со школьных лет, — происходит абиогенно. Однако откуда берутся соли в природном растворе? В аридных районах, где происходит соленакопление, растения характеризуются высоким содержанием золы (до 40—55%) и необычным ее составом. Содержание натрия в золе иногда составляет 65%, хлора — до 48, SO42− — до 36, магния — до 4,5%. Сбрасывая листву, растения засоляют почву. При разложении отмершей органики NaCl, Na2SO4, MgCl2 выщелачиваются, мигрируют с водами поверхностного и подземного стока и собираются в бессточных впадинах или в Мировом океане. По подсчетам В. А. Ковды (1944), 16 млн. т солей (половина всего солевого привноса) в Аральское море поступает ежегодно из необиогенного вещества. Жизнь сбрасывает туда ненужный ей избыток солей — хлоридов и сульфатов. Эти соли выпадают абиогенно из пересыщенных растворов в соленых озерах или в отшнурованных от моря лагунах. Таким же путем образуются и отложения соды — с той лишь разницей, что натрий в этом случае в процессе миграции связывается с углекислотой воздуха. Что же касается калия, то он в процессе миграции быстро перехватывается живым веществом. Поэтому при наличии развитого растительного покрова крупные месторождения калийных солей таким путем формироваться не могут. Приуроченность крупнейшего в Союзе месторождения калийных солей — Непского — к кембрийским отложениям обусловлено, по мнению А. Л. Яншина, именно отсутствием в то время наземной растительности. Мы рассмотрели последовательно восемь групп осадочных пород и везде находили в той или иной форме проявления деятельности живого вещества. Из всех существующих групп остались только две — обломочные и глинистые. Эти группы, как известно, образуются в результате выветривания и переотложения первичных вулканических, а также осадочных пород. Какую роль играет в этих процессах живое вещество, мы рассмотрели в предыдущей главе. Могут ли обломочные и глинистые породы образоваться при отсутствии живого вещества? Вряд ли. Для глин это практически исключено: они слишком дисперсны, чтобы какой-либо другой механизм, помимо живого вещества, мог обеспечить столь мелкий размер слагающих их частиц («Мы не имеем на земле более могучего дробителя материи, чем живое вещество» — эти слова Вернадского хорошо известны). Они слишком дисперсны и для того, чтобы выпадать в осадок без участия живого вещества: накопление глинистых пород в биосфере происходит путем биоседиментации. При этом каждая глинистая частица неоднократно проходила через «кишечную тюрьму» многоклеточных животных. В поэтической форме эту особенность глинистых пород выразил еще Омар Хайям:
Я к гончару зашел: он за комком комок
Клал глину влажную на круглый свой станок.
Лепил он горлышки и ручки для сосудов
Из царских черепов и из пастушьих ног.
Последние комментарии
23 часов 33 минут назад
1 день 7 часов назад
1 день 22 часов назад
2 дней 2 часов назад
2 дней 2 часов назад
2 дней 2 часов назад