Биофизика познает рак [Инал Георгиевич Акоев] (fb2) читать онлайн
- Биофизика познает рак (и.с. От молекулы до организма) 1.32 Мб, 178с. скачать: (fb2) - (исправленную) читать: (полностью) - (постранично) - Инал Георгиевич Акоев
[Настройки текста] [Cбросить фильтры]
[Оглавление]
И. Г. Акоев Биофизика познает рак
АКАДЕМИЯ НАУК СССР Ответственный редактор член-корреспондент АН СССР А. М. КузинРецензенты: доктор биологических наук Ю. Б. Кудряшов, кандидат биологических наук М. М. Виленчик
Введение
Бурное развитие физики в XX в., проникновение как ее подходов к сложным проблемам, так и ее методов и интерпретаций результатов исследований в биологию породили быстро прогрессирующую ветвь знания — биологическую физику. Объектами исследований биофизики являются низко- и высокомолекулярные соединения, важные в биологическом отношении, разнообразные одно-и многоклеточные организмы животного и растительного мира, биоценозы. Исследуются самые общие закономерности функционирования биологических структур и осуществления биологических процессов с точки зрения физики и математики, на основе которых в дальнейшем изучаются более конкретные вопросы. Среди этих закономерностей природа сил взаимодействия, кинетика процессов, самоорганизация и эволюция систем, механизмы преобразования энергии, механизмы подвижности, восприятия, переработки и хранения информации, механизмы влияния физических факторов, пути управления системами (рис. 1). Биологическая физика все более становится теоретической основой современной биологии и медицины. Она состоит из трех основных разделов: молекулярной биофизики, биофизики клетки и биофизики сложных процессов. В изучение рака много полезного вносят исследования всех трех разделов современной биофизики. Сложность изучения причин и механизмов развития злокачественных заболеваний заставляет использовать различные подходы к познанию нераскрытых тайн рака. Биофизика сложных процессов предложила свои подходы к изучению проблемы рака. Биофизика сложных процессов исследует механизмы взаимодействия разных систем как в горизонтальном плане, т. е. между системами одного уровня организации, так и в вертикальном плане — между системами разных уровней организации.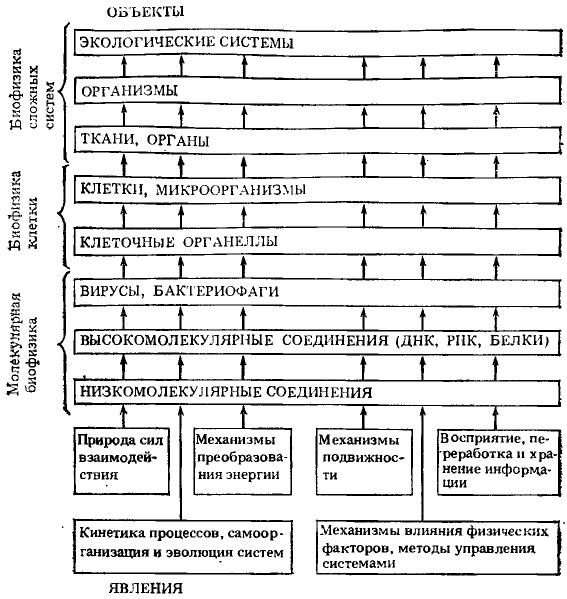 Рис. 1. Явления и объекты изучения современной биологической физики (по Г. Р. Иваницкому)
Рис. 1. Явления и объекты изучения современной биологической физики (по Г. Р. Иваницкому)
В условиях все продолжающейся дифференцировки наук и углубления знаний о физических и физико-химических основах структуры и функции отдельных биологических процессов значение биофизических исследований взаимодействия их друг с другом, а также взаимовлияния их возрастает и становится необходимым при изучении явлений, возникших в результате изменения в нескольких и разных взаимодействующих системах. Многие болезни как раз и являются следствиями нарушений не только в каких-либо отдельных системах организма, но и нарушений взаимодействия между ними. Обычно специалисты хорошо знают изучаемую ими систему и слабо — другие системы (например, гематолог хорошо знает систему кроветворения, но хуже — эндокринную систему). В то же время даже в пределах одной системы имеется также четкая дифференциация знаний по процессам, происходящим на разных уровнях ее организации. Гематолог-клиницист — специалист по заболеваниям крови — хорошо знает проявления таких болезней в клинической картине и в традиционных показателях лабораторного анализа периферической крови и пунктатов костного мозга, но нередко он менее хорошо знает гистологическую и морфологическую основу болезни (это прерогатива морфологов и патологоанатомов) и может недостаточно знать тонкости дифференцировки и пролиферации морфологически неразличимых стволовых клеток. Еще сложнее обстоит дело с использованием в клинике достижений экспериментальной цитологии, цитогенетики, молекулярной биофизики и молекулярной генетики. Здесь имеются свои узкие направления знаний, объединить которые с целью интерпретации состояний целого организма чрезвычайно сложно. Незнание или слабое знание биофизики межсистемных взаимодействий в механизмах развития болезней затрудняет получение ответов на основные вопросы — почему происходят те или иные явления, почему они происходят в данной последовательности, а не в иной и т. д. Особую сложность представляют предпатологические состояния, когда нет очевидных проявлений заболевания, когда организм уже не здоров и еще не болен. Среди заболеваний, имеющих неблагоприятный прогноз, первое место занимает рак — разнообразные злокачественные опухолевые заболевания, и среди них лейкоз — рак крови (или лейкемия — белокровие), самое распространенное заболевание после воздействия радиации и многих токсических соединений. Характерной чертой раковых заболеваний является длительный, иногда чрезвычайно длительный (до двух десятков лет и более) скрытый период до появления клинических признаков и жалоб больного. Подходы биофизики к сложным проблемам, а также ее методы исследований сложных процессов оказались особенно ценными в изучении состояния организма в период до появления клинических форм заболевания, т. е. в предпатологический период, когда еще возможно эффективное лечение и возвращение организма к норме. Именно с позиции биофизики сложных процессов и будут ниже рассмотрены проблема рака на примере лейкоза и проблема предраковых и предлейкозных состояний.
Лейкоз — генерализованная форма рака
Рак — проблема XX века
Строго научное определение «рак» объединяет лишь злокачественные опухолевые заболевания кожи и производных его зачаткового листка. Более широкое понятие рака, распространенное, в частности, у неспециалистов и среди населения и используемое нами здесь, объединяет все злокачественные опухолевые заболевания, число которых превышает сто наименований. Среди них и быстро завершающиеся смертельным исходом саркомы некостной ткани, и вяло текущие хронические заболевания лимфатической системы, сопровождаемые нередко длительными ремиссиями (т. е. временными улучшениями здоровья). Канцерогенез (от латинского слова «канцер» — рак) описывает многоэтапный процесс перехода здоровой ткани в злокачественную, важнейший этап которого — малигнизация связан с трансформацией, т. е. с непосредственным приобретением клетками свойств злокачественности. Главные среди этих свойств следующие: способность к безудержному размножению клеток, переставших подчиняться регулирующим механизмам организма-хозяина; способность проникать, прорастать в окружающие здоровые ткани и вызывать в них нарушения в функционировании и повреждения; способность отрываться от основной опухоли и переноситься с током крови и лимфы в другие органы и ткани и образовывать в них вторичные опухоли — метастазы; способность вызывать общие нарушения обмена веществ и отравлять организм продуктами распада опухолевых клеток. Коварство рака связано с тем, что организм не чувствует появления начальных этапов канцерогенеза, и обычно впервые обращаются к врачу, когда началась уже прогрессия сформировавшейся опухоли (ее быстрый рост), препятствующий нормальному функционированию ряда органов. Из этого следует чрезвычайная важность исследований механизмов самых ранних этапов канцерогенеза в доклинический период. Коварство рака связано и с бесконечным разнообразием факторов, способствующих и вызывающих рак. Поиск общих для них закономерностей также относится к первоочередным задачам исследователей. Рак стал болезнью XX в. По числу смертельных исходов рак занимает второе место после сердечно-сосудистых заболеваний. Однако страх у населения перед угрозой заболеть раком выше, чем каким-либо другим заболеванием. Это связано как с действительным повышением частоты злокачественных заболеваний за последние 50—100 лет, так и с определенной неинформированностью населения о возможности современными лечебными средствами противостоять неотвратимому смертельному исходу, особенно в случае раннего обращения больного к врачу. Повышение частоты злокачественных заболеваний связано с улучшением выявления рака, с увеличением средней продолжительности жизни людей и с увеличением контингента людей, имеющих контакт с различными потенциально канцерогенными факторами на работе и в окружающей среде. Рак не есть болезнь только XX в. и только человека. Различными формами злокачественных опухолевых заболеваний практически болеют все виды многоклеточных организмов животного и растительного мира. Рак старше человека. Рак (остеосаркомы и другие формы) обнаружен и среди ископаемых остатков растений и животных, живших задолго до появления человека на Земле, например у динозавров. Следовательно, эволюционно-биологический подход к проблеме рака может оказаться очень полезным в изучении его загадок, в изучении этиологии (причин) и механизмов его развития. О причинах возникновения рака и возможностях его лечения среди неспециалистов бытует много легенд, далеких от реальной действительности. На самом деле имеется большое количество разнообразных опухолевых злокачественных заболеваний, которым тем не менее присущи некоторые общие черты. В случае несвоевременной диагностики и лечения действительно неотвратимо наступает смертельный исход. Однако нельзя к диагнозу рака относиться как к смертельному приговору. К настоящему времени накоплено много материалов, свидетельствующих о важности участия самого больного в определении исхода заболевания, о еще большей важности состояния организма в предшествующий период, когда человек был еще здоровым и не подозревал, что многое из его образа жизни, его привычек и поведения может привести к возникновению изменений в организме, способствующих развитию рака. Эти изменения накапливались в так называемый предпатологический период. Большинство форм рака носит локальный характер, но оказывает пагубное влияние на весь организм. Наиболее генерализованной формой опухолевых заболеваний является лейкоз — рак крови, что связало с интегрирующей ролью системы крови. Лейкозным клеткам присущи те же основные отличия от нормальных клеток, что и клеткам других злокачественных опухолей, и те же причины их возникновения. По лейкозу имеются надежные статистические данные, особенно по лейкозам радиационной этиологии, и более изучены взаимоотношения опухоли и организма. Наконец, именно на примере лейкоза наиболее подробно исследован период предпатологических изменений, определяющих в реальных условиях вероятность злокачественного заболевания человека. Поэтому в дальнейшем мы будем в основном излагать материал, относящийся к лейкозам, обобщая основные клинические сведения о лейкозах человека и подчеркивая определяющую роль нарушений взаимосвязи различных систем организма в предпатологический период, имеющих неспецифический по отношению к лейкозу характер, механизм их возникновения и пути предотвращения или устранения их. По вопросам этиологии и возможным механизмам развития рака имеется большое число работ. В них обсуждается вопрос, почему может произойти малигнизация клеток у отдельных лиц, но нет ответа на другой — почему у абсолютного большинства людей, подвергающихся такому же воздействию канцерогенных факторов и условий, злокачественные заболевания не возникают. Кроме того, они не дают возможности понять всю последовательность событий, происходящих на уровнях молекулярно-клеточном и тканевом, на уровне физиологических систем организма, формирующих предраковые и предлейкозные состояния с последующим возможным переходом в рак и лейкоз. Характерной особенностью последних лет стали серьезное обсуждение и поиск доказательств обратимости злокачественного процесса. Поднимается вопрос: является ли соматическая мутация, т. е. возникшее наследственное изменение, основным механизмом злокачественной трансформации? На факты, не совместимые с мутационной гипотезой, обращал внимание В. С. Шапот [1975]. Он писал: «Отсутствие в раковой клетке какого-либо специфического белка делает неправдоподобным предположение об участии мутировавших структурных генов в неопластическом превращении. Вся сложность проблемы состоит в том, что раковая клетка создается из нормальных элементов» (с. 237). Об этом писали и другие авторы. Аналогичное положение и с хромосомными изменениями. В большинстве случаев не выявляется однозначной закономерной связи между хромосомной характеристикой опухоли или клеточной линии и ее морфологическими и физиологическими свойствами (Е. А. Погосьянц, А. Д. Захаров). Так называемая филадельфийская хромосома встречается не у всех больных лейкозом (даже при хроническом лимфолейкозе), но в то же время она может быть и в клетках, не подвергающихся малигнизации. В последние годы удалось доказать возможность получения нормальных поколений клеток из некоторых опухолевых тканей. На основе анализа факта наличия у опухолевых клеток иммунных, обменных и ультраструктурных признаков, характерных для эмбриональных клеток, М. Г. Эренпрейс делает категорическое заключение о том, что ничего другого, кроме эмбрионизации, при канцерогенезе не происходит и какими-либо особыми свойствами, кроме эмбриональных, опухолевые клетки не обладают. Эта крайняя точка зрения также отвергает основополагающую роль в первичных механизмах малигнизации мутаций, генных и хромосомных нарушений. Как же быть тогда с гипотезой о вирусной природе происхождения злокачественных опухолей и лейкоза? Последние обзоры по этой проблеме (например, Н. П. Мазуренко) подтвердили реальность вирусной этиологии ряда злокачественных заболеваний у животных, но не у человека (кроме редко встречаемой в отдельных районах Африки лимфомы Беркита). Для человека рак — незаразная болезнь. Для него признаются ведущими факторами, по мнению Н. П. Мазуренко, процессы, происходящие в самом организме, и прежде всего связанные с его старением или с дисбалансом его систем в более молодом возрасте. У животных опухолевые процессы вирусного происхождения в эксперименте развиваются только при определенном гормональном статусе организма, при определенном повреждении и раздражении тканевой системы, в которую инокулируется вирус, или при искусственном изолировании группы клеток от влияния нормальной тканевой и организменной регуляции. Не следует также забывать, что злокачественная трансформация определенного числа клеток еще не означает неизбежного развития злокачественного заболевания. При сохранении системы иммунологического надзора организм способен обезвреживать и уничтожать большое число клеток, приобретших черты иммунологической чужеродности.Эпидемиология лейкоза и других необластозов
Эпидемиология изучает географию заболевания и вероятность ого связи с окружающей средой, традициями быта, питания и поведения. По официальным статистическим данным разных стран, на злокачественные опухолевые заболевания кроветворной системы (они объединяются общим термином «гемобластозы») приходится около 50 % всех случаев смерти в возрасте до 30 лет. Это связано с тем, что в отличие от ряда других опухолей многие формы гемобластозов встречаются и в детском и в юношеском возрасте. У человека гемобластозы встречаются у 10—12 % онкологических больных. С учетом потери трудоспособности гемобластозы занимают обычно третье место среди новообразований после рака желудка и легких. В течение многих десятилетий наблюдалось увеличение заболеваний гемобластозами в СССР и в Европе. Однако в последнее десятилетие замедлился рост частоты заболеваний лейкозом. Для населения США это стало заметно примерно с 1965 г. Тем не менее в 1977 г. из пяти наиболее распространенных злокачественных заболеваний среди мужской части населения США в возрастных группах до 15 лет и 15—34 года больше всего погибло от лейкоза. В 1979 г. число больных гемобластозами в США составляло около 88 300, а в мире — около 1 млн 766 тыс. человек. В СССР общее число больных злокачественными заболеваниями лимфатической и кроветворной ткани составило в 1979 г. 23 139 человек, или 8,8 случая на 100 000 жителей [Напалков и др., 1981]. Доля этих заболеваний среди всех онкологических болезней была практически одинаковой для городского (4,4%) и сельского (4,3%) населения при очень больших различиях в отдельных республиках (2,6 и 13,6 случая на 100 000 населения соответственно для Таджикистана и Литвы). По прогнозам до 1990 г. заболеваемость раком крови еще будет расти. Для человека наиболее надежные статистические сведения получены о неопластических процессах радиационной этиологии (медицинское применение ионизирующей радиации и радиационные последствия взрывов атомных бомб в Хиросиме и Нагасаки). Джон Бойс обработал и обобщил в Национальном институте рака США накопленные в разных странах к 1980 г. сведения о частоте возникновения различных неопластических заболеваний за длительные отрезки времени (десятилетия) после медицинского применения радиации (радиотерапия анкилозоспондилита, острого мастита, гипертрофии тимуса у детей, тонзиллита, туберкулезного аденита, гемангиомы; радиационная кастрация женщин; облучение волосяного покрова головы детей; флюороскопия груди; радиационная пельвиметрия беременных женщин; общая рентгенодиагностика; радиотерапия шейки матки; введение в организм радиоактивных веществ: радия-224 для лечения костного туберкулеза и анкилозоспондилита, фосфора-32 при полицетимии, йода-131 при тиреотоксикозе, тория-232 в качестве контрастного средства при рентгенодиагностике). Обрабатывались единым методом данные, имеющие достаточное дозиметрическое описание. Сведения относятся к более чем 150 тыс. человек, получавших радиационные воздействия. На основе обобщения автором сделаны следующие основные заключения: 1) однократное воздействие радиации в определенной дозе вызывает увеличение частоты злокачественных заболеваний спустя много лет; 2) радиационный рак не отличается от спонтанно возникающего и вызванного другими причинами; 3) молочная железа, щитовидная железа и кроветворение являются наиболее чувствительными к индукции злокачественных опухолей радиационной этиологии; 4) злокачественные заболевания крови (лейкозы) являются наиболее вероятными среди других типов злокачественных заболеваний, индуцируемых радиацией; 5) увеличение частоты лейкозов начинается спустя 2—4 года после воздействия радиации. Максимум заболеваний отмечен спустя 6—8 лет после воздействия радиации, затем до 25-летнего периода уменьшалась частота индуцированных лейкозов; 6) солидные опухоли имели минимальный латентный период, равный примерно 10 годам, а некоторые из них появлялись лишь по достижении облученным организмом того возраста, когда обычно появляются спонтанные опухоли; 7) возраст человека в момент облучения является самым важным фактором организма, влияющим на последующую вероятность индукции злокачественного заболевания; 8) процент увеличения злокачественных заболеваний в расчете на дозу 0,01 Гр неодинаковый для всех типов заболеваний. Некоторые виды заболеваний, спонтанно возникающие с высокой частотой (рак толстой кишки), обладали относительно низкой индукцией при воздействии радиации, в то время как другие, возникающие спонтанно сравнительно редко, показали высокую индукцию их радиацией (рак щитовидной железы); 9) кривые доза—эффект имели часто линейный характер, но также наблюдали и кривые с оптимумом дозы для индукции злокачественных заболеваний, т. е. с неэффективным облучением при высоких дозах, что могло быть связано как с гибелью наиболее радиочувствительных клеток, так и (или) с влиянием факторов, определяющих выраженность заболевания. Последние обобщения Национальным институтом рака США материалов последствии ядерных взрывов в Хиросиме и Нагасаки выявили также много нового. В дополнение к ранее сообщавшимся видам злокачественных заболеваний, таким, как лейкоз, рак щитовидной железы, молочной железы и легких, теперь надо добавить еще рак пищевода, желудка, мочевых органов и лимфомы как последствия воздействия ионизирующей радиации в событиях 1945 г. Обоснованность доказательств радиационной природы этих заболеваний возросла. Лейкоз был самой частой радиационной причиной гибели человека по сравнению с другими видами злокачественных заболеваний, имевших место у жителей Хиросимы и Нагасаки, подвергшихся воздействию повреждающих факторов атомного взрыва. За 24-летний период от лейкоза погибло людей больше, чем от других злокачественных заболеваний, вместе взятых. Максимум лейкозов наблюдался в период 1950—1954 гг., т. е. через 5—10 лет после радиационного воздействия. В течение 1970—1974 гг., т. е. спустя 25—30 лет после взрывов атомных бомб, еще отмечались случаи радиационного лейкоза. Однако вероятность их возникновения стала существенно ниже, чем, например, вероятность возникновения рака органов пищеварения у пострадавших жителей Хиросимы и Нагасаки. Среднее превышение частоты лейкозов в Хиросиме над этим же показателем в Нагасаки (примерно в 2—3 раза выше) связывают с более выраженным лейкозогенным эффектом нейтронов, которых в ионизирующей радиации было больше в Хиросиме — до 30% в радиационной дозе. При теоретическом анализе кривых доза— эффект допускается вид кривой с оптимумом дозы, превышение которой уменьшает канцерогенный эффект. На риск канцерогенного действия радиации влияет целый ряд факторов во время облучения и в последующий период, в частности возраст и состояние организма. Считается также, что состояние медицинского обеспечения, санитарии, питания в пострадиационный период являются факторами, влияющими на риск злокачественного заболевания радиационной этиологии. В результате анализа рака молочной железы радиогенного происхождения при малых дозах радиации установлено, что интервал времени между облучением человека и появлением клиники заболевания может существенно изменяться гормональным статусом и другими зависящими от возраста организма факторами, а не дозой воздействия. Радиационно вызванный рак молочной железы не выявляется, пока организм не достигнет возраста, при котором начинает появляться спонтанный рак молочной железы. Повышение дозы воздействия не ускоряет развитие рака молочной железы. Интересно и другое. Аденомы легких и некоторые другие опухоли легких возникают спонтанно сравнительно часто. Однако при действии радиации их частота обычно уменьшается. Оптимум радиационных доз для индукции ряда опухолей близок к тем дозам, которые начинают вызывать деструктивные процессы. На примере рака молочной железы человека это показано особо четко. По статистическим данным, среди всех гемобластозов (злокачественные заболевания крови) лейкозы и эритремии составляли 61—65%, а лимфомы — 35—37%. Из них эритремии (краснокровие) возникали крайне редко (всего 4%), или примерно в 6 раз реже, чем острые и хронические миело- и лимфолейкозы (белокровие). Характерно значительное преобладание лейкозов с выраженной картиной изменений одного из ростков кроветворения. Лейкозы с выраженными изменениями двух или более ростков встречаются реже — менее 2% от общего числа гемобластозов. По другим данным, эритремия обнаруживается в 10—20 раз реже острого лейкоза. Удивляет полиэтиологичность лейкозов. В числе возможных причин их возникновения называют радиационный фактор, химические канцерогены, вирусы, дисгормональные расстройства, злоупотребление лекарствами, нейрогенные дисфункции, стресс, хронические инфекционные процессы, хронические раздражения и др. Сопоставление клинических и экспериментальных данных о возможных классах канцерогенов показало их огромное разнообразие. Перечислим некоторые из них: 1) химические стимулы — органические соединения (нитрозоамины, бензпирены и др.), природные соединения (афлатоксины и др.), гормоны, нуклеиновые кислоты, неорганические соединения (асбест, соединения свинца, бериллия и др.); 2) физические стимулы — ионизирующие излучения, механическое раздражение (хронические раны и ожоги), полимерные включения; 3) внешние биологические стимулы — некоторые паразиты и бактерии, опухолевые вирусы (ДНК- и РНК-вирусы). Описаны также лейкозы после применения иммунодепрессантов, цитостатиков, некоторых антибиотиков, пирамидона. Известны лейкозы на почве бензольной интоксикации. Кроме того, имеется определенная предрасположенность наследственного характера к злокачественным заболеваниям. Имеются интересные данные о том, что соотношение частот заболеваемости хроническим лимфолейкозом у взрослых европейцев (около 30% всех лейкозов) и у китайцев и японцев (1,5%) не изменяется, несмотря на иммиграцию населения, что указывает на большую роль наследственных конституционных факторов. Имеется связь между неспецифической устойчивостью детей и предрасположенностью их к заболеванию острым лейкозом. Среди различных наследственных заболеваний человека и осложнений их определенными видами злокачественных заболеваний лейкоз, включая злокачественную лимфому, наблюдается при ряде наследственных заболеваний. Примерно 9% из 2000 наследственных заболеваний человека связано со злокачественными новообразованиями. Однако из общего количества злокачественных заболеваний лишь 5—10% случаев имеют наследственную природу. Остальные случаи связаны с влиянием множества эндогенных и экзогенных факторов. Подверженность к злокачественному заболеванию определена как комбинация генетической чувствительности и воздействия факторов окружающей среды. Среди внешних для организма факторов рассматриваются и такие, как характер питания и стрессовая обстановка. В специальных обзорах обсуждаются различные вопросы влияния пищи и расстройств питания, включая те, которые связаны с алкоголизмом и курением, на возникновение различных форм рака. Пища как один из факторов внешней среды может влиять на частоту рака, изменяя в организме состояние отдельных биохимических систем и содержание гормонов. 80—90% случаев заболеваний раком в США связано с воздействием на человека многообразных факторов внешней среды. Среди них на долю факторов питания приходится до 60% У женщин и до 40% у мужчин. Имеются обобщающие работы о роли стресса в развитии малигнизации. Проведен анализ большого числа работ по оценке отклонений в состоянии центральной нервной системы или ее особенностей у онкологических больных, включая лейкозных. Изучались также психофизические и психосоциальные особенности больных и делались попытки понять возможный механизм связи стресса и заболевания. Стресс независимо от причины, его вызвавшей, может существенно влиять на развитие различных заболеваний, включая неопластические, хотя механизм такого влияния еще не ясен. Предполагается, что стресс индуцирует серию биохимических изменений в организме, опосредованных через нейроэндокринную систему. Имеются данные, что направленным воздействием на центральную нервную систему можно вызвать определенные изменения в иммунологической реактивности и тем самым влиять на условия возникновения и развития опухолей. Р. Е. Кавецкий указывал, что в создании предопухолевой предрасположенности могут большую роль играть иммунодепрессивные состояния, вызванные большой группой факторов, среди них наследственное, возрастное и постинфекционное снижение реактивности. Существование механизмов эндокринного канцерогенеза не вызывает сомнения. Естественное или искусственное значительное и длительное усиление действия гормонов-стимуляторов пролиферативной активности на ткани-мишени приводит к малигнизации клеток этой ткани. Так, избыточное и длительное выделение гипофизом тиреотропного гормона может вызывать злокачественные опухоли щитовидной железы. Нарушения в психофизиологических функциях центральной нервной системы, вызванные стрессом, через нейроэндокринные регуляции ингибируют или извращают функции иммунной системы. Вследствие этого ослабляется иммунологическая защита организма, что способствует возникновению инфекционных и злокачественных заболеваний, а также аутоиммунных и аллергических расстройств. Предполагается, что в механизмах таких взаимосвязей большую роль играют реакции макрофагов на гормональный дисбаланс. Прямыми экспериментальными данными показано, что у животных с пониженной или резко повышенной реактивностью гипофизарно-надпочечниковой системы значительно ускоряется развитие опухолей молочных желез при введении химических канцерогенов. У крыс, резистентных к введению канцерогена, реактивность коры надпочечников и ее андрогенная и глюкокортикоидная функции в предопухолевом периоде остаются без существенных отклонений. У животных с относительно высокими показателями андрогенов и глюкокортикоидов в организме и нормальной реактивностью коры надпочечников опухоли или совсем не возникают на введение химического канцерогена, либо возникают в более поздние сроки и являются доброкачественными (фибромы, фиброаденомы). На частоту возникновения злокачественных заболеваний влияют и конституциональные особенности организма. Еще Галену было известно, что женщины-меланхолики заболевают раком чаще, чем сангвиники. Иммунологии канцерогенеза посвящено много работ, так же как и рассмотрению процесса канцерогенеза как проблемы биологии развития. Следует, однако, иметь в виду, что система иммунологической защиты на самом деле начинает активно действовать лишь в том случае, когда опухолевые клетки уже имеются в организме, т. е. после того, как уже произошла малигнизация какой-то группы клеток. Поэтому система иммунитета очень важна для ограничения размножения клеток, для предупреждения их прогрессии, но она не играет сама ведущей роли в истинном индуцировании малигнизации клеток. При врожденных дефектах иммунной системы частота злокачественных заболеваний возрастает в сотни раз. Следует обратить внимание на тот факт, что в популяции с повышенным риском канцерогенеза или лейкозогенеза в реальных условиях заболевает лишь незначительная ее пасть. Организм абсолютного большинства людей, находящихся в таких же условиях канцерогенного и лейкозогенного соприкосновения, справляется с неблагоприятным фактором и остается здоровым. В эксперименте у нормальных животных рост прививной опухоли возникает только в том случае, если было введено в организм более 1000 живых опухолевых клеток, хотя каждая из них способна дать начало опухолевому процессу. Из изложенного следует важность исследования состояния организма, определяющего возможность или невозможность возникновения лейкоза в условиях соприкосновения организма с лейкозогенными условиями. Следует иметь в виду, что основная масса опухолей человека связана с принципиально устранимыми факторами поведения и образа его жизни. Большое внимание по-прежнему уделяется вирусной этиологии лейкоза и рака. На этом направлении исследований достигнуты крупные успехи в экспериментах, особенно на мышах и курах. Вместе с тем пока не удалось преодолеть все сложности в изучении возможной вирусной природы опухолей и лейкоза человека. Имеющиеся у приверженцев этого направления исследований данные не могут объяснить весь комплекс эпидемиологических, клинических, патофизиологических, цитологических и биохимических данных о лейкозной болезни человека. Необходимо учитывать невозможность перевивки лейкозов у человека при случайном переливании крови от больных к здоровым, не передается лейкоз от больной матери к плоду и, наоборот, матери от ребенка с врожденным острым лейкозом. Все это говорит против вирусной этнологии лейкоза человека. В связи с изложенным лойкозогенные вирусы представляются не столько возможными этиологическими, сколько патогенетическими агентами среди других таких же факторов, способных вызвать развитие лейкоза. Однако исследования с вирусами позволили открыть новую страницу в изучении возможных молекулярных механизмов канцерогенеза. Большие надежды ныне возлагаются на так называемые вирусные онкогены, играющие, по мнению Ф. Л. Киселева и других исследователей, ключевую роль в процессах трансформации нормальных клеток в опухолевые. Но здесь встретились непреодолимые трудности: открыто слишком много вирусных онкогенов — более 20, такие же онкогены выявлены и в нормальных клетках, причем белки онкогенов ретровирусов и клеток не отличаются, по существу, ни по аминокислотной последовательности, ни по биохимическим, ни по иммунологическим свойствам, часть из них относится к регуляторам роста нормальной клетки и участвует в нормальных процессах пролиферации и дифференцировки. Предполагается, что в связи с нестабильностью генома под влиянием разных причин может осуществляться усиленная экспрессия (активация) онкогенов — тех генетических структур, которые способны трансформировать нормальные клетки в злокачественные в определенных условиях. Детальные исследования молекулярной структуры онкогенов ретровирусов и клеточных онкогенов показали их идентичность, так же как и идентичность их белков. Более того, они обнаружены и в геноме нормальных клеток, где их функция в основном связана с регуляцией роста клеток и процессов пролиферации и дифференцировки. К тому же для ряда онкогенов четко показана и неспецифичность их в вызывании определенных форм рака. Так, вирус острого лейкоза птиц вызывает наряду с лейкозом и опухоли совсем другого типа в печени и почках. В связи с изложенным возникает сомнение в том, являются ли онкогены первичным механизмом малигнизации клеток, или активация их является вторичной, производной от клеточных изменений и возникшей нестабильности генома, о которых речь будет позднее. Говоря об экспериментальном изучении возможных механизмов образования злокачественных заболеваний, следует иметь в виду следующие обстоятельства. В настоящее время довольно успешно исследуются вопросы злокачественной трансформации клеток в культуре. Однако необходимо отметить, что существование популяции клеток в организме и в культуре принципиально отличается. В последнем случае клетки находятся вне влияния тканевого и организменного окружения. Ю. М. Васильев и И. М. Гельфанд [1981] пишут, что «после перенесения фибробластов и эпителиальных клеток в культуру свойства этих клеток могут претерпевать серию генетически стойких изменений как спонтанных, так и индуцированных. Эта эволюция свойств в культуре обладает рядом черт сходства с неопластической эволюцией клеток в организме» (с. 40). Следовательно, сам факт переноса клеток в культуру заставляет их приобретать определенные первоначальные черты, приближающие их к злокачественной трансформации (изменение формы клеток и их поверхности, появление способности размножаться в условиях, в которых ранее они не размножались, изменение чувствительности к росторегулирующим веществам и др.). Клетки, размножающиеся в культуре,— это клетки, уже вступившие на путь, ведущий к трансформации. Надо быть очень осторожным при попытках использовать данные, полученные в культуре, для исследования первичных механизмов злокачественной трансформации клеток в организме. То же самое и с оценкой результатов экспериментов на животных. Абсолютное большинство опытов проводится или на специально подготовленных, или генетически дефектных животных, или при очень далеких условиях перевивки лейкозогенного фактора от условий, в которых находится заболевший человек. Поэтому в настоящее время ряд исследователей основное внимание уделяют попыткам понять возможные механизмы канцерогенеза для реальных условий окружения человека. При этом подчеркивается необходимость изучения прежде всего предраковых состояний. Наиболее подробно в клиническом и экспериментально-морфологическом аспекте Л. М. Шабадом изучено предраковое состояние. Он различает следующие друг за другом четыре стадии опухолевого процесса: 1) диффузная или неравномерная гиперплазия (т. е. разрастание ткани за счет клеточного размножения); 2) очаговые пролифераты (т. е. узлы таких разрастаний ткани); 3) доброкачественная опухоль (не имеющая тенденции к безудержному росту); 4) злокачественная опухоль. Его взгляд на роль предварительной гиперплазии с очаговой (узелковой) пролиферацией подтвержден последующими работами. Следовательно, вопросы нарушения роста и размножения клеток, нарушения дифференцировки клеток — приобретения ими специфических функций, необходимых организму, должны исследоваться в первую очередь. Особое внимание при этом надо уделять тем изменениям, которые являются характерными для реального злокачественного заболевания человека, в частности лейкоза.Некоторые клинические особенности лейкоза
Лейкоз относится к гемобластозам, т. е. к группе опухолей, возникающих из кроветворных клеток. Лейкоз — это гемобластоз, при котором костный мозг повсеместно заселен опухолевыми клетками. Термин лейкемия менее целесообразен но двум причинам: 1) к лейкозам относятся опухоли, состоящие не только из лейкоцитов, но и из других клеток (эритрокариоцитов или мегакариоцитов), и 2) появление в крови избытка лейкоцитов не обязательно в начале болезни. Кроме лейкозов, в группу гемобластозов входят гематосаркомы — солидные опухоли, возникшие вне костного мозга из кроветворных бластных клеток. Лимфомы могут состоять из зрелых лимфоцитов или образуются разрастаниями, идентичными лимфатическому узлу. Клетки гематосарком и лимфом могут распространяться по системе кроветворения и поражать костный мозг. Тогда они не отличаются от острого лейкоза или хронического лимфолейкоза. Практика показала, что диагноз острого лейкоза может быть уверенно установлен только по обнаружению в периферической крови или в костном мозге бластных опухолевых клеток. Сколько-нибудь характерного начала со стороны клиники или жалоб больного, свойственных острым лейкозам, найти не удается. Клинические признаки лейкоза описаны достаточно хорошо только для той стадии развития заболевания, когда больной обращается к врачу, как правило, по поводу не специфических для лейкозов жалоб. В большинстве случаев к этому времени у больного уже определились специфические изменения в крови и клинические симптомы, позволяющие поставить диагноз острого или хронического лейкоза определенной формы в соответствии с принятой классификацией их, основанной на выделении ведущего ростка кроветворения и стадии созревания клеток в нем, затронутых болезнью. Большое значение в современной диагностике лейкозов имеют и определенные изменения хромосомного аппарата.Отдельные особенности течения развитого лейкоза
В пестрой и многообразной картине каждой из форм лейкозов все же можно выделить некоторые особенности, представляющие интерес для последующего обсуждения. Весьма часто можно наблюдать неодинаковую выраженность изменений содержания зрелых форм различных ростков в периферической крови. Преобладание числа зрелых форм одного ростка может сопровождаться уменьшением в крови зрелых форм других ростков кроветворения. Так, при остром миелолейкозе нередко могут наблюдаться явления анемии, тромбоцитопении, а при хроническом лимфолейкозе — анемии, тромбоцитопении, гранулоцитопении. Среди хронических миелоидных лейкемий иногда различают эозинофильную, базофильную и нейтрофильную лейкемии. При ряде форм лейкозов (острый и хронический миелобластный и хронический лимфолейкоз) может отмечаться увеличение селезенки и реже печени. В начале заболевания хроническим миелолейкозом селезенка увеличена у 85% больных. Продолжительность жизни эритроцитов и лейкоцитов, особенно при хронических формах, сокращена. При этом обнаружена прямая корреляция между увеличением размера селезенки (гиперспленизм) и сокращением продолжительности жизни эритроцитов. Берлин и соавторы, обобщая сведения о продолжительности жизни эритроцитов при различных заболеваниях крови, пришли к выводу, что 2/3 больных хронической миелоидной лейкемией имеют укороченную продолжительность жизни эритроцитов. При этом эритроциты, перелитые от здорового человека в кровь таких больных, также имели укороченную продолжительность жизни. Авторы делают вывод, что укорочение продолжительности жизни эритроцитов у этих больных не связано с качеством эритроцитов, а определяется внеклеточными для них факторами. Сокращение продолжительности жизни эритроцитов (а также лейкоцитов) нередко связано с развитием аутоиммунного гемолиза, например при хроническом лимфолейкозе, и может проходить и без увеличения селезенки и даже без классических признаков усиления гемолиза. При лейкозах, сопровождаемых явлениями гиперспленизма, удаление селезенки оказывает хотя и временное, но значительное клиническое улучшение. При этом продолжительность жизни эритроцитов существенно возрастает и приближается к нормальной. На основании эффекта удаления селезенки И. А. Кассирский и Г. А. Алексеев делают заключение о важной роли селезенки в ускоренном разрушении эритроцитов у больных миелолейкозом, у которых в период обострения в 80% случаев продолжительность жизни эритроцитов укорочена. Следовательно, одной из важных особенностей течения лейкоза является вовлечение в патологический процесс эритроидного ростка костного мозга, изменения в котором имеют большое прогностическое значение. В классических работах, вышедших до широкого применения кортикостероидных препаратов и цитостатических иммунодепрессантов, более определенно, чем в гематологических руководствах последнего времени, оценивалось значение изменений красной крови в миело- и лимфолейкозах для прогноза заболеваний, а также сама возможность выздоровления. Прежде считалось, что для ранних этапов острого лейкоза типично развитие анемии. Однако позднее сложилось убеждение, что симптом анемии, во всяком случае для острого лейкоза, может долго не выявляться. Важнейшей задачей лечения лейкозов И. А. Кассирский и Г. А. Алексеев [1970] считают сохранение и восстановление красного кроветворения и борьбу с анемией (в том числе при миелоидном и лимфоидном лейкозах), а критерием положительного лечебного эффекта — улучшение в первую очередь состава красной крови. В старых руководствах лейкоз однозначно считался неизлечимым заболеванием, летальный исход которого можно было лишь отдалить. Случаи самопроизвольного или под влиянием терапии излечения считалиськазуистическими, и правильность диагноза ставилась под сомнение. С введением в практику лечения лейкозов кортикостероидных и специальных цитостатических препаратов взгляд на возможность излечения от лейкоза принципиально изменился. Появление полных гематологических ремиссий, исчисляемых многими месяцами и годами, с восстановлением физиологической регенерации крови позволило поставить вопрос о принципиальной и практической обратимости острого лейкоза. В отношении хронических лейкозов это стало понятным намного раньше. В связи с изложенным изменилось и отношение к случаям самопроизвольного излечения от лейкоза или возникновения спонтанных и длительных ремиссий. Эти сведения стали серьезно изучаться. И. А. Кассирский и Г. А. Алексеев [1970] собрали имевшиеся в литературе сведения о самопроизвольных ремиссиях, которые возникли у 104 больных острым лейкозом, в том числе у 67 из них причиной ремиссии были нагноения, у 35 — инфекция, у 2 — пневмония. Имеются отдельные случаи очень длительных ремиссий острого лейкоза, вызванные инфекцией (до 10 лет) и нагноениями (до 12 лет). Описаны два случая ремиссии острого лейкоза до 1—3 лет, вызванные беременностью. Однако имеются и другие, более многочисленные примеры отягощающего действия инфекции, которая является главной непосредственной причиной смертельных исходов при лейкозах. У обезьян как видовая особенность течения гемобластозов отмечено сравнительно частое возникновение спонтанных ремиссий, иногда до 4 лет и более, т. е. практически полное самовыздоровление. В литературе описано около 600 случаев ремиссий и самоизлечения от рака под влиянием лихорадки, инфекции, вакцинации, гормональных изменений, хирургических операций и сделан вывод, что это связано с изменением взаимоотношений между организмом и опухолевым процессом. И. А. Кассирский и Г. А. Алексеев [1970] приводят сведения и о том, что имелись случаи, когда больных хроническим лимфолейкозом выписывали из стационара как безнадежных, однако у них в дальнейшем исчезали клинические и гематологические проявления заболевания. Влияние указанных факторов на возникновение спонтанных ремиссий и самовыздоровления при лейкозах, по-видимому, обусловлено воздействием их на собственные физиологические защитные силы организма с активизацией их, и в первую очередь активизацией механизмов иммунологической защиты. Имеется прямая связь между частотой возникновения неоплазм и иммунологическим статусом организма. Для нас существенно, что концепция иммунологического надзора привлекает основное внимание не к самому опухолевому процессу, а к состоянию организма, его защитных сил. Таким образом, в отдельных случаях лейкоз не является заболеванием с неотвратимо неблагоприятным исходом. В тяжести течения и в исходе заболевания значительную роль играют собственные физиологические защитные силы организма. Сохранение избыточной или извращенной функции их при некоторых формах лейкоза предполагает возможность подхода к лейкозам как к своеобразным болезням компенсации.Клинические проявления предлейкоза
Особое внимание в настоящее время уделяется начальным проявлениям лейкоза и предлейкозным состояниям, которые, как правило, остаются вне наблюдения специалистов, но могут быть определены в ряде случаев ретроспективно. В последнее десятилетие укоренился термин «предлейкозный синдром» для характеристики состояния организма перед развитием безусловно специфических для лейкоза клинических проявлений. Однако оказалось очень трудным отделить начальные проявления лейкоза от тех проявлений, которые можно отнести к предвестникам возможного, но не обязательного развития в последующем лейкоза. К проявлениям предлейкозного синдрома относят прежде всего явления анемии, которая может быть разного типа. Эта анемия сопровождается часто сокращением продолжительности жизни эритроцитов и увеличением селезенки. Иногда отмечаются изменения размеров и формы эритроцитов. Часто наблюдается гиперпластичный костный мозг с преобладанием красного ростка. В периферической крови, кроме уменьшения числа эритроцитов, нередко обнаруживают уменьшение числа лейкоцитов или тромбоцитов и тогда говорят о явлениях цитопении. В отдельных случаях отмечается сохранение числа эритроцитов и гемоглобина на уровне верхних показателей нормы при нестабильности числа лейкоцитов или уменьшении их и некотором увеличении числа ретикулоцитов. Может развиваться типичная миелопролиферативная реакция костного мозга, неотличимая от реакции, возникающей по другим причинам. Появление дефектов созревания предшественников эритроцитов, гранулоцитов или моноцитов или дефектов в мегакариоцитах при нормо- или гиперпластичном костном мозге обычно относят к начальным признакам лейкоза, хотя и они не являются абсолютно надежным свидетельством начавшегося развития лейкоза. М. Саарни и Дж. Линман в 1973 г. обобщили литературные сведения о 143 случаях достаточно надежного диагноза «предлейкозный синдром». Этот диагноз ставился редко. Так, в одной группе из 322 больных острым лейкозом достаточных сведений для ретроспективной постановки диагноза «предлейкозный синдром» было только в 21 случае, когда острый лейкоз развился спустя 3—39 мес. В другой группе из 580 больных ни у одного больного не оказались достаточными ретроспективные данные для рассмотрения вопроса о наличии или отсутствии у них симптомов предлейкозного синдрома. У тех 143 больных, у которых можно было поставить диагноз «предлейкозный синдром», наблюдались разнообразные, но совершенно неспецифические проявления; среди них в 10—15% случаев — увеличение селезенки и печени, инфекция. Анемия изолированно или в сочетании с нейтропенией и тромбоцитопенией в различных комбинациях была в большинстве случаев. Отмечались нередко изменения размеров и формы эритроцитов. Однако в большинстве случаев наблюдали нормохромные эритроциты. В небольшом числе в крови появлялись ядерные эритроциты. Число ретикулоцитов — в норме или слегка увеличено. В большинстве случаев костный мозг был гиперплазирован (увеличен в размерах) в основном за счет красного ростка с возникновением в ряде случаев мегалобластических изменений — появлением наиболее ранней морфологически различаемой клетки. Гранулоцитарный отдел костного мозга обычно характеризовался смещением влево гранулоцитарной формулы, отмечали умеренное увеличение ранних предшественников и запрет на созревание клеток. Выживаемость меченных 51Cr эритроцитов или нормальна, или сокращена. Фетальный гемоглобин может быть увеличен. Уровень в плазме крови витамина B12 и солей фолиевой кислоты не уменьшен, и терапия этими препаратами неэффективна. Продолжительность периода от постановки диагноза «предлейкозный синдром» до развития лейкоза — чаще всего от 6 мес до 5 лет, но может быть и значительно больше, например до 20 лет. Авторы представили и результаты собственных клинических наблюдений. Из 132 больных с обостренной миеломопоцитарной лейкемией только в 34 случаях было достаточно данных для детального анализа состояния перед развитием лейкоза. Среди них увеличение селезенки или печени отмечено у 10 человек, анемия в сочетании с лейкопенией и тромбоцитопенией или без них — у абсолютного большинства (29 больных), нормальный или несколько увеличенный процент гемоглобина — у 25 человек, изменения размеров или формы эритроцитов — у всех, появление ядерных эритроцитов в крови — у 23, гиперплазия костного мозга — у 26 (гипоплазия лишь у одного), эритроцитарная гиперплазия — у 17 больных (эритроцитарная гипоплазия — лишь у трех). Среди причин обращения к врачу в пяти случаях — инфекционные процессы. В эритроцитарном отделе костного мозга нарушения созревания предшественников выявлены у 30 человек, столько же — в других отделах. В заключение авторы указывают, что приведенные материалы по предлейкозным симптомам многие относят к заболеваниям, не несущим плохой прогноз злокачественной болезни, например к рефрактерной анемии, сидеробластической анемии, апластической анемии, пиридоксин-связанной анемии, пароксизмальной гемоглобинурии, гиперспленизму. Имеются обобщенные данные о 131 случае предлейкоза. В 114 случаях (87%) встречалась анемия в чистом виде (в 30 случаях) или в сочетании с лейкопенией, тромбопенией или панцитопенией. Лейкопению наблюдали лишь в восьми случаях (6%), и еще реже только тромбопению — в пяти случаях (менее 4%). Следовательно, наиболее частый гематологический синдром в предлейкозный период — анемия в различных ее формах. Имеются данные, что у больных арегенеративной или рефрактерной анемией с гиперплазией костного мозга и патологическим накоплением промежуточных сидеробластов более чем в 50% случаев болезнь заканчивается острым лейкозом. Обобщая данные наблюдений других авторов о предлейкозном состоянии детей, у которых в дальнейшем развилась острая лимфобластная лейкемия, можно считать, что во всех случаях у них регистрировалась анемия гипопластического типа. В этот период никаких оснований для диагноза лейкоза не было. Анемия проходила сама или под влиянием преднизолона. Таким образом, ведущие события в период, предшествующий появлению симптомов, достаточных для постановки диагноза «лейкоз», связаны с нарушениями в системе красной крови. Симптомы описанного выше предлейкозного синдрома относятся к проявлениям патологического характера. В то же время для наиболее изученной по этиологии форме лейкоза — радиационного лейкоза известен длительный период полного клинического благополучия от воздействующего лейкозогенного агента (радиации) до появления симптомов предлейкозного синдрома. Этот период может исчисляться многими годами и даже десятилетиями. Так у лиц, благополучно перенесших острый период после воздействия радиации в Хиросиме, через 2 года в периферической крови никаких отклонений от контрольной группы не обнаружено. В дальнейшем среди этих людей отмечены случаи заболевания лейкозом радиационной этиологии, т. е. было длительное клиническое и гематологическое благополучие перед появлением начальных признаков радиационного лейкоза.Особенности эритремии и возможность общей гемобластозной болезни
Следует также остановиться на более доброкачественной форме гемобластоза — эритремии, которая встречается очень редко. Болезнь характеризуется увеличенным содержанием числа эритроцитов и их массы, циркулирующей в крови, увеличением селезенки, гиперпластичным костным мозгом. Отмечают, что переход эритремий в острый лейкоз происходит редко. Под влиянием радиотерапии (рентгеновское облучение или радиоактивный фосфор) вероятность перехода в острый лейкоз возрастает примерно в 10 раз. Чаще всего происходит переход в эритромиелоз с явлениями панмиелоза и реже — в хронический миело- или лимфолейкоз. Обращает на себя внимание наличие переходных форм заболеваний между «чистым» миелоидным или лимфоидным лейкозом (белокровием) и типичной эритремией (краснокровием). Клиницисты давно обратили на это внимание. Г. А. Алексеевым в 1950 г. описаны клинические случаи постепенного развития болезни от типичной эритремии (краснокровия) к хроническому миелолейкозу (белокровию). Он рассматривал их как последовательные стадии одного болезненного процесса и обращал внимание на случаи эритролейкемий, сочетающих в себе одновременно признаки эритремии и хронического миелолейкоза. Эритролейкемию можно рассматривать как переходную стадию в развитии острой миелобластной и миеломоноцитарной лейкемий. Обобщив собранный материал с учетом приведенных экспериментальных материалов, далее мы излагаем сложившиеся представления об основных периодах развития такой общей гемобластозной болезни в типичный белокровный лейкоз в виде упрощенной схемы (табл. 1).Таблица 1. Основные периоды клинического состояния при общей гемобластозной болезни после воздействия лейкозогенного фактора (радиации и др.)

Первый период — период клинического и гематологического благополучия (период А). После воздействия лейкозогенного агента (например, радиации) отмечается длительный период (измеряемый годами и иногда десятилетиями) полного благополучия. Отсутствуют клинические и гематологические расстройства при использовании традиционных клинико-лабораторных методов обследования больных. Нет никаких оснований предполагать возможность каких-либо отдаленных последствий у абсолютного большинства лиц, подвергавшихся воздействию лейкозогенного фактора. Далее возможны три исхода: а) сохранение полного клинического и гематологического благополучия без каких-либо отдаленных последствий; б) развитие неспецифических по отношению к лейкозу потенциально предпатологических состояний, выявляемых в первый период времени лишь при функциональных нагрузках или при использования специально разработанных методик исследований. У большинства лиц наблюдается обратное развитие в сторону полного клинического и гематологического благополучия без каких-либо последствий. При неблагоприятном течении этих состояний могут начать проявляться некоторые не ярко выраженные отклонения в гематологических показателях, однако с благоприятным дальнейшим исходом; в) развитие такого же неспецифического по отношению к лейкозу потенциально предпатологического состояния, переходящего в следующий период предлейкозного состояния. Это характерно лишь для небольшой части лиц, находившихся в лейкозогенной обстановке. Второй период — период предлейкоза (период Б). В этот период медленно нарастают изменения со стороны показателей периферической крови и костного мозга. Ведущее значение имеют изменения со стороны красной крови. Эти изменения вначале носят скрытый характер, выявляемый лишь функциональными нагрузками или специальными методиками исследований. Затем они приобретают клинически заметные формы — неустойчивость содержания лейкоцитов, гранулоцитов и тромбоцитов в крови, уменьшение их числа по одному или нескольким росткам, реже панцитопения. В костном мозге соотношение бластных форм красного и белого ряда смещается чаще в сторону красного ряда. Все эти изменения не дают еще четкой картины для уверенной постановки диагноза какого-либо заболевания. При благоприятном течении изменений, что наблюдается у большинства лиц, каких-либо последствий не ожидается. В организме восстанавливается полное клиническое и гематологическое благополучие. Однако при неблагоприятном течении возникших изменений эти состояния могут постепенно перерастать в клинически выраженную форму, достаточную для постановки диагноза анемии. В этот период ускоренно разрушаются эритроциты в селезенке и печени. В костном мозге чаще развивается компенсаторная гиперплазия с преобладанием пролиферативных процессов в красном ростке. К анемии может присоединяться снижение числа клеток других ростков крови. Но ведущими и наиболее стойкими симптомами являются изменения со стороны красной крови. Анемия практически трудно курабельна, но может проходить и без лечения. Однако она может перерастать и в следующий период общей гемобластозной болезни. Иногда рассматриваемый период не имеет четко выраженных изменений и остается вне внимания исследователя, когда он впервые обращает внимание лишь на последующие периоды. Третий период — период эритремиоидного состояния (период В). В этот период происходит внешне благоприятное изменение со стороны красной крови. Явления анемии сглаживаются. Число эритроцитов в крови восстанавливается до нормы или слегка превышает ее. Аналогичное положение с содержанием гемоглобина. Такое состояние можно рассматривать как следствие успешного развития реакций компенсации в ответ на недостаток эритроцитов и ускоренное разрушение их. Однако компенсация крайне неустойчива и может переходить в сверхкомпенсацию, когда появляется избыток эритроцитов в периферической крови (это может завершаться эритремией как самостоятельным заболеванием), или в срыв компенсации, когда появляется панцитопения. Костный мозг может быть разным: от гиперплазированного по красному ростку до тотальной гиперплазии (что бывает реже) и до гипопластического состояния (что встречается очень редко). Этот период может остаться для клинициста и незамеченным, если не проводятся исследования костного мозга, а изменения в крови не достигли серьезных отклонений. При отсутствии исхода в эритремию неустойчивое состояние кроветворения не может оставаться неизменным длительные сроки. При тотальной гиперплазии по красному и белому росткам может начаться следующий период развития общей гемобластозной болезни. Четвертый период — период переходных клинических форм (период Г). Этот период также неустойчивый. Он характеризуется промежуточным состоянием кроветворения, когда нет выраженного преобладания клеток одного из ростков, когда костный мозг одновременно гиперплазирован по красному и белому росткам, но может отмечаться и гипоплазия по всем росткам. Предпочтительное производство клеток красной крови начинает сменяться, но еще не сменилось предпочтительным производством клеток другого ряда. В редких случаях такое промежуточное состояние проявляется клиническим выходом в форме такого самостоятельного заболевания, как эритромиелоз. Клетки крови продолжают ускоренно разрушаться в крови. Однако чаще именно на этой стадии завершается переключение примата производства с красного ростка на миелоидный росток (гранулоцитарный, моноцитарный или на производство их общего предшественника) или на лимфоцитарный росток. Соответственно начинает изменяться и периферическая кровь. Наблюдается ограничение пролиферативной активности красного ростка костного мозга, последующее подавление или истощение его с гиперплазией других ростков, уменьшение числа эритроцитов до нормы и ниже ее, увеличение числа клеток белой крови до нормы. Пятый период — период типичного лейкоза (период Д). Это завершающий период развития общей гемобластозной болезни в типичные «белокровные» лейкозы. Значительное увеличение числа клеток белой крови в начальный период не обязательно, в дальнейшем — характерно. Развитию лейкоза не обязательно предшествует гиперплазия костного мозга. Он может быть и истощенным вследствие предшествующего длительного периода гиперплазии. Чаще всего тот росток, из которого развивается в дальнейшем «белокровный» лейкоз, находится в угнетенном состоянии. Это первое правило лейкозной опухолевой прогрессии по А. И. Воробьеву. Содержание клеток других ростков в крови уменьшено в различной степени. Эта идеализированная схема необходима, чтобы на ее основе попытаться понять закономерную последовательность появления разнообразных симптомов. Схема поясняет периоды развития общей болезни, закономерность промежуточных выходов в клинические формы, как будто бы не имеющие прямой патогенетической связи с типичным лейкозом. В то же время схема поясняет возможность скрытого с точки зрения заметных внешних клинических симптомов прохождения заболевания непосредственно до «белокровных» лейкозов. Главное, по нашему мнению, заключается в том, что схема предусматривает длительный период неспецифических предпатологических состояний, который, как будет показано далее, позволит понять многие вопросы патогенеза лейкоза и их связь с нормальными возрастными изменениями. В реальной клинической картине правильность этой упрощенной схемы проверить трудно из-за огромного разнообразия возможных форм заболеваний и их проявлений, обусловленных, во-первых, тесной взаимосвязью всех ростков кроветворения, объединяющей происходящие в них процессы в одну общую системную гемобластозную болезнь, и, во-вторых, клоновым характером кроветворения и воспроизводством возникших нарушений в последующих генерациях клеток, разъединяющих процессы клеток одного ростка от клеток других ростков и создающих в ряде случаев конкурентные отношения между ними за преимущественную продукцию своих клеток. Конкретная форма заболевания определяется не только местом первоначальной бластной трансформации в ряду конкретных стадий созревания клеток данного ростка или их предшественников, но и возможностями развития замещающих и компенсаторно-восстановительных реакций организма. Эти реакции компенсации могут приобретать также патологический характер. В связи с этим следует обратить внимание на своеобразную группу хронических лимфолейкозов — парапротеинемических гемобластозов, отличительной чертой которых является способность вырабатывать в большом количестве иммуноглобулины — одни из важнейших факторов гуморального иммунитета организма. Кроме того, имеются большие группы незлокачественных заболеваний крови (агранулоцитозы и анемии), в патогенезе многих форм которых избыточные или извращенные реакции гуморального или клеточного иммунитета играют важную роль. В этиологии этих заболеваний инфекционному фактору, стимулирующему иммунные реакции, или химико-токсическому фактору, извращающему их, отводится важное место. Это подчеркивает необходимость исследования в механизмах возникновения предлейкозных состояний возможных избыточных и извращенных реакций компенсации.
Экспериментальные лейкозы собак
Частота спонтанных лейкозов у собак в отличие от наблюдений на мышах и крысах близка к частоте их развития у человека. Кроме того, в условиях хронического радиационного воздействия в суммарной дозе 40—70 Гр, вызывающей гибель собак через 2—6 лет после начала облучения, риск смерти этих животных от лейкоза весьма близок к такому риску у человека. В то же время естественный риск смерти от лейкоза у мышей в 10—100 раз выше, чем у человека. Из этого следует особая ценность исследования механизмов развития лейкоза и предлейкозных состояний на экспериментальной модели собак. Сведения о лейкозах у собак очень ограниченны. Вероятно, это связано и с тем, что постановка массовых и длительных опытов на собаках недоступна для широкого экспериментирования. Тем не менее некоторые характерные данные о лейкозах у собак оказались очень ценными. Наиболее подробное описание проявлений лейкоза и, что особенно важно, предшествующих ему проявлений предлейкоза приводит М. С. Лаптева-Попова. Ей удалось воспроизвести экспериментальные лейкозы у собак путем систематического воздействия малых доз рентгеновских лучей. В начальный период развития лейкоза число лейкоцитов не изменялось, и лишь за 1—2 недели до смертельного исхода содержание лейкоцитов начинало нарастать. Во всех случаях одновременно отмечали гиперхромную макроцитарную анемию. Систематические исследования крови и костного мозга позволили автору отметить определенные изменения, предшествовавшие выраженным проявлениям заболевания. За 4—7 мес до начала клинического заболевания в периферической крови стали изредка появляться единичные незрелые клетки костного мозга (миелобласты, промиелоциты). Наряду с уменьшением числа эритроцитов появлялись в периферической крови и эритробласты, иногда в значительном количестве. Число тромбоцитов резко колебалось. Интересны наблюдения автора и в более ранний период, когда еще отсутствовали в периферической крови какие-либо патологические или незрелые формы клеток. Появление в периферической крови эритробластов на фоне снижения числа эритроцитов и гемоглобина свидетельствовало о крайнем напряжении эритропоэза. В случае двухмесячного перерыва в облучении число эритроцитов повысилось с 3,5 до 5 млн/мм3 крови, повышенное количество лейкоцитов возвратилось к норме, резкие колебания в числе тромбоцитов прекратились, исчезли миелобласты, число которых в крови до перерыва в облучении доходило до 2%. Базофилы тоже исчезали. Однако через месяц после возобновления облучения появилась четкая картина подострой формы миелоидного лейкоза. Интересное наблюдение привел Е. Н. Антипенко. У одной собаки из тех, которые еще жили 27,5 мес после воздействия облучения в дозе 6,5 Гр с последующим симптоматическим лечением в течение месяца, обнаружили лимфосаркому. Эта собака при обследовании до 8 мес после воздействия радиации ничем не выделялась в своей группе. Через 8 мес все показатели периферической крови были в норме (число эритроцитов, лейкоцитов, ретикулоцитов, тромбоцитов, содержание гемоглобина, лейкоцитарная формула). Однако по величине отношения клеток миелоидного ряда к клеткам эритроидного ряда в костном мозге она значительно выделялась. Это соотношение было равно лишь 0,5, в то время как у всех остальных собак отношение превышало единицу. Повторное исследование костного мозга через 9,5 мес после облучения снова показало низкую величину этого отношения; оно стало еще меньше — 0,4. Через 1,5 года после этого обследования собака погибла от лимфосаркомы. Следовательно, в этом опыте было отмечено значительное преобладание эритроидных элементов над миелоидными в костном мозге задолго (за 1,5 года) до развития лимфосаркомы. И в этом случае норма показателей периферической крови сопровождалась существенной гиперплазией красного костного мозга в предлейкозный период. Значительный интерес представляют исследования на чистопородных гончих собаках, систематически облучавшихся гамма-радиацией в малых дозах, У них лейкоз появляется в среднем через 24—35 мес с начала облучения. Анемия обнаруживалась значительно раньше и сопровождалась глубокой тромбоцитопенией. Имеются сообщения о результатах других работ на чистопородных гончих собаках, которых систематически облучали. Из восьми облученных собак шесть погибли от лейкемии. В предлейкемический период у них отмечали анемию, изменение формы эритроцитов, появление ядерных эритроцитов, левый сдвиг гранулоцитов, увеличение числа моноцитов и др. В случае эритролейкемии за три педели до смерти в периферической крови впервые обнаружили выраженный эритробластоз. Пункция костного мозга за три недели до гибели у одной собаки показала дефекты созревания клеток эритроидного ряда с полиплоидией фигур деления, почкующимися ядрами, хромосомными мостами. У другой собаки с посмертным диагнозом «хронический грануло-мегакариоцитарный миелоз» анемия появилась еще за 13 мес до смерти. У всех собак масса селезенки и печени была увеличена. * * * Завершая данный раздел, отметим, что лейкоз относится к наиболее изученным формам злокачественных образований, по которым имеются надежные эпидемиологические данные и наиболее массовые случаи заболеваний, особенно радиационной этиологии. Выяснены некоторые особенности клинических проявлений заболевания как в период развитого лейкоза, так и в предлейкозный период. Обращено внимание на большое значение изменений со стороны красной крови и определенную связь в возникновении лейкоза с состоянием организма и неспецифическими по отношению к лейкозу изменениями в его важнейших системах. Эти изменения являются общими и предшествуют ряду разнообразных клинических форм патологии, включая возрастную, т. е. определяют состояние предпатологии.Общие черты предпатологии радиационной и нерадиационной природы
Наличие длительного скрытого периода развития радиационного лейкоза, так же как и других отдаленных радиационных последствий, заставило обратить наше внимание на более тщательное изучение того внешне клинически благополучного состояния организма, которое наступает вслед за практически полным восстановлением организма, перенесшего воздействие радиации. Радиационная модель, как известно, удобна для исследования общих закономерностей развития ряда предпатологических состояний вообще, так как клинические проявления и, по-видимому, патогенез отдаленной радиационной патологии принципиально не отличаются от патологии хронических радиационных и многих токсических воздействий. В частности, выяснилось, что в механизмах развития многих проявлений этих видов патологии важную роль играют ослабление и истощение одних и тех же компенсаторно-восстановительных реакций и искажение взаимосвязей различных систем организма, определяющих возникновение и функционирование так называемых порочных патогенетических кругов и имеющих связь с возрастными изменениями организма.Радиационные изменения кроветворения в ближайшие и отдаленные сроки
При анализе процессов восстановления гематологических показателей у человека после острой лучевой болезни (средней степени тяжести) мы обратили внимание на то, что они восстанавливаются значительно позднее клинического выздоровления организма. Восстановление цитологического состава крови обычно наблюдается через 4—6 мес и касается не всех ростков кроветворения. В первую очередь восстанавливается красная кровь, если ее показатели были снижены. Восстановление числа лейкоцитов и тромбоцитов происходит в более поздние сроки и не носит характера устойчивой нормализации. Особенно долго не восстанавливается число лимфоцитов в крови. Так, через 1,5—2 года после радиационного воздействия еще находили пониженное содержание лимфоцитов в крови человека. Следует сказать, что восстановление цитологического состава крови часто сопровождается сохранением извращенных реакций белой крови на различные воздействия: инфекции, травмы, функциональные нагрузки. Более подробно эти вопросы изучены в экспериментах. Так, в опытах И. Н. Усачевой последовательность восстановления различных ростков кроветворения у облученных собак была следующей. После воздействия рентгеновскими лучами в дозах 6 и 8 Гр через 45 сут у выживших собак еще были значительные нарушения в периферической крови и костном мозге, несмотря на внешнее клиническое благополучие. Они выражались в уменьшении числа эритроцитов, тромбоцитов, лейкоцитов и лимфоцитов и в отсутствии нормального количественного соотношения эритробластических и миелоидных клеток костного мозга. Одновременно отмечалось значительное повышение числа ретикулоцитов с последующим нарастанием числа эритроцитов периферической крови и увеличением числа кроветворных клеток в костном мозге, среди которых преобладают эритробластические. Через 4—6 мес у большинства собак наблюдалось первоочередное восстановление эритропоэза. Стойкого восстановления тромбопоэза не обнаруживали и через 18 мес. К этому сроку еще отмечали значительные колебания их числа в сторону как повышения, так и понижения. Еще медленнее шло восстановление белой крови. Даже через 12 мес после облучения в дозе 7 Гр у четырех собак из десяти обследованных в этот срок число лейкоцитов было в пределах 3—6 тыс. клеток в 1 мм3 вместо исходных 9—15 тыс. клеток. У двух собак, облученных в дозе 8 Гр и наблюдавшихся, и через 18—20 мес число лейкоцитов не достигло исходного уровня. Эти изменения в значительной мере были обусловлены недостаточностью лимфопоэза. Особый интерес представляют эксперименты Е. Н. Антипенко, специально поставленные для изучения последовательности восстановления показателей гемопоэза у собак после воздействия радиацией в дозах 4 и 6,5 Гр (часть собак при облучении в этих дозах подвергалась симптоматическому лечению). В группе выживших собак, которых облучали в дозе 4,5 Гр (ЛД 50/30), восстановление гемопоэза происходило медленно. Быстрее всего восстанавливалось число эритроцитов. С 75-го дня после облучения их количество начинало превышать исходный уровень или равнялось ему. Число лейкоцитов за весь годовой период наблюдений не достигало исходных величин. Восстановление числа лимфоцитов шло наиболее замедленными темпами, и в последние месяцы эксперимента лимфоцитов в среднем оказалось в 2 раза ниже исходного уровня. Содержание тромбоцитов было значительно снижено — в большинстве случаев больше, чем нейтрофилов. Число тромбоцитов достигло уровня 90% исходной величины лишь на 12-м месяце. При исследовании костного мозга на 4-м и 10-м месяце отношение клеток миелоидного ряда к клеткам эритроидного ряда было смещено в сторону красного ряда. Рис. 2. Типичные изменения показателей крови у собак в отдаленный период после облучения
1 — эритроциты, млн/мм3 крови; 2 — гемоглобин, г%; 3 — ретикулоциты, %; 4 — лейкоциты, тыс/мм3; 5 — нейтрофилы, тыс/мм3; 6 — лимфоциты, тыс/мм3; 7 — тромбоциты, тыс/мм3
Рис. 2. Типичные изменения показателей крови у собак в отдаленный период после облучения
1 — эритроциты, млн/мм3 крови; 2 — гемоглобин, г%; 3 — ретикулоциты, %; 4 — лейкоциты, тыс/мм3; 5 — нейтрофилы, тыс/мм3; 6 — лимфоциты, тыс/мм3; 7 — тромбоциты, тыс/мм3
Наблюдали значительное преобладание молодых форм клеток над зрелыми формами в нейтрофильном ряду, что указывало на сохранение в этом ряду сильной стимуляции к ускорению пролиферации клеток, способных к делению, в ущерб их созреванию. Собаки, облученные в дозе 6 Гр с последующим симптоматическим лечением (в течение месяца), через 8—10 мес по общеклиническим показателям не отличались от здоровых животных. Исключение составили показатели крови и костного мозга (рис. 2). Во все сроки дальнейшего исследования (до трех лет после облучения) число лейкоцитов у них было снижено на 20%, так и не достигнув исходных величин. Это снижение происходило за счет еще большего уменьшения числа лимфоцитов — на 33—50%. В то же время показатели красной крови находились в пределах нормы или даже несколько превышали ее. Следовательно, по срокам пострадиационного восстановления клетки крови располагаются в следующем убывающем порядке: эритроциты, нейтрофилы, тромбоциты и лимфоциты. Такая последовательность совпадает с разрозненными данными о последовательности восстановления клеток крови у людей, перенесших лучевую болезнь. Далее следует еще отметить, что ни у одной из наблюдавшихся собак не удалось и через 3 года после облучения выявить нормализации лимфопоэза. В то же время показатели красной крови первыми достигали нормальных величин. Эта нормализация сопровождалась заметным увеличением числа ретикулоцитов, т. е. увеличенным выбросом из костного мозга в периферическую кровь недозревших эритроцитов. Это, а также сохранившийся сдвиг отношения бластных клеток белого и красного рядов в сторону красного свидетельствует о том, что нормализация содержания эритроцитов и гемоглобина в периферической крови происходит за счет напряжения эритропоэза. По мнению многих специалистов, изменения красной крови в большинстве случаев прогностически более важны, чем белой крови. В организме отдается предпочтение восстановлению той крайне необходимой для организма дыхательной функции, какую выполняет красная кровь. Это позволило считать, что без истинной нормализации эритропоэза в преобладающем большинстве случаев трудно ожидать стабильного восстановления других ростков кроветворения и их функций, в частности иммунитета. Однако представления о состоянии эритропоэза, полученные по данным общепринятых исследований цитологического состава крови и костного мозга, не всегда могут правильно отражать состояние кроветворения. Миелограммы в ряде случаев не позволяют судить о функциональной активности всей гемопоэтической ткани в силу возможной очаговости изменений в ней и в случаях изменения плацдарма кроветворения (т. е. массы кроветворной ткани). По данным пунктатов костного мозга трудно судить о количественных изменениях в скорости кроветворения, особенно когда нет изменений в соотношениях отдельных стадий созревания клеток. Поэтому суммарную продуктивность эритропоэза определяли по включению 59Fe в эритроциты. Меченое железо включалось на стадии гемоглобинизации эритроцитов. Расчеты меченого железа, поглощенного эритроцитами 1 мл крови, 1 млн эритроцитов или 1 г гемоглобина, давали идентичные результаты. В этих условиях радиоактивность циркулирующих эритроцитов характеризовала продукцию их всей массой кроветворной ткани и поступление их в периферическую кровь. Примененная методика исключала влияние возможных часовых, суточных и ситуационных колебаний в течение указанных процессов на конечный результат, поскольку определялось интегральное поглощение 59Fe за четырехсуточный период. Изучены суммарная интенсивность эритропоэза у собак через 1; 2,5; 12 и 42 мес после облучения в дозе 4 Гр и показатели у других групп собак, подвергавшихся облучению в разных дозах. Выявлены наиболее характерные результаты для собак, у которых после перенесения острого лучевого поражения цитологические показатели крови не имели сколько-нибудь выраженных изменений или наблюдались умеренные изменения со стороны белой крови. После однократных облучений с конца первого месяца после лучевого воздействия и далее в различные сроки до 3,5 лет у многих, а в определенных условиях у большинства, собак отмечался повышенный темп эритропоэза при нормальном или несколько пониженном уровне лейкоцитов и значительно реже эритроцитов и гемоглобина. Часто, но не всегда обнаруживался ретикулоцитоз. Нередко наибольшее включение 59Fe в эритроциты наблюдалось у тех животных, у которых была более выраженная эритропения. Особенно значительное усиление темпа кроветворения (в 2 раза и более) отмечалось после облучения в дозах 2,0—3,5 Гр (дозы, близкие или несколько превышающие максимальные несмертельные, но вызывающие клинически выраженную лучевую болезнь). Повышенный темп эритропоэза при нормальной периферической крови можно рассматривать как подтверждение того, что восстановление и нормализация системы красной крови, связанной с функциями, жизненно необходимыми для существования организма, происходят в первую очередь. Отсутствие изменений в цитологическом составе крови не должно служить доказательством истинного благополучия гемоноэза. Напряжение эритропоэза, отмеченное в течение весьма длительного времени после лучевого воздействия, может быть одной из причин задержки в восстановлении числа лейкоцитов (лимфоцитов) или тромбоцитов у перенесших лучевое поражение или отсутствия устойчивой нормализации в их уровне. Другими словами, без истинной нормализации эритропоэза не следует ожидать стабильного восстановления других ростков кроветворения и выполняемых ими функций, в первую очередь иммунитета. Отмечаемые в течение длительных периодов пострадиационные лейкопенические состояния с этих позиций следует, по-видимому, рассматривать как обусловленные преимущественной пролиферацией эритробластических элементов, что вызывалось необходимостью первоочередного восстановления и поддержания функций красной крови в пределах физиологической нормы. Выявленные изменения в интенсивности гемопоэза нельзя однозначно связывать с глубиной радиационного поражения системы крови. Повышение темпа эритропоэза можно рассматривать как компенсаторно-приспособительную реакцию, направленную на поддержание функций красной крови в пределах физиологической нормы. Степень выраженности этой реакции определяется двумя главными факторами: необходимостью и возможностью ее развития. Необходимость повышения темпа эритропоэза возникает при ускоренной убыли эритроцитов из сосудистого русла. Выраженность этой реакции зависит от пределов физиологических возможностей регенеративного аппарата эритропоэза и глубины его радиационного поражения. Продолжительность жизни эритроцитов в сосудистом русле у изучавшихся собак, определявшаяся с помощью метки собственных эритроцитов 51Cr, оказалась сокращенной. У одной опытной и у одной контрольной здоровой собаки одновременно определяли и интенсивность эритропоэза, и эритрофагоцитарную функцию селезенки через 190 сут после окончания многократного лучевого воздействия, используя 51Cr в качестве метки. С указанной целью через 2 нед после введения в организм меченных изотопом собственных эритроцитов извлекали селезенку и готовили срезы толщиной около 0,5 см. Срезы промывали физиологическим раствором для удаления крови, после чего определяли их радиоактивность, средняя величина которой выражалась в процентах от радиоактивности селезенки контрольной собаки. У облученной собаки радиоактивность была в 1,5 раза больше, что свидетельствовало об усилении фагоцитоза эритроцитов клетками селезенки. Подробное описание состояния указанных групп собак дано в наших монографиях [Акоев, 1970; Акоев и др., 1981]. Средние данные по 12 собакам через 190 сут после окончания многократного облучения: внешнее клиническое благополучие, морфологический состав крови у облученных собак не выходил за пределы нормы. Однако отмечалось относительное снижение числа лейкоцитов (на 2,9 тыс. клеток в 1 мм3 крови) и повышение содержания ретикулоцитов (3,1 % против 1,2 % в контрольной группе). Это давало повод предполагать скрытое напряжение эритропоэза. И действительно, оно подтвердилось исследованиями с меченым железом. Суммарное поступление эритроцитов в русло крови оказалось в 1,75 раза выше нормального, что, в свою очередь, могло служить косвенным доказательством ускоренного разрушения эритроцитов. Возможность очень длительного сохранения сокращенной продолжительности жизни эритроцитов у человека после радиационных воздействий была показана Хубером и Ширакурой на примере исследований, проводившихся не менее чем через 6 лет после успешной лучевой терапии 25 женщин по поводу рака матки и обнаруживших у них уменьшенную продолжительность пребывания эритроцитов в сосудистом русле. Изложенные исследования, выполненные нами на собаках, демонстрируют определенную последовательность включения и выключения разных механизмов усиления продуктивной способности гемопоэза, необходимых для поддержания в первую очередь функции красной крови. Из анализа представленных данных можно заключить, что примат производства клеток красного ростка существует тогда, когда имеется значимая для организма недостаточность количества эритроцитов и гемоглобина. Первым мобилизуемым резервом является расширение плацдарма кроветворения, т. е. увеличение массы гемопоэтической ткани. Этот резерв — расширенный плацдарм кроветворения — используется организмом наиболее длительно. У собак в этот период «костный мозг» грудины и эпифизов на секции становится красным, т. е. происходит замещение его эритропоэтической тканью. Вторым возможным мобилизуемым резервом является ускорение прохождения клетками делящегося и делящегося-созревающего пулов. Наконец,позже всех и на самое короткое время (на месяц-полтора) может увеличиваться митотический индекс эритробластов, непосредственно отражающий в этих условиях сокращение их генерационного цикла. Основные наблюдения за состоянием кроветворения у собак в отдаленный пострадиационный период (изменения периферической крови и интенсивность эритропоэза) были подтверждены нами и в экспериментах на крысах. Возможность сокращения продолжительности генеративного цикла бластных клеток белого и красного ряда была доказана А. В. Илюхиным с соавторами при длительном хроническом воздействии радиации на собак. После введения фенилгидразина или массивного кровопускания также обнаружено сокращение генерационного времени делящихся эритробластов, которое в основном происходило за счет стадии G1. Была подробно изучена цитокинетика кроветворения у собак на протяжении трехлетнего слабого хронического воздействия радиации. Собак в течение трех лет непрерывно (кроме времени на кормление и обследование) облучали: суммарные дозы от 0,63 до 5,70 Гр. В периферической крови на 4—12-м месяце облучения отмечали ретикулоцитоз, сокращение времени пребывания эритроцитов в крови (на 12—24-м месяце) при сохранении числа эритроцитов и содержании гемоглобина в пределах контрольных значений (табл. 2). Это могло указывать па скрытое напряжение эритропоэза для компенсации сокращения продолжительности жизни эритроцитов и поддержания увеличенной скорости обновления эритроцитов. Оценка костно-мозговой продукции эритроцитов показала статистически значимое ее увеличение, сохранявшееся в течение всего второго года облучения. Тем не менее качество эритроцитов (насыщенность гемоглобином, кривые Прайс—Джонса) не ухудшалось. К концу третьего года все показатели были в пределах нормы. Со стороны белой крови отмечали стойкое снижение лейкоцитов и нейтрофилов на протяжении всего третьего года облучения. Продолжительность пребывания гранулоцитов была снижена. К концу третьего года облучения костно-мозговая продукция гранулоцитов в группе, имевшей суммарную дозу воздействия 5,7 Гр, снизилась примерно наполовину. Осмотическая стойкость лейкоцитов была сниженной на протяжении последних 2,5 лет наблюдений. В отличие от показателей красной крови, которые нормализовались на протяжении третьего года облучения, в белой крови продолжала к этому сроку нарастать недостаточность миелопоэза. Изменения показателей красной крови в условиях слабого хронического воздействия были менее выраженными и восстанавливались быстрее, явно отдавая приоритет восстановлению эритропоэза. Авторы пишут о возможном дефиците стволовых клеток и их направленной дифференцировке в эритроидном направлении.
Таблица 2. Сроки (в месяцах) выявления статистически достоверных отклонений в цитокинетических показателях эритропоэза и лейкогранулопоэза у собак на протяжении трехлетнего хронического радиационного воздействия в малых дозах (наша обработка данных А. В. Илюхина и др.)
| Показатель | Эритропоэз | Лейкогранулопоэз | ||
|---|---|---|---|---|
| Увеличение | Снижение | Увеличение | Снижение | |
| Число клеток в периферической крови | 4-12 (ретикулоциты) | — | 24-36 | |
| Период циркуляции клеток в крови | 12-24 | 32 | ||
| Скорость обновления клеток в крови | 24 | 32 | ||
| Костно-мозговая продукция зрелых клеток | 12-24 | 36 | ||
| Качество зрелых клеток | 12-36 | |||
| Число клеток костного мозга общее | 28-36 | 24-36 | ||
| Скорость обновления клеток костного мозга | 28 | 28-26 (до 2 раз) | ||
| То же для отдельных властных форм | 24-32 | |||
| Длительность генерационного цикла | 28 (G1) | 28-36(G1, S, G2) | ||
| Костно-мозговой резерв | 5-36 | |||
Регуляция размножения и дифференцировки кроветворных клеток
В связи с успехами исследования процессов кроветворения и функциональных возможностей стволовых клеток при использовании метода селезеночных колоний и культур на полутвердых средах и других методов представления о регуляции путей их пролиферации и дифференцировки существенно изменились. Экспериментально подтвердилось существование единой полипотентной стволовой клетки, способной к дифференцировкам по всем направлениям гемопоэза, включая лимфопоэз (рис. 3). Казалось бы, это непосредственно доказывает правильность исходного положения о конкуренции различных ростков кроветворения за направление дифференцировки стволовой клетки в соответствии с потребностями организма. Однако вопрос оказался значительно сложнее. Наряду с исходной полипотентной стволовой кроветворной клеткой могут существовать и обладающие стволовыми свойствами предшественники, коммитированные (т. е. ограниченные к выбору направления дифференцировки) предшественники двух типов — миелопоэза и лимфопоэза. Это первое и главное разделение направлений возможной дифференцировки единой полипотентной стволовой клетки. Как осуществляется регуляция выбора дифференцировки — в сторону миелопоэза или лимфопоэза, не очень ясно. Стволовые клетки, коммитированные в сторону лимфопоэза, дифференцируются сначала в общих, а затем в раздельных предшественников Т- и В-лимфоцитов, из которых через ряд промежуточных стадий образуются Т-лимфоциты (приобретая определенные клеточные рецепторы) и В-лимфоциты (приобретая рецепторы к комплементу, к фрагментам и комплексам иммуноглобулинов и комплексам антиген—антитело). Кроме того, из них образуются плазматические клетки. Лимфопоэз наименее изучен. До сих пор неясно, имеются ли в лимфоцитарном ряду клетки, не способные к пролиферации. Среди лимфоидных клеток трудно определить принадлежность их к пролиферирующему, созревающему или функциональному пулам, так как возможны взаимные перекрытия. Стволовые клетки, коммитированные в сторону миелопоэза, в дальнейшем через поэтинчувствительные предшественники разделяются на три ростка: эритроцитарный, гранулоцито-макрофагальный и тромбоцитарный. В эритроцитарном направлении выявлено несколько промежуточных клеток (в том числе и бурстообразующая клетка) между общим предшественником миелопоэза и эритропоэтинчувствительными клетками. Последующие морфологически различаемые клетки относятся к проэритробласту, трем видам эритробластов, ретикулоциту и зрелому эритроциту. Обычно происходит 5—6 митозов в морфологически различимых клетках, и каждый эритробласт дает 30—60 эритроцитов. Эритробласты теряют способность к делению, как только в клетке накопится 27 нг гемоглобина. В случае замедления синтеза гемоглобина увеличивается число делений эритробластов. Из одной полипотентной стволовой клетки, начавшей дифференцировку в сторону миелопоэза, может образоваться около 1 млн эритропоэтинчувствительных клеток и около 0,1 млн клеток — предшественников гранулоцитов и макрофагов. Рис. 3. Основные пути дифференцировки полипотентной стволовой клетки, клеточные формы и их главные функции
Рис. 3. Основные пути дифференцировки полипотентной стволовой клетки, клеточные формы и их главные функции
 Рис. 4. Возможные пути регуляции пролиферации и дифференцировки полипотентных стволовых клеток в соответствии с запросами организма
Рис. 4. Возможные пути регуляции пролиферации и дифференцировки полипотентных стволовых клеток в соответствии с запросами организма
Развитие гранулоцито-макрофагального направления миелопоэза проходит на стадии образования клеток, дающих колонии на полутвердых средах (KOEK), кластерообразующих клеток и предшественников гранулоцитов и моноцитов-макрофагов. Последующие клетки морфологически различимы. Миелобласт нейтрофильный далее дает клетки гранулоцитарного ряда (эезинофил), нейтрофильного ряда (нейтрофил) и базофильного ряда (базофил и тучная клетка). Монобласт дает моноциты, макрофаги, гистиоциты, купферовы клетки печени и остеокласты. Клетки моноцитопоэтического ряда объединены в систему фагоцитирующих мононуклеаров как способные к фагоцитозу и осуществляющие в норме функции клеточного фагоцитоза. Ранее входившие вместе с указанными клетками в ретикуло-эндотелиальную систему ретикулярные клетки и эндотелиальные клетки исключены из системы фагоцитирующих мононуклеаров как являющиеся лишь факультативными фагоцитами, не имеющими рецепторов для иммуноглобулинов и не способными к иммунному фагоцитозу. Клетки же системы фагоцитирующих мононуклеаров в процессе дифференцировки приобретают рецепторы для иммуноглобулинов и комплемента и становятся способными к активному иммунному фагоцитозу. Тромбопоэтинчувствительные клетки формируют морфологически различимые клетки: мегакариобласт, промегакариоцит и тромбоциты. Для предшественников характерны 4—5 эндомитозов. Таким образом, по современным данным, клетки костного мозга, способные к делению, можно условно разделить на морфологически различимые клетки, как правило способные к очень ограниченному числу делений (4—8 делений), и на морфологически неразличимые клетки, выявляемые лишь по способности их давать колонии в селезенке облученных мышей или пролиферировать на полутвердых средах (и другими методами), способные к чрезвычайно активной пролиферации. Число делений, которые может совершить полипотентная стволовая клетка, на порядок больше, чем для морфологически различимых клеток. В норме их в костном мозге лишь около 0,5% от числа всех клеток. Однако достаточно сохраниться 0,1% стволовых клеток после сильного радиационного воздействия, чтобы обеспечить восстановление нормального пула стволовых клеток и резкое повышение дифференцировки клеток в последующих отделах. При нормальном кроветворении абсолютное большинство стволовых клеток находится вне клеточного цикла (период G0 или длительный период G1). Стабильное поддержание величины отдела стволовых клеток обеспечивается небольшой долей клеток, вступивших в митотический цикл. В случае уменьшения отдела стволовых клеток в пролиферацию может вовлекаться вся популяция сохранившихся стволовых клеток. При этом время удвоения популяции стволовых клеток оказывается очень коротким, порядка 15—20 ч. По данным ряда авторов, укорочение или удлинение стадии G1 стволовых клеток соответственно сопровождается пролиферацией (т. е. оставлением в пуле стволовых клеток) или дифференцировкой (т. е. уходом в последующие отделы). Следовательно, основной вклад в восполнение дефицита клеточности костного мозга вносит пролиферация морфологически неразличимых клеток и потому исследование регуляции их численности и регуляции выбора направления дифференцировки является чрезвычайно важным. К сожалению, в этих вопросах еще очень много неясного. Как решается для стволовых клеток первый выбор направления дифференцировки (в сторону лимфопоэза или миелопоэза), тоже неясно. По многочисленным данным, в опытах на собаках и обезьянах аллогенная кроветворная ткань вызывала появление огромного количества лимфоцитов, специфически иммунных против реципиента, при полной или частичной блокаде остального кроветворения. Следовательно, возможно переключение части стволовых клеток с эритроидного на лимфоидный путь дифференцировки. Характер дифференцировки кроветворной ткани в условиях трансплантации указывает на конкуренцию миелоидных и лимфоидных клеток за общий предшественник. У полицитемических радиохимер, у которых снижен эритропоэз, ускоряется восстановление лимфопоэза. Приведенные нами данные о существенной задержке пострадиационного восстановления числа лимфоцитов в крови человека в период, когда эритропоэз напряжен, также свидетельствуют о конкурентных отношениях ростков кроветворения. Об этом непосредственно говорит и проведенное нами изучение соотношений в морфологически различаемых клетках разных ростков костного мозга у облучавшихся собак. Тем не менее некоторыми учеными предполагается, что направление дифференцировки стволовых клеток или закреплено генетически постоянно, или осуществляется стохастически, в обоих случаях — независимо от запросов организма. Возможным доказательством этого может служить одно и то же распределение маркерных клеток по разным росткам гемопоэза при изменении общих запросов организма. В связи с изложенным, вероятно, более правильно считать существование конкурентных отношений между ростками кроветворения за преимущественную продукцию конечных зрелых форм клеток, а не примат дифференцировки полипотентной стволовой клетки в тот или иной росток гемопоэза. При этом влияние дальнодействующей для гемопоэза регуляции с целью реакций на запросы организма в экстремальных ситуациях может осуществляться не только через поэтинподобные регуляторы, но и через микроокружение в локальных участках гемопоэтических тканей, которое может зависеть также от запросов организма. Эритропоэтин для красного ростка и колониестимулирующий фактор для гранулоцитарного ростка стимулируют пролиферацию клеток и укорачивают средний период генерации клеток. При этом удлинение или укорочение стадии G1 клеточного цикла может определять уход клеток в пролиферацию или дифференцировку (рис. 4). В случае вступления клетки в деление сокращается генерационный цикл и клетка не успевает осуществить дифференцировку. Пролиферация прекращается, как только весь объем микроокружения окажется занятым стволовыми клетками. Таким образом, исследования реакции отдела стволовых клеток на экстремальные воздействия и пострадиационные изменения гемопоэза позволили показать ряд общих закономерностей в реакции этой системы на экстремальные воздействия и в процессах последующего восстановления. Влияние запросов организма на гемопоэз осуществляется дальнодействующей регуляцией посредством гуморальных факторов. Поэтому следует кратко рассмотреть отдельные радиационные изменения со стороны эндокринной и других систем организма.
Другие проявления отдаленных радиационных последствий в предпатологический период
Обобщение и анализ остаточных и отдаленных последствий воздействия радиации проводились неоднократно. Наряду с изменениями в системе крови, изложенными выше, отмечались последствия и в других системах организма. К ним в первую очередь необходимо отнести нарушения иммунологической реактивности, проявляемые наиболее четко в снижении устойчивости к инфекционным агентам. Так, устойчивость к живому возбудителю газовой гангрены и его токсину в экспериментах была снижена у мышей через 190 сут, а у крыс — через 150 сут после воздействия радиации. Имеются также данные, что для полного восстановления иммунобиологической реактивности у облученных грызунов необходимы чрезвычайно длительные сроки, составляющие значительную часть всей нормальной продолжительности жизни животных. Это хорошо соответствует изложенным выше данным о длительной задержке восстановления лимфоидной и гранулоцитарной части кроветворения после облучения. Цикл исследований был выполнен нами на собаках через 190 сут после окончания многократного воздействия (осуществлявшегося в 2—4 приема на протяжении 2,5—14 мес) при полном клиническом благополучии животных и гематологических показателях в пределах видовой нормы к моменту исследования. О скрытой неполноценности восстановления крови можно было только предполагать: число лейкоцитов несколько ниже, чем в контроле, фагоцитарная активность лейкоцитов (число фагоцитирующих клеток) и фагоцитарный индекс (количество микробов на один фагоцит) недостоверно выше, а абсолютный фагоцитарный показатель (число введенных в кровь клеток золотистого стафилококка, фагоцитированных лейкоцитами 1 мм3 крови) был на 25% ниже контрольного уровня. Кроме того, у одной из собак при некотором снижении числа эритроцитов обнаружена в 1,75 раза более высокая суммарная продуктивность эритропоэза. Следовательно, отмечалась типичная картина скрытого напряжения гемопоэза, прикрытого стандартными показателями периферической крови в пределах видовой нормы или несущественно отличаясь от нее. Эффективной проверкой состояния систем организма собак спустя 190 сут после окончания многократного воздействия радиации являлась реакция на травматическое повреждение и скорость посттравматической регенерации. С этой целью на спину собак в области между лопатками наносили кожную рану путем отсечения кожного лоскута. Рану антисептической обработке не подвергали. Оценивали период времени, необходимый для уменьшения площади раневой поверхности наполовину. Период полурепарации раны у ранее облучавшихся собак был больше, чем у контрольных животных, имея тенденцию к возрастанию с увеличением суммарной дозы облучения. Количество лейкоцитов, участвовавших в фагоцитозе введенного в кровь золотистого стафилококка, у некоторых животных резко увеличилось, в 1,5—2,5 раза превысив исходный уровень. У ранее облучавшихся собак, наоборот, отмечено уменьшение почти в 2 раза исходного и контрольного уровней фагоцитарной активности лейкоцитов. Еще большие различия выявлены в абсолютном фагоцитарном показателе: например, на 21-е сутки после ранения он был равен у контрольных животных 29,8 тыс., а у облученных собак — 5,5 тыс. микробных тел, фагоцитированных лейкоцитами 1 мм3 крови. В экспериментах на облученных мышах с закрытым переломом костей голени или стандартной кожной раной также обнаружено замедление посттравматической регенерации, в том числе и в случае, когда животным наносились травмы через месяц после четырехкратного с месячным интервалом облучения в несмертельных дозах. Относительная величина удельного замедления репарации при заживлении костной ткани была больше, чем кожной. Сниженную устойчивость собак к повторному воздействию радиации наблюдали через 10 мес после окончания многократного воздействия радиации. Известно, что изменения со стороны центральной нервной системы могут отмечаться у перенесших лучевое воздействие в течение длительных периодов времени. Более того, со временем они у ряда лиц могут быть более выраженными. У некоторых лиц, подвергавшихся субтотальной рентгенотерапии, явления астенизации прогрессивно нарастали в течение 3—4 лет и в дальнейшем на протяжении 5—7 лет и более перерастали в симптомы органического поражения нервной системы. К последствиям, для которых характерен длительный латентный период после воздействия радиации, относят дегенеративно дистрофические и склеротические изменения в различных органах и системах. Атрофические, склеротические и язвенные изменения слизистой желудочно-кишечного тракта, циррозы печени, нейросклерозы и др. В. Н. Стрельцова и Ю. И. Москалев относят к неопухолевым формам поздних эффектов действия радиации. О развитии в поздние сроки радиационных функциональных и морфологических изменений эндокринных желез сообщалось в ряде работ. В. В. Шиходыров и соавторы отмечают некоторые различия в реакции ткани на грубое структурное поражение (обычно в критических органах) и на менее выраженное поражение (обычно в некритических органах). В обоих случаях изменения не зависят от характера поврежденного агента. В первом случае начальным звеном процесса служат некробиотические изменения ткани. Наряду с процессом дистрофии развиваются явления склероза. Разрастание соединительной ткани оценивается вторичным, заместительным процессом в ответ на уменьшение числа клеток паренхимы органа. Определенное значение в развитии упомянутых нарушений имеют изменения кровеносных сосудов. Во втором случае не возникают первоначальные грубые структурные изменения паренхимы органа. В течение длительного времени отсутствуют какие-либо морфологические повреждения. Происходят постепенное развитие патологического процесса, уменьшение клеточности паренхимы органа и нарастание атрофии и склероза. Изложенные процессы в целом имеют ту же направленность, что и при старении. У облученных животных эти процессы происходят более ускоренно. Так, однотипные изменения в центральной нервной системе наблюдаются у интактных собак в 7—12 лет и у облучавшихся — в 5—7 лет. Важно отметить, что дистрофические изменения и нарушение процессов физиологической регенерации часто могут сопровождаться развитием очагов узловатой гиперплазии. Отмечается, что в условиях длительно текущих и выраженных дистрофических нарушений во внутренних органах возникает повышенная нагрузка на органы эндокринной системы, связанная с возможным перенапряжением и истощением их. В менее демонстративной форме такое влияние на старение оказывают и другие факторы (инфекционные заболевания, интоксикация, нарушения питания и др.). Для пострадиационных изменений иммунитета наиболее характерны сочетания иммунодефицитного и аутоагрессивного состояний, недостаточность механизмов неспецифической и специфической иммунной защиты организма. Общая иммунологическая реактивность после введения в организм ряда радионуклидов изменялась при таких низких суммарных дозах, как 0,3 Гр за 30 сут для стронция и 0,6 Гр за 30 сут для цезия. Фагоцитарные реакции изменялись примерно при таких же низких дозах воздействия. Еще более низкие дозы необходимы, чтобы началась атрофия лимфоидной ткани и плазматизация селезенки (0,10—0,35 Гр за год воздействия). Примерно при таких же суммарных поглощенных дозах происходили изменения в образовании аутоантител (увеличение их). Изложенное свидетельствует о чрезвычайной чувствительности отдельных реакций иммунитета к внешним воздействиям. Изменение соотношения числа Т- и В-лимфоцитов также отмечали при суммарных поглощенных дозах порядка 0,1 Гр. Нелетальное воздействие радиации может вызывать ряд аутоиммунных и аллергических расстройств (данные В. М. Шубина). Различают первичную (генетически обусловленную) и вторичную (индуцированную) иммунологическую недостаточность. Последняя может развиваться как после воздействия ионизирующих излучений, так и после приема иммунодепрессантов, а также при старении и некоторых заболеваниях. Все эти средства и состояния неспецифически угнетают и клеточный и гуморальный иммунитет. Особенностью развития аутоиммунной реакции при действии ионизирующей радиации и других воздействий в отличие от иммунного ответа на чужеродные антигены является ее формирование не в сторону развития невосприимчивости, а в сторону сенсибилизации, т. е. повышения чувствительности к тканевым продуктам распада. Образование аутоантител происходит не только при действии излучения, но и при иммунизации микробными антигенами. Иммуногенез после облучения угнетен по отношению к чужеродным агентам, а по отношению к собственным тканям, наоборот, активно развивается. Известно, что в тканях (особенно в селезенке) здорового организма, не имеющего контакта с инфекцией или предупреждающими воздействиями, всегда можно обнаружить нормальные клетки, образующие аутогемолизины. В крови их число доходит до 3% ядросодержащих клеток, а в паренхиматозных органах — в пределах 5—10%. Они выполняют важную физиологическую роль по удалению старых клеток и обезвреживанию продуктов их распада. Однако под влиянием радиации, других повреждающих факторов и старения эта функция перерастает в аутоагрессивную и становится сама повреждающим фактором. Многие из изложенных в этом и предыдущих разделах клинических форм и патологических состояний не являются специфическими для радиационных последствий. Они известны и для возрастной патологии. В связи с этим широко обсуждаются и вопросы ускорения и увеличения частоты возрастных изменений в результате действия радиации. Особый интерес вызывает определенная последовательность изменений в эндокринной системе, играющей важную роль в регуляторных механизмах взаимодействия систем организма. Анализ временной последовательности изменений в эндокринной системе и становления клинических форм отдаленных радиационных последствий позволил нам прийти к выводу о зависимости многих проявлений радиационных последствий от недостаточности эндокринных органов. По данным Е. Н. Антипенко, изучавшего функциональное состояние и структуру щитовидной железы у собак, через 3,5 мес после лучевого воздействия в дозе 4 Гр йодфиксирующая функция еще не отличалась от нормы. Однако при гистологическом исследовании были обнаружены участки с пониженной и повышенной тиреоидной активностью. Через 6—13 мес после облучения деятельность железы усиливалась, о чем свидетельствовали кривые накопления радиойода в железе, содержание органически связанного радиойода в плазме крови и строение железы. Характерно, что явлений тиреотоксикоза не было. Однако в этот период продолжали нарастать со временем частота, а также выраженность дистрофических и некробиологических изменений. Масса железы снижалась. Выше приводились данные о состоянии кроветворения у собак в эти же временные отрезки: скрытое напряжение эритропоэза при нормальной, но нестабильной картине крови или при незначительном снижении числа лейкоцитов и лимфоцитов или эритроцитов при увеличении числа ретикулоцитов, костный мозг гиперпластичен по красному ростку, продолжительность жизни эритроцитов сокращена. Через 5 мес после облучения крыс в дозе 6 Гр нами проводился комбинированный опыт. Величина максимального накопления радиойода в железе крыс была в 1,5 раза выше, чем в контроле. Скорость накопления и выведения его также оказалась выше. Относительная масса железа на 100 г массы тела была ниже, чем у контрольных крыс. Усиленная йодфиксирующая способность железы, относительно невысокое содержание в ней коллоида и его повышенная резорбция, а также полнокровие железы свидетельствовали о значительном усиление гормонообразования и особенно гормоновыделения без явлений тиреотоксикоза. Одновременно у части крыс анализировали состояние кроветворения. У облученных животных отмечали тенденцию к снижению числа эритроцитов и гемоглобина, повышение ретикулоцитов в крови и увеличение суммарной продуктивности эритроцитов по данным включения радиоактивного железа. Эта тенденция к напряжению эритропоэза оказалась менее выраженной, чем у более молодых крыс (опыт ставили на крысах в возрасте 1,5 лет, у которых можно было ожидать более яркого проявления гормональной терапии в связи с возможным начинающимся возрастным угасанием эндокринных функций). У отдельных животных отмечалось, наоборот, значительное снижение скорости включения меченого железа и уменьшение количества эритроцитов. У другой группы крыс определяли устойчивость к повторному облучению. Полулетальная доза повторного облучения, которое проводили через 5 мес после первого воздействия радиации, оказалась на 1,47 Гр меньше, чем в контроле. Устойчивость ранее облучавшихся крыс к токсину возбудителя газовой гангрены в дозе, близкой к минимальной абсолютно смертельной, оказалась также сниженной. Среднее время выживаемости сократилось по сравнению с контролем. Учитывая, что усиленная функция щитовидной железы у облучавшихся животных в указанных опытах не сопровождалась явлениями токсикоза и могла быть расценена как компенсаторная в связи с возросшими потребностями организма, в последнем опыте на крысах нами проверялась эффективность введения в организм гормонов щитовидной железы (тиреоидин, который давали с пищей ежедневно на протяжении четвертого и пятого месяцев после облучения, или диэтилстильбестрол, который добавляли в пищу в те же сроки). У ранее облучавшихся крыс, получавших гормоны щитовидной железы, отмечали значительное снижение функций щитовидной железы (йодфиксирующая функция резко снижена, относительная масса железы ниже контрольной, железы образованы крупными фолликулами с большим количеством гомогенного коллоида); повышение устойчивости к повторному воздействию радиации, которое, однако, не достигло контроля; повышение устойчивости крыс к токсину возбудителя газовой гангрены (но не достигло контрольного уровня); снятие напряжения эритропоэза (число эритроцитов и содержание гемоглобина повысились, число ретикулоцитов снизилось, продукция эритроцитов уменьшилась до контрольного уровня). В этом опыте введение тиреоидина ранее облучавшимся крысам оказало благотворное действие на устойчивость организма к внешним воздействия, на щитовидную железу, на гемопоэз, несмотря на усиление гормонообразовательной и инкреторной функций железы у не получавших тиреоидина крыс. Это доказало, что в организме в этот период не хватало гормонов щитовидной железы, несмотря на усиленное поступление их из железы, а усиление функций железы носило компенсаторно-приспособительный характер в ответ на повышенное расходование их в облученном организме. Дополнительное введение гормонов в организм по этой причине не вызывало токсикоза, а оказывало благоприятное действие. Гормональный дефицит и указанные отдаленные последствия оказались в определенной связи. Другая группа крыс получала половой гормон. У крыс этой группы йодфиксирующая функция щитовидной железы была чрезмерно увеличена. Относительная масса железы увеличилась, фоликулы были образованы гипертрофированными клетками, запас коллоида практически отсутствовал. Это чрезмерное напряжение гормонообразовательной и инкреторной функций щитовидной железы могло быть обусловлено значительным усилением тиреотропной функции гипофиза за счет снижения гонадотропной в связи с насыщением организма половым гормоном. Устойчивость к повторному воздействию радиации повысилась. Введение в организм полового гормона привело не только к уменьшению недостатка в нем (крысы были к этому времени в возрасте двух лет, т. е. нерепродуктивного возраста), но и через дальнейшее усиление функций щитовидной железы к снижению дефицита в гормоне последней. У животных под влиянием введения полового гормона наблюдали увеличение содержания гемоглобина при сохранении числа эритроцитов на прежнем уровне, увеличение числа ретикулоцитов и тенденцию к дальнейшему увеличению суммарной продукции эритроцитов костным мозгом. Таким образом, если введение тиреоидина в организм крыс через 5 мес после облучения способствовало ликвидации напряжения эритропоэза и гиперфункционального состояния щитовидной железы, предотвращая возможность их функционального истощения, то введение полового гормона, наоборот, вызывало дальнейшее функциональное напряжение гемопоэза и щитовидной железы, усугубляло возможность истощения функций или перехода их в патологическую сверхкомпенсацию. В наших исследованиях на собаках, которым наносили через 190 сут после окончания многократного облучения хирургическую травму, также испытывали действие предварительного 15-суточного введения тиреоидина с пищей или преднизолона (препарат коры надпочечников). В группе собак, получавших тиреоидин перед нанесением травмы, количество эритроцитов и гемоглобина осталось таким же, как и в группе облученных, но не получавших гормонального препарата. Число ретикулоцитов снизилось до нормального уровня. Продукция эритроцитов, определявшаяся по включению меченого железа, снизилась. Число лейкоцитов в 1 мм3 крови поднялось. Резко возросли показатели фагоцитарной активности лейкоцитов: число фагоцитирующих золотистый стафилококк клеток, количество поглощенных микробов на один фагоцит и число микробов, фагоцитированных лейкоцитами 1 мм3 крови, увеличились в 2—2,5 раза и более по сравнению с облученным контролем и здоровыми животными. В группе собак, получавших преднизолон, несколько увеличилось число эритроцитов и лейкоцитов и уменьшилась продукция костным мозгом эритроцитов. Содержание гемоглобина, число ретикулоцитов не изменились. Показатели фагоцитарной активности лейкоцитов крови также существенно были более высокими. Однако после ранения собак выявилась неполноценность функции фагоцитоза. Высокая фагоцитарная активность лейкоцитов до нанесения раны, вызванная введением в облученный организм преднизолона, оказалась менее полноценной, чем более умеренное участие лейкоцитов в фагоцитозе микробных тел у здорового организма. Это обусловлено тем, что положительное влияние эндокринных препаратов на фагоцитарный индекс, т. е. на количество микробов, поглощаемых каждым лейкоцитом, на фоне сниженного числа лейкоцитов в крови оказалось недостаточным, чтобы абсолютный фагоцитарный показатель после ранения был таким же, как у здоровых животных. Тем не менее гормонотерапия все же позволила весьма существенно поднять этот показатель (например, на 21-й день после ранения с 5,5 до 12,2 тыс. и до 20,2 тыс. микробов, фагоцитированных лейкоцитами 1 мм3 крови соответственно в группах, получавших тиреоидин и преднизолон). В другом опыте, проводившемся на живых мышах через 5 мес после воздействия радиации в дозе 5,75 Гр, вызвавшем гибель 2/3 животных в течение 4 мес, также определяли функциональное состояние щитовидной железы. У этих животных поглощение радиойода щитовидной железой составляло 60% от введенного количества изотопа, что существенно ниже по сравнению с контролем. Активность связанного с белком йода также была ниже контроля. Все это позволило предполагать наступающее истощение функций щитовидной железы. Морфологические исследования подтвердили это заключение. Следует сказать, что смену повышенной деятельности щитовидной железы гипофункциональным состоянием после радиационного воздействия отмечали многие исследователи. Так, у крыс через 40 сут после однократного введения 210Ро наблюдали морфологические признаки гиперфункционального состояния щитовидной железы, а через 70—100 сут — явления истощения ее. Смену признаков гипертиреоидного состояния на признаки гипофункционального выявляли у собак через 2—3 года после общего облучения или облучения верхней части туловища (накопление железой радиойода, содержание связанного с йодом белка в крови, реакция на тиреотропную стимуляцию). У части животных развилась микседема. Аналогичная последовательность изменений функции коры надпочечников может быть прослежена после воздействия радиации. При лучевой болезни у собак вслед за усилением их функции первые признаки недостаточности коры надпочечников стали появляться спустя 3 мес. Наблюдались все возрастающие патологические изменения в надпочечниках по мере отдаления срока после лучевого воздействия. Э. Бетц первый обратил внимание на то, что состояние некоторых инкреторных функций после воздействия радиации такое, какое может наблюдаться при постоянном поддержании реакции напряжения, и при этом особое внимание было обращено на длительно сохраняющееся состояние гиперкортицизма. Об отдаленных изменениях в эндокринных функциях у людей, переживших ядерный взрыв в Хиросиме и Нагасаки, много сообщалось. При этом подчеркивалась определенная однотипность этих изменений в различных эндокринных органах, нередко доходивших до стадии истощения. Известно, что значение дисгормональных расстройств особенно возрастает в период отдаленных лучевых последствий. Возникновение ряда опухолей эндокринных органов (в первую очередь щитовидной железы) в отдаленный период отнесено к наиболее радиочувствительному виду радиационных неоплазм. Характерно, что для развития этих опухолей необходим определенный оптимум дозы, который бы не приводил к гибели и позволил в течение длительного времени сохранять напряжение функций эндокринных желез. Общую последовательность развития отдаленных изменений эндокринного статуса облученного организма можно представить так. У благополучно перенесших лучевое воздействие наступает полное клиническое выздоровление, несмотря на то что в отдельных системах имеются остаточные повреждения и нарушения. Они не проявляются и могут никогда не проявиться, пока компенсаторно-приспособительные реакции организма находятся на адекватном уровне для поддержания состояния стабилизация здоровья. Однако в ряде случаев через определенный латентный период повышается функциональное состояние периферических эндокринных желез (щитовидная железа и, возможно, другие), в организм поступает все большее количество гормонов этих желез без явлений гормональных токсикозов. Организм не испытывает их избытка. Наоборот, имеются основания предполагать, что в этих условиях ткани-потребители гормонов испытывают их недостаток, так как дополнительное введение их в организм тоже не вызывало токсикоза и оказывалось во многих отношениях благоприятным. Длительное состояние гиперфункции желез компенсаторного плана в связи с дефицитом гормонов в конце концов может переходить в гипофункциональное состояние и истощение. Возрастающий со временем недостаток гормонов, т. е. срыв компенсации, создает благоприятные условия для клинического проявления остаточных и возникновения отдаленных радиационных последствий. Как показали наши исследования, такие последствия, как напряжение эритропоэза и сниженная иммунобиологическая реактивность, проявлялись в большей степени в те периоды, когда дефицит в гормоне щитовидной железы предполагался более выраженным. В качестве примера возможного экспериментального подтверждения указанных положений сошлемся на опыт по изучению эритропоэза. Были получены следующие результаты: у крыс напряженность гемопоэза была более выражена через 4 мес, а не через 2 мес после облучения. Свидетельством того, что через 4 мес организм испытывал более выраженный недостаток в гормонах щитовидкой железы, может, по-видимому, служить более высокая эффективность 30-суточного введения тиреодина в малых физиологических дозах через 4 мес после облучения, чем через 2 мес. Наблюдения над людьми, перенесшими радиационное воздействие, показывают, что нередко следует говорить лишь о практическом выздоровлении. Различные факторы нерадиационной природы часто служат внешним поводом для проявления отдаленных радиационных последствий. Так, у лиц, подвергавшихся радиотерапии по поводу злокачественных заболеваний, отмечали, что толчком к появлению астенизации иногда служили внешние факторы нерадиационной природы. Они выводили больных из состояния компенсации. Есть данные о том, что через 9 лет у большинства лиц, перенесших воздействие радиации ядерного взрыва, несмотря на восстановление здоровья, реакция их на всякое напряжение изменяется в сторону возможности появления хронических расстройств. Это связано с тем, что патогенетически более поздние проявления радиационного поражения являются в определенной мере универсальными и их выраженность зависит во многих отношениях от одних и тех же компенсаторно-приспособительных реакций, а срыву компенсации могут способствовать разные факторы. Взаимно отягощающее влияние разных факторов на состояние щитовидной железы показано и на примере воздействия радиации и инфекции. Значительное повышение функции железы у крыс вызывали раздельно радиацией (через 4 мес после облучения в дозах 7,5 и 8,5 Гр) и заражением газовой гангреной (исследовали морфологию железы, массу секретирующей тиреоидной ткани, содержание меченого тиреоглобулина в железе, массу железы). У крыс, которых заражали газовой гангреной через 4 мес после воздействия радиации, гистологическое состояние щитовидной железы указывало на значительное усиление ее функции. Однако оно не сопровождалось увеличением массы железы, а содержание тиреоглобулина в железе было существенно ниже, чем у здоровых животных. Такимобразом, инфекция, сама по себе вызывающая у здоровых животных гиперфункцию железы, привела у облученных крыс к состоянию, близкому к ее истощению. При дефиците гормонов периферических желез (на примере недостаточности щитовидной железы) выявляется временная связь с рядом проявлений как радиационных последствий, так и нарушений, не имеющих отношения к радиации. Высказано мнение, например, о том, что ускоренное разрушение эритроцитов может быть не связано с качеством их, а обусловлено недостатком гормонов в организме. Известно, что при гипотиреоидных состояниях продолжительность жизни эритроцитов уменьшается почти в 2 раза по сравнению с продолжительностью их жизни у здорового человека. Клиницисты-гематологи выделяют специальную форму анемии — тиреопривную анемию, развивающуюся на почве гипотериоза и микседемы, основным лечением которой является введение в организм тиреоидных гормонов. При этом для нас важным представляется тот факт, что эритроциты больных микседемой, перелитые здоровым реципиентам, стали иметь нормальную продолжительность жизни. Обнаружено также, что и при ряде других болезней, сопровождаемых укорочением продолжительности жизни эритроцитов, переливание их в здоровый организм обусловливало нормальную продолжительность их жизни (серповидноклеточная анемия, нефриты) и, наоборот, переливание эритроцитов от здоровых людей в организм больных укорачивало продолжительность их пребывания в русле крови (ревматоидный артрит, острые инфекционные процессы). Все это свидетельствовало о том, что основной и непосредственной причиной укорочения продолжительности жизни эритроцитов в данных случаях было не ухудшение их качества, а не связанные с ними факторы организма. Эти факторы могут быть разными, но в абсолютном большинстве случаев они непосредственно связаны с селезенкой, с гиперфункциональным ее состоянием. Для нас важно, что действие этих факторов по времени совпадает с недостаточностью периферических эндокринных желез. Указанные в этом разделе отдаленные радиационные последствия, в том числе и злокачественные, не отличаются от таких же заболеваний, вызванных другими факторами. Более того, они характерны для возрастной патологии и часто возникают только тогда, когда возраст организма приблизился к периодам проявления аналогичной спонтанной патологии (дегенеративно-дистрофические нарушения, склеротические, циррозы печени, нефросклерозы, неопластические заболевания и т. п.). В этот период также наблюдаются определенные возрастные изменения в эндокринной системе. Изложенное породило специальные направления исследований, таких, например, как «Старение, климакс и рак» [Дильман, 1968] и «Эндокринологическая онкология» [Дильман, 1974].Возрастные изменения и гормональная регуляция
Два обстоятельства в исследовании возрастных изменений обращают на себя внимание: 1) некоторая общность проявлений отдаленной радиационной и возрастной патологии; 2) взаимосвязь многих проявлений возрастной патологии с состоянием эндокринных желез. Старение — процесс универсальный, характеризующийся своими внутренними закономерностями и захватывающий все системы организма. Появляется функциональная неполноценность клеток разного типа, уменьшается их число и на оставшиеся ложится повышенная функциональная нагрузка. Существенно изменяются реактивность клеток, их чувствительность к регуляторным влияниям. Возрастные изменения затрагивают процессы генерации энергии и ограничивают ее возможности. На примере мышечных волокон показано, что при старении наступает перераспределение различных путей энергетического обеспечения клеток, все большее значение начинают приобретать гликолитические процессы. Уменьшается число белковых рецепторов на мембране, и изменяется их состояние, зависимое от фосфолипидного окружения. В. В. Фролькис считает, что частичная дерецепция клеток при старении нарушает межклеточные взаимоотношения и участие клеток в системных, общерегуляторных реакциях, что является важным механизмом расстройства их функции. Многие изменения в клетках в процессе старения сходны со сдвигами в денервированной ткани. Ослабляется нервный и гуморальный контроль деятельности клеток. Однако в ответ на возрастание изменения и падение способности клеток отвечать адекватной реакцией развиваются в первый период старения достаточно эффективные компенсаторно-приспособительные и адаптивные реакции. Так, известно, что в этот период происходит возрастание чувствительности клеток некоторых органов к катехоламинам, может наблюдаться даже рост содержания в них цАМФ. Тем не менее общие адаптационные возможности организма и его важнейших систем к старости значительно сужаются. Возрастные изменения в таких системах, как сердечно-сосудистая, дыхательная и пищеварительная, способствуют развитию явлений кислородного голодания организма и недостатка ряда субстратов обмена. Наиболее ярко видны возрастные изменения в иммунной и эндокринной системах. Прогрессивное уменьшение массы лимфоидной ткани после рождения и на протяжении всей последующей жизни хорошо известно. Это заметно по уменьшению тимуса, лимфатических узлов, селезенки и числа лимфоцитов в крови. Наблюдения на людях и в экспериментах на животных дали много доказательств связи между болезнями старения и нарушением иммунных функций. В эту группу заболеваний включают воспалительные заболевания, васкулярные и особенно кардиоваскулярные и почечные заболевания, так называемые дегенеративно-сосудистые заболевания, аутоиммунные заболевания, предрасположенность к инфекционным и злокачественным заболеваниям и амилоидоз.| Возраст | Число лиц с аутоантителами, % | Средний титр |
|---|---|---|
| 12-14 лет | Нет | Нет |
| 17-22 года | 13 | 1 : 2 |
| 46-59 лет | 38 | 1 : 6 |
| 60—75 лет | 50 | 1 : 10 |
Общие закономерности изменений клеток при их ускоренном размножении
Наиболее подробно эти закономерности изложены в монографии И. Г. Акоева и Н. Н. Мотлоха «Биофизический анализ предпатологических и предлейкозных состояний» [1984]. Ниже конспективно изложены материалы из этой монографии.Нарушения клеточного цикла пролиферирующих тканей
Развитие предпатологических и патологических состояний в той или иной форме связано с изменениями нормального соотношения процессов клеточного размножения и специализации (дифференцировки). Нарушение регуляции этих процессов предшествует и развитию опухолей. Функция дифференцировки присуща всем живым клеткам. При развитии и совершенствовании многоклеточных организмов клетки дифференцируются с целью обеспечения выполнения в интересах целого организма так называемых специфических функций, т. е. тех функций, в прямых или косвенных результатах которых нуждаются другие клетки организма, прежде всего клетки иных типов дифференцировки. В настоящее время считается доказанным, что ядро дифференцированной клетки в значительной мере сохраняет мультипотентность, т. е. способность давать информацию, необходимую для развития всех потенциально возможных функций клетки. Процесс дифференцировки, вероятно, происходит главным образом через регулируемую извне репрессию—дерепрессию последовательно сменяющихся ведущих генов, т. е. эпигенетическим путем, а не за счет главенствования необратимых изменений генетического аппарата. Этот во многом не решенный вопрос общебиологической значимости имеет самое непосредственное отношение и к проблеме опухолевой трансформации. Такой эпигенетический характер регуляции предполагает, что процесс дифференцировки клетки, исходно развертываемый на основе наследственного материала, через посредство внутриклеточных медиаторов должен управляться внеклеточными факторами, или репрессорами, будь то ионный состав среды, наличие в ней определенных химических соединений, контактных механических взаимодействий и др. Правильный запуск любого очередного этапа дифференцировки клетки вообще невозможен без адекватной реализации обратной связи структурных генов с внеядерной и внеклеточной средой (в их широком понимании) для предшествующего этапа. Существует определенный антагонизм между функциональной и митотической активностью, между процессами дифференцировки и пролиферации. На тканевом уровне, как и на уровне клеточных популяций, обратная зависимость между степенью дифференцировки и пролиферативной активностью остается в целом верной. Известно важное обстоятельство, вытекающее из указанной закономерности: по мере дифференцировки снижается реализуемая способность клеток к делению. Достигнув некоторой пороговой степени дифференцировки, клетки могут вообще перестать делиться и продолжают далее дифференцироваться без митозов, как это происходит у млекопитающих с корковыми нейронами и миоцитами. Потеря способности клеток к делению часто есть следствие дифференцировки клетки, заключающееся в утрате еще одной клеточной функции. Применение различных способов получения синхронно делящихся клеточных популяций и метода радиоавтографии привело к обнаружению морфологически неявно различимых периодов клеточного онтогенеза. Эти периоды, обозначенные как G1 (пресинтетический — до основного синтеза ДНК), S (синтетический — идет синтез ДНК) и G2 (постсинтетический, или премитотический, — подготовка к делению клетки), совместно с митозом в представленной последовательности составляют митотический цикл. Величина пролиферативной активности определяется не только количеством делящихся клеток, но и скоростью их продвижения по периодам митотического цикла. Для большинства растущих клеточных линий и тканевых культур интервал между делениями, или длительность цикла, составляет 10—30 ч. Наибольшее количество экспериментальных работ проведено на культивируемых клетках Hela. Время генерации этих клеток около 24 ч. В среднем они находятся 15—16 ч в период G1, 6—7 ч — в стадии синтеза ДНК (S-период), 2 ч — в периоде G2 и митозе. Большинство исследователей пришли к убеждению, что для различных клеточных типов изменения продолжительности митотического цикла в основном происходят за счет вариаций его начального, пресинтетического G1-периода. Его длительность меняется от неуловимо малых значений до нескольких суток и более. Клетки могут находиться в двух альтернативных состояниях — в митотической активности или покое. В последние годы к покоящимся принято относить клетки, которые неопределенно долгое время могут не размножаться и при этом полностью сохранять как жизнеспособность, так и способность к пролиферации (т. е. к делению) независимо от степени своей специфической функциональной нагрузки. Покоящиеся клетки всего лишь часть непролиферирующей фракции клеточной популяции. Выход клетки в состояние пролиферативного покоя не является необратимым. В определенных физиологических или патологических ситуациях клетки могут вернуться в митотический цикл. Однако по мере пребывания клетки в состоянии покоя ее метаболизм становится все более консервативным и требуется все больше усилий и времени, чтобы инвертировать его на пути деления. Во время продвижения клеток по циклу от одного митоза до другого встречается важнейшая критическая фаза в периоде G1, в которой осуществляется выбор клеткой дальнейшей судьбы — идти к делению или к дифференцировке. Она находится в середине или в завершающей части периода G1. В целом по мере клеточной дифференцировки в пределах сформированных органов в течение эмбриогенеза и в ходе постнатального онтогенеза темп репродукции клеток постепенно уменьшается. При этом продолжительность каждой последующей генерации клеток, как правило, возрастает (например, в ходе дифференцировки клеток миелоидного ряда — с 25 до 604 ч), а пролиферативный пул и матричная активность ДНК снижаются. Вместе с тем в любой стадии развития существуют промежуточные, бластные элементы с интенсивной пролиферацией и коротким циклом: нейробласты, миобласты, эритробласты и др. Между возрастом животного и средней продолжительностью цикла существует определенная зависимость. Так, пролиферативный пул, рассчитанный для зачатка печени, составил 64,1%. С возрастом эта величина уменьшается, составляя у новорожденных крыс около 39%, через 21 сут от рождения — 15, через 51 сут — около 3%. Максимальный пролиферативный пул при регенерации равен 69,3%. Эта величина близка к величине пула в зачатке печени эмбрионов, равной 64,1%. В соответствии с этим эмбриональная и регенерирующая печень имеет схожие короткие времена средней продолжительности митотического цикла, которые близки к этому показателю для быстро делящихся клеток других эмбриональных тканей, злокачественных опухолей, клеток нормальной слизистой оболочки кишечника. Следует подчеркнуть, что и кинетика процессов пролиферации в зачатке печени и при ее регенерации в зрелом организме также имеет значительное сходство.Изменения клеточного цикла после воздействия радиации
Многочисленными исследованиями установлено значительное нарушение митотического цикла клеток при воздействии на них разнообразных факторов. Наиболее полно изучено действие ионизирующих излучений. Первоначальная задержка клеточного деления и соответствующее удлинение митотического цикла при воздействии радиации сменяются в дальнейшем сокращением продолжительности митотического цикла. Имеется большое число исследований, направленных на изучение реакций отдельных периодов клеточного цикла на воздействие радиации. Для разных линий клеток получены различные данные. Обобщение этих данных, проведенное во многих работах, показало, что для большинства изученных клеток в культуре митоз и конец периода G1 (переход в период S) наиболее чувствительны к действию радиации, а период S — наиболее устойчив. Задержка деления клеток наиболее значительна, если облучение осуществляли во время митоза, и мала для клеток, облученных в период S. Однако имеются линии клеток, где соотношение задержек периодов клеточного цикла иное. Обращено внимание также на то, что реакция клеток на облучение в различные периоды клеточного цикла прямо противоположна, если сравнивать задержку деления клеток и их гибель: чем больше задержка деления, тем меньше гибель клеток. Так, клетки, облученные в периоде S, могут иметь высокую выживаемость потому, что они на длительный срок задерживают свое деление. В этот удлиненный период до деления клетки могут успеть отрепарировать возникшие в них повреждения. Обращено внимание на то, что радиозащитное средство цистеамин не оказывает защитного эффекта при воздействии радиации в период G1 в отличие от облучений во время других периодов клеточного цикла. Данные о влиянии радиации на митотический цикл клеток опухолей in vivo противоречивы, что объясняют различным временем проведения исследований после облучения. Работая на тех же клетках четырехсуточной асцитной фибросаркомы, обнаружили сначала удлинение митотического цикла (в 2 раза через 20 ч после облучения), а затем укорочение (через 48 ч). Если исследовать клеточный цикл асцитной карциномы Эрлиха сразу после облучения, то наблюдается увеличение продолжительности периодов G1 и G2 в 2 раза; продолжительность митоза не изменена. Значительно меньше данных об изменении стадии митотического цикла in vivo у млекопитающих при действии радиации. Прежде всего выяснилось, что периоды G1 и G2 наиболее сильно реагируют на воздействие радиации, а периоды М и S — мало. При усилении пролиферативной активности тканей организма сокращения продолжительности митотического цикла в основном происходят за счет периода G1. В опытах на собаках, подвергавшихся длительному слабому хроническому воздействию, показано, что средняя продолжительность генерационного цикла миелоидных элементов может сокращаться в 2 раза за счет сокращения всех периодов интерфазы. В эритроидном ростке (эритробласты) в этот период сокращение генеративного цикла не было столь значительным и происходило только за счет укорочения периода G1. При одновременном исследовании митотического цикла у клеток кишечника и костного мозга установлено, что изменения продолжительности цикла клеток кишечника были менее выраженными. По-видимому, это является отражением того факта, что клетки кишечника делятся чаще, чем бластные клетки костного мозга. Возможности к сокращению интерфазы клеток кишечника меньше, чем у клеток костного мозга. Имеется взаимосвязь между нормальной продолжительностью митотического цикла клеток и чувствительностью их к хроническому облучению — чем короче интерфаза митотического цикла, тем чувствительнее клетки к радиации. Это обусловлено продолжительностью периода G1. Дальнейший наш анализ возможной роли периода G1 в определении устойчивости к радиации показал много интересного. Широко распространенный феномен приостановки клеточного деления в пролиферирующих тканях (первоначальная реакция) относится к более общей реакции биологических систем в ответ на повреждение. Происходит временное ингибирование многих функций, не необходимых для репарации (восстановления) повреждений, и отдаление по времени переходов в другие функциональные состояния, когда эти невосстановленные повреждения могут проявиться неблагоприятным для биологической системы эффектом. Такое однотипное поведение биологических систем наблюдается на разных уровнях: организм, физиологическая система, клеточная система, ткань. Продолжительность возможных задержек периодов клеточного цикла зависит не только от характера ткани, к которой клетки относятся. Так, в эпителии роговицы глаза мышей в период G1 возникают более продолжительные задержки, чем в эпителии кишечника, в расчете на единицу дозы нейтронного излучения: для эпителия роговицы — 17 мин на 0,01 Гр, а для эпителия кишечника — только 4 мин на 0,01 Гр. Продолжительность митотического цикла клеток эпителия роговицы в норме в 6,5 раза больше, чем у клеток эпителия кишечника. К тому же клетки эпителия роговицы способны блокировать митотический цикл в периоде G1 на значительно большие сроки, чем это возможно для эпителия кишечника. Установлено, что способность к удлинению продолжительности интерфазы (в основном в периоде G1) митотического цикла является одним из основных условий эффективного завершения репарации лучевых повреждений. Существует определенная пропорциональность продолжительности блоковых задержек периодов интерфазы митотического цикла и интенсивности повреждения. Четкие доказательства последовательности выхода клеток из блоков различных периодов интерфазы митотического цикла, зависимость продолжительности их от дозы облучения, относительное значение блоковых задержек были получены в нашей лаборатории В. Г. Тяжеловой и др. Основной результат аналитических исследований представлен в виде пространственной трехкоординатной картины дозовых кривых митотической активности, позволяющих извлекать ряд интересных закономерностей. Всего выделено пять пиков митотической активности. Экстраполяция дозовых зависимостей к нулевой дозе характеризует нормальную продолжительность периодов митотического цикла: G1 — 8 ч, S — 8, G2 — 1 и М — 1 ч при суммарной длительности цикла 18 ч. Это соответствует данным, полученным другим путем для эпителия кишечника крыс. Длительность блока в данном эксперименте учитывается через максимум митотической активности, который здесь представляет собой сумму нормальной длительности периода и блоковой задержки. Первые три пика соответствуют первому послеблоковому делению клеток. Выход клеток из блоков происходит в естественной последовательности. Раньше клетки выходят из блоков тех периодов, которые в момент облучения были ближе по ходу естественного созревания клеток к периоду митоза. Первая кривая характеризует выход из блока G2M, вторая — из блока SG2, третья — из блока G1S. С увеличением дозы облучения длительность всех периодов интерфазы митотического цикла растет, но не беспредельно. Она стремится к некоторому пределу. С одной стороны, удлинение периодов интерфазы необходимо для репарации увеличенного объема повреждений с увеличением дозы облучения, с другой — клетка, по-видимому, не может находиться неограниченное время в каком-либо одном периоде интерфазы и задерживать естественный для нее процесс созревания. Для каждого периода развития клетки характерно преобладание одних структурнофункциональных процессов и ингибирование других. В этом причина ограничения продолжительности блоковых задержек периодов интерфазы митотического цикла. При этом скорость достижения указанного предела для разных периодов неодинакова. Она последовательно уменьшается для блоков G2M, SG2 и G1S. Длительность периодов с учетом блоковых задержек после облучения максимальна для G1, меньше для S и еще меньше для стадии G2. В то же время относительные изменения периодов (в процентах от нормальной продолжительности стадии) иные: максимальное для G2, менее сильное для G1 и слабое для периода S. Сравнительный анализ площадей, охватываемых кривыми митотической активности, дал возможность при некоторых допущениях (число клеток пропорционально площади) оценить относительное число выживающих клеток, облученных в определенном периоде митотического цикла, и построить дозовые кривые выживаемости. Клетки, облученные в периоде S, не погибали, пока доза облучения не превысила ~5 Гр, Даже после облучения в дозе 9 Гр еще выживало не менее половины клеток. Кривая выживаемости для клеток, облученных в периоде G2, имеет небольшое плечо, затем число выживающих клеток снижалось и достигало примерно 50% в районе дозы облучения 5 Гр. Поведение клеток, облученных в стадии G1, существенно отличалось. Гибель клеток происходила при любой дозе и достигала примерно 50% при облучении в дозе порядка 3,5 Гр, а при облучении в дозе 9 Гр оставалось в живых лишь около 20% клеток. Наибольшие различия обнаруживаются при сравнении доз облучения, вызывающих небольшой процент гибели. Так, для того чтобы получить примерно одинаковую 80%-ную выживаемость клеток, необходимо облучать клетки в периоде S в дозе около 7,5 Гр, в периоде G2 — в дозе около 2,5 Гр и в периоде G2 — в дозе 0,5—1,0 Гр. Хотя эти выводы были получены на основе анализа ограниченного материала (эпителий тонкого кишечника крыс при воздействии на них радиации с мощностью дозы около 0,4 Гр/мин), они позволяют делать более общие выводы, так как не противоречат отрывочным данным других авторов. Итак, проводя дальнейшие сопоставления полученных выводов для эпителия кишечника крыс, необходимо обратить внимание на следующее. 1. Период S среди других периодов интерфазы митотического цикла обладает наибольшей устойчивостью к радиации и имеет на кривой выживаемости плечо, свидетельствующее об огромных возможностях репарационных процессов, характерных для этой стадии. Однако при воздействии радиации период S имеет наименьшее относительное увеличение своей продолжительности за счет блоковых задержек и по достижении предельно возможной длительности блоковой задержки начинается гибель клеток. При этом кривая уменьшения выживаемости при дальнейшем увеличении дозы облучения имеет наибольший наклон. При ускорении процессов деления клеток период S относится к наименее укорачиваемому. 2. Период G1 отличается наибольшей чувствительностью к радиации и не имеет на кривой выживаемости плеча, которое свидетельствовало бы о сколько-нибудь заметных возможностях репарационных процессов, характерных для этого периода. Однако период G1 при действии радиации имеет наибольшее абсолютное увеличение своей продолжительности и не имеет ограничений в этом (по крайней мере, до дозы 9 Гр нет такого ограничения). Гибель клеток отмечена даже при дозах менее 1 Гр. При ускорении процессов деления период G1 наиболее вариабелен и может весьма существенно укорачиваться. В случае сокращения генеративного цикла это происходит в основном за счет периода G1. 3. Период G2 наиболее короткий в норме, и увеличение его при воздействии радиации также небольшое, хотя в процентном отношении и является наибольшим. Этот период быстрее других выходит на предел возможного увеличения своей продолжительности. По количеству клеток, погибающих после облучения в периоде G2, он занимает промежуточное положение между периодами S и G1. Изменения длительности этого периода не отражаются существенно на общей продолжительности митотического цикла. Интересные данные получены Ямагучи и Табачником при исследовании кинетики клеток эпидермиса кожи морских свинок в норме и после локального воздействия бета-радиации. Основные исследования проводились в период 12—14 сут после облучения, когда вслед за клеточной депопуляцией развивалась активная пролиферация и отмечался наиболее высокий митотический индекс. Результаты исследований приведены ниже. Указана продолжительность всего цикла и отдельных его периодов в часах.| G1 | S | G2 | M | Сумма | |
|---|---|---|---|---|---|
| Без облучения | 76-77 | 9,3-9,6 | 5,0-5,6 | 0,4-0,6 | 91-92 |
| После облучения | 3-4 | (5,7-7,2) | 2,3-2,6 | (0,4-0,6) | 16 |
| Во сколько раз сокращено | ~20 | ~1,5 | ~2 | 1,0 | ~5 |
Изменения клеточного цикла в предопухолевых состояниях и в опухолях
При канцерогенезе на первый план всегда выступают количественные изменения пропорции дифференцирующихся и размножающихся клеток, связанные с уменьшением доли дифференцирующихся клеток среди дочерних генераций. Интенсивность пролиферации злокачественной ткани часто превышает нормальную этих же тканей. Это различие, однако, не относится к числу абсолютных критериев. Известно достаточное количество примеров обратных взаимоотношений, когда скорость деления злокачественных клеток снижена по сравнению с исходными нормальными значениями. К тому же в массе опухолевой ткани могут существовать и длительно не пролиферирующие клетки, пребывающие в состояниях покоя или дифференцировки. Опухоли чаще всего возникают в местах длительной и (или) интенсивной физиологической или репаративной (раневой) регенерации, сопровождаемой увеличением клеточной массы. Между канцерогенезом и предшествующей ему длительной и (или) интенсивной пролиферацией существует, как правило, положительная корреляция. Однако после завершения малигнизации пролиферативная активность может быть разной. Важная пролиферативная характеристика гетерогенности популяции — коэффициент вариации длительности митотического цикла составляющих ее клеток. Вариабельность длительности цикла и отдельных его периодов увеличена для клеток в ходе предопухолевой активной пролиферации и в злокачественных опухолях. Нормальные обновляющиеся ткани, обладающие очень высокой митотической активностью, имеют минимальный резерв ее дальнейшего увеличения. Поэтому стимулы, направленные на дальнейшее увеличение интенсивности деления клеток этих тканей, в процессе канцерогенеза могут вести к срыву компенсаторного резерва, который находится и в норме «на пределе», и в результате к концу канцерогенеза — к уменьшению скорости деления. В тканях, в которых скорость нормального размножения клеток далека от максимальной, величина компенсаторного резерва пролиферативной активности может оказаться достаточно высокой. Поэтому в этих тканях неэкстремальные стимулы к интенсификации клеточного деления вызывают в них прямой эффект усиления митотической активности, сохраняющейся и после окончания канцерогенеза. Время генерации пролиферирующих лейкозных бластов обычно значительно меньше, чем ранних кроветворных предшественников, т. е. они размножаются быстрее и с укороченным митотическим циклом. Вероятность опухолевой трансформации наиболее велика для клеток активно пролиферирующих тканей. Возникновение процесса трансформации клеток, по нашему мнению, можно рассматривать как результат естественной метаболической перестройки клетки в ответ на внешние для нее воздействия и условия, вызывающие длительную и непрерывную стимуляцию клеточного деления, происходящего при сокращении митотического цикла клеток и особенно периода G1.Биохимические и биофизические изменения, связанные с усилением пролиферативной активности
Имеется большое количество книг и обзоров, рассматривающих биохимические процессы в тканях и популяциях клеток со стимулированной пролиферацией. Существуют общие закономерности, свидетельствующие об одинаковой направленности большинства биохимических изменений в эмбриональных, регенерирующих и опухолевых тканях по сравнению с нормальной тканью, хотя встречаются и отдельные исключения. Пролиферирующие активно делящиеся клетки обычно отличаются усиленным расходованиемлипидов, ускоренным гликолизом, нуклеосинтезом и протеосинтезом, повышением активности пентозофосфатного шунта. В таких клетках, как правило, соотношение циклаз смещается в сторону гуанилатциклаз, соотношение циклонуклеотидов — в сторону цГМФ над цАМФ. Соответственно увеличивается активность цГМФ-зависимых протеинкиназ и уменьшается активность цАМФ-зависимых протеинкиназ. В митохондриях увеличивается активность АТФаз и снижается активность АТФ-синтетаз, накапливаются ионы Ga2+ и уменьшается содержание ионов Mg2+. Активность НАДФ-зависимых дегидрогеназ повышается, а НАД-зависимых — падает. В цитоплазме уменьшается соотношение белковых SH- и S—S-групп, а в ядре и ядрышке — увеличивается. В отношении небелковых SH- и S—S-групп картина обратная. При длительном усилении пролиферативной активности ткани и снижении степени дифференцировки составляющих ее клеток однозначно увеличивается активность систем, определяющих основные биологические характеристики делящихся клеток и являющихся более древними в эволюционном плане. Одновременно или несколько позже в таких случаях снижается активность эволюционно более молодых систем, связанных с ведущими признаками дифференцированного состояния клетки. Эта закономерность общая для всех ускоренно размножающихся клеток, включая и клетки злокачественных опухолей. Не обнаружено никаких специфических опухолевых ферментов или каких-либо белков, липидов, гликолипидов или других химических соединений, которые не были бы свойственны нормальным клеткам на различных этапах их жизненного пути. При длительном состоянии ускоренной пролиферации любых клеток происходит определенная редукция клеточного метаболизма. Так, Уоррен и Бак отметили, что злокачественные, активно пролиферирующие клетки содержат больше гликолипидов с короткими углеводными цепями, чем нормальные (из-за сниженного содержания сиаловой кислоты), но меньше гликолипидов с длинными цепями. Такие клетки в силу укорочения митотического цикла не успевают завершать синтез углеводных компонентов своих гликолипидов. Уменьшено среди белков и гликопротеидов поверхности клеток количество фибронектина и коллагена, неспецифических и адгезивных гликопротеидов как при стимуляции пролиферации в норме, так и при опухолевом росте. Подобные аномалии могут нести к снижению чувствительности клеток к контактному угнетению митотической активности и к изменению иммунной специфичности гликопротеинов клеточной поверхности. При длительном состоянии ускоренной пролиферации в существующих в норме последовательностях реакции, преимущественно имеющих специализированное значение, возможно также выпадение промежуточных звеньев. Оно начинается со снижения активности и содержания фермента, а затем приводит к исчезновению последнего. Соответственно происходит упрощение состава метаболитов и укорочение метаболического пути. Клетка в связи с ускоренным размножением как бы освобождается от уже ненужных метаболических реакций. Некоторые специализированные ферменты, принимающие участие в функции нормальных тканей (аргипаза, каталаза, цитохромоксидаза, цитохром с, эстераза и др.), в опухолях отсутствуют, или их активность очень низка. Иными словами, нарушается или утрачивается специфический ферментный профиль. Например, в митохондриях с увеличением скорости роста гепатом Морриса появляется дефицит моноаминооксидазы на внешней мембране, аденилатциклазы в межмембранном пространстве и глутаматдегидрогеназы в матриксе. Аналогичные данные получены и для нормальной печени, регенерирующей после частичной гепатэктомии. Некоторые опухоли имеют митохондрии с недостатком одного или двух компонентов дыхательной цепи с дефицитом ферментов клеток крови. При редукции метаболизма в ускоренно размножающихся клетках в реакциях катаболизма (распада) происходит снижение эффективности работы метаболической цепи в целом. В реакциях анаболических (синтеза) в условиях энергетического дефицита может происходить неполное завершение последовательности реакций и(или) переход к «крупноблочному строительству», т. е. утилизации высокомолекулярных недорасщепленных субстратов. Сокращение этапов синтетических процессов наиболее вероятно в условиях обостренной конкуренции за макроэргические соединения, выработка которых может затрудняться падением общей продуктивности митохондриального окислительного фосфорилирования при редукции митохондриального аппарата либо при внутритканевой гипоксии в далеко зашедших пролиферативных состояниях. Так, есть некоторые экспериментальные доказательства возможности использования активно делящейся клеткой крупных блоков для синтеза ДНК. Например, в опухолевых клетках по сравнению с нормальными в десятки раз более активны ферментные системы, направляющие биосинтез нуклеотидов на энергетически выгодные пути утилизации готовых пуриновых и пиримидиновых оснований без предварительного их синтеза. Такой механизм может обеспечить клетке повышенную скорость обновления нуклеиновых кислот независимо от наличия простых предшественников энергоемкого синтеза азотистых оснований (из глицина, аспартата, глутамина, формиата, рибозы). В то же время «крупноблочное строительство» может уменьшить эффективность репарации ДНК и увеличить накопление в ней ошибок. В быстро пролиферирующих тканях и опухолях низка активность не только деполимераз нуклеиновых кислот, но и ряда катаболических ферментов пиримидинового, пуринового и аминокислотного обмена. Создаются условия для снижения субстратной специфичности ферментов. Известны случаи замещения аминокислот в полипептидных цепях в процессе их синтеза, вероятность которых с усилением пролиферации нормальных клеток, как мы предполагаем, может повышаться. Такие же замены аминокислот часто делают ферменты более чувствительными к тепловой денатурации. При этом доля термолабильной фракции белков возрастает. С другой стороны, термолабильная фракция ферментов обладает сниженной субстратной специфичностью и реагирует с аналогами субстратов с большей эффективностью, чем нормальные ферменты катализа. Немногочисленные биофизические данные также свидетельствуют о существенной роли нарушений содержания элементов кальция, хлора, меди, йода, железа, магния, марганца, фосфора, калия, натрия, цинка и соотношений их концентраций в процессах пролиферации в дифференцировки и опухолевой трансформации. По мнению ряда ученых, ключевая роль в этом процессе принадлежит ионам Mg2+. Удаление Mg2+ из среды инкубации сказывалось в торможении синтеза ДНК, а снижение концентрации Са2+ действовало на синтез только при одновременном четырехкратном уменьшении содержания Mg2+. Предполагается, что роль кальция в регуляции деления клеток опосредована магнием. В настоящее время наибольшее признание имеет точка зрения, согласно которой модификация ферментов, участвующих в обмене циклических мононуклеотидов, и внутриклеточная концентрация последних являются одними из ранних реакций, связанных со стимулированием пролиферации или дифференцировки. В различных нормальных и опухолевых клеточных популяциях и тканях отмечается общая направленность изменения циклазных систем при увеличении содержания пролиферирующих и дисдифференцированных клеток. Это увеличение отношений цГМФ/цАМФ, гуанилатциклаза/аденилатциклаза, цГМФ-зависимые протеинкиназы/цАМФ-зависимые протеинкиназы. В нормальных дифференцированных тканях взрослых животных активность аденилатциклазной системы, как правило, существенно превышает активность гуанилатциклазной. С увеличением пролиферативной и особенно дисдифференцированной компонент клеток эти соотношения начинают возрастать в сторону гуанилатциклазной системы. В связи с этим широко распространено мнение, что цАМФ участвует в фосфорилировании белков, отвечающих за специализированную функцию дифференцированной клетки, а цГМФ — за фосфорилирование белков, участвующих в пролиферации. Для гепатом Морриса отмечена корреляция перечисленных изменений циклазных систем со скоростью роста. Интересны также данные о том, что гуанилатциклаза печени может стимулироваться некоторыми химическими канцерогенами. Так называемые связанные сульфгидрильные (SH-) и дисульфидные (S—S-) группы находятся в белках благодаря наличию в них серосодержащих взаимопревращающихся аминокислот цистеина (SH-) и цистина (S—S-), а свободные группы (если не учитывать SH-группы свободных аминокислот) принадлежат в основном глутатиону. Отмечено, что общеклеточный сдвиг равновесия групп SH↔SS влево стимулирует деление клеток, а смещение вправо — задерживает. В целом в клетках животных, растений и микроорганизмов для прохождения митоза необходима относительно высокая концентрация SH-групп. Таким образом, в ряде случаев прослеживается положительная корреляция между общим содержанием SH-групп в клетках и интенсивностью их пролиферации. Существенный вклад в это увеличение, по-видимому, вносит и синтез de novo глутатиона. Есть ряд данных, указывающих, что внутритканевое содержание убихинонов, токоферолов, некоторых стероидных гормонов, фосфолипидов и SH-содержащих белков при усиленной пролиферативной активности возрастает в нормальных и злокачественных клетках. Концентрация водородных ионов (pH) внутри и вне клетки является фактором, регулирующим направленность и интенсивность множества внутриклеточных процессов. Как в нормальных, так и в малигнизированных тканях в состоянии активной пролиферации нередко регистрируется снижение pH. Это особенно характерно для тех случаев, когда ускоренная пролиферация сопровождалась гипоксией и дисфункцией митохондриального аппарата, что способствует внутриклеточной аккумуляции метаболических кислот, например молочной кислоты. Измерения pH в раковых тканях показали, что они в среднем на 0.5 единиц pH имеют более кислую реакцию, чем нормальные ткани, хотя в отдельных случаях pH снижается на 1—2 единицы. Если нормальные клетки имеют внутриклеточный pH около 7,4, а раковые в обычных условиях ~7,0, то с усилением потребления последними глюкозы pH может достигать 5,8—6,0. С интенсификацией клеточного деления и соответственно снижением степени дифференцировки клеток, как правило, уменьшаются электропроводность, теплопроводность, мембранный потенциал покоя, ионселективность плазматической мембраны, степень кальцификации, механическая прочность мембран, степень адгезии (прилипания) в системах «клетка+клетка», «клетка+подложка», контактное торможение деления, антиокислительная активность липидов, вязкость цитозоля и повышаются перекисное окисление липидов, способность белков и ДНК к денатурации, неспецифическая проницаемость мембран, подвижность органических молекул в мембране, электрофоретическая подвижность, диффузия веществ внутри клетки, гидрофильность мембран (степень гидратации), степень текучести, «разжижения» мембран. Среди перечисленных характеристик нет ни одной, которую можно было бы строго доказательно отнести к специфическим признакам опухолевого роста. Все эти изменения обусловлены повышенной интенсивностью клеточной пролиферации. В условиях нормальной пролиферации они обратимы. Изменения не являются ведущими в опухолевой трансформации и в малигнизации, а служат функциональным отражением или сопутствующим фоном реакций исполнительного аппарата клетки в ответ на регулирующие пролиферацию сигналы и изменения окружающей ее среды. То же самое можно сказать и в отношении морфологических изменений. Изменения, выявляемые различными методами световой и электронной микроскопии, обычно тем глубже, чем выше уровень пролиферативной активности и степень нарушения дифференцировки. При длительной интенсивной пролиферации повышается количество аномалий клеточного деления как в нормальных, так и в опухолевых тканях. Электронная плотность клеток и части их органелл (митохондрии, ядерного матрикса, цитозоля) обычно снижается в состоянии активной пролиферации. Объем ядра, ядрышек и ядерно-цитоплазматическое отношение часто значительно увеличиваются. Возрастает количество пор в ядерной мембране. Понижена степень конденсации хроматина. Расширяются эндоплазматические цистерны. Соответственно уменьшается число связанных рибосом и увеличивается количество свободных рибосом. Размеры полисом сокращаются. Пониженное содержание гранулярной сети в эмбриональных, регенерирующих и других тканях с интенсивным делением клеток отражает тот закономерный факт, что их клетки заняты собственным воспроизводством в большей степени, чем выполнением специализированных функций для удовлетворения внеклеточного функционального запроса в данный момент. Это же обстоятельство объясняет снижение количества и размеров секреторных пузырьков и гранул, а также иногда наблюдаемую частичную редукцию секреторных структур аппарата Гольджи. Запасы резервных полимеров, капель липидов и особенно гранул гликогена в животных клетках резко истощаются. Эти морфологические наблюдения совпадают с биохимическими данными о существенной стимуляции гликолиза и липолиза при недостаточности соответствующих анаболических процессов. В среднем может быть увеличено количество и размеры некоторых цитоплазматических включений: липофусциновых гранул, миэлиновых фигур, Бизаровых телец и т. д. Предполагается, в частности, что липофусциновые гранулы (каротиноксисомы) могут способствовать активации реликтовых путей поддержания энергетического и окислительного обмена в условиях ограничения функции митохондрий и развивающейся в связи с этим клеточной гипоксии. Известно появление так называемой токсической зернистости нейтрофилов при состояниях, сопровождающихся при токсических отравлениях лейкопенией с последующей стимуляцией лейкопоэза. Увеличение числа дегенеративных клеток характерно для абортивного подъема числа лейкоцитов при лучевой болезни, когда наблюдается первый пик регенерации лейкопоэза и предполагается укороченным митотический цикл бластных клеток. Тельца Бизара появляются в предлейкозный период, когда обычна гиперплазия эритроидного ростка костного мозга. Претерпевают изменения и митохондрии: матрикс светлеет, часто объем их возрастает, количество крист уменьшается, содержание осмиофильных гранул повышается. Последнее обстоятельство имеет биохимическое подтверждение в увеличении количества внутримитохондриального кальция, что характерно как для регенерирующих, так и для опухолевых тканей. Относительно быстро истощается компенсаторный резерв окислительного фосфорилирования и становится невозможным его эффективное функционирование в связи с низким напряжением кислорода и по другим причинам. Митохондрии клеток быстро пролиферирующих тканей по сравнению с неделящимися более чувствительны к повреждению в различных неблагоприятных для клетки условиях. Уменьшается степень связи клеток в ткани, о чем свидетельствует увеличение размеров межклеточных пространств, снижение числа и размеров щелевых мостиков и других структур межклеточных контактов. Сокращение межклеточных контактов совпадает с изложенными биофизическими данными. В далеко зашедших случаях хронической ускоренной пролиферации может происходить потеря контактного торможения деления клеток. Важно обратить внимание на то, что в клетках, длительное время ускоренно размножающихся, уменьшается число специализированных выростов и выпячиваний плазматической мембраны, содержащих различные рецепторные комплексы на гормоны белковой природы, антигены, гормональные регуляторы и др. Этот признак является морфологическим отражением утраты клеткой многих поверхностных глюко- и протеоконъюгатов, падения чувствительности делящейся клетки к внешним регуляторным влияниям. Происходит относительная «регуляторная глухота», снижение чувствительности ко многим гормонам белковой природы, действующим на уровне внешней стороны плазматической мембраны. Средняя картина морфологической перестройки клетки в связи с длительной интенсификацией деления характеризуется сокращением количества и объема мембранных структур, выполняющих специализированные функции в интересах целого организма, и одновременно усилением структур, непосредственно связанных с функцией деления. Клетка морфологически упрощается и становится более похожей на независимо существующий одноклеточный организм (рис. 5). Все изложенное приближает изначально дифференцированную зрелую клетку к ее эволюционно более древним предкам. Сокращение общей площади мембран клетки сочетается с биохимическими и биофизическими данными о нарушении интегральных функций мембран и связанных с ними структур и первичных регуляторных процессов. Рис. 5. Цитологическая перестройка «идеальной» животной клетки при переходе от состояния «покоя» (специфической функции в дифференцированном состоянии) к пролиферации (снижению специфической функции и дисдифференцировке)
1 — ядро; 2 — ядрышко; 3 — хроматин; 4 — митохондрии; 5 — лизосомы; 6 — микротельца; 7 — секреторные пузырьки; 8 — агранулярный эндоплазматический ретикулум; 9 — гранулярный эндоплазматический ретикулум; 10 — гранулы гликогена; 11 — капли липидов; 12 — свободные рибосомы; 13 — связанные рибосомы; 14 — полисомы; 15 — гранулы неорганических веществ; 16 — центриоль; 17 — микроворсинки; 18 — межклеточные контакты; 19 — межклеточные пространства; 20 — базальная мембрана
Рис. 5. Цитологическая перестройка «идеальной» животной клетки при переходе от состояния «покоя» (специфической функции в дифференцированном состоянии) к пролиферации (снижению специфической функции и дисдифференцировке)
1 — ядро; 2 — ядрышко; 3 — хроматин; 4 — митохондрии; 5 — лизосомы; 6 — микротельца; 7 — секреторные пузырьки; 8 — агранулярный эндоплазматический ретикулум; 9 — гранулярный эндоплазматический ретикулум; 10 — гранулы гликогена; 11 — капли липидов; 12 — свободные рибосомы; 13 — связанные рибосомы; 14 — полисомы; 15 — гранулы неорганических веществ; 16 — центриоль; 17 — микроворсинки; 18 — межклеточные контакты; 19 — межклеточные пространства; 20 — базальная мембрана
Способность к пролиферации — древнейшее свойство клеточного уровня организации биологических систем. Специфические функции клетки, возникшие в ходе дифференцировки у многоклеточных организмов, относятся к более поздним эволюционным приобретениям. Эти функции связаны с разделением обязанностей и специализацией отдельных клеток в интересах целого организма. Переход клетки и ткани от выполнения специализированной функции в системе целостного организма, т. е. от дифференцированного состояния, к снижению дифференцировки и усилению пролиферации означает переход на эволюционно более древние и более устойчивые пути метаболизма. Обнаружение в активно или длительно пролиферирующих тканях каких-либо эмбриональных свойств (ферментов, антител и т. п.) следует рассматривать как проявление эволюционно-древних признаков.
Ошибки синтеза белка при усилении пролиферативной активности ткани
В процессе канцерогенеза могут появляться не характерные для данной нормальной ткани белки, изменяться спектры изоферментов и антигенов, что и используется для ранней диагностики опухолевых заболеваний. Однако указанные изменения оказались не строго специфичными, поскольку аналогичные белки, их модификации и аналогичные изменения спектров изоферментов и антигенов характерны как для периода эмбрионального развития, так и для периодов длительной и значительной активации размножения клеток нормальной ткани. Так, открытие в свое время α-фетопротеина как специфического ракового белка оказалось ошибочным. Позднее он был обнаружен и в нормальных тканях в случаях стимулированной пролиферации. В этих случаях также изменяется соотношение изоформ белков, что отражается в изменении, например, спектра гемоглобинов и спектра изоферментов. Изменяется и антигенная характеристика белков. Важно, что имеются общие особенности всех указанных изменений. 1. Эти изменения приводят к качественному подобию спектров изоформ белка нормальной ткани в состоянии активной пролиферации и нормальной регенерирующей ткани к спектрам изоформ белка эмбриональной и активно растущей злокачественной ткани. Такое же подобие наблюдается и по спектру антигенов. 2. Указанные изменения в нормальной ткани возникают вслед за стимуляцией пролиферативной активности и исчезают, как только пролиферативная активность нормализуется. Это не зависит от типа ткани. 3. Указанные изменения происходят и при воздействиях физических факторов на организм человека и животных, но только в тех случаях и в то время, когда происходит усиление пролиферативной активности ткани. Новые белки в этом случае, как правило, не появляются, но может происходить их модификация. Следовательно, во всех указанных случаях можно говорить прежде всего о генетической регуляции, о процессах репрессии — дерепрессии определенных локусов генома в зависимости от состояния клетки, об однотипности и неспецифичности процессов генной регуляции для разных тканей организма и при воздействии разных факторов, вызывающих состояние активной пролиферации ткани. Указанные изменения не требуют возникновения мутации, т. е. изменений в структурных генах. Необходимые процессы генной регуляции синтеза белка описаны достаточно подробно во многих руководствах, и на них мы останавливаться не будем. Однако механизмы возможной связи между изменением пролиферативной активности и изменением изоформ белков не ясны. В связи с изложенным следует рассмотреть ошибки рибосомального синтеза, не связанные и связанные с генетической регуляцией. Особое внимание было обращено на тот факт, что закономерные ошибки рибосомального синтеза белка обнаруживаются и в опытах in vitro в бесклеточной среде при отсутствии генной регуляции. Наиболее распространенное мнение о механизме работы рибосомы предполагает, что рибосома, состоящая из двух неравных субчастиц, ползет по матричной РНК от 5'-конца к 3'-концу, считывает информацию об аминокислотной последовательности и присоединяет соответствующие аминокислоты в полипептидную цепь. При этом малая субчастица рибосомы осуществляет контакт с мРНК. Полипептидная цепь собирается на особых центрах большой субчастицы. При этом механизм сборки полипептида, динамика процесса остаются неясными. А. С. Спирин и Л. П. Гаврилова предложили свою схему строения рибосомы, которая лучше соответствует наблюдаемым фактам, вскрывает движущие силы и механизм работы рибосомы. В основу положено разделение функций между субчастицами: большой отведена роль полимерного носителя, удерживающего последовательно наращиваемый пептид, а малой субчастице — роль «подносчика» аминокислот. Обе субчастицы соединены подвижным шарниром и связаны с одной и той же матрицей. Выделен рабочий цикл рибосомы, состоящий из пяти «шагов». Сначала «подносчик» связывается с мРНК, отодвигается от носителя (рибосома открывается) и вылавливает из цитоплазмы аминоацилтранспортную РНК, соответствующую очередному кодону матрицы. Пока рибосома открыта, совершается перебор соответствия кодонов антикодонам и, следовательно, правильного выбора очередной аминокислоты. Когда аминокислота выбрана, рибосома закрывается и приводит аатРНК в контакт с наращиваемым полипептидом и очередная аминокислота (точнее, ее остаток) занимает свое место в цепи, а освобожденная тРНК уходит в цитоплазму и находит себе новую молекулу этой же аминокислоты. Периодическое размыкание и смыкание рибосомальных субчастиц является приводным механизмом, обеспечивающим все перемещения тРНК, мРНК и аминокислот в процессе синтеза пептидной цепи. Каждому кодону мРНК соответствует антикодон на аатРНК. Каждый кодон состоит из триплета нуклеотидов (разные сочетания урацила, аденина, цитозина и гуанина). Каждая из 20 аминокислот имеет характерные только для нее кодоны (табл. 3). Например, тирозин кодируется следующими кодонами: УАУ (урацил—аденин—урацил) и УАЦ (урацил—аденин—цитозин). Метионин кодируется одним кодоном, в то время как ряд других аминокислот — большим числом, например шестью кодонами (лейцин, серин, аргинин). Для каждой аминокислоты все кодирующие ее кодоны равнозначны. Рибосомальный синтез белка оказался чрезвычайно устойчивым к непосредственному действию радиации. Однако его интенсивность очень чувствительна к регуляторным влияниям организма. В соответствии с предложенным А. С. Спириным механизмом работы рибосомы стали в определенной мере понятны и молекулярные механизмы известных давно ошибок рибосомального синтеза белка, зависящие от самой рибосомы. Известно, что противобактериальное действие ряда аминоглюкозидных антибиотиков (стрептомицин, канамицин, неомицин и др.) связывается со способностью их вызывать ошибки кодирования рибосомального синтеза белка, и это было показано экспериментально на бесклеточных системах. В работах А. С. Спирина и его коллег было изучено влияние указанных антибиотиков на цикл работы рибосомы и показано, что все они ускоряли стабилизацию ассоциированных субчастиц рибосомы. Кроме того, установлено, что бивалентные ионы магния, кальция, марганца и ряд некоторых агентов также увеличивали (ускоряли) стабилизацию (т. е. смыкание) субчастиц рибосомы. Так, изменение концентрации магния на 1 мМ существенно (примерно на 10%) изменяло вероятность нахождения рибосомальных субчастиц в связанном состоянии (т. е. в сомкнутом). Наоборот, одновалентные катионы натрия, калия и других уменьшали вероятность нахождения субчастиц рибосомы в сомкнутом состоянии. Увеличение концентрации их в бесклеточной среде примерно на 10 мМ уменьшало долю связанных субчастиц на 10—20%. Все факторы, которые ускоряли стабилизацию субчастиц рибосомы, способствовали увеличению ошибок кодирования рибосомального синтеза белка. Это было особенно четко показано в экспериментах с бактериальными рибосомами в бесклеточной среде на примере использования полиуридиновой матрицы, кодирующей аминокислоту фенилаланина. Вместо фенилаланина рибосома в небольшом проценте случаев включала в полипептидную цепь лейцин и изолейцин и значительно реже серин, тирозин и валин. Однако при добавлении в среду, например, стрептомицина и ионов магния ложное кодирование, определявшееся по отношению включенного в полипептидную цепь лейцина к фенилаланину, увеличивалось с 5 до 32%, т. е. в 6 раз. Особенно сильно влияние ионов магния и натрия. Так, по кривым зависимости числа ошибок включения лейцина видно, что изменение концентрации Mg с 1,0 до 2,0 мМ увеличило ошибки более чем в 3 раза (до 17%). В терминах модели А. С. Спирина повышенная вероятность сомкнутого состояния рибосомы должна приводить к повышению ложного кодирования. Все факторы, ускоряющие стабилизацию сомкнутого состояния рибосомы, должны увеличивать ошибочное включение аминокислот в пептидную цепь. Среди них изучено влияние ионов магния, кальция, марганца и некоторых других. Противоположным действием обладают ионы натрия, калия и другие одновалентные катионы.Таблица 3. Аминокислотный код (указаны нуклеотиды кодонов и соответствующие им аминокислоты)
| Второй нуклеотид кодона | |||||||
|---|---|---|---|---|---|---|---|
| У | Ц | А | Г | ||||
| Первый нуклеотид кодона | У | (УУУ, УУЦ) Фенилалацин; (УУА, УУГ) Лейцин | (УЦУ, УЦЦ, УЦА, УЦГ) Серин | (УАУ, УАЦ) Тирозин; (УАА) —; (УАГ) — | (УГУ, УГЦ) Цистеин; (УГА) —; (УГГ) Триптофан | УЦАГ | Третий нуклеотид кодона |
| Ц | (ЦУУ, ЦУЦ, ЦУА, ЦУГ) Лейцин | (ЦЦУ, ЦЦЦ, ЦЦА, ЦЦГ) Пролин | (ЦАУ, ЦАЦ) Гистидин; (ЦАА, ЦАГ) Глютаминовая кислота | (ЦГУ, ЦГЦ, ЦГА, ЦГГ) Аргинин | УЦАГ | ||
| А | (АУУ, АУЦ, АУА) Изолейцин; (АУГ) Метионин | (АЦУ, АЦЦ, АЦА, АЦГ) Треонин | (ААУ, ААЦ) Аспарагиновая кислота; (ААА, ААГ) Лизин | (АГУ, АГЦ) Серин; (АГА, АГГ) Аргинин | УЦАГ | ||
| Г | (ГУУ, ГУЦ, ГУА, ГУГ) Валин | (ГЦУ, ГЦЦ, ГЦА, ГЦГ) Аланин | (ГАУ, ГАЦ) Аспарагин; (ГАА, ГАГ) Глютамин | (ГГУ, ГГЦ, ГГА, ГГГ) Глицин | УЦАГ | ||
Таблица 4. Принципиально возможные ошибки чтения кодонов, обнаруженные в экспериментах in vitro в бесклеточной среде на синтетических матрицах
| Категория ошибки | Исходный кодон | Ошибочное чтение |
|---|---|---|
| Ошибка в чтении одного из нуклеотидов кодона | УУУ (фенилаланин) | УУГ (лейцин) УЦУ (серин)ГУУ (валин) |
| Сдвиг рамки чтения | ЦАУ (гистидин) | АУЦ (изолейцин) УЦА (серин) |
| Ошибка в чтении двух нуклеотидов (очень редко встречавшаяся) | УУУ (фенилаланин) | УГЦ (цистеин) ААУ (аспарагиновая кислота)АУГ (метионин) |
 Рис. 6. Схема, поясняющая возможные механизмы увеличения ошибок рибосомального синтеза белка при ускоренном ритме клеточного деления и отсутствии нарушений во всех звеньях аппарата синтеза белка
Рис. 6. Схема, поясняющая возможные механизмы увеличения ошибок рибосомального синтеза белка при ускоренном ритме клеточного деления и отсутствии нарушений во всех звеньях аппарата синтеза белка
Главным условием правильного узнавания кодона является достаточное время экспонирования кодона, т. е. длительность периода разомкнутого состояния субчастиц рибосомы. Чем короче время экспонирования кодона, тем меньше времени на узнавание кодона, осуществляемое путем перебора антикодонов на аминоацилтранспортных РНК по законам стохастики, и тем больше вероятность ошибочного подбора антикодона и включения ошибочной, но близкой по кодовой специфичности аминокислоты (рис. 6). Изложенные особенности работы рибосомы были выявлены в модельных опытах на рибосомном ассоциате субчастиц в условиях так называемой бесфакторной трансляции. В условиях, более приближенных к естественной работе рибосомы, процесс сборки пептидной цепи на искусственной матрице ускоряется и количество ошибок возрастает. Общепринято суждение о том, что закономерности, полученные на модельных бесклеточных системах, справедливы и для процесса синтеза белка в живой клетке. Гемоглобин эмбрионального типа, или фетальный HbF, человека содержит по сравнению с основной формой гемоглобина HbA больше изолейцина (кодоны АУУ, АУЦ и АУА)[1] и меньше валина (ГУУ, ГУЦ, ГУА) и пролина (ЦЦУ, ЦЦЦ, ЦЦА), по данным В. Штейна и соавторов. Далее представим, что часть основного гемоглобина HbA превращается в гемоглобин HbF за счет замен указанных аминокислот в тех условиях, в которых содержание его снижается, а минорного, наоборот, повышается. Простой анализ, выполненный нами, показал, что тогда замену аминокислот в данном случае можно представить по принципу рибосомальных ошибок синтеза глобина, так как вместо валина может включаться изолейцин (у них второй и третий нуклеотиды соответственно одинаковые) и вместо пролина — также изолейцин (у них в трех кодонах одинаковые третьи нуклеотиды). После облучения кроликов в дозе 8,5 Гр, по данным Б. Ф. Сухомлинова, состав аминокислот гемоглобина изменился, уменьшилось количество цистеина (УГУ, УГЦ) и фенилаланина (УУУ, УУЦ) при увеличении аспарагиновой кислоты (ААУ, ААЦ) и гистидина (ЦАУ, ЦАЦ) — при отсутствии заметных изменений в содержании других аминокислот. В этом случае каждая пара кодонов у всех четырех аминокислот имеет одинаковые третьи нуклеотиды и, следовательно, у них имеется также определенная близость кодовой специфичности. Замена этих аминокислот могла произойти по принципу ложного кодирования. У собак на 15-е сутки после облучения в дозе 3,8 Гр минорный гемоглобин HbA1 увеличивался с 7,5 до 26%, а основной гемоглобин HbA упал с 86,5 до 65—68%. У минорного гемоглобина по сравнению с основным меньше содержание серина (кодоны УЦА, УЦГ, АГУ, АГЦ) и лейцина (УУА, УУГ) и увеличено содержание цистеина (УГУ, УГЦ), глутамина (ГАА), валина (ГУУ, ГУЦ), аргинина (ЦГУ, ЦГЦ) при отсутствии заметных изменений со стороны других аминокислот. Анализ кодонов этих аминокислот нам показал, что замена серина на цистеин и аргинин возможна, так как у них есть общие второй и третий нуклеотиды (ГЦ), серина на валин — по сдвигу рамки чтения (УЦГ на ГУЦ), лейцина на валин — тоже по сдвигу рамки чтения (УУГ на ГУУ), серина и лейцина на глутамин, валин и аргинин — по общности третьего нуклеотида (А). В указанном эксперименте на собаках прослежена динамика спектра гемоглобинов при одновременном исследовании красной крови. Наибольшее изменение в соотношении указанных двух фракций гемоглобина отмечалось в период 7—46 сут после облучения. Ранее 7 сут исследования, к сожалению, не проводились. Доля HbA с 86,5% упала до 65—68%, а доля HbA1 увеличилась с 7,5 до 24—27%. Доля других минорных фракций гемоглобина M1 и М2 в этот период была также увеличена. Снижение числа эритроцитов и гемоглобина в крови стало заметным с 12-х суток после облучения и было наибольшим на 20-е и 30-е сутки. У одной из старых собак, которая к моменту обследования имела выраженную анемию и увеличенную селезенку, через 8 лет после облучения почти весь гемоглобин был заменен на HbA1. Состояние костного мозга у них не исследовалось, но в наших экспериментах, описанных выше, были собаки, облученные примерно в такой же дозе, у которых анализировали и состояние костно-мозгового гемопоэза. У них отмечены изменения, позволяющие говорить об ускорении пролиферации и созревании эритробластов в период наблюдавшихся изменений в спектро гемоглобинов. Оно часто сопровождается укорочением клеточного цикла эритробластов в основном за счет сокращения стадии G1, во время которой идет массовый рибосомальный синтез белка. У эритробластов, потерявших способность к делению, может происходить ускорение созревания и укорочение времени на завершение процесса гемоглобинизации нормобластов. В специальных экспериментах на анемизированных крысах было показано сокращение времени пребывания на каждой стадии дифференцировки бластных клеток красного ряда: проэритробластов — в 1,27 раза, базофильных эритробластов — в 1,45 раза, полихроматофильных эритробластов — в 1,8 раза, ортохромных эритробластов — в 2,1 раза, ретикулоцитов — в 2,6 раза. Наибольшее сокращение времени созревания клеток было для тех стадий дифференцировки, на которых идет накопление гемоглобина и, следовательно, рибосомальный синтез молекул белка глобина, а ускорение рибосомального синтеза белка может быть связано с укорочением времени на один рибосомальный цикл с ошибочным включением в пептидную цепь глобина аминокислот с близкой кодовой специфичностью. Если изложенное верно, то с улучшением состояния красной крови и выполняемой ею главной дыхательной функции снимается напряжение эритропоэза, восстанавливается нормальная продолжительность клеточных циклов и периодов созревания, нормализуется продолжительность рибосомального цикла, падает ошибочное включение аминокислот в молекулу глобина, восстанавливается нормальный спектр гемоглобинов. Действительно, у указанных собак по мере улучшения красной крови восстанавливается нормальный спектр гемоглобинов. Наиболее интересные данные о спектре гемоглобинов человека связаны с появлением гемоглобина эмбрионального типа — фетального гемоглобина — при всех заболеваниях, при которых выявлено или предполагается напряжение эритропоэза. Достоверное повышение фетального гемоглобина выявлено у людей, подвергшихся хроническому воздействию свинца, ДДТ, при хронической желтухе, при различных заболеваниях крови — анемиях, лейкозах. Увеличение фетального гемоглобина — распространенная реакция на внешние химические и физические воздействия. Наконец, увеличение выделения в кровь эритропоэтина сопровождается изменением спектра гемоглобина. Эритропоэтин, как известно, ускоряет пролиферацию и дифференцировку клеток эритроидного ряда. Изучено соотношение активностей двух форм лактатдегидрогеназы — ЛДГ5 и ЛДГ1 — в зависимости от стимуляции пролиферативной активности и синтеза белка в селезенке облученных мышей. Действительно, при стимуляции указанных процессов закономерно преобладала активность ЛДГ5 над активностью ЛДГ1. Имеются более детальные данные о биохимических различиях непосредственно ЛДГ1 (чистая Н-форма) и ЛДГ5 (чистая М-форма) у свиньи. Поскольку с увеличением пролиферативной активности ткани увеличивается содержание ЛДГ5 и уменьшается содержание ЛДГ1, необходимо анализировать возможность замен тех аминокислот, которых больше в ЛДГ1 и меньше в ЛДГ5 и наоборот. В ЛДГ1 больше глутамина, треонина, серина, валина, аспарагина и фенилаланина. В ЛДГ5, наоборот, больше гистидина, лизина, аргинина, глицина и изолейцина. Предполагали, что увеличение содержания ЛДГ5 идет за счет ошибочных замен глутамина, треонина, серина, валила, аспарагина и фенилаланина, характерных для ЛДГ1, на гистидин, лизин, аргинин, глицин, изолейцин, характерные для ЛДГ5. Возможность таких замен по принципу ошибочного чтения одного из нуклеотидов кодона или по принципу сдвига рамки чтения представлена в табл. 5. Анализ показал принципиальную возможность всех указанных замен аминокислот, если усиление пролиферативной активности будет сокращать продолжительность рибосомального цикла.
Таблица 5. Возможные замены характерных аминокислот ЛДГ1 на характерные аминокислоты ЛДГ5 и их механизм (анализ данных работы Вахсмита и соавторов)
| Заменяемая аминокислота | Включаемая аминокислота | Механизм замены | |
|---|---|---|---|
| Замена одного нуклеотида | Сдвиг рамки чтения | ||
| Глутамин | Глицин | ГАГ → ГГГ | ГАГ → ГГА |
| Лизин | ГАА → ААА | ГАА → ААГ | |
| Аргинин | ГАА → АГА | ||
| ГАГ → АГГ | |||
| Треонин | Изолейцин | АЦУ → АУУ | |
| АЦЦ → АУЦ | |||
| АЦА → АУА | |||
| Аргинин | АЦА → АГА | ||
| АЦГ → АГГ | |||
| Лизин | АЦА → ААА | ||
| АЦГ → ААГ | |||
| Гистидин | АЦЦ → ЦАЦ | ||
| Серин | Аргинин | АГЦ → АГГ | АГЦ → ЦГА |
| АГУ → АГА | |||
| Глицин | АГУ → ГГУ | ||
| АГЦ → ГГЦ | |||
| Изолейцин | АГУ → АУУ | ||
| АГЦ → АУЦ | |||
| Гистидин | УЦА → ЦАУ | ||
| Валин | Изолейцин | ГУУ → АУУ | |
| "top" >ГУЦ → АУЦ | |||
| ГУА → АУА | |||
| Аргинин | ГУЦ → ЦГУ | ||
| Глицин | ГУУ → ГГУ | ||
| ГУЦ → ГГЦ | |||
| ГУГ → ГГГ | |||
| Аспарагин | Гистидин | ГАУ → ЦАУ | |
| ГАЦ → ЦАЦ | |||
| Глицин | ГАУ → ГГУ | ||
| ГАЦ → ГГЦ | |||
| Аргинин | ГАЦ → ЦГА | ||
| Фенилаланин | Изолейцин | УУУ → АУУ | |
| УУЦ → АУЦ | |||
Таблица 6. Различия в аминокислотной последовательности М- и Н-форм ЛДГ у цыплят (выписка из работы Торфа и соавторов)
| 2 | 5 | 6 | 9 | 10 | 12 | 13 | 14 | 15 | 17 | 18 | 19 | 20 | 24 | 30 | 41 | 43 | 45 | |
| M | УЦА | ГАУ | ЦАУ | ЦАЦ | ААУ | ЦАУ | AAA | ГАГ | ГАА | ЦАЦ | ЦАЦ | ГЦУ | ЦАУ | УЦА | ГЦГ | АУГ | ГАУ | ГЦУ |
| Ser | Asp | His | His | Asn | His | Lys | Glu | Glu | His | His | Ala | His | Ser | Ala | Met | Asp | Ala | |
| H | Thr | Glu | Lys | Thr | Pro | Ala | Ala | Gly | Ser | Thr | Val | Pro | Ser | Thr | Gly | Gly | Gly | Cys |
| АЦА | ГГУ | AAA | АЦЦ | ЦЦУ | ГЦУ | ГЦУ | ГГГ | УЦУ | АЦЦ | ГУЦ | ЦЦУ | УЦА | АЦА | ГГГ | ГГУ | ГГУ | УГЦ | |
| 49 | 54 | 73 | 75 | 78 | 79 | 80 | 84 | 88 | 91 | 93 | 98 | 116 | 123 | 124 | 130 | 132 | 133 | |
| M | АЦУ | ГУУ | AAA | ЦЦЦ | АЦУ | УЦА | ГГУ | УЦА | ЦАУ | ЦУУ | АУУ | ГЦУ | АУУ | ААЦ | ГУУ | ГАУ | AAA | ЦУУ |
| Thr | Val | Lys | Pro | Phr | Ser | Gly | Ser | His | Leu | He | Ala | He | Asn | Val | Asp | Lys | Leu | |
| H | Ala | Leu | Gin | His | Val | Ala | Asp | Ala | Asn | He | Val | Val | Val | Gin | He | Asn | Val | He |
| ГЦУ | ЦУУ | ЦАА | ЦАЦ | ГУУ | ГЦА | ГАУ | ГЦА | ААУ | АУУ | ГУУ | ГУУ | ГУУ | ЦАА | АУУ | ААУ | ГУА | АУУ | |
| 135 | 147 | 150 | 153 | 167 | 172 | 175 | 183 | 190 | 192 | 199 | 212 | 213 | 215 | 217 | 222 | 224 | 226 | |
| M | АУУ | ГЦУ | АУУ | УУУ | УЦУ | ЦАУ | ГГУ | ЦУА | ГУУ | ЦАГ | ЦЦУ | ААГ | ГЦА | ЦАУ | ГАУ | ГЦА | AAA | ЦАУ |
| Не | Ala | He | Phe | Ser | His | Gly | Leu | Val | Gln | Pro | Lys | Ala | His | Asp | Ala | Lys | His | |
| H | Val | Thr | Leu | Leu | Thr | Tyr | Ala | Thr | Leu | Glu | Ala | Gln | Gln | Asp | Ala | Lys | Ser | Asn |
| ГУУ | АЦУ | ЦУУ | УУА | АЦУ | УАУ | ГЦУ | АЦУ | ЦУУ | ГАГ | ГЦУ | ЦАГ | ЦАГ | ГАУ | ГЦУ | АЛА | АГУ | ААУ | |
| 236 | 243 | 249 | 258 | 260 | 263 | 264 | 268 | 272 | 273 | 276 | 280 | 231 | 284 | 285 | 296 | 297 | 300 | |
| M | ГАУ | AAA | АГУ | ГАУ | ГЦУ | АУА | АУГ | ЦГУ | ЦЦУ | АУУ | ГЦУ | АУГ | ЦАУ | AAA | ГАУ | ГГУ | УЦА | АУУ |
| Asp | Lys | Ser | Asp | Ala | He | Met | Arg | Pro | He | Ala | Met | His | Lys | Asp | Gly | Ser | He | |
| H | Glu | Arg | Asn | Glu | Cys | Met | Leu | Tyr | Ser | Val | Leu | Thr | Tyr | Gln | Asn | Ser | Ala | Leu |
| ГАА | АГА | ААУ | ГАА | УГЦ | АУГ | ЦУГ | УАУ | УЦУ | ГУУ | ЦУГ | АЦГ | УАУ | ЦАА | ААУ | АГУ | ГЦА | ЦУУ | |
| 302 | 304 | 305 | 306 | 307 | 310 | 314 | 315 | 316 | 325 | 329 | 331 | 333 | ||||||
| M | ГАУ | ГУУ | AAA | АУГ | АУА | ЦЦУ | ГАА | ЦАГ | АУУ | ГГУ | ГАА | ЦАА | УУУ | |||||
| Asp | Val | Lys | Met | He | Pro | Glu | Gln | He | Gly | Glu | Gln | Phe | ||||||
| H | Ser | He | Asn | Gln | Lys | Asp | Ala | Lys | Leu | Ser | Asp | Lys | Leu | |||||
| АГУ | АУУ | ААУ | ЦАГ | AAA | ГАУ | ГЦА | ААГ | ЦУУ | АГУ | ГАУ | AAA | УУА | ||||||
| Примечание. Цифры в таблице — номера аминокислот по порядку, буквы — сравниваемые нами триплеты кодонов. | ||||||||||||||||||
Таблица 7. Основные звенья цепи процессов производства функционирующего белка
| Процесс | Основное проявление | Зависимость от состояния пролиферативной активности ткани |
|---|---|---|
| Изменение структуры ДНК (мутация) | Стойкая замена аминокислот | Нет зависимости |
| Изменение функции ДНК (генная регуляция) | Временная замена аминокислот | Четкая зависимость предполагается |
| Транскрипция ДНК | Нарушение синтеза белка | Зависимость ожидается |
| Созревание иРНК | То же | То же |
| Транспорт иРНК из ядра в цитоплазму | '' | '' |
| Рибосомальный синтез белка | Закономерная замена аминокислот | Четкая зависимость доказана |
| Посттрансляционные изменения структуры белка | Модификация аминокислот | Зависимость можно предполагать |
Порочные потенциально патогенетические круги в физиологических и внутриклеточных системах
Биофизический анализ совокупности экспериментальных и клинических материалов о некоторых пострадиационных, возрастных и предлейкозных изменениях позволил обнаружить и описать определенную последовательность развития взаимосвязей в физиологических и внутриклеточных системах, формирующих порочные потенциально патогенетические круги. Кругами они названы потому, что обратные связи осуществляются через несколько промежуточных процессов или систем. Они являются патогенетическими, поскольку их функционирование может вызывать патологические явления, проявляемые определенной клиникой или клеточными нарушениями. Однако это происходит не всегда, а только в отягчающих условиях, и потому можно говорить лишь о потенциально патогенетическом характере этих кругов. Порочны они потому, что организму и его клеткам очень трудно самим без внешней помощи разорвать возникшие связи. Ниже выделены основные потенциально патогенетические круги и основные связи, их формирующие. Безусловно, можно найти и многие другие связи между системами и процессами, которые будут вовлекать и другие системы и процессы. Для нас важно было выделить минимальное их число, но достаточное, чтобы понять взаимодействие, формирующее неспецифические предпатологические состояния, понять биофизические механизмы, объясняющие определенную последовательность событий, выяснить пути возможного вмешательства с целью разорвать или ослабить неблагоприятное воздействие возникших порочных межсистемных взаимосвязей в клетках и организме.Порочные круги в физиологических системах как основа клинической предпатологии
Первый порочный круг. Ведущие процессы: уменьшение эффективности действия гормонов и недостаток гормонов. Имеются данные о возрастном повышении порога чувствительности гипоталамуса и желез внутренней секреции к соединениям (глюкоза, кортизон, эстрогены, гонадотропины), являющимся звеньями функциональной межсистемной регуляции по принципу прямых и обратных связей. Имеются и данные, свидетельствующие о возможном пострадиационном снижении чувствительности тканей к гормонам некоторых периферических эндокринных желез (тиреоидным, кортикостероидным). Показано также возрастное снижение чувствительности семенников к гонадотропной стимуляции и понижение в старости эффекта действия тестостерона на обменные процессы и морфологию тканей. Ускоренное снижение чувствительности тканей к медиаторам межсистемных связей может происходить и при воздействии различных внешних факторов и при различных заболеваниях. Снижение эффективности действия гормонов (в общем случае — медиаторов межсистемных связей) может происходить по трем причинам: 1) ухудшение качества гормонов; 2) изменения в акцепции гормонов; 3) снижение реактивности эффекторного органа в ответ на гормональный сигнал. В большинстве случаев нет оснований предполагать ухудшение качества гормонов в начальный период зарождения и развития порочных кругов. Усиление выработки собственной железой гормонов (того же качества) успешно компенсирует снижение их сродства с акцептором. Нарушения в акцепции гормонов могут зависеть от изменения качества и количества гормональных рецепторов. В обоих случаях необходимо анализировать состояния прежде всего плазматической мембраны клеток. Здесь только укажем, что нарушения в акцепции гормонов в начальный период зарождения порочных кругов наиболее вероятны и они связаны, по-видимому, с тонкими изменениями поверхности плазматической мембраны. Получены прямые экспериментальные доказательства возможности изменять порог чувствительности к гормонам (в общем виде — к медиаторам межсистемных связей) воспринимающих их тканей, воздействуя на состояние плазматической мембраны. Снижение реактивности эффекторного органа в ответ на гормональный стимул, т. е. снижение способности воспринимающих их тканей ответить адекватной реакцией, характерно для возрастных изменений. При снижении эффективности действия гормонов для выполнения их регулирующей роли организму требуется все большее количество гормонов. Создается состояние повышенных потребностей организма в гормонах. Соответственно стимулируется гормонообразование для осуществления нормальных регуляторных функций со стороны эндокринной системы. Происходит постоянно нарастающее повышение гормональной активности основных периферических эндокринных желез (щитовидной, коры надпочечников, половой). Так, женские яичники могут увеличивать в 2 раза выработку стероидных гормонов каждые 10 лет. Радиационное воздействие и другие факторы могут ускорять эти процессы. При достаточных физиологических резервах это не приводит к истощению эндокринных функций. Однако в отягчающих условиях длительное напряжение эндокринных функций может приводить к начальным явлениям истощения их и может отмечаться недостаточное количество гормонов в организме. Это вызывает новую стимуляцию гормонообразования. Так замкнулся первый порочный потенциально патогенетический круг (рис. 7). Рис. 7. Схема первого порочного потенциально патогенетического круга
Ведущие процессы: уменьшение эффективности действия гормонов и недостаток в организме гормонов
Рис. 7. Схема первого порочного потенциально патогенетического круга
Ведущие процессы: уменьшение эффективности действия гормонов и недостаток в организме гормонов
Длительное функционирование первого порочного круга может приводить к различным последствиям. 1. В сложившихся условиях крайне затруднена оценка функционального состояния эндокринной функции путем измерения концентрации гормонов в крови (тем более в моче). В крови, как в проточной системе, при ускоренном расходовании гормонов и напряжении в функционировании эндокринной железы реальная концентрация гормонов будет сильно флюктуировать и не отражать состояние железы. 2. Недостаток в организме гормона определяется разницей в скоростях его продукции и расходования и непосредственно не связан с состоянием гормонообразования. Недостаток гормона в организме может быть и при повышенном уровне гормонообразования, и при увеличении в размерах железы. Следовательно, нормальные показатели функционирования железы или признаки усиления ее деятельности могут скрывать реальный дефицит гормона в организме. 3. В рассматриваемых условиях могут быть расхождения в оценке состояния железы по гормональной продуктивности и по морфологическим признакам на основе биопсии. В условиях длительного напряжения функций железы общий уровень гормонообразования может складываться за счет работы разных участков железы, находящихся в различных функциональных состояниях (от гипофункциональных до состояний сильной гиперфункции и явного истощения). И наоборот, при сниженном уровне гормонообразования отдельные участки железы могут находиться в состоянии высокой функциональной активности, а отдельные узелки — в состоянии высокой пролиферативной активности. Указанное особенно ярко проявляется при развитии возрастной патологии. Свидетельством недостатка в организме гормонов, несмотря на усиленное гормонообразование, является отсутствие признаков гормонального токсикоза в этот период, в том числе и при дополнительном введении в организм гормонов. Наоборот, в этих случаях введение в организм гормонов оказывало благотворное действие по целому ряду показателей. Основной результат функционирования первого порочного круга, имеющий важнейшее значение для создания последующих потенциально патогенетических кругов, — это формирование состояния скрытой эндокринной недостаточности в организме.
Второй порочный круг. Ведущие процессы: усиленная гемоклазия и увеличение массы кроветворной ткани. При длительном функционировании первого порочного круга и при дальнейшем увеличении недостатка гормонов в организме может развиться общая неспецифическая реакция неблагополучия в ответ на скрытую эндокринную недостаточность. Она сопровождается активизацией защитно-охранительных систем организма, в том числе систем клеточного фагоцитоза. Нас интересуют в первую очередь процессы деструкции и фагоцитоза (пожирания) клеток крови, в первую очередь эритроцитов, при эндокринной недостаточности. В принципе полезная гемокластическая (кроверазрушительная) функция селезенки и других органов, участвующих в деструкции и фагоцитозе эритроцитов, направлена на очищение кровотока от устаревших и поврежденных клеток. Однако при длительной стимуляции ее происходит ускоренная убыль клеток из русла крови. Известно, что в физиологических условиях у человека лишь 10% старых эритроцитов разрушается в кровяном русле. Основная масса эритроцитов разрушается внутриклеточно после захвата их макрофагами печени, селезенки, костного мозга и легких. Кроме того, в эритрофагоцитозе могут участвовать ретикулярные клетки, гистиоциты и полинуклеарные лейкоциты. Селезенка не является основным местом деструкции эритроцитов в норме. В ней разрушается лишь 13—23% этих эритроцитов. Однако в определенных и патологических условиях селезенка становится настоящим кладбищем эритроцитов. Так, достаточно перевязать селезеночную вену или денервировать ее, как у экспериментальных животных развивается анемия. Мы приводили уже данные о синдроме гиперспленизма и о прямой зависимости продолжительности жизни эритроцитов от размеров селезенки при ряде заболеваний. Всякое замедление кровотока в селезенке и усиление секвестрации крови (т. е. задержки ее в полостях селезенки) ведут к увеличению вероятности контакта с макрофагами и другими эритрофагоцитирующими клетками, что в условиях селезенки нормальных размеров способствует первичной деструкции эритроцитов. Даже отсутствие первичной усиленной деструкции эритроцитов в селезенке в этих условиях еще не говорит о том, что она не оказывает на них повреждающего действия. В период замедленного прохождения эритроцитов по пульпе селезенки в эритроцитах могут возникать неблагоприятные изменения, которые лишь в дальнейшем приведут их к ускоренному разрушению. При этом прежде всего происходят изменения в гликопротеидах, белках и липидах поверхностных слоев мембраны, облегчающих контакт эритроцитов с макрофагами и другими эритрофагоцитирующими клетками. Еще раз напомним, что указанные изменения происходят в нормальном организме лишь с целью удаления старых и поврежденных клеток крови. Удаление селезенки не увеличивает продолжительности жизни всей популяции эритроцитов. Однако при стимуляции этих функций в условиях неспецифических реакций неблагополучия организма происходит ускорение разрушения эритроцитов.
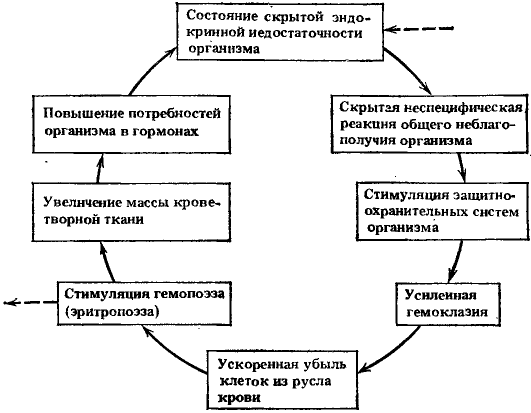 Рис. 8. Схема второго порочного потенциально патогенетического круга
Ведущие процессы: усиленная гемоклазия и увеличение массы кроветворной ткани
Рис. 8. Схема второго порочного потенциально патогенетического круга
Ведущие процессы: усиленная гемоклазия и увеличение массы кроветворной ткани
Вызывать такие реакции могут разные причины, в том числе и скрытая или явная недостаточность эндокринной системы. Имеются прямые доказательства этому. Ранее постулированное нами состояние скрытой (для клинициста) неспецифической реакции общего неблагополучия организма, ведущее через стимуляцию гемоклазии к ускоренной убыли клеток из русла крови, получило прямое экспериментальное доказательство. Ускоренная убыль клеток из русла крови вызывает соответствующую стимуляцию гемопоэза. В случаях несильной, но длительной по времени стимуляции гемопоэза, длящейся месяцами, в начале увеличивается продуктивность костного мозга за счет увеличения массы кроветворной ткани. «Желтый» жировой костный мозг трубчатых костей заменяется «красным» костным мозгом, активно продуцирующим эритроциты. В этот период показатели периферической крови и данные миелограммы остаются в пределах нормы. Такие состояния могут продолжаться длительное время и оставаться скрытыми для гематолога, использующего традиционные клинико-лабораторные методы исследования крови. Поскольку эти изменения вначале выражены не резко, они могут быть выявлены лишь специальными изотопными методами, позволяющими количественно определять суммарную продуктивность всего костного мозга организма за период времени, превышающий несколько суток. Увеличение массы гемопоэтической ткани повышает ее потребность в пластических веществах и в регуляторах обмена, в том числе и в гормонах. Это вновь способствует повышению потребностей организма в гормонах и обостряет уже имеющееся состояние скрытой эндокринной недостаточности. Так сформировался второй порочный круг, схема которого приведена на рис. 8. Одновременное функционирование обоих порочных кругов может приводить к более явным явлениям недостаточности эндокринной системы и напряжения гемопоэза.
Третий порочный круг. Ведущие процессы: недостаточность лейкопоэза и снижение иммунобиологической реактивности. Длительное функционирование первых двух порочных кругов в конце концов может приводить в глубоко зашедших случаях к зарождению нового, третьего порочного круга. В основе начала его лежит продолжающаяся стимуляция гемопоэза (эритропоэза). Организм уже не может поддерживать нормальный цитологический состав крови путем повышения продуктивности костного мозга за счет увеличения массы кроветворной ткани. Исчерпан также резерв репопуляции полипотентных стволовых клеток при нормальном распределении их дифференцировок в различные ростки кроветворения. В таких случаях в соотношении продукции клеток разных ростков происходит смещение в сторону клеток красного ряда и осуществляется примат продукции клеток этого ростка, так как он обеспечивает главную, т. е. дыхательную, функцию крови. Обоснования этого уже рассматривали. Уменьшение производства клеток белого ряда приводит к развитию недостаточности лейкопоэза (прежде всего лимфопоэза) со всеми вытекающими из этого последствиями, особенно со стороны иммунитета. Выраженность этих последствий может быть различной. На первом этапе нормальное количество эритроцитов и гемоглобина в крови обеспечивается за счет несущественного перераспределения пролиферирующих и дифференцирующихся клеток костного мозга в сторону производства эритроцитов с некоторым ущербом для белого ростка. В этот период в периферической крови число лейкоцитов неустойчиво находится в границах, близких к норме. При различных нагрузках и функциональных пробах на лейкопоэз происходят хорошо известные извращения лейкоцитарных реакций (вместо лейкоцитоза наблюдается ареактивность или кратковременное уменьшение их числа). На втором этапе при более значительном перераспределении пролиферирующих и дифференцирующихся клеток костного мозга в сторону производства эритроцитов продолжает поддерживаться норма функций красной крови при снижении числа лейкоцитов (гранулоцитов и лимфоцитов). Формы проявления недостаточности лейкопоэза могут быть разными. Это зависит от степени недостаточности поступления ранних пролиферирующих предшественников в различные ростки кроветворения. Если следовать по пути возможных направлений дифференцировок полипотентной стволовой клетки, то на первой развилке выбора направлений дифференцировок уменьшается поток клеток в лимфопозз и, следовательно, в первую очередь будет ослаблена система иммунной защиты организма. Имеются прямые клинические и экспериментальные доказательства этого положения. Наиболее чувствительным показателем недостаточности иммунной защиты организма является снижение устойчивости его к инфекционным факторам, бактериальным и другим токсинам, действию радиации; ослабляется и противораковая устойчивость организма. Недостаточность иммунной защиты организма может вызывать неспецифическую скрытую реакцию общего неблагополучия организма, вновь стимулирующую защитно-охранительную систему организма, включая гемоклазию и эритрофагоцитоз. Это, в свою очередь, дает дополнительный стимул к дальнейшей активизации гемопоэза. В условиях стимулированного эритропоэза такой стимул означает дальнейшую активизацию его и еще большее преобладание эритропоэза над другими ростками костного мозга. Замкнулся новый, уже третий порочный потенциально патогенетический круг, представленный в упрощенном виде на рис. 9. В этих условиях можно предполагать усиление потока клеток-предшественников в сторону миелопоэза, обеспечивающего эритроцитарный, грануло-макрофагальный и тромбоцитарный ростки гемопоэза. На уровне предшественника миелопоэза идет новый выбор направления дальнейшей дифференцировки. Необходимость примата производства клеток красного ростка приводит к развитию недостаточности грануло-макрофагального и тромбоцитарного ростков. Здесь могут складываться многообразные соотношения между этими ростками. По-видимому, меньше должен страдать или совсем не снижать своей активности макрофагальный росток, поскольку активация его эритрофагоцитарной активности происходила раньше в ответ на недостаток в организме гормонов и предшествовала активации эритропоэза.
 Рис. 9. Схема третьего порочного потенциально патогенетического круга
Ведущие процессы: недостаточность лейкопоэза и снижение иммунобиологической реактивности
Рис. 9. Схема третьего порочного потенциально патогенетического круга
Ведущие процессы: недостаточность лейкопоэза и снижение иммунобиологической реактивности
Ослабление иммунологической защиты организма должно также стимулировать усиление процессов гуморального и клеточного иммунитета и вызывать опять состояние скрытого неблагополучия организма. Однако в сложившихся условиях (активированный эритропоэз, активированная функция эритрофагоцитоза), по-видимому, всякое иммунологическое обострение уже не будет давать нормально выраженной реакции во всех звеньях гуморального и клеточного иммунитета. Будут прежде всего сказываться недостаточное поступление ранних предшественников в лимфопоэз и примат производства клеток эритроцитарного ряда. Поэтому в ответ на стимуляцию иммунной защиты организма система иммунитета, как мы полагаем, сможет ответить прежде всего теми реакциями, которые были до этого стимулированы, т. е. реакциями усиления гемоклазии, эритрофагоцитоза (см. рис. 9). Все события, охваченные первыми тремя порочными кругами, еще не дают ярких клинических изменений. При использовании традиционных клинико-лабораторных исследований, обычно проводимых при массовых профилактических осмотрах населения и отсутствии жалоб на здоровье, указанные изменения остаются вне внимания врача. Четкие отклонения от нормы не обнаруживаются: они являются клинически скрытыми и могут быть выявлены лишь при специальных нагрузочных тестах и при использовании особых методов исследования или при обращении к врачу по поводу какой-либо травмы или острой болезни, что вызвало более длительное обследование больного. Однако длительное функционирование описанных трех порочных кругов может приводить к более глубоким изменениям, обращающим на себя внимание врача и при традиционных методах клинико-лабораторного обследования. Возникает следующий, четвертый порочный потенциально патогенетический круг.
Четвертый порочный круг. Ведущие процессы: дальнейшая интенсификация эритропоэза и ухудшение качества эритроцитов. Длительное и одновременное функционирование трех порочных потенциально патогенетических кругов может в неблагоприятном исходе привести к тому, что показатели красной крови уже не смогут поддерживаться в пределах нормы за счет увеличении массы кроветворной ткани и примата производства клеток красного ряда. Появляются клинически заметные изменения как в системе крови, так и в эндокринной и других системах. Развитие указанных изменений можно разбить на несколько периодов или этапов. 1. В первый период еще не происходит ухудшение качества эритроцитов. Для сохранения дыхательной функции крови в норме начинают использоваться такие возможности, как ускоренный выброс из костного мозга недозревших эритроцитов (ретикулоцитов) в периферическую кровь и увеличение содержания гемоглобина в эритроцитах (увеличение цветного показателя). Общее число эритроцитов в пределах нормы, но число ретикулоцитов увеличено или значительно колеблется. Цветной показатель часто может быть выше нормы. Содержание лейкоцитов — на уровне нижней границы нормы или ниже ее (часто за счет лимфоцитов), может наблюдаться сдвиг лейкоцитарной формулы влево. Эритрофагоцитоз облигатными и факультативными системами клеточного фагоцитоза заметно усилен. В эритрофагоцитозе начинают участвовать ретикулярные и эндотелиальные клетки: их фагоцитоз не связан с состоянием иммунитета. Продолжительность жизни эритроцитов сокращена, иногда весьма значительно. Перекрестные переливания крови показывают, что это сокращение пребывания эритроцитов в русле крови происходит за счет внеклеточных для них факторов. В этот период устойчивость организма к инфекции, токсинам и радиации снижена. При возникновении травматических ран репаративная реакция вялая и скорость их заживления снижена. При исследовании периферических эндокринных желез выявляется повышенная функциональная активность щитовидной железы, коры надпочечников и эндокринной половой железы без явлений гормонального токсикоза. При морфологических исследованиях в щитовидной железе и коре надпочечников выявляются очаговые узлы функционального истощения наряду с очагами активной пролиферации. Изложенное состояние наиболее подробно описано применительно к периоду отдаленных последствий радиационного воздействия после состояния полного клинического выздоровления или к периоду формирования последствий слабых хронических воздействий радиации. Однако они не являются специфическими и возникают при различных иных воздействиях, а также могут быть элементами возрастной патологии. Часто наиболее чувствительным тестом развития указанных состояний выступает сниженная устойчивость к инфекции и бактериальным токсинам — проявления слабости иммунной защиты организма. Эксперименты показали, что восстановление такой устойчивости может затягиваться на чрезвычайно длительные сроки, составляющие значительную часть всей продолжительности жизни. Это происходит потому, что функции иммунитета являются второстепенными после дыхательной функции крови. Привлекает внимание в этих состояниях и замедление посттравматической репарации. Нами это обнаружено при травмах мягких тканей и костей. Это может быть также следствием перераспределения потоков клеток в различных ростках кроветворения, в том числе и в макрофагально-моноцитарном ростке. Изменения дифференцировок моноцитов в направлении тканевых макрофагов, участвующих в защитно-охранительных реакциях организма в ответ на недостаточность эндокринной системы, в ущерб гистиоцитам соединительной ткани могут объяснить феномен замедления посттравматической репарации. Важно подчеркнуть, что при возникновении описанных состояний длительное введение в организм в субфизиологических дозах гормональных препаратов щитовидной железы, коры надпочечников и реже половой железы в определенных условиях снимает или существенно ослабляет все перечисленные характерные явления. Наоборот, разнообразные дополнительные нагрузки (например, инфекция, токсины, антигормоны) утяжеляют эти явления и способствуют дальнейшему развитию порочных кругов. 2. Возможным вторым этапом функционирования четвертого порочного потенциально патогенетического круга является состояние с ухудшением качества эритроцитов. С целью сохранения жизненно важной дыхательной функции крови организм вынужден использовать новые резервы компенсации. Повышается пролиферативная активность клеток эритропоэза еще и за счет сокращения продолжительности клеточного цикла эритробластов и возможно более ранних предшественников. Возможность сокращения продолжительности цикла бластных клеток красного и особенно белого ряда костного мозга экспериментально доказана. Для ряда пролиферирующих клеток других тканей также имеются доказательства или это можно предполагать по аналогии. Имеются общие закономерности для всех клеточных популяций, реагирующих на внешние сигналы и повреждения изменением продолжительности клеточного цикла. Продолжительность собственно митоза, т. е. самого процесса деления клеток, практически не изменяется. Изменяется продолжительность интерфазы, т. е. периодов G1, S и G2. Наибольшее значение имеет продолжительность периода G1. На этой стадии идут массовый рибосомальный синтез белка, основной рост массы клетки, накопление структурно-мембранных компонентов клетки, все подготовительные мероприятия, необходимые для последующего удвоения молекулы ДНК. Наиболее значительные изменения в своей продолжительности имеет период G1. Продолжительность периодов G1, S и G2 при различных повреждениях клеток и в ответ на сигналы тревоги и необходимости подготовки к репарации предстоящих повреждений (при стрессовых ситуациях, при введении в организм радиопрофилактических средств и т. п.) увеличивается за счет блоковых задержек. При этом длительность блоковых задержек пропорциональна глубине повреждения клеток. Чем сильнее повреждение, тем больше времени необходимо на их репарацию, которая должна завершиться до перехода клетки в следующую стадию своего развития. Играет роль и изменение скорости репарации повреждений. Следует сказать, что имеются общие закономерности для гомойотермных организмов, определяющие эффективность восстановления, т. е. полноту устранения возникших спонтанных и индуцированных внешним воздействием повреждений. Под повреждениями здесь понимается вся совокупность возможных разнообразных повреждений, ошибок обмена и синтеза. Эти закономерности представлены на рис. 10. Эффективность работы восстановительных механизмов сложных биологических систем зависит в основном от двух факторов: скорости восстановительных процессов и продолжительности времени, в течение которого могут эффективно осуществляться восстановительные процессы. Детально изучались возможные границы изменения этих факторов у млекопитающих. Здесь только укажем, что в условиях гомойотермного организма млекопитающих из двух указанных факторов, определяющих эффективность и полноту восстановления, в организме более оперативно и в течение более длительных периодов используется фактор времени. Так, блоковые задержки на стадиях клеточного цикла могут наступать немедленно вслед за повреждающим воздействием. В то же время клетки могут в определенных условиях уходить в период покоя G0 практически на неограниченное время. Установлено, что в активно пролиферирующих реальных популяциях организма по мере усиления повреждения увеличивается продолжительность блоковой задержки. Однако она происходит лишь до определенного предела. Пока не достигнут предел продолжительности блоковой задержки, клетки, облученные в этой стадии, не имеют гибели ни интерфазного, ни митотического типа (в первом и последующих митозах). При отсутствии у клеток возможностей к дальнейшему увеличению продолжительности блоковой задержки в связи с усилением их повреждения (увеличение дозы облучения) они начинают гибнуть до митоза или во время последующих митозов.
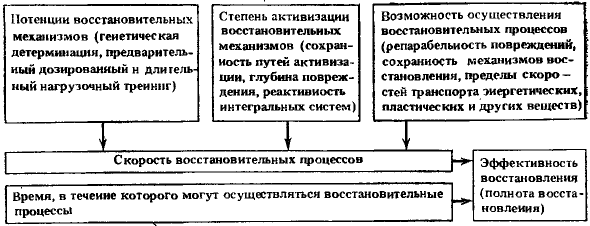 Рис. 10. Факторы, определяющие эффективность и полноту восстановления спонтанных и индуцированных внешним воздействием повреждений
Рис. 10. Факторы, определяющие эффективность и полноту восстановления спонтанных и индуцированных внешним воздействием повреждений
 Рис. 11. Схема четвертого порочного потенциально патогенетического круга
Ведущие процессы: дальнейшая интенсификация эритропоэза и ухудшение качества эритроцитов
Рис. 11. Схема четвертого порочного потенциально патогенетического круга
Ведущие процессы: дальнейшая интенсификация эритропоэза и ухудшение качества эритроцитов
Следовательно, недостаток времени для осуществления восстановления клетки на данной стадии клеточного цикла от индуцированных повреждений заставляет клетку переходить в следующую стадию с сохранением части повреждения. Остаточные повреждения часто не мешают проходить даже очередной митоз, но они могут кумулироваться до уровня, несовместимого с жизнью клетки, и она погибает через один или несколько митозов. При значительном усилении пролиферативной активности не всегда адекватный ответ ткани достигается только за счет вовлечения в делящийся пул клеток из покоящегося резерва. Происходит и сокращение продолжительности генеративного цикла клеток, в основном за счет периода G1. При этом повышается чувствительность клеток к повреждающим воздействиям, например к радиации. Увеличивается и вероятность неполного восстановления спонтанных и индуцированных повреждений из-за недостатка времени на их репарацию. Клетки могут переходить из стадии в стадию и вступать в митоз с остаточными повреждениями, способными к кумуляции. В этом мы видим одну из главных причин появления морфологических изменений (включений), совместимых с жизнью клетки и не имеющих определенного функционального значения. Вторая принципиально возможная причина появления таких изменений может заключаться в отставании поступления пластических и энергетических веществ от значительно ускоренных процессов роста и созревания клетки. Выше были специально рассмотрены биохимические и биофизические изменения, сопровождающие ускорение процессов пролиферации и созревания клеток. Закономерные изменения, происходящие при значительном усилении пролиферативной активности, могут быть связаны с процессами генетической регуляции и могут носить внегеномный характер. Наше внимание привлекли ошибки рибосомального синтеза белка, зависящие от изменения рибосомального окружения (ионный гомеостаз, pH и др.) и уменьшения времени на перебор кодон—антикодонных соотношений, происходящих, как предполагается, при сокращении продолжительности периода G1. В этих случаях может включаться ошибочная аминокислота, близкая по своей кодовой специфичности «правильной» аминокислоте, информацию о которой несет матричная РНК. В таких случаях, несмотря на то что ни в ДНК, ни в матричной РНК, ни в рибосоме никаких изменений не произошло, происходит синтез белка с ошибками. Сократилось лишь время на один цикл работы рибосомы. Одна или несколько таких ошибок не могут существенно повлиять на активный центр фермента, но могут изменять антигенные свойства белка и модифицировать ряд физических характеристик белка и белоксодержащих структур (плазматических мембран и др.). Анализ всего пути следования информации от ДНК до функционирующего белка показал, что имеются и другие, но менее вероятные возможности внесения ошибок, влекущих за собой включение ошибочных аминокислот в пептидную цепь белка. Они во многом зависят от изменения окружения информационных молекул, от ионного гомеостаза, от pH и других локальных изменений, которые происходят при значительном усилении пролиферативной активности клеточной популяции и значительном ускорении клеточного деления. При накоплении проходящих через митоз изменений могут уже ухудшаться качества белковой части гемоглобина, теряться сродство гемоглобина к кислороду, в связи с чем будет снижаться дыхательная функция крови. Это, в свою очередь, будет вновь усиливать стимуляцию эритропоэза. Таким образом, сформировался новый порочный патогенетический круг, схема которого в упрощенном виде представлена на рис. 11.
Пятый порочный круг. Ведущие процессы: истощение эритропоэза и стимуляция лейкопоэза. Длительное функционирование четырех описанных выше порочных потенциально патогенетических кругов создает все возрастающую стимуляцию функций красной крови по изложенным уже механизмам межсистемных связей. В конце концов это может приводить к начинающемуся истощению эритропоэтической функции и явлениям стойкой анемии различной выраженности. Эта анемия некурабельна при применении гемостимуляторов. Уменьшение числа эритроцитов и гемоглобина в периферической крови, снижение дыхательной функции крови приводят к мобилизации всех возможных резервов кроветворения для улучшения дыхательной функции. Продукция других ростков кроветворения снижена, начинает проявляться недостаточность лимфопоэза, гранулопоэза и тромбопоэза. В далеко зашедших случаях в крови может отмечаться снижение числа гранулоцитов, лимфоцитов, включая иммунокомпетентные клетки, и реже тромбоцитов. Картина изменений цитологического состава может быть весьма разнообразной. Чаще преобладает снижение зрелых клеточных форм одного из указанных ростков, но могут наблюдаться и сложные отношения, включая панцитопению. Проявления подобных состояний достаточно хорошо известны клиницистам. Указанные процессы развиваются на фоне скрытой или уже явной эндокринной недостаточности, скрытой или уже ставшей явной недостаточности иммунологической защиты организма. Начинает проявляться накопление модифицированного белка в разных белоксодержащих структурах, и в первую очередь в мембранах, в том числе в наружной клеточной мембране, что может отражаться на акцепторных свойствах ее по отношению к медиаторам межклеточных и межсистемных связей. Эндокринная недостаточность начинает усугубляться и ухудшением качества гормонов в связи с невозможностью в течение очень длительных сроков отвечать на постоянный стимуляционный сигнал увеличенной продукцией полнокачественных гормонов. Железа начинает выделять недозревшие и некачественные гормоны или их малоактивные аналоги. Эти процессы наиболее подробно изучены применительно к продукции гормонов женским яичником. Яичник начинает вместе с эстрадиолом вырабатывать и неклассические фенолстероиды, уже не обладающие всеми свойствами женского полового гормона. По каналам обратной связи это приводит к активизации выделения гипофизом гонадотропного гормона, что вновь стимулирует яичник к усиленной выработке гормона. Возникают стойкие очаги активной клеточной пролиферации в паренхиме железы. Сложные отношения возникают и в системе иммунитета. Известно определенное противостояние клеточного и гуморального иммунитета, особенно в условиях недостаточного поступления клеток — ранних предшественников в лимфопоэз. При усилении клеточного иммунитета может ослабляться гуморальный иммунитет и наоборот. При значительном примате клеточной продукции миелопоэза могут быть ослаблены все иммунные функции и клеточный и гуморальный иммунитет. В условиях функционирования описанных порочных кругов, по-видимому, клеточный иммунитет может быть ослаблен меньше, чем гуморальный, поскольку первый в определенной мере участвует в усиленной эритроклазии и эритрофагоцитозе. Недостаток иммунных функций создает также состояние, вероятно, общего неблагополучия организма и вызывает по каналам обратной связи стимул к усилению их.
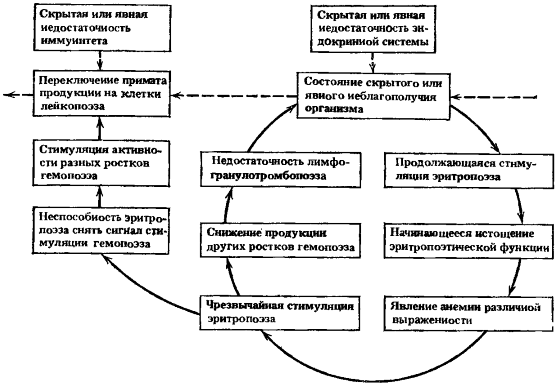 Рис. 12. Схема пятого порочного потенциально патогенетического круга
Ведущие процессы: истощение эритропоэза и стимуляция лейкопоэза
Рис. 12. Схема пятого порочного потенциально патогенетического круга
Ведущие процессы: истощение эритропоэза и стимуляция лейкопоэза
Усугубление всех указанных процессов при возникшей неспособности эритропоэза снять или существенно ослабить стимуляционный сигнал к активации гемопоэза выправлением дыхательной емкости крови приводит к стимуляции разных ростков кроветворения. В одних случаях появляется полицитемия, в других — начинает преобладать один из неэритроцитарных ростков. Чаще происходит переключение примата производства клеток в направлении лейкопоэза (лимфопоэза или гранулопоэза), но возможно и в направлении тромбопоэза и моноцитопоэза. Все это происходит в условиях реакции скрытого или явного неблагополучия организма при определенных нарушениях межсистемных взаимодействий. Схема основных событий пятого порочного круга представлена на рис. 12.
Шестой порочный круг. Ведущие процессы: примат лейкопоэза и нарушения качества и количества лейкоцитов. Шестой порочный потенциально патогенетический круг рассмотрен на примере клеток белого ряда. Значительная стимуляция пролиферативной активности клеток белого ряда, продолжающаяся длительные сроки, может приводить к сокращению продолжительности клеточного цикла делящихся и созревающих клеток, участвующих в лейкопоэзе (полипотентная стволовая клетка, коммитированная в сторону лимфопоэза или миелопоэза с дальнейшим направлением дифференцировки в сторону гранулоцитарного или моноцитарного ростков, миелобласты и миелоциты или монобласты и моноциты). Известно, что в случае такой стимуляции клетки белого ряда реагируют более значительным укорочением продолжительности клеточного цикла, чем клетки красного ряда. Возможность снижения эффективности и полноты репарации спонтанных и индуцированных повреждений, ошибок обмена и синтеза мы также уже обсуждали. По-видимому, клетки белого ряда, как выполняющие менее значимую функцию, чем клетки красного ряда, не имеют столь мощных систем репарации. Известно, что эритропоэз может увеличивать свою продуктивность в 6—8 раз после массивных кровопусканий, сокращать при этом продолжительность клеточного цикла для клеток делящегося и делящегося—созревающего пула. Тем не менее очень часто не наблюдаются какие-либо морфологические изменения клеток или включения в них. Для клеток белого ряда характерно появление разнообразных морфологических отклонений или включений при стимулированном лейкопоэзе. Поэтому имеются основания считать, что в клетках белого ряда в отличие от клеток красного ряда быстрее будут накапливаться недорепарированные повреждения, ошибки обмена и синтеза, чем в клетках красного ряда. Имеются в виду повреждения, переходящие из одной стадии в следующую и через митоз. Быстрее будут ухудшаться качество лейкоцитов, их функциональные свойства. Изложенное приводит к снижению иммунологической защиты организма и к новой стимуляции лейкопоэза. Указанные события замкнули новый, уже шестой порочный патогенетический круг (рис. 13), функционирование которого может наращивать силу стимуляционного сигнала к дальнейшей активации лейкопоэза. Продолжающаяся стимуляция лейкопоэза, ускоренное деление клеток, накопление в них повреждений, сохраняющиеся во многих поколениях клеток биофизические и биохимические изменения, характерные для усиленно пролиферирующих клеток, в конце концов могут приводить к автономности ритма ускоренного клеточного деления, к редуцированному клеточному метаболизму и дедифференцированному состоянию. Возникают порочные потенциально патогенетические круги во внутриклеточных процессах, которые будут рассмотрены ниже. Все изложенное формирует состояния, способствующие малигнизации клеток.
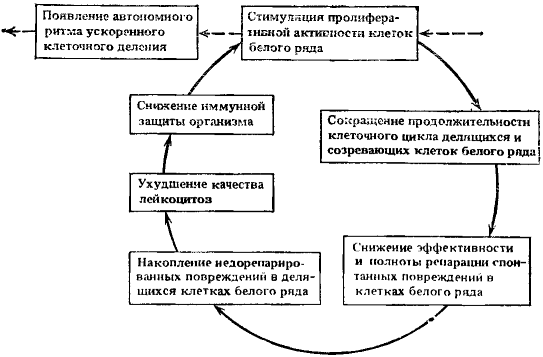 Рис. 13. Схема шестого порочного потенциально патогенетического круга
Ведущие процессы: примат лейкопоэза и нарушения качества и числа лейкоцитов
Рис. 13. Схема шестого порочного потенциально патогенетического круга
Ведущие процессы: примат лейкопоэза и нарушения качества и числа лейкоцитов
Рассмотренные порочные потенциально патогенетические круги в физиологических системах не исчерпывают всех возможных взаимосвязей, последовательно вовлекающих новые процессы и новые физиологические структуры, способствующие переходу предпатологии в клинически выраженные формы. Внутри каждого порочного круга также не исчерпаны все активно действующие взаимосвязи. Тем не менее сделанные упрощения позволили описать шесть последовательно возникающих порочных кругов в физиологических системах. Для нас было важно получить такую последовательность событий в результате функционирования порочных кругов, которая соответствовала бы: 1) последовательности клинических проявлений в период между воздействием радиации или другого лейкозогенного агента и заболеванием лейкозом; 2) возможной последовательности развития клинических синдромов общей гемобластозной болезни; 3) последовательности изменений в различных системах (кроветворения, эндокринная, иммунная и др.), предшествующих появлению отдаленной радиационной патологии, и могла их объяснить. Кроме того, порочные круги должны были иметь определенную связь с возрастными изменениями, поскольку частота возникновения лейкозов, так же как и других злокачественных заболеваний, зависит от возраста организма. Следовательно, процессы и события, ведущие к предлейкозу, должны быть неспецифическими и общими с целым рядом других предпатологических состояний. Описанные взаимосвязи, создающие порочные потенциально патогенетические круги, отвечают указанным требованиям и хорошо объясняют всю последовательность событии. Возможность диагностического использования этой последовательности событий будет рассмотрена в последних разделах. Изложение порочных кругов мы начали с изменений в эндокринной системе, в которой раньше, чем в других системах нормального человека, выявляются значимые для организма возрастные изменения. Это не значит, что развитие порочных кругов всегда должно начинаться с изменений в этой системе. Источником зарождения порочных кругов могут быть и другие самые разнообразные причины. Они могут начинать порочные круги в различных пунктах. Например, хронические и токсические воздействия — с реакций общего неблагополучия организма и стимуляции защитно-охранительных систем организма; хронические нейрогенные и некоторые лекарственные — с явлений стойкой гиперсеквестрации крови в селезенке и ускоренного разрушения клеток крови; различные иммуннопатологические факторы — также с ускоренной гемоклазии; онкогенные вирусы, по-видимому, будут ускорять накопление повреждений в бластных клетках белой крови и их предшественниках. Вероятность прогрессивного развития каждого патогенетического круга зависит от наступающего истощения репаративных и компенсаторно-восстановительных возможностей системы. Последние генетически предопределены. Различная предопределенность их обусловливает индивидуальные отличия в формировании порочных кругов и в вероятности завершения их переходом от состояния предпатологии к клинически выраженной патологии. Многое зависит от процессов, происходящих на субклеточном уровне, которые также могут формировать свои порочные патогенетические круги.
Порочные круги во внутриклеточных системах как основа клеточной патологии
Вводные замечания можно распространить и на порочные потенциально патогенетические круги во внутриклеточных системах. Следует сказать, что в клетке труднее, чем в физиологических системах, выделить те существующие связи между различными системами, которые могли бы в соответствующих условиях формировать порочные круги. Здесь труднее отделить одни процессы от других ввиду тесных взаимосвязей различных внутриклеточных систем, опосредованных прежде всего непосредственно биохимическими регуляциями. Тем не менее ниже сделана попытка выделить основные взаимосвязи, возникновение которых, по нашим представлениям, ведет к формированию порочных потенциально патогенетических кругов под влиянием непрерывной стимуляции клеток к ускоренной пролиферации, к ускоренному размножению. В отличие от порочных кругов в физиологических системах здесь ведущие связи оказались более сложными. Их трудно описать простым кругом. Приходится прибегать к разветвленным и параллельно существующим связям. Тем не менее и их оказалось возможным замкнуть в конечном итоге в порочный круг, препятствующий возвращению клеток к состоянию, в котором они находились до воздействующего сигнала к ускоренной пролиферации.Первый порочный круг. Ведущие процессы: ускоренный ритм деления клеток и редуцированный метаболизм. Как следует из ранее изложенных материалов, конечным результатом функционирования порочных потенциально патогенетических кругов в физиологических системах может быть длительное время продолжающаяся стимуляция пролиферативной активности отдельных тканей или части клеток этой ткани. Такая стимуляция вызывает не только вовлечение в делящийся пул новых, ранее покоящихся клеток, но и ускорение самого ритма прохождения клетками митотического цикла. Сокращение митотического цикла, происходящее за счет укорочения периодов интерфазы и продолжающееся во многих поколениях клеток, влечет за собой целый комплекс изменений, характеризующих недодифференцированное — дисдифференцированное состояние с определенной редукцией метаболизма. Для нормального завершения структурообразующих дифференцировочных реакций необходимо определенное время. При укорочении интерфазы митотического цикла сокращается и время для указанных реакций. Уже одно это может в определенной мере препятствовать осуществлению дифференцировки клеток. В настоящее время считается доказанной даже для морфологически неразличаемых костно-мозговых стволовых клеток зависимость степени дифференцировки (уход в коммитированное состояние или в пролиферацию) от продолжительности периода G1 интерфазы. Укорочение или удлинение периода G1 сопровождалось соответственно пролиферацией или дифференцировкой стволовых клеток. Сокращение периодов интерфазы митотического цикла сокращает и время для осуществления процессов репарации и восстановления спонтанных и индуцированных внешними для клетки влияниями повреждений, ошибок обмена и синтеза. Известно, что биологические системы в ответ на возросший объем повреждений (если он не превышает критического уровня) обычно отвечают повышением эффективности восстановительных процессов за счет ускорения их и увеличения времени на их осуществление. При этом большее значение имеет фактор времени, чем ускорение восстановительных процессов. На уровне клеточных популяций эта закономерность проявляется в увеличении продолжительности периодов интерфазы митотического цикла за счет блоковых задержек и в определенном соотношении их степени повреждения клеток. В случае сохранения сокращенных периодов интерфазы митотического цикла в последующих поколениях клеток недорепарируемые и недовосстановленные повреждения, ошибки обмена и синтеза, не приводящие к гибели клеток в первом митозе, будут накапливаться с каждым поколением клеток и в конце концов начнут мешать течению нормальных метаболических процессов и в крайнем случае вызывать гибель клеток. В этом мы видим причину гибели части поврежденных клеток не в первом митозе, а в последующих, а также причину срыва так называемого абортивного подъема числа лейкоцитов после их снижения при воздействии ионизирующих излучений. С этим, видимо, связаны и другие наблюдения. Так, при длительной стимуляции пролиферативной активности ткани в клетках могут появляться различные морфологические аномалии и включения. Например, в период, предшествующий появлению клиники типичного лейкоза, наиболее характерны гиперпластические состояния костного мозга, в основном по красному ростку. Такие состояния сопровождаются ускоренным эритропоэзом с сокращением митотического цикла клеток делящегося пула. Именно для предлейкозного синдрома характерно появление телец Бизара. Они могут встречаться у 79% лиц, находящихся в предлейкозном состоянии. Другой пример связан с упоминавшимся абортивным подъемом числа лейкоцитов в крови в пострадиационном периоде. В развитии лейкопении имеются два максимума снижения числа лейкоцитов в периферической крови: у человека на 8—12-е и 27—32-е сутки и у собаки на 4—8-е и 19—20-е сутки после воздействия радиации. Между этими периодами снижения числа лейкоцитов отмечается временный подъем их числа с последующим абортивным снижением уровня лейкоцитов. В этот период бурного, но неудавшегося восстановления лейкопоэза выявляют значительное повышение митотического индекса и сокращение цикла, а также появление клеток со структурными аномалиями, пикнотическими ядрами и дегенеративными изменениями цитоплазмы. В том числе отмечают изменения, связанные с характером конденсации ядерного хроматина, и изменения типа и степени сегментации ядра гранулоцитов. Подробно проблема биохимических, биофизических и цитологических изменений, связанных с усилением пролиферативной активности ткани, рассмотрена выше. Здесь мы хотели только напомнить, что имеются общие биохимические, биофизические и цитологические особенности для совершенно разных клеточных популяций: 1) в эмбриональной ткани по сравнению с происходящей от нее более дифференцированной тканью; 2) в ткани в период репаративной регенерации по сравнению с этой же тканью в период нормальной физиологической регенерации; 3) в активно растущей злокачественной ткани по сравнению с предшествующей ей нормальной тканью. Общими особенностями для них являются состояние активной пролиферации, сопровождаемое преобладанием анаэробного гликолиза над дыханием, накопление лактата, падение окислительно-восстановительного потенциала (при достатке углеводного питания), закономерные изменения изоэнзимного и антигенного спектра белков, цитологическое упрощение морфологии клеток. На первых порах после создания прекрасной модели — гепатом Морриса с различной степенью злокачественности (начиная с состояния, близкого к норме) стали искать закономерности изложенных изменений в зависимости от степени дифференцировки ткани. Была обнаружена корреляция между степенью дифференцировки и скоростью роста ткани, выраженностью указанных биохимических особенностей и изменением изоэнзимного состава ферментов. Такая расстановка последовательности событий не давала возможности подойти к изучению причинно-следственных отношений. Поэтому позднеестали искать зависимость указанных изменений от скорости роста, т. е. от степени пролиферативной активности ткани. Следовательно, изложенные изменения стали связывать и с сокращением интерфазы митотического цикла, в частности периода G1 со всеми вытекающими последствиями, рассмотренными выше. Возрастание ошибок синтеза описано и на примере ошибочного включения аминокислот в пептидную цепь белка при усилении пролиферативной активности ткани, сокращений стадии G1 митотического цикла и ускорении рибосомального цикла. На возможность такой взаимосвязи процессов мы указывали еще в 1974 г. Вопрос большой принципиальной важности: могут ли сохраняться те многообразные изменения в клетках, которые возникают при длительной стимуляции пролиферативной активности, в случае снятия такой стимуляции? Морфологическое упрощение и редукция метаболических путей, навязанные клеткам длительно сохраняющейся стимуляцией к ускоренному делению клеток, становятся ли настолько привычными, что клеткам уже невыгодно в энергетическом и в других отношениях возвращаться к исходному состоянию? Материалы, обобщенные выше, говорят в пользу возможности сохранения указанных изменении и при последующем снятии стимуляционного сигнала. Мы обращаем внимание при этом на следующие обстоятельства. Во-первых, при длительно сохраняющейся стимуляции пролиферативной активности происходит одновременно в силу указанных уже выше причин сокращение периодов интерфазы митотического цикла и накопление проходящих через митоз спонтанных и индуцированных внешними для клеток факторами повреждений, ошибок обмена и синтеза. В этих условиях, затрудняющих осуществление полноценной дифференцировки клеток, ее потенциальные возможности к нормальной дифференцировке не реализуются во многих поколениях клеток. Соответствующие локусы генома не дерепрессируются и не реализуют свою информацию. Многократное повторение такой ситуации во многих поколениях клеток при возрастающей нестабильности генома в конце концов, как считают, может закрепить постоянную репрессию их на уровне регуляторных генов. Это будет препятствовать возвращению клеток в нормальное дифференцированное состояние и после снятия стимуляционного сигнала. Клетки будут продолжать находиться в состоянии сокращенного митотического цикла и ускоренного деления. Во-вторых, при длительно сохраняющемся ускоренном делении клеток могут быть затруднения в энергетическом обмене. Превалирование анаэробного обмена над дыханием существенно уменьшает синтез макроэнергетических соединений. Недостаток времени и энергетических и пластических веществ (имеются серьезные ограничения в увеличении скорости их транспорта) упрощает и редуцирует клеточный метаболизм. Начинают играть все большую роль короткие, упрощенные и менее энергоемкие метаболические пути. Все большее значение приобретает «крупноблочный» синтез ряда макромолекул. Известно, что синтез органических соединений из неорганического сырья обходится организму энергетически дорого. Так, при синтезе одной молекулы глутаматной кислоты (с преобразования которой идет последующий синтез ряда других аминокислот) расходуется 28,5 молекулы АТФ. Для синтеза одной молекулы пиримидинового нуклеотида уридинмонофосфата из аспарагиновой кислоты затрачивается 53,5 молекулы АТФ. В норме у высших млекопитающих нуклеиновые кислоты распадаются до мононуклеотидов, пуриновых и пиримидиновых оснований и далее часть их — до мочевины и аммиака. В условиях ускоренного деления клеток, смещения pH в «кислую» сторону и недостатка макроэргов распад нуклеиновых кислот происходит лишь до моно- или даже олигонуклеотидов, а синтез их — из блоков готовых нуклеотидов без предварительного синтеза их оснований. Синтез белка в основном идет из готовых аминокислот. Такие изменения метаболизма становятся энергетически более выгодными, и клетки по этой причине также сохраняют эти изменения и после снятия стимула к ускоренному делению клеток. Сохраняется и недодифференцированное состояние. Основные связи между указанными процессами представлены на рис. 14 в максимально упрощенном виде. Они сформировали первый внутриклеточный порочный круг, препятствующий возвращению клеток к состоянию нормальной дифференцировки и спокойной пролиферации.
 Рис. 14. Первый круг
Ведущие процессы: ускоренный ритм деления клеток и пролиферативно-редуцированный метаболизм
Рис. 14. Первый круг
Ведущие процессы: ускоренный ритм деления клеток и пролиферативно-редуцированный метаболизм
Второй порочный круг. Ведущие процессы: избыточный синтез низкомолекулярных тиолов и уменьшение специфичности структуры поверхности клетки. Навязанный клеткам ускоренный ритм деления, помимо рассмотренных событий, упомянутых при изложении первого порочного круга, вызывет одновременно и много других биофизических и биохимических изменений. Рассмотрим связи, в которых ведущее значение имеют синтез низкомолекулярных тиолов и изменение специфичности клеточной поверхности и которые формируют второй порочный круг (рис. 15). На возможности изменения соотношений анаэробного и аэробного метаболизма, прежде всего глюкозы, и на связанном с этим снижении окислительно-восстановительного потенциала мы уже останавливались. Наблюдаются также изменения обмена низкомолекулярных тиолов. По мнению Е. Е. Селькова, перестройка всего клеточного метаболизма может происходить со сменой стационарных состояний обмена низкомолекулярных тиолов, сопровождаемой большим количеством изменений в различных звеньях метаболизма. Активность анаэробного гликолиза и пентозофосфатного шунта увеличивается.
 Рис. 15. Второй круг
Ведущие процессы: повышенный синтез низкомолекулярных тиолов и уменьшение специфичности структуры поверхности клеток
Рис. 15. Второй круг
Ведущие процессы: повышенный синтез низкомолекулярных тиолов и уменьшение специфичности структуры поверхности клеток
Увеличение транспорта кислорода и активности окислительного фосфорилирования в то же время имеет свои ограничения. Скорость глюконеогенеза и транспорта глюкозы в клетку отстает от скорости ее потребления. Концентрация гексозомонофосфатов уменьшается. Уменьшается активность ряда ферментов синтеза гликолипидов и гликопротеидов. Активность липолиза и синтеза фосфолипидов и белков в то же время увеличивается. Однако уменьшен синтез разнообразных производных N-ацотилглюкозамина, сиаловой кислоты и глюкоконъюгатов клеточной поверхности. Это существенно отражается на молекулярной структуре внешней поверхности плазматической мембраны клеток. Уменьшаются ее специфичность и адгезивность (способность к прилипанию). Указанные глубокие изменения метаболизма, по мнению Е. Е. Селькова, происходят в связи с переключением SH-систем клетки из одного стационарного состояния, характеризующегося низким уровнем тиолов, в альтернативно другое стационарное состояние с повышенным уровнем тиолов. Выполненный им анализ кинетики поведения биохимических систем показал, что следствием нового стационарного состояния метаболизма низкомолекулярных тиолов должен быть ускоренный ритм клеточного деления. Упомянутые изменения клеточной поверхности приводят к ослаблению контактного торможения деления клеток и к изменению иммунной специфичности гликопротеидов плазматической мембраны и ее акцепторных свойств по отношению к медиаторам межклеточных связей. Изложенные взаимосвязи изменений в различных процессах формируют второй внутриклеточный порочный круг, сохраняющий, так же как и первый круг, ускоренное клеточное деление и в случае снятия внешнего для клетки сигнала стимуляции пролиферативной активности.
Третий порочный круг. Ведущие процессы: усиление перекисного окисления липидов мембран и частичная декомпартментализация низкомолекулярных веществ. Длительное повторяющееся во многих поколениях клеток ускоренное их деление существенно отражается и на состоянии липидного обмена. На рис. 16 выделены некоторые из них, приводящие к формированию третьего порочного круга. Основное внимание уделено тем процессам в липидном обмене, которые изменяют неспецифическую мембранную проницаемость и которые выше уже рассматривались. Здесь только мы пытаемся составить логическую последовательность их изменений. Длительная стимуляция пролиферативной активности и ускоренное деление клеток приводят к уменьшению образования новых липидных соединений и, наоборот, увеличивают использование ранее синтезированных липидов. Липонеогенез в значительной мере заменяется липолизом. Количество ненасыщенных жирных кислот увеличивается, а насыщенных — уменьшается. Происходят существенные изменения и в системе оксидантов, т. е. природных соединений, участвующих в окислительных процессах. Содержание прооксидантов увеличивается при уменьшении антиоксидантов. Активируются процессы окисления.
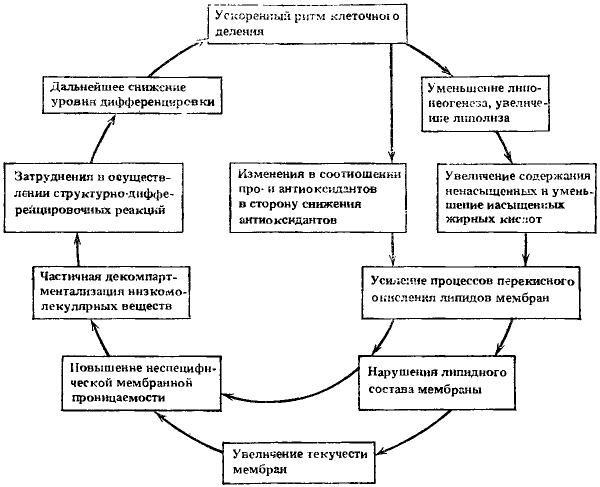 Рис. 16. Третий круг
Ведущие процессы: усиление перекисного окисления липидов мембран и частичная декомпартментализация низкомолекулярных веществ
Рис. 16. Третий круг
Ведущие процессы: усиление перекисного окисления липидов мембран и частичная декомпартментализация низкомолекулярных веществ
Все изложенное приводит к усилению процессов перекисного окисления липидов, изменению состава липидов мембран и повышению «текучести» мембран; их вязкость снижается. Проницаемость мембран для многих веществ повышается. Происходит нарушение нормального распределения и разграничения различных веществ в цитоплазме, т. е. происходит определенная декомпартментализация их, особенно низкомолекулярных веществ. Указанная частичная декомпартментализация усложняет осуществление структурно-дифференцировочных реакций в клетке, создает новые затруднения в дифференцировке клеток. Это наряду с другими причинами, изложенными выше, приводит к дальнейшему снижению уровня дифференцировки клеток, что по причинам, ранее изложенным, смещает пролиферативно-дифференцировочное отношение в сторону активации пролиферации, т. е. ускоренного размножения клеток. Снятие первоначального внешнего для клетки сигнала к пролиферации в этих условиях не снимает ускоренного деления клеток. Появились новые внутриклеточные условия, которым более соответствует ускоренный ритм деления. Порочный круг вновь замкнулся. В этом разделе рассмотрены процессы, наиболее характерные для цитоплазмы клеток. Рассмотрение близких процессов для ядра клеток представлено следующим порочным кругом.
Четвертый порочный круг. Ведущие процессы: нарушение проницаемости ядерной мембраны и фиксирование на уровне генетической регуляции изменений метаболизма и дедифференцированного состояния. Описанные выше навязанные ускоренной пролиферацией изменения метаболизма, снижение окислительно-восстановительного потенциала, усиленное перекисное окисление липидов влияют и на состояние ядерной мембраны клеток. Вязкость ядерной мембраны падает, а неспецифическая проницаемость ее повышается. Физиологический барьер между содержимым цитоплазмы и ядра ослабевает. В ядро из цитоплазмы начинают проникать чуждые для него вещества. Среди них могут быть и вещества, способные активно вмешиваться в работу генетического аппарата. Известно, например, что нарушение проницаемости ядерной мембраны может приводить к проникновению в ядро из цитоплазмы некоторых эндогенных мутагенов фенольной природы. Такие вещества могут взаимодействовать с комплексом ДНК—белок и затруднять нормальные процессы репрессии — дерепрессии генетической информации. Длительно поддерживаемый внешней для клетки стимуляцией ускоренный ритм клеточного деления одновременно приводит также к накоплению повреждений, ошибок обмена и синтеза. Могут, по-видимому, накапливаться и спонтанные и индуцированные повреждения комплекса ДНК—белок, в первую очередь той его части, которая связана с функциями регуляторных генов, зависимых от целого ряда других процессов в клетке, в частности от состояния ближайшего окружения ДНК-белкового комплекса. Часть повреждений и изменений фиксируется в трудно репарируемые, и некоторые из них могут трансформироваться в изменения суперструктуры ДНК или в изменения структуры хроматина. Это уменьшает репарируемость возникших изменений в ДНК и хроматине. Недорепарированные по тем или иным причинам изменения и повреждения ДНК могут накапливаться.
 Рис. 17. Четвертый круг
Ведущие процессы: нарушение проницаемости ядерной мембраны и фиксирование на уровне генетической регуляции изменений метаболизма и степени дифференцировки
Рис. 17. Четвертый круг
Ведущие процессы: нарушение проницаемости ядерной мембраны и фиксирование на уровне генетической регуляции изменений метаболизма и степени дифференцировки
Все это может вызывать затруднения в процессах репрессии — дерепрессии нормальной генетической информации и в процессах транскрипции ДНК. В случае многократного повторения указанных изменений в регуляторных функциях ДНК во многих поколениях клеток при нарастании затруднений в процессах выбора и транскрипции локусов генома, по-видимому, может происходить фиксирование на уровне генетической регуляции происшедших изменений метаболизма и дедифференцированного состояния, хорошо соответствующего ускоренному ритму деления клеток. Поэтому снятие внешнего стимула к усилению пролиферации в этих условиях не снимает ускоренный ритм деления. Начал функционировать четвертый внутриклеточный порочный круг (рис. 17), также препятствующий возвращению клеток к нормальному уровню дифференцировки и к спокойной пролиферации. Рассмотренные порочные круги не исчерпывают всех возможных взаимосвязей, возникающих при длительной стимуляции пролиферативной активности, которые затрудняют возвращение клеток в исходное дифференцированное состояние. Можно найти и другие взаимосвязанные процессы в разных внутриклеточных структурах, создающие аналогичные порочные круги. Задача заключалась не в этом, а в поиске более общих закономерностей. Стало ясно, что длительная по времени стимуляция клеточного деления смещает пролиферативно-дифференцировочное отношение в сторону недостаточной дифференцировки. Повышается активность эволюционно старых метаболических процессов, направленных на обеспечение подготовки и деления клеток. В конечном итоге это приводит к уменьшению компенсаторного резерва внутриклеточных систем, к гипотрофии и гипоплазии структур, ответственных за дифференцировку. Это вновь открывает пути к ускоренной пролиферации. Ускоренный ритм деления хорошо соответствует недостаточной дифференцировке клеток, и потому он сохраняется и в случае снятия внешнего для клетки сигнала к ускоренной пролиферации. В последние несколько лет появились принципиально новые данные по взаимосвязанным внутриклеточным процессам, в которых участвуют циклические нуклеотиды, протеинкиназы, фосфоинозитолы, диацилглицерины и фосфолипазы и которые в порядке положительной обратной связи могут создавать эффект длительного присутствия внешнего стимулирующего пролиферацию фактора. Главной особенностью рассмотренных порочных кругов во внутриклеточных системах является их тесная, неразрывная связь друг с другом. Одни и те же процессы могут присутствовать в различных кругах. Разделение кругов весьма условно. Вторая их особенность заключается в том, что все они функционируют одновременно. Здесь нет той последовательности вовлечения одного порочного круга за другим по мере развития событий в первом. Некоторая последовательность событий в каждом порочном круге имеется, и в этом их специфика. Однако начало функционирования кругов происходит одновременно и вызывается одной и той же причиной. События одного порочного круга усугубляют события других порочных кругов. Каждый из порочных кругов можно рассматривать как отражение разных углов зрения для принципиально одних и тех же процессов. Третья особенность внутриклеточных порочных кругов связана с тем, что основная причина их возникновения общая: длительное время сохраняющееся ускоренное деление клеток в связи с непрерывной, хронической внешней для клеток стимуляцией пролиферативной активности. Это причина всех возникающих последующих событий. Следующая особенность внутриклеточных порочных кругов заключается в общем возможном следствии их функционирования: создание в клетке внутренних условий, поддерживающих ускоренный ритм деления клеток и в случае снятия сигнала к стимуляции пролиферативной активности. Тогда уже ускоренный ритм клеточного деления начинает сам поддерживать все изложенные биохимические, биофизические и цитологические изменения, а последние, в свою очередь,— поддерживать ускоренный ритм деления клеток. Поэтому появляется определенная автономность самоуправления скоростью пролиферации, которая в силу рассмотренных причин может закрепиться в генетической регуляции, т. е. вызвать стойкие изменения в регуляторных генах, а это, по-видимому, и есть первый шаг в сторону малигнизации клеток. Подчеркиваем, что для малигнизации клеток с этой точки зрения чрезвычайно важно, чтобы указанные изменения продолжались в течение очень длительного времени. Здесь фактор времени, вероятно, является определяющим. Необходимо постепенное накопление проходящих через митоз многих поколений клеток неспецифических изменений, закрепляемых в силу этого в регуляторных генах. Известно, что для абсолютного большинства видов лейкоза и других злокачественных заболеваний необходим очень длительный период накопления изменений, многократно превышающий продолжительность жизни клеток пролиферирующих тканей. С такими же, но сравнительно кратковременными изменениями в условиях гомойтермного организма млекопитающих клетки, как правило, успешно справляются, и малигнизации не происходит. Однако в случае малигнизации вследствие большой нестабильности генома появляются злокачественные клетки с широким набором важнейших цитологических характеристик. Часть из них находится длительное время в периоде G0 и редко вступает в деление.
Последние комментарии
4 часов 42 минут назад
4 часов 50 минут назад
4 часов 59 минут назад
5 часов 5 минут назад
6 часов 34 минут назад
6 часов 37 минут назад